JOuRNALOF VIRoLOGY, Aug. 1979, p. 325-333 0022-538X/79/08-0325/09$02.00/0
Vol. 31, No. 2
Temperature-Sensitive
Mutants
of
Complementation
Group
E
of
Vesicular Stomatitis Virus New Jersey
Serotype
Possess
Altered
NS
Polypeptides
D.EVANS, C. R. PRINGLE,ANDJ. F. SZILAGYI*
Medical Research CouncilVirologyUnit,Instituteof Virology, University of Glasgow, GlasgowGll5JR,
Scotland
Received for publication 29 March1979
Invesicular stomatitis virus NewJerseyserotypepolyacrylamide gel
electro-phoresiswasunabletodistinguishthepolypeptidesof thetemperature-sensitive
(ts) mutants ofcomplementationgroupsA,B,C, and F from those of the wild-type virus. However, the NSpolypeptide of therepresentativemutantof group E, tsEl, had asignificantlygreaterelectrophoretic mobilitythan that of the wild-typevirus NSpolypeptide.Theelectrophoretic mobilitiesoftheNSpolypeptides
ofthe three mutants of complementation group Evaried, being greatest in the caseof tsEl,slightlyless for tsE2,andonlyalittle greater than that ofwild-type
virus NS polypeptide in the case of ts E3. Since the NS polypeptides ofthe revertantclones ts El/Rl and tsE3/Rl have mobilities identicaltothatof
wild-typeNSpolypeptide,theobservedaltered mobilities of the group Emutants are
almost certainly the direct result of the ts mutations in the E locus. The
electrophoretic mobilities ofthe intracellular NS polypeptides of the group E
mutantswereindistinguishable from those of their virion NSpolypeptides. The
electrophoreticmobilities of theNS polypeptidesof the group E mutants
synthe-sized in vitro using mRNA synthesynthe-sized in vitro by TNP were identical to those of theNSpolypeptides of their purified virions. The NSpolypeptidesofall three mutants werelabeled with
'Pi
toapproximately thesame extent as wild-typevirus NSpolypeptide, indicatingthatgrossdifferences inphosphorylationofthispolypeptide areunlikelytoaccountfor thealtered mobilities. We proposea
model in whichthe NS polypeptide consists ofatleast three loopsheld in this configuration by hydrophobic orionicforces or both andstabilized by phospho-diester bridges. If a mutation affects one of the amino acids to which the
phosphateiscovalently linked,thephosphodiester bridgecannotbeformed, and,
as aresult,in the presence of sodiumdodecylsulfate theaffectedloopopensand
thus the NSpolypeptidemigrates further into thegel.Suchaconfigurationmay
alsoexplainthemultifunctionalnatureof the NSpolypeptide.
Vesicularstomatitis virus (VSV)NewJersey
serotype consistsofasingle-strandedRNA mol-ecule and five polypeptides, L, glycosylated G,
N,phosphorylated NS,and M(12, 13,
15,22,28-30). PolypeptidesN,NS, andM,aswellas the
unglycosylated precursor to G, have been
syn-thesized in the rabbitreticulocyte lysate system
by usingRNAsynthesizedin vitroby the
tran-scribing nucleoprotein (TNP)complex (15).
The temperature-sensitive (ts) mutants of VSV New Jersey fall into six nonoverlapping complementation groups; the majorityisolated
belong to groups A and B, whereas group C
contains only four mutants, group E contains three, groupFcontains two,and groupD con-tainsonlyone(16, 17).
AttherestrictivetemperatureviralRNA
syn-thesis was defective in BHK-21 cells infected with mutants ofgroups A, B, and F, butnot in thoseinfectedwith mutants of groups C and D (17). Mutants of group E were heterogeneous; ts El and ts E3 did not synthesize viralRNA at
390C,
whereastsE2did (17). Viral protein syn-thesisat390C
ininfected BHK-21cellsexhibited a similar pattern (30). Subsequent analysisshowed thatprimary transcriptionwasinhibited at39.5°C in cells infected with ts Bl and ts Fl but not incells infectedwith tsAl,tsCl,tsDl, andtsEl (9).
Recently, Szilagyi and Pringle (26) showed
that in vitro RNA synthesis at
390C
by the RNA-negative ts Al and the RNA-positivets Cl and tsDlwas similartoRNA synthesis in wild-type virus, whereas the RNA-negative ts Bl325
on November 10, 2019 by guest
http://jvi.asm.org/
326 EVANS, PRINGLE, AND SZILAGYI
synthesizedonlysmallamounts of RNA in vitro
at 39°C. Mutant ts El synthesized very little RNA,tsE2 synthesizedmoderate amounts, and ts E3 was not inhibited. The group F mutants werealso dissimilar;tsFlsynthesized only very little RNAin vitro at39°C,whereas ts F2 syn-thesized as much RNA as did wild-type virus. We haveanalyzedthepolypeptidesofsomeof these ts mutants of VSV NewJersey to
deter-mine whether differences in theelectrophoretic mobilities of any of the structuralpolypeptides areassociated withanyof thecomplementation groups. Initially, we examined one representa-tive mutant from each complementation group and found that the mobility of the NS poly-peptideof ts El differedfrom that ofwild-type NS polypeptide. The present paper describes ourconsequent studyof the NSpolypeptide of all threecomplementation group Emutants.
MATERIALS AND METHODS Mutants. The tsmutantsof VSV NewJersey de-scribed previously by Pringle etal. (17) were used. The revertant clone ts El/Ri has been described previously bySzilagyiandPringle (26),andtsE3/R1
was isolated from BS-C-1 monolayers incubated at
390C.
Growth and purification of virus. Virus was grown inBHK-21 clone13monolayersandpurified by
the meothod of Sziligyi and Pringle (24). Purified virus was suspended in 20 mM Tris-hydrochloride
buffer, pH 8.0, and theproteinwasdeterminedbythe method ofLowryetal.(10).
Preparationofradioactivelylabeled virus. Vi-ruswasgrown in 80-ounce (2,400-nil) Burrler bottles
asdescribed above except that (i) the medium con-tainedonly20%oof itsnormal amino acidcomposition
when theviruswas tobelabeled with[3S]methionine,
or(ii)the mediumwasphosphate-freewhen the virus wastobe labeled with32P.[35S]methionineor32Pi(200
ytCi in35 ml of medium perbottle) wasadded 6h
postinfection. Afterharvesting, the medium contain-ing the virus wasclarifiedby centrifugation at 2,500
rpmfor 30 minto removecellulardebris,andthen the virus waspelletedbycentrifugationat21,000 rpm for
90minthrougha30%glycerolcushioncontaining20
mMTris-hydrochloride,pH8.0,1mMEDTA,and0.1
MNaCl.Thepelletswerethenresuspendedinabuffer consistingof 20 mMTris-hydrochloride buffer, pH8.0,
1mMEDTA, and 100mMNaCl.
Preparation ofintracellular
[35Slmethionine-labeled polypeptides. Throughout theexperimentthe medium contained only20% of its normal amino acid composition. Each BHK-21 cellmonolayer ina Burrlerbottlewasinfectedwithalowmultiplicityof purified virus (24) andincubated at310Cfor22h in 35 mlofmedium. After this incubationthemedium was removed, and the surface of thecell monolayer
waswashed twicewithfresh medium. Then 20ml of medium containing 300 ,uCi of [3S]methionine was added, and incubationwascontinued at310C. After
90minofincubation, themediumwasremoved, and thecell surfacewaswashed twice with fresh medium.
Finally, 15 ml of 20 mM Tris-hydrochloride buffer,pH 8.0, wasadded, the cell monolayer was removed by sterileglass beads, and a homogeneous suspension was obtainedby extensive sonication.
Amock-infected cell suspension was obtained in the sameway except that the virus was omitted.
Preparation of TNP. TNP complexes were pre-pared by the method ofSzilagyi and Uryvayev (27), incorporating the modifications of Szilagyi and Pringle
(25).
In vitro RNA and protein synthesis. Comple-mentary RNA wassynthesized in vitro at310Cfor 90 min in a 0.2-ml incubation mixture containing 0.02 ml ofTNP,0.05 ml ofreagent mixture (containing 400 mMTris-hydrochloride buffer, pH 8.0,400 mM NaCl, 3.2Mig of actinomycin D per ml, 2.56 mM ATP, 2.56 mMCTP, 2.56 mM GTP, 0.256 mM UTP, and 12,tCi
of[5,6-3H]UTP), 3.5 mMdithiothreitol, 2 mM addi-tionalATP, 25,iM S-adenosylmethionine, and 1 U of ratliver RNase inhibitor (Amersham/Searle) per ml. An unfractionated rabbit reticulocyte lysate was prepared as described by Preston and Szilagyi (15). Theendogenous RNA from the lysate was removed by treating it with micrococcal nuclease as described by Pelham and Jackson (14).
The cell-free translation system (25 ,u) contained 100 mMHEPES (N-2-hydroxyethyl piperazine-N'-2-ethanesulfonic acid) buffer (pH 7.7), 40 mMKCl,0.8 mMspermidinetrihydrochloride, 1 mM ATP, 0.1 mM GTP, 4mg of creatinephosphate per ml, 200 Mg of creatinephosphokinaseperml, each of the 19 common amino acids except methionine at a concentration of 50
AM,
500ACi
ofL-[35S]methionine, 12.5% lysate,and200MuMhemin.Finally,2.5,l of the incubation mixture wasadded, and it was incubated at310Cfor 60min. Protein synthesis was terminated by the addition of 25 Il of a solution containing 100 mM EDTA, 2% methionine and 300 ,ug of RNase A per ml, followed
byafurtherincubation at310Cfor 15 min.
Polyacrylamidegel electrophoresis. Samples of
purifiedvirus or cell-free translation system were an-alyzed by discontinuous sodium dodecyl sulfate
(SDS)-polyacrylamidegradient gel electrophoresis as described by Marsden et al. (11). To obtain greater separationof the N and NS polypeptides, the electro-phoresis was continued for an additional 2 h after the bromophenol blue dye front had reached the bottom of thegel. Kodirexfilm was used for the autoradiog-raphy.
RESULTS
Polypeptide composition of the virions of thets mutantsof VSV NewJersey.We chose arepresentative mutantfrom eachofthe com-plementation groups A, B, C, E, and F and analyzedtheirpolypeptide compositions by dis-continuouspolyacrylamide gradient gel electro-phoresis. Group D was excluded for reasons outlinedbelow.
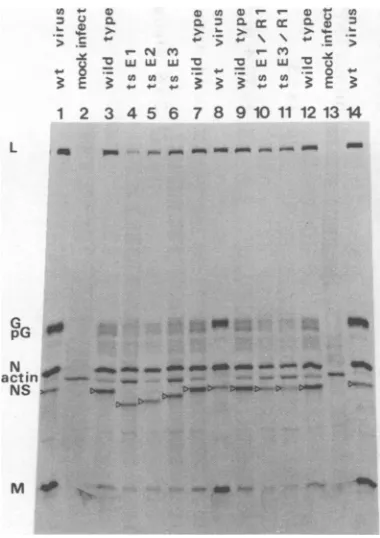
Figure 1shows thatwild-typeVSV New Jer-seycontains fivepolypeptides, L, G, N, NS,and M, although at least two minor bands (MB1, MB2) of unknownoriginwerealso detected. The
J. VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
NS POLYPEPTIDES OF VSV ts MUTANTS 327
polypeptides of themutantstsAl,tsBl,tsCi, andtsFl had electrophoretic mobilities identi-caltothose of the wild-type virus polypeptides. However, the NS polypeptide oftsEl hasa
significantly greater electrophoretic mobility than that of the wild-type virus NS polypeptide, whereas the mobilities of the other polypeptides
werenotdifferent (Fig. 1). This indicated that
the ts El mutation might have affected the electrophoretic mobility of the NS polypeptide. Polypeptide composition of thevirionsof
the group E mutants. Since the NS poly-peptideof tsEl hadanalteredelectrophoretic
mobility inapolyacrylamide gel,wedecidedto
examine allthreemutantsof complementation
group E,as wellastherevertantclones tsEl/
Rl andtsE3/Rl.
01)
-CL
3 <2
n 4U:
0) Q)
X -_ i. LL
U) 0 0
4- 4-1 4- 4t
.. :. :: .... :. :}:.:..: .:
^.=
Ls_w_-
. rg*.:...::...
.... .w *.:hi,s. s i . ''... ... ... .. ..
:. .: ::
.D ..
..:...:::::::::
:: :: .:: ::. :: .. : ...
.. .. :.
:. ... .. ..
.:
...:...:.
....,S,.--;-'
.*...:
.. . . ..
.: .: .5 .... ...
.,
.. .: :: .. ....
G ^6i__wlw _ _- t_ __
N NS
MB2 >4; *4
MB1
M e4 f
FIG. 1. Virionpolypeptides of the ts mutants of VSV New Jersey. [ Sjmethionine-labeled purified
virionsofwild-typevirus andts mutantswere
ana-lyzedon a discontinuousSDS-polyacrylamide
gra-dientgelasdescribed inthe text. Each track
con-tainedapproximately60),000cpmof[3Slmethionine.
ThepolpeptidesL,G, N, NS,andM,as well astwo
minor bandsofunknown origins (MB1, MB2), are
indicated. Toemphasizetheposition ofthe NS poly-peptides, theyaremarkedbyopentriangles.
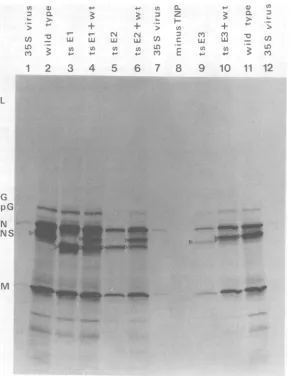
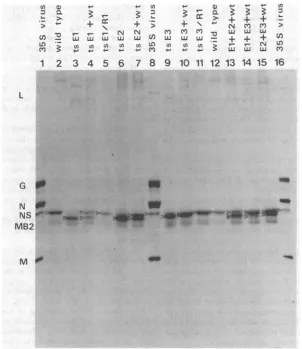
Figure 2 shows thatthe mobilities of
polypep-tides L, G, N, and M of each mutant and the
two revertants wereindistinguishable from those of wild-type virus polypeptides although the NS polypeptides of the threetsmutantshad differ-entmobilities.
Because theelectrophoretic mobility of N was
identical in eachcase, it was convenient to
ex-presstherelative mobilities of the NS polypep-tides of the mutants by comparing theextentof theseparation of their N and NSpolypeptides
with theseparationobserved inwild-typevirus.
Theseparation ofpolypeptides Nand NSwas
approximately 1.6timesgreaterfor ts El than
for wild-type virus (Fig.2, tracks 1 through3),
it wasslightly less in the case of ts E2
(approxi-mately 1.4 times greater, tracks 5 through 7),
and in tsE3 the separation was only 1.3 times greaterthan inwild-type virus (tracks 8 through
10).
Thedifferencesintheelectrophoretic
mobili-ties of theNSpolypeptides ofthe three ts mu-tants wereclearly resolvedin thosetracks where
mutantsandwild-typevirusweremixed in
var-ious combinations (Fig. 2, tracks 13 through 15).
Thus,the order of the mutants, according to
the extentofmigrationof their NSpolypeptide,
is tsEl, ts E2, ts E3.
Theelectrophoretic mobilitiesof theNS
poly-peptides of the revertant clones ts
El/Ri
and tsE3/Rlappear tobe identicaltothat of the NS
polypeptide ofthewild-typevirus(Fig. 2, tracks
4, 5, 11and 12). The simultaneousreversion of the tsphenotypeand theelectrophoretic mobil-ity ofthe NSpolypeptide stronglysuggeststhat the ts mutation is responsible for the altered
mobilityof the NSpolypeptidesof themutants
ts El and tsE3,andprobablyof ts E2 aswell.
Polypeptide synthesis in
cefls
infectedwith the group E mutants. BHK-21
celis
were infected with the three group E mutants and the two revertants, and the intracellularpolypep-tideslabeled duringa 90-minpulse of
[3S]me-thionine were analyzed by polyacrylamide gel
electrophoresis (Fig. 3).
Actin was the predominant labeled
poly-peptide in mock-infected cells (Fig. 3, tracks 2
and 13). In cells infected with the wild-type
(tracks3,7, 8, and11),mutant(tracks4
through
6), andrevertant(tracks10and11)virusesthere were,besides theactin, sixmajorlabeled
poly-peptides. These were the viral polypeptides L, G, N, NS, and M, as well as a further
poly-peptide,pG,which ispresumablyaprecursorof G.Ineach casepolypeptides L,G, pG, N,and M were
indistinguishable
from thepolypeptides of the wild-type virus(tracks1 and 14). However,theelectrophoreticmobilitiesofpolypeptideNS
in cellsinfectedwith thethree mutants(tracks
-VOL. 31,1979
on November 10, 2019 by guest
http://jvi.asm.org/
[image:3.504.82.226.263.573.2]328 EVANS, PRINGLE, AND SZILAGYI
a 4r,4)
e- r- ._ N
LU IL L - wU
U,0 U- 4t
4-1 2 3 4 5
* ::
L i r¢z.
+ 4- + \
X - L UJ L
u) : z u: u
6 7 8 9 10 11 12 13 14 1516
*:.
0% 44^_N
S
NS t
Cr~*"
~~
D [image:4.504.114.404.72.414.2]M -~ up_ .qoe- e-4I~
FIG. 2. Virionpolypeptides ofthetsmutantsofcomplementationgroupE.[35SJmethionine-labeledpurified
virions ofwild-type VSV New Jersey (wt), the threets mutants(tsEl,tsE2,tsE3) ofcomplementationgroup
E, and therevertant clones (ts El/RI, tsE3/RI) wereanalyzedon adiscontinuous SDS-polyacrylamide
gradient gelindividuallyorin mixedtracks,asindicated.Each track contained approximately60,000cpmof
[35S]methionine. Therefore, 60,000,30,000,or20,000cpmof[S]methionine for each viruswasplacedonthe
tracks,dependingonwhether the viruseswerealoneorincombination.Thepositionsof the NS polypeptides
areindicatedbytheopentriangles.
4 through 6) were different from that of NS
polypeptide synthesized in cells infected with
wild-type virus(tracks 3,7, 9,and 12) andwere
alsodifferent fromeach other. Theseparations ofthe Nand NSpolypeptides forts El,ts E2, and ts E3 were 1.6, 1.4, and 1.3 times greater, respectively, than the separation for wild-type virus. Thus, the mobilities ofthe intracellular NS polypeptides ofthe group E mutants were
indistinguishablefrom thoseof thepurified
vir-ionNSpolypeptides.
The intracellular NSpolypeptidessynthesized in cellsinfected with the revertant clones(Fig. 3,tracks 10 and11)wereindistinguishablefrom
the NSpolypeptideofthewild-typevirus(tracks
9and12), which strengthensthesuggestionthat thealteredelectrophoreticmobilitiesofthe NS
polypeptidesofthemutantsarethe result ofthe
tsmutations.
Invitro-synthesizedpolypeptides. Asthe aberrant electrophoretic mobilities of the NS polypeptidesingroupEappeartobe the result of mutation, these differences should also be observed in polypeptides synthesized in vitro. We tested this bysynthesizing mRNA'sinvitro, using TNP complexesandtranslatingthem ina
rabbitreticulocytelysate cell-freesystem.
Figure 4shows that four major polypeptides
were translatedwhen the RNA synthesized by
wild-typeTNPwasused. Three of these
corre-spondtothe N, NS, and M polypeptides of the virion, whereas the fourth polypeptide (pG) is presumably the unglycosylated G polypeptide (Fig. 4, tracks 1, 2, 11, and 12). In
vitro-synthe-N
-a
13-
LU+
r-LUJ
LU LLU
+ + 7
LU
NU
J. VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
NS POLYPEPTIDES OF VSV ts MUTANTS 329
Q .
I._
-
=-r.L CL cc ClC rn CL)m
>- t. >. w + : *,\ \ - .C -T- N M r-M X Y
0 ._ E 3
1 2 3 4 5 6 7 8 9 10 11 12 13 14
m PM _ _ _ _ _ _ _
pG
X oLe-
" ="M aN do_^, am___ Om am _
actin __ ,__
NS
FIG. 3. Intracellular polypeptides ofcelsinfected
with thegroup E mutants. [55],methionine-labeled infected cellextractswereobtainedandanalyzed by
SDS-polyacrylamidegradient gel electrophoresisas
described in thetext. Eachtractcontained
approxi-mately5,000cpmof labeledcellextract.For
compar-ison, [35S]methionine-labeledpurified wild-typeVSV
NewJersey virions (wt virus)or amock-infected ceU
preparation wasused. The NSpolypeptidesare
in-dicatedby theopentriangles.
sized polypeptides pG, N, and M of all three
mutantswereidenticaltothose of the wild-type virus (tracks3, 5, and 9).
Theelectrophoretic mobilities of the in vitro-synthesized NS polypeptides oftsEl andtsE2
were significantly greaterthan the mobility of
wild-type virus NS polypeptide (tracks 3 through 6); the mobility of the NS polypeptide oftsE3, although lower, wasstill greaterthan the mobility of the wild-type NS polypeptide (tracks9and 10).
The differences in theelectrophoretic mobili-ties ofthe NS polypeptides of themutantsare
easierseenherethan inthevirionanalysis,since
the NSpolypeptide isamajor product of the in
vitrotranslation. The in vitro results fully
con-firm the differences in the electrophoretic
mo-bilities ofthe NS polypeptides of the group E
mutants.
Since themobilitiesofthe NSpolypeptides of thevirions andinfectedcells,aswellasofthein
vitro-synthesized NS polypeptides of all of the mutants, were identical, it seems unlikely that the altered mobilitiesof the NS polypeptides are due to aposttranslational cleavage. The lack of low-molecular-weight cleavage products ob-served ingels where electrophoresis was stopped before the dye front ran off the gel is further evidence that cleavage does not take place.
Phosphoprotein composition ofthe group E mutants. The mutants were grown in the presenceof32Pito determine whether the alter-ations in the electrophoretic mobilities of the
NS polypeptides of the group E mutants were
due todifferencesinthe degree of phosphoryl-ation. Equal amounts of protein were placed on each track todetermine whether there were any differences in the degree of phosphorylation of the mutants.
Figure5shows thatonly NS was
phosphoryl-ated(tracks1, 2, 8, 12, and 16).One of the minor
polypeptidesofunknown origin (MB2) was also
phosphorylated. The 3P-labeled NS
polypep-tidesof the three mutants showed differences in
mobilities similar to those obtained previously
with[3S]methionine-labeled virus.
The chief conclusion from Fig. 5 is that the NS polypeptides of all three mutants and the two revertants arephosphorylated.The degree
ofphosphorylationof the NS polypeptide of the
individual mutants is approximately of the same order ofmagnitudeasthatof thewild-typevirus
NS polypeptide. This is clearly illustrated in
mixed tracks whereequalamountsof themutant
and wild-type virus were present (tracks 4, 7,
and10).
DISCUSSION
Polyacrylamide gel electrophoresis was
un-abletodistinguishdifferences between the
poly-peptidesof thets mutantsofcomplementation
groups A,B, C, and F andthe polypeptides of
the wild-type virus. Previously Wunner and
Pringle(31)reportedthat the G and N
polypep-tides ofts Dl, the onlymutantof
complemen-tation group D, exhibited aberrant
electropho-reticmobilities and that theNpolypeptide de-fect was associated with the ts lesion. Subse-quent work suggests that neither polypeptide
defect may beresponsiblefortemperature sen-sitivityoftsDl.This workwillbe thesubjectof aseparate report andistherefore excluded from this discussion.
Theexperiments describedinthispaper show that the NS polypeptides ofall threemutants
have altered electrophoretic mobilities in rela-tionto oneanother andtowild-typevirus.Since the electrophoretic mobilities ofthe NS
poly-peptides ofthe revertantclonestsEl/Rlandts
L
VOL. 31,1979
on November 10, 2019 by guest
http://jvi.asm.org/
[image:5.504.55.245.71.340.2]330 EVANS, PRINGLE, AND SZILAGYI
a a 2
.-4 N N
> + + > VI
C) - Li LU LU LU
LO g TO !In 0 S _
1 2 3 4 5 6 7 8
LU Ln- '7
9 10 11 12
L
G pG
N '-NS cI'
-.4S.
_;uif
[image:6.504.109.402.71.447.2]m om o
FIG. 4. Polypeptidessynthesized in vitrousingRNAproduced in vitroby TNP complexes of the group E mutants.The preparationofthe TNPcomplexes and the in vitro RNA and protein syntheses aredescribedin the text. Tracks 1, 7, and12contained[35S]methionine-labeledpurified virions ofwild-typeVSV New Jersey
(35Svirus). Track8containedcell-freetranslation systemfromwhich the TNPwasomitted. The other tracks containedcell-freetranslation systemsofthevarious mutants andwild-type virus,asindicated.Singletracks contained 20 ttl. Mixed tracks contained 10 ul from each of the cell-free translation systems. The NS
polypeptidesareindicatedby the opentriangles.
E3/Rl wereidenticaltothat of
wild-type
virusNSpolypeptide,the alteredmobilitiesof the NS
polypeptidesofthemutantstsEl andtsE3are
almost certainly the result of the ts mutation.
Presumablythis isalsotrueforts
E2, although
as yet we have been unable to obtain a stable spontaneous revertantfromthismutant. Thus,
theinhibitionof virusdevelopmentof thegroup E mutants at the restrictive temperature is
al-mostcertainlyduetoinabilityoftheirmutated
NS polypeptides to function normally at this temperature.
Recently,SzilagyiandPringle(26)foundthat thevirion-associatedtranscriptase
activity
oftsE1 is temperature sensitive, although in vitro RNA synthesisis not inhibitedat 390C in the
caseoftsE2 andtsE3. In vivo studies showed thatts E3 isdefective in thereplicationofthe virion RNA (17), whereas in the case of ts E2 replication did notappear to beinhibited (17).
Therefore, Szilagyiand Pringle suggested that
the polypeptide affected by the mutation in group E is a multifunctional polypeptide in-volved intranscription, replication,and possibly
somelate stage of virusdevelopment (26).Since the results presented in this paper showthat the mutatedpolypeptideisalmostcertainlyNS, we conclude that NS is the multifunctional
poly-J. VIROL.
NOW
on November 10, 2019 by guest
http://jvi.asm.org/
NS POLYPEPTIDES OF VSV ts MUTANTS 331
4)(1-' -4-'cn ) tt Ut
L- a>
t-L-> I= ,- L > + + +
> + 3 ~~~~~~N~ >~
-~~~~~- ~~~ N N CL WL Uii
Cl) LWLLJ LX LAU)LL X U- + + + l LO >LOU) XJa U) V1 U:*-, N LO
5 6 7 8 9 10 11 12 13 14 15 16
L
NWS
[image:7.504.97.399.72.421.2]MB2 >HJ
FIG. 5. Phosphoproteincomposition of complementationgroupEmutants. 32P-labeled purifiedvirionsof
thegroupEmutantsandtheirrevertantswereanalyzed bydiscontinuousSDS-polyacrylamide gradient gel
electrophoresisasdescribed in thetext.Each trackcontained 9 ,ugofvirusprotein.Intracks whichcontained
morethanonevirus,theamountsofeach viruswereproportionately reduced in ordertomaintain thetotal
protein concentrationataconstantlevel. The NSpolypeptidesareindicated by theopentriangles.wt, wild-typevirus.
peptide. That the NS polypeptide of VSV
Indi-anaisinvolved intranscription has already been
shownby dissociationandreconstitution
exper-iments (4, 7). Our evidencesuggeststhat the NS polypeptide is also involved in thereplicationof virionRNA, althoughwecannotsaywhether it
isthereplicaseitselforpartofa replicase
com-plex. Since viral RNA synthesis takes place ints
E2-infectedcellsattherestrictivetemperature,
the involvement of NS polypeptide in a late
stage ofvirusdevelopment is suggested. Previ-ously, Szilfigyi and Pringle (26) showed that the heat stability of the transcriptase ofts El was
dependent on an interaction between the viral
coatandthecore.
At present it is not certain whatcauses the
marked alterationintheelectrophoretic mobil-ities of theNS polypeptides of the threegroup
Emutants. Twopossible explanations arise: (i) either the NSpolypeptides of thegroup E
mu-tantsareshorterthan theNSpolypeptide of the
wild-type virus, or (ii) the phosphorylation of
mutant NS differs from that of the wild-type NS.
The occurrence of a shortened polypeptide canbeexplained in severalways,although there
areobjectionstoallof them.(i) A section of the
NS cistronmayhave been deleted by the
mu-tation. However,weconsiderthisunlikely since
the base analog 5-fluorouracil, which favors point mutation, was used as the mutagen and
sincerevertantswereobtained fromtsElandts
E3athigh frequency.
(ii) The mRNA for NS may have been
pre-maturely terminated. However, all of the evi-dence available suggests that transcription is
1 2 3 4
VOL. 31,1979
on November 10, 2019 by guest
http://jvi.asm.org/
332 EVANS, PRINGLE, AND SZILAGYI
initiated only at the 3' end ofthe genome (1, 2). Thus,premature termination of the mRNA syn-thesiswould producepolareffects whereby the subsequent messengers for the M, G, and L polypeptides would notbe transcribed. To ex-clude this possibility, the mRNA's of the mu-tantswill have tobecomparedwith those ofthe wild-type virus by gel electrophoresis and
oli-gonucleotidefingerprinting.
(iii) The mutant mRNA may contain an early termination or late initiation codon resulting in a shorterNSpolypeptide.To determinewhether this isthe case, thesequence of themRNA's of
the various group E mutants will have to be
determined.
However, itisdifficulttoreconcile early ter-mination or late initiation of transcription or
translation with the loss of functions of the
various group E mutants.
(iv) Another possibility is that mutation al-tered the NS polypeptide insuch a way as to render it moresusceptibletocleavage by
prote-olyticenzymes.This is alsounlikely since the in
vitro-synthesized NSpolypeptideshadsimilarly alteredmobilities. Also, it isgenerallyheld that in the rabbit reticulocyte lysate system the in
vitro-synthesized polypeptides are not cleaved
(3, 18, 21), and there was no evidence for the presence of a low-molecular-weight cleavage
product. Furthermore, the mobilitiesof the
in-tracellularNSpolypeptides of the group E
mu-tants were indistinguishable from those of the purified virion NS polypeptides,making prote-olytic cleavage even more unlikely.
SinceNS is aphosphorylatedpolypeptide, the secondpossibility is that the aberrant mobilities of this polypeptide in thevarious group E mu-tants are aconsequence of the differences in the extentofphosphorylation. However, the exper-iment with32P-labeled virions indicated that, in the case ofall three mutants,polypeptide NS is
phosphorylated toapproximately the same
ex-tent asit is inwild-type virus. Therefore, gross
differencesin the level ofphosphorylationinthe
variousmutants areunlikelyto bethe cause of thedifferencesinthemobilitiesofthe NS poly-peptide.
Wepropose a model which couldexplain both thealteredmobility of the NSpolypeptide and the nature ofthegroup Emutation.According tothismodelthe NSpolypeptide chain is folded into atleast three loops. Each loop is held to-gether by ionic orhydrophobic bonds or both
and stabilized in this configuration by a
phos-phodiester bridge. Phosphate diesters cannot be hydrolyzed by alkaline phosphatase (6, 19), which explains why polypeptide NS was not dephosphorylated by this enzyme (12, 22; un-published observations on VSV New Jersey).
J. VIROL.
Therefore, the structure ofpolypeptide NS can-not bedenatured by boiling in SDS and 2-mer-captoethanol during preparation for polyacryl-amide gel electrophoresis, and hence it would notmigrate asfar into the gel as afully dena-turedpolypeptide. This would also explain why the NS appears as ahigh-molecular-weight poly-peptide inpolyacrylamidegels despite the rela-tivelysmall coding capacity of its mRNA and
also would account for the differences in the
mobilitiesof the NSpolypeptideindifferentgel
systems (5,8, 20,23).
Ifmutation results in the substitution ofone of the amino acids to which the phosphate is
covalently bonded, the phosphodiester bridge
cannot be formed and the stabilization of the
loop doesnottakeplace. Therefore, under
de-naturing conditions this loop opens, and the
mobility of the NS
polypeptide
in the gelin-creases.The extentofthis increasepresumably
dependsonthe size of theloopaffected bythe
mutation,whichwouldexplain the differing
mo-bilities of the NS polypeptides of the group E
mutants.
Our model can alsoexplain the ts nature of
themutation.Atthepermissivetemperature the
ionic and hydrophobic bonds alone are
suffi-cient, but at the restrictive temperature the
phosphodiester bond is required to hold the
configuration of the loop. Thus, a mutation
which results in the breaking of the
phospho-diester bridgewill be temperature sensitive for the function of that loop. Mutations affecting
any other amino acid notlinked by the
phos-phodiester bond will presumably be either a
silentor alethalmutation.
Accordingtothismodeleachloop corresponds
to a different function of the NS polypeptide.
One of theloops (loop ts
El)
isinvolved in thetranscription of mRNA's, the second (loop ts
E3) isinvolved in thereplicationof virusRNA,
and the third (looptsE2)isinvolved in the late
functioninwhich themutant tsE2is defective.
The model would explain why there is no
correspondencebetween the loss of functionsby
the mutants andthe differencesin the electro-phoreticmobiitiesof theNSpolypeptide.
Althoughatpresentwehavenoexperimental
evidence tosubstantiate our model,weare ac-tivelyengagedinthedetermination of the struc-ture ofthe NS polypeptide.
Also,
we willhave to isolate morerevertantclones,especiallyfrom tsE2,sinceattemptstoisolate stablerevertants from thismutanthave sofar beenunsuccessful.ACKNOWLLEDGMENTS
We thank J. H.Subak-Sharpeforacriticalreadingof the manuscript. We also thank C.Cunninghamfor hisexcellent technicalassistance, C. M.Prestonforhelping with the in
on November 10, 2019 by guest
http://jvi.asm.org/
NS POLYPEPTIDES OF VSV ts MUTANTS 333
vitrotranslation, and J. T. Poyner for helping with the prep-arationofthe manuscript.
D.E. was arecipient ofaScience Research Council scholar-ship for training in research methods.
LITERATURE CITED
1. Abraham, G.,and A. K. Banerjee. 1976. Sequential transcriptionof the genesofvesicular stomatitis virus. Proc. Natl. Acad. Sci. U.S.A.73:1504-1508.
2. Ball,LA.,andC.N.White.1976.Orderoftranscription ofgenesofvesicularstomatitis virus. Proc.Natl. Acad. Sci.U.S.A. 73:442-446.
3. Cancedda,R.,and M. J.Schlesinger.1974.Formation ofSindbis viruscapsidprotein inmammalian cell-free extractsprogrammed with viral messenger RNA. Proc. Natl.Acad. Sci. U.S.A. 71:1843-1847.
4. Emerson,S. U., and Y.-H. Yu. 1975. Both NS and L proteins are required for in vitro RNA synthesisby vesicular stomatitisvirus.J.Virol.15:1348-1356. 5. Freeman, G. J., J. K. Rose, G. M Clinton, and A.S.
Huang. 1977. RNA synthesis ofvesicular stomatitis virus.VII. Complete separation of the mRNA's of ve-sicularstomatitis virusby duplexformation.J. Virol. 21:1094-1104.
6. Heppel,LA., D.R.Harkness, and R. J. Hilmoe.1962. Astudyofthesubstrate specificity and otherproperties ofthealkalinephosphatase of Escherichia coli. J. Biol. Chem. 237:841-846.
7. Imblum,R.D.,and R. R.Wagner.1975. Inhibition of viral transcriptasebyimmunoglobulindirectedagainst the nucleocapsid NS protein of vesicular stomatitis virus. J. Virol. 15:1357-1366.
8. Knipe, D., J.K.Rose, andILF.Lodish 1975. Trans-lation of individual species of vesicular stomatitis viral mRNA.J.Virol. 15:1004-1011.
9.Lesnaw, J. A., and M. E. Reichmann. 1975. RNA synthesis bytemperature-sensitive mutants of vesicular stomatitisvirus, New Jerseyserotype.Virology 63:492-504.
10. Lowry,0.FL,N.J.Rosebrough, A. L Farr, and R. J. Randall. 1951. Proteinmeasurement with the Folin phenolreagent.J. Biol. Chem. 193:265-275.
11. Marsden, H. S.,T. K. Crombie, and J. H. Subak-Sharpe.1976. Control ofprotein synthesis inherpes virus-infectedcells:analysis of the polypeptides induced by wildtypeandsixteen temperature-sensitivemutants ofHSV strain17.J. Gen. Virol.31:347-372.
12. Moyer, S. A., andD.F.Summers. 1974. Phosphoryla-tionofvesicularstomatitis virus invivo and in vitro. J. Virol.13:455-465.
13. Obijeski,J.F.,A. T.Marchenko,D. H. LBishop,B. W. Cann, and F. A. Murphy. 1974. Comparative electrophoretic analysis of the virusproteins of four rhabdoviruses.J.Gen.Virol.22:21-33.
14. Pelham,IL R.B.,and R. J.Jackson.1976. Anefficient mRNA-dependent translationsystem fromreticulocyte lysates.Eur. J.Biochem.67:247-256.
15. Preston, C. M.,and J. F.Szilgyi.1977.Cell-free trans-lationof RNA synthesized in vitro by a transcribing
nucleoprotein complex preparedfrom purifiedvesicular stomatitis virus. J. Virol. 21:1002-1009.
16. Pringle, C.R. 1977.Geneticsofrhabdoviruses,p. 239-289.In H. Fraenkel-Conrat andR. R. Wagner (ed.), Comprehensive virology, vol. 9. Plenum Press, New York.
17.Pringle,C.R., I. B. Duncan, and M. Stevenson. 1971. Isolationandcharacterization of temperature-sensitive mutantsof vesicularstomatitis virus, New Jersey sero-type.J. Virol.8:836-841.
18. Purchio, A. F.,E.Erikson, andR.L. Erikson. 1977. Translation of35S and ofsubgenomicregions of avian sarcoma virus RNA. Proc. Natl. Acad.Sci. U.S.A.74: 4661-4664.
19. Reid, T. W.,and T. B.Wilson. 1971. E. coli alkaline phosphatase, p. 373-415. In P. D. Boyer (ed.), The enzymes, vol.4,3rd ed. Academic Press Inc., New York. 20. Rhodes,D.P., G. Abraham,R. J.Colonno,W.
Jeli-nek, and A.K.Banerjee. 1977. Characterizationof vesicularstomatitis virus mRNAspecies synthesizedin vitro. J.Virol. 21:1105-1112.
21. Simmons,D.T.,and J. H. Strauss. 1974. Translation of Sindbisvirus 26S RNA and 49S RNA inlysatesof rabbitreticulocytes. J. Mol. Biol. 86:397-409. 22. Sokol, F.,and H. F.Clark.1973.Phosphoproteins,
struc-tural components ofrhabdoviruses. Virology 52:246-263.
23. Stampfer, M.,and D. Baltimore. 1973. Identification of thevesicularstomatitis viruslarge proteinas aunique viralprotein. J.Virol.11:520-526.
24. Szilagyi,J.F., and C.RIPringle.1972.Effect of tem-perature-sensitive mutations on thevirion-associated RNAtranscriptaseofvesicular stomatitisvirus. J. Mol. Biol. 71:281-291.
25.Szilagyi,J.F.,andC.R.Pringle.1975.Virion transcrip-tase activitydifferences in host range mutants of vesic-ular stomatitis virus. J.Virol.16:927-936.
26. Szilagyi,J.F., and C.R.Pringle.1979.Effect of
tem-perature-sensitive mutation onactivity of the RNA transcriptase of vesicularstomatitis virus NewJersey. J. Virol. 30:692-700.
27.Szilagyi,J.F.,andLUryvayev.1973.Isolation ofan
infectious ribonucleoprotienfromvesicular stomatitis viruscontaininganactiveRNAtranscriptase.J. Virol. 11:279-286.
28. Wagner,R. R. 1975.Reproductionofrhabdoviruses,p. 1-93.In H. Fraenkel-Conratand R. R.Wagner(ed.),
Comprehensive virology, vol. 4. Plenum Press, New York.
29. Wagner,R.R., LPrevec,F.Brown,D.F.Summers, F. Sokol, and R.MacLeod. 1972. Classification of rhabdovirusproteins:aproposal.J.Virol.10:1228-1230. 30. Wunner,W.IL,andC.R.Pringle.1972.Protein syn-thesisin BHKcelLsinfected withvesicularstomatitis virus. II. ts mutants of the NewJerseyserotype. Virol-ogy50:250-253.
31. Wunner,W.H.,and C.R.Pringle.1974. A temperature-sensitive mutantofvesicular stomatitis virus withtwo
abnormal virusproteins.J. Gen. Virol. 23:97-106. VOL. 31,1979
on November 10, 2019 by guest
http://jvi.asm.org/

![FIG. 2.gradient[35S]methionine.E,tracks,arevirions and Virionpolypeptides ofthe ts mutants ofcomplementation group E](https://thumb-us.123doks.com/thumbv2/123dok_us/1510047.103671/4.504.114.404.72.414/gradient-methionine-tracks-arevirions-virionpolypeptides-ofthe-mutants-ofcomplementation.webp)