JOURNALoFVIROLOGY,May 1975,p.1049-1056 Copyright X 1975 AmericanSocietyforMicrobiology
Vol. 15, No. 5 Printedin U.SA.
Genetic
Analysis of Adenovirus Type
2
II.
Preliminary Phenotypic Characterization
of
Temperature-Sensitive
Mutants
J. WEBER,* M. BEGIN, AND G. KHITTOO
Departement deMicrobiologie, Centre Hospitalier Universitaire, Sherbrooke, Quebec, Canada Receivedfor publication 21 October 1974
The properties oftemperature-sensitive mutants ofadenovirus type 2
repre-senting12complementationgroups werestudied. Allmutantswerenormalwith
respect to adsorption as measured by viral inclusion formation and viral DNA
syntbeFsis asshownby velocity sedimentation in alkalinesucrosegradients. One
mutant, however, formed viral inclusions of altered morphology atthe
nonper-missive temperature. Thesynthesis of themajor capsid proteins wasexamined
by immunodiffusion. On this basis, the complementation groups could be arranged asfollows: (i) onegroup wasnegative for allthree proteins; (ii) three groups failed to synthesize penton bases; (iii) eight groups were positive for
hexons, pentons, and fibers. The assembly of virus particles at 39C was
examined by equilibrium sedimentation in CsCl; three groups were found
defective, whereas two of the penton-negative groups were positive for virion
production. Tests of the thermolability of virions at 50C revealed eightgroups
labile whereas theremainderwere insensitive toheat inactivation. Noneof five
mutantsinoculated in newbornratsinducedtumors, although three ofthemwere
capableof invitrotransformation.
In a previous communication we have
re-ported the isolation of temperature-sensitive
(ts) mutants of adenovirus type 2 (2). These mutants were classed into 12 complementation groups and arranged in a linear sequence on a tentative ge-ieticmap by recombination
analy-sis. In this report we examine some of the functional characteristicsof these mutants.
(Preliminary reports of this work were pre-sented atthe Annual Meeting of the American
SocietyforMicrobiology, Chicago, Ill., 12 to 17
May 1974, and at the Canadian Society of
Microbiologists, Montreal, 18to 21 June1974.)
MATERIALS AND METHODS
Virusandcels.Humanadenovirustype 2(Ad 2) designatedasthewild type(WT)waspropagatedin KBcells and titratedbytheplaqueassayonKB cells orby the infected cell unit assayonHEp-2cells, as
described previously (2, 19). The isolation ofthets mutants and their preliminary genetic characteriza-tion have also been described previously (2). In the interest of abiding by uniform nomenclature, the mutant numbers published in the preceding paper which are greater than 99 have been revised as follows:ts241is ts1; ts 496 is ts 2; ts 235 is ts 3;ts 500 is ts4; ts 458 ists5;ts155is ts6; ts 517 is ts 7; ts 166 is ts 8; ts 186 is ts 9; ts 231 is ts 11; ts 469 is ts 12. Henceforth the mutants shall be referred to by this newnomenclature, in conformity with other
adenovi-ruslaboratories (7).The KBcellswereobtainedfrom
Flow Laboratories (Rockville, Md.) and were never
usedbeyondthesixthpassage. BothHEp-2 andKB
cellswereculturedinDulbecco-modified,Eagle mini-mal essential medium containing 10% calf serum.
Virus-infected cells were culturedinminimal
essen-tial mediumsupplementedwith 2.5% calfserum and
0.4mM arginine.
DNAsynthesis.Confluent monolayersof KB cells grown in 50-mm diameter Falcon petri dishes were
inoculatedwith ts mutants at a multiplicity of input of 5 PFU/cell. After a 50-min adsorption period at 33 C, the cells were washed three times with
Tris-buffered saline (pH 7.4). Fresh medium was then added and the infected cultures wereincubatedat33 and 39C, then labeled with5
,Ci
of ['Hlthymidineper ml(40 to 60Ci/mM) 24 and 16 h later,
respec-tively. At 40 hpostinfection, the mediumof the cul-tures incubated at 39 C was removed, and the cells were washed with Tris-buffered saline, scraped off
with a rubber policeman, and resuspended in colQ
Tris-buffered saline. The cultures incubated at 33 C
received the same treatment 65 h postinfection. In someexperimentsthecellswere labeled for 1honly andeitherlysedimmediately or further incubated in medium without ['H]thymidine. Linear, 5 to 20% sucrose gradients (in 1.0 M NaCl, 0.01 M EDTA, and 0.3 M NaOH, pH 12.9) were prepared in poly-allomer centrifuge tubes essentially as described by
Doerfler (5). A lysing layer (0.20 ml) consisting of 0.5 MNaOH and 0.01 M EDTA was carefully applied ontop of thepreformed gradients. Samples of about 1049
on November 10, 2019 by guest
http://jvi.asm.org/
WEBER, BEGIN,ANDKHITTOO
10'
cells (contained in 0.05 ml) were mixed with[l'C
lthymidine-labeled marker virus and gently pipetted onto the lysing layer. The cells wereallowed to lyse for 1 h at room temperature or overnight at 4 C. The preparations were then centrifuged in the B60 International ultracentrifuge using the SB-405 rotor, at 60,000 rpm for 84min at 4 C. At the end of the run the gradients were collected dropwise on Whatman (GF-81) fiber glass filter disks from the bottom of the tubes. The filters were washed with ice cold 5% trichloroacetic acid solutionfollowed by ethanol, and then dried and counted in a toluenebasedscintillation fluid.
Immunodiffusion. Double diffusion in agar gels was carried outby a modification of the microtech-nique ofCrowle (4)and alsobythestandard ouchter-lony macrotechnique using the Gelman (Gelman
Instruments, AnnArbor, Mich.)apparatus(14).After
developmentfor1to2days,the slidesweresoakedin 0.15 MNaClfor 1 to 3days,then dried atadistance of 25 cm under a 250-W infrared lamp. To prevent
shrinkage during drying,the wells of the macrotech-niquegels were filled withwater.Theprecipitinlines werestained with amido black (1 g of stain in450ml of 1M acetic acid, 450ml of 0.1 M sodium acetate, and 100 ml ofglycerol).
Antisera. Whole virion antiserum waspreparedby
immunizingrabbitswith virions whichwerepurified by two extractions with fluorocarbon (Freon 113,
DupontCo.) followedbytwocyclesofcentrifugations
toequilibriuminCsCl. Hexon,penton,andfiberwere
purified by chromatography ofthe top components from CsCl gradients on DEAE-cellulose (9). The purity of these proteins were verified by sodium
dodecylsulfate-polyacrylamide gelelectrophoresis
(1)
and by immunodiffusion against whole virion anti-serum. Antisera against theseproteins was also
pre-pared by immunizing rabbits. The rabbit antisera wererecheckedby immunodiffusion.
Virusassembly.KB cell monolayers wereinfected with virus, incubated at 33C, andlabeledat 24hor
incubatedat 39C andlabeledat 18hafter infection with 1MCiof3H-labeledaminoacids mixture per ml
(International Chemical and Nuclear Corporation,
Irvine,Calif.).Thelabel was left on the cells until the cells were harvested inTris-buffered saline 2 and 3
daysafterinfectionat39and 33C, respectively, lysed,
and extracted with trichlorotrifluoroethane (Freon
113, Dupont of Canada, Ltd.). This material was gentlypipetted on top of a CsCl solution (1.40g/cm3)
and centrifuged to equilibrium overnight. The tubes were photographed; then fractions were collected through the bottom. The density of the CsCl was determinedbyrefractometry whereas the radioactiv-ity was determined as 5% cold trichloroacetic acid-precipitable counts in toluene based scintillation fluid. Thenegatives of the photographs were scanned at 540 nm in a Gilfordspectrophotometer equipped
with a linearscanning device.
A second series of KB cell cultures were infected with virus and incubated at 33, 39, and 40.5C, then labeled with 2 uCi of [8H]thymidine per ml (60Ci/
nM, NewEngland Nuclear) at 6, 4, and3h after in-fection, respectively. The labelwas left on the cells
until the end of the infection period at 3 days (33 C) and 2 days (39 and 40.5C) postinfection. The virus was extracted with Freon as above andtreated with 50
Ag
of pancreatic DNase I per ml (Calbiochem) at 37Cfor 30min,and then 0.1 ml was layered on top of a preformed CsCl gradient (1.2 to 1.5 g/cm3) and centrifuged at 60,000 rpm for 1 h in rotorSB-405 of an International B60 ultracentrifuge. Thefractions were treated as described above.RESULTS
Adsorption and inclusion body formation. Theefficiency of adsorption of the mutants was tested by the formation of inclusion bodies (15, 19, 20) at the permissive and nonpermissive temperatures. Single cover slips (1-cm diame-ter) with a confluent monolayer of HEp-2 cells wereplaced in separate petri dishes and prein-cubated at 33 or 39 C, and then 25
Al
of prewarmedvirus at several dilutions was added to corresponding cover slips and allowed to adsorbat 33 or 39C.After 1 h,unadsorbedvirus wasrinsed off, mediumprewarmed to 33 C was added, and all cover slips were incubated at 33 C for 3days. At this time they were fixed and stained,and the number ofvirus-induced inclu-sionbodieswascountedasdescribedpreviously (19). Since several dilutions of virus were tested, it was possible to choose a dilution which produced between 0.1 to 1% inclusion bodies (19), andtoread the cover slipscorresponding to this dilution at the two adsorption tempera-tures. The ratio of inclusion bodies at 33 C to inclusion bodies at 39C did not significantly deviate from unity for the 15 mutants tested (belonging to the 12complementation groups). As all the mutantsappeared normalfor adsorp-tion, the actual dataare not shown.In a second experiment we wished to deter-mine whether the mutants were capable of forming inclusion bodies at the nonpermissive temperature. The experiment was essentially conducted asabove, exceptthatadsorptionwas at 33C and incubation at both33and39 C.
As in the adsorption experiment, the ratio of inclusion bodies at 33 C to inclusion bodies at 39 C was notsignificantly differentfromunity. With the exception of ts 3, the morphologyof theinclusionswasalsoindependentof tempera-ture.Although at 33Cthe ts3-infected cellsare indistinguishable from the WT, incubation at 39C causes the cells to appearcrenated.
It was concludedthat these ts mutants were not defective in adsorption and inclusion body formation.
Viral DNA synthesis. The ability of the mutants to synthesize DNA at the nonpermis-sive temperature was determined by velocity
1050 J. VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
VOL.15.1975 PHENOTYPIC CHARACTERIZATION OF TS AD 2 MUTANTS
sedimentation of ['H
jthymidine-labeled
in-fected cell DNA in alkaline sucrose gradients.Under the conditions employed, viral DNA
from infected cells cosedimented with purified Ad 2 marker DNA, whereas cellular DNA was pelleted. Figure 1 shows typical profiles ob-tained for ts 53,74, 6, 8, 9, 11, 3, 1, 5, 2, 4, 7,and WT. Similar results were obtained when using the following modifications of the basic proce-dure: (i) thelabeling periodwith [8H
]thymidine
was reduced to 1 h and the cells were lysedimmediately or after a 24-h chase; (ii) the alkali lysis period was extended to overnightat 4 C;
(iii) the temperature ofincubationwasraisedto 40.5 C. In all cases there wasalways somewhat more DNA synthesized at the nonpermissive than at the permissive temperature. Reversion
can be ruled out since we are dealing with a single growth cycle and the rate of reversion was never greater than 10-'(2). Although leak-iness cannot be ruled out, it appears highly
improbable that DNA synthesis during a 1-h labeling period should be appreciably affected by it. From this result it may be concluded that these mutants are not defective in viral DNA synthesis.
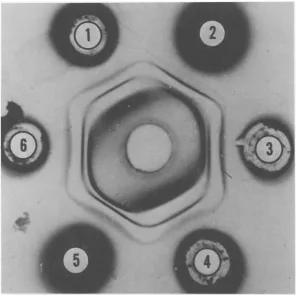
Synthesis of viral capsid antigens. Using the standard macroimmunodiffusion tech-nique, three distinct precipitin lines were
ob-tained with infected cell lysates and antiserum prepared againsthighlypurified completevirus
12
-10
x
*-0
6FRACTION
FIG. 1. Viral DNAsynthesis. Alkalinesucrose
gra-dient velocity sedimentation of
[3H]thymidine-labeledDNA from KBcellsinfected withts mutants.
This profile isrepresentative of all thets mutantsand WT tested. The arrow indicates the position of ["C]thymidine-labeled Ad2markerDNA. Symbols: A, ['H]thymidine counts/min labeled at 33C; A,
['H]thymidinecounts/min labeledat39C.
(Fig. 2). A fourth faint line was also present sometimes. By means of the microtechnique, two tothree additional, fainter precipitin lines canalsobedetected. The three majorprecipitin lines were identified by means of monospecific antisera to hexons and pentons (penton base andfiber). At the present time we do not know the nature of the extra precipitin lines. The ability of the ts mutants to synthesize immuno-logically cross-reactive material of the three major capsid antigens (hexon, penton, and fiber) was tested underpermissive and nonper-missive conditions by both these techniques. The results are summarized in Table 1. Itcan be seen that ts3 fails to synthesize any of the capsid antigens; ts 11, ts 4, and ts 7 representing three distinct complementation groups pro-duced normal precipitin lines correspondingto hexon and fiber antigens but reducedprecipitin
lines corresponding to pentons, whereas the remaining mutants representing eight com-plementation groups appear to synthesize nor-mal levels of theantigens.
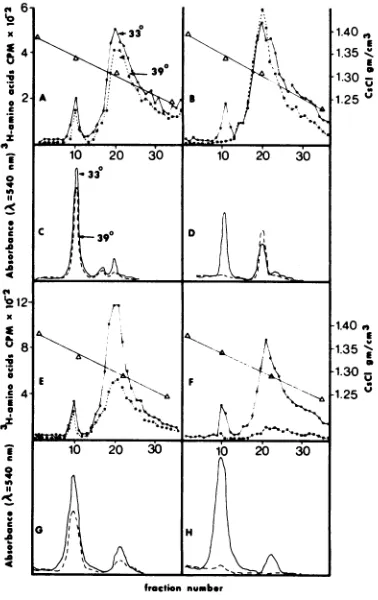
Virus assembly. The immunodiffusion ex-periments described above have demonstrated that some ts mutants were defective in the synthesis of one ormorecapsid antigens at the nonpermissive temperature, whereas the major-ity had apparently synthesized a pattern of antigens indistinguishable from the WT virus. Since the synthesis of cross-reactive material is noguarantee of the functionalintegrity of the antigen, northeabsence of cross-reactive mate-rial fortheabsenceofvirions, theproblemof the assembly of virus particles was examined by the following experiments. KB cell cultures weresingly infected with mutants or WTat 33 and 39 C and labeled with 'H-labeled amino acids, and the infected cell extracts were cen-trifuged to equilibrium in CsCl. The virus bands were photographed in the tubes, and then fractions were collected and measured for den-sity andradioactivity. Figure3shows the radio-activity profilesandthe densitometric tracings
of the photographs of three mutants and the WT. The profiles show thesharpvirus band in the 1.34 density region and a broader band of material which includes empty virions, virus
capsids (top components), and probably some cellularproteinsaround the 1.30density region (3, 8, 17). The disproportionate virus band in the densitometric tracings is due to the strong diffractionofvisible lightbyvirions. Of the mu-tants tested, ts 53, 74, 6, 8, 9, and 1 had pro-files identical toWT; ts 2, 7, and 11produced a normal amount ofvirus but a reduced quan-tity of top components; ts 4 and 5 produced 1051
on November 10, 2019 by guest
http://jvi.asm.org/
[image:3.505.49.247.411.584.2]WEBER, B1GIN,ANDKH1TTOO
FIG. 2. Doublediffusion inagar. Centerwell containsrabbit anti,erum againstAd2andperipheralwell's
extractsfromcellsinfectedwithvirus aisfollows:1and4,ts at39C; 2, WTat39C;3and6,ts at33C; 5,
at33C.
no virus but an apparently normal amount of
top components;andts3producedneither virus
nortop components. Since ts3was apparently
defectiveintheproductionofcapsid
cross-reac-tivematerial, it issatisfyingto notethe drastic reduction of both virionsandtopcomponents at
thenonpermissive temperature.Although ts4, 11, and 7 were defective in the production of
pentons,only ts4appearedalsotobedefective in virionproduction. The defects ints5 and 2
may involve a minor virion component or a
nonstructural virus function. The defects in
mutantswhichwereindistinguishable from the
WT will need to be investigated by other methods.
To test someof thepropositionsenumerated
above, the experimentwasrepeated withsome
modifications. Although the above experiment dealt with changes detected on the basis of
following the proteins, the present one was
designed tostudytheassemblyof nucleic acid-containing particles. Virus was labeled with [3H1]thymidine at33, 39, and 40.5 C and puri-fied asdescribed above. The radioactivity
pro-filesareshowninFig.4.Underthese conditions
ts1, 2, 6, 8, 9, 53, and74 wereidenticaltoWTas expected; ts 3 and 4 remained
unchanged
whereas ts 5, 11, and 7 (panel B) showed aconsiderabledrop invirion
assembly
or mainte-nance at 40.5C. This result appears to beconsistent with the notion ofleakiness ints 11
and7.
Heatinactivation. Figure 5gives the results of experiments testing the
thermolability
of mutant and WT virus particles at 50C. Theslopeoftheinactivationcurvevaried
depending
onthe stateofpurification andthesuspension
medium. To have uniform conditions, crude virussampleswerediluted with thesamebuffer and all viruses were tested at the same time. The mutants appeared to cluster in three groups: those indistinguishable from WT (ts 2,
4, 7, 8); those inactivated 10- to50-fold faster than WT (ts 1, 3, 5, 6, 9, 11); and those inactivated 1,000-fold faster than WT (ts 53,
74). To check if the survivors contained revert-ants, some plaques from the 32-min exposure were picked and replaqued at 33 and 39C; all were shown to be ts mutants. This result suggests that mutants of eight
complementa-V:i.
J. VIROL.
1052
on November 10, 2019 by guest
http://jvi.asm.org/
[image:4.505.106.402.67.362.2]VOL. 15, 1975 PHENOTYPIC CHARACTERIZATION OF TS AD 2 MUTANTS
TAau 1. Immunodiffusionoflysates of cells infected
with ts mutants at 39Ca
Complementa- Mutant Virus capsid
antigens
tion group Hexon Fiber Penton
J ts 3
E tal11
F ts1 (+) (+) (4)
C ts 7~
B ts53
H ts 74
H ts 26
G ts 6
L ts 8
Ats 9 (+ (+ ()
D tal1
D tsl12
I tas
K ts2
Unassigned ts 10
aThisrepresents aqualitativesummaryofseveral
experiments using both the macro- and microim-munodiffusion methods with complete virus anti-serum and monospecific antiserum to hexon and pentonantigens.
tion groups
(A, B, D,
E,
G, H, I, J)
mayhave defects in genes for structural components of the virion.Tumorigenesis. WT Ad 2 has been shown tobeoncogenic
indirectly by
thetransplantationof in vitro transformed cells(6). To our
knowl-edge there exists no recent report to induce tumors by direct inoculation ofthe virus into
newborn rodents. To fill this gap, and more importantly to test the oncogenicity of ts mu-tants,newbornrats(Sprague-Dawley, Madison, Wis.) were inoculated with WT and 5 ts mu-tants. At about6 to 8 weeksof age, and also 1
week later, each rat received intramuscular injections of 2.5 mg of cortisone acetate as an immunosuppressive. The rats were palpated weekly for the presence of tumors up until a maximum of 14 weeks of age. Although the
rat's body temperature of 38.2 C closely
ap-proaches the nonpermissive temperature, in
this experiment neither the WT nor the ts mutants induced tumors at the site of inocula-tion (Table 2). Preliminary results of trans-formation experiments using primary cultures ofrat embryo cells indicate that WT andts 1, 3, and4arecapable oftransformation ateither 33or39 C. Thereasonforthe failure ofinvivo transformation remains unclearatpresent.
DISCUSSION
The present study showed that Ad 2 ts mutants representing 12 complementation
[image:5.505.51.245.79.302.2]groups were all defective in late functions.
Table 3presentsasummaryof the properties of the mutants studied. All of the mutants ad-sorbed normally, synthesized viral DNA, and produced a cytologically recognizable intranu-clear inclusion body. The morphology of the inclusion ofts3 atthenonpermissive tempera-ture was atypical. This was also the only mu-tantwhich failedtoproduce hexon,penton,and fiber antigens as evidenced by immunodiffu-sion. Working with Ad 31tsmutants, Suzukiet al. (18) have observed similar alterations in inclusion body morphology, accompanied by a reduction in the accumulation of the capsid
antigens. The drastically altered phenotype of ts 3 however is not likely due to a multiple mutational event as evidenced by the com-plementation, recombination, and reversion tests (2).
Immunodiffusionanalysis also revealed three complementation groups which showed reduced accumulation of the penton antigen, whereas
E
Pv"I
u
; 3
I I v
3
frocton number
FIG. 3. Radioactivity and absorbance profiles of cesium chloride equilibrium gradients ofextracts of
cells infected in the presence of 'H-labeled amino acids with WT and ts mutants at the permissive and
nonpermissivetemperatures.AandC, WT, ts 53, 74, 6, 8, 9, 1; B andD,ts4and5; E andG,ts11, 2, 7; F andH,ts 3. - , 33C; ---, 39 C.
1053
on November 10, 2019 by guest
http://jvi.asm.org/
[image:5.505.260.449.314.614.2]WEBER,
BgGIN,
ANDKHITTOOE
I.-,E
.
, U
froaion number
FIG. 4. Radioactivity profiles of cesium chloride equilibrium gradients ofextracts ofcellsinfectedin
the presence of [3H]thymidine with WT and ts
mutants atthepermissiveandnonpermissive temper-atures.Panel A:ts1, 2, 6, 8, 9, 53, 74,and WT.Panel B:ts5, 7, 11. Panel C:ts4. Panel D:ts3.Symbols: *, 33C; *, 39 C; A,40.5 C.
normal levels of hexon and fiber antigen were
present. The precipitin line corresponding to
the penton cross-reacts with that of the fiber
and has been showntocontaintheentirepenton capsid, consisting of penton base and fiber(9). Since the fiberantigen ispresentineach ofthe three mutants, and current data suggests that the penton base may consist ofmore than one copy of a single molecular weight class of
polypeptide (10), the existence of three
com-plementationgroupsappearstobeanintriguing
finding. Two of these complementation groups mayinfact be involved withregulatoryaspects
affecting the synthesis, processing, or
metabo-lism ofpenton bases.Alternatively, wemaybe dealing with intracistroniccomplementation,or
multiple-step mutants. The first possibility is
unlikely as the complementation indices were
greaterthan10for thecombinationsts 11x ts4 orts11 x ts7and, though that betweents4 x
ts7wasonly2,thesemutantsmapped distantly byrecombination.The genetic evidence and the highrateof reversion (2) alsoargueagainstthe
second possibility. Russell et al. (12, 13) have demonstrated six complementation groups
which affect hexon synthesis, and three
com-plementation groups which affect fiber
synthe-sis, whereas only one group affected penton
synthesis.
Of the three mutants defective in penton
synthesis, onlythe leastleakyone, ts4,wasalso
10~~~~~~~~~N
8 3
U~~~ ~lUE AT 50°
10
ts4
ts7
86316s32
FIG. 5. Heat inactivation of virions of ts mutants.
Crudesamples of virus grown at 33 C were sonically
treated to disperse possible aggregates, and diluted
1:10 in Thris-buffered saline, pH 7.4. Samples were
exposed to 50 C for the indicated times, then cooled
rapidlyin an ice bath. Survival was assayed by plaque
formation at 33 C.
TABLE 2. Attempts to inducetumors in rats with WT
and tsmutantst
Virus dose No. ofbaby Number of Virus (PFU/rat) rats injected free
I017_10 25 13-14
ts 1 107 27 9
ts 6 107 8 8
ts 4 106 8 11
ts 3 107 5 10
ts74 106 15 10
aNewborn litters were injected subcutaneously
with0.1ml of virus within 24hofbirth and checked for tumorsweekly. At about6 to8weeks of age and1 weeklater, eachratreceivedtwointramuscular injec-tions of 2.5mg of cortisone.
1054 J. VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:6.505.71.253.76.394.2] [image:6.505.263.461.219.414.2]VOL.15, 1975 PHENOTYPIC CHARACTERIZATION OF TS AD 2 MUTANTS TABLE 3. Properties ofts mutantsoftype2adenovirusa
DNA Inclusion Antigensproduced (39C) Virion Thermo- Invivo Invitro
Mutant synthesis assembly sensitivity tumor for-
-(3 (39C)
C
)
Hexon Fiber Penton 39 or (50C) mation(m3a9tC)
H2tsJ 3 1 + + + +
H2tsE 1 1 + + + + 4 4 + NT NT
H2tsF 4 + + + + 4 _- _ +
H2tsC 7 + + + + 1 - NT NT
H2tsB 53 + + + + + + + NT NT
H2tsH 74 + + + + + + + _ NT
H2tsH 26 NT + + + + NT NT NT NT
H2tsG 6 + + + + + + + _ NT
H2tsL 8 + + + + + + _ NT NT
H2tsA 9 + + + + + + + NT NT
H2tsD 1 + + + + + + + _ +
H2tsD 12 NT + + + + NT NT NT NT
H2tsI 5 + + + + + 4 + NT NT
H2tsK 2 + + + + + + _ NT NT
H2ts 10 NT + + + + NT NT NT NT
WT + + + + + +
_
_
+aAbbreviations: NT, nottested;H2, human type 2adenovirus; J,E,F, etc., denoted the complementation
group.
defective in virion assembly at 39C. The two
leaky mutants, ts 11 and ts 7,
only
showedreduced assembly at40.5 C.
The assembly of virions in the absence of
adequate numbersofpentons isnot as surpris-ing in view ofthe experiments of Prage et al. (11) which showed that virions lacking pentons and even peripentonal hexons arestableinCsCl
gradientsand canbe visualizedbymeansofthe electron microscope. Further phenotypic sepa-ration of these mutants was obtained by the
demonstration of thethermolabilityofonly one ofthese three mutants. Increased thermolabil-ity of the mutant virions has been taken to suggest thedefectivenessofastructural compo-nent or aprotein implicated in the determina-tion of the structure of the virion. Unaltered
lability however, fails to indicate if the ts lesion is in astructural or a nonstructuralprotein.By this criterion, ts 3 and 5 are twofurther com-plementation groups which fail to produce vi-rions andareprobably defective inastructural protein. Intotal, eight complementation groups
showed increased thermosensitivity, thus sug-gestingeightcistronsspecifyingproteins
impli-cated in the architecture of the virion. The remaining four complementation groups failed to show increased thermolability. According to currentdata(1), Ad 2maysynthesize upto 14 molecular weight classes of structural proteins and perhaps seven nonstructural proteins. As-sumingthattheseproteinsareindeed codedfor
byAd2, the total molecularweight of 920,000of these21proteinsaccountsfor80% ofthecoding
capacity of the virus. The 12 complementation groups discussed in this paper may represent
about half of the total genetic complement of Ad 2.
ACKNOWLEDGMENTS
The technical assistance ofKen Harrison and in part Diane Duranleauare gratefully acknowledged.
J.W. is ResearchScholarofthe Medical Research Council of Canada. M.B. is recipient of a studentship from the Conseil de RecherchesM6dicales duQu6bec.Thiswork was supported bygrants from the National Cancer Instituteof Canada and MA-4164 from the MedicalResearch Councilof Canada.
LITERATURE CITED
1. Anderson, C. W.,P. R.Baum,andR.F.Gesteland.1973. Processingofadenovirus 2-induced proteins.J.Virol. 12:241-252.
2. Begin, M., and J. Weber. 1975. Genetic analysis of adenovirustype2. I.Isolation andgenetic characteriza-tion oftemperature-sensitivemutants.J.Virol.15:1-7. 3. Burlingham, B. T., D. T. Brown, and W. Doerfler.1974.
Incomplete particles of adenovirus. I. Characteriza-tion of the DNA associated with incomplete adeno-virionsoftypes2 and 12.Virology60:419-430. 4. Crowle,A.J. 1958. Asimplified micro double-diffusion
agar precipitin technique. J. Lab. Clin. Med. 52:784-786.
5.Doerfler, W. 1969. Nonproductive infection of baby hamsterkidney cells(BHK 21)with adenovirustype
12.Virology38:587-606.
6. Gallimore,P. H.1972. Tumorproductionin immunosup-pressedratswith cellstransformedinvitroby adenovi-rustype2.J.Gen.Virol.16:99-102.
7. Ginsberg, H.S.,J. F.Williams,W. H.Doerfler,and H. Shimojo. 1973.Proposednomenclature for mutants of adenoviruses. J. Virol. 12:663-664.
8; Ishibashi, M., and J. V.Maizel, Jr. 1974. The polypep-tides ofadenovirusV. Youngvirions,structural
inter-1055
on November 10, 2019 by guest
http://jvi.asm.org/
WEBER,
B.AGIN,
ANDKHITTOOmediate between top components and aged virions. Virology 57:409-424.
9. Pereira, H. G. 1960. Antigenicstructureof non-infectious adenovirus materials. Nature (London) 186:571-572. 10. Philipson, L., and U. Pettersson. 1973. Structure and
function of virion proteins ofadenoviruses. Prog. Exp. TumorRes. 18:1-55.
11.Prage, L., U. Pettersson, S.Hoglund, K. Lonberg-Holm, and L. Philipson. 1970. Structural proteins of
adenovi-ruses. IV. Sequential degradation of the adenovirus
type2virion.Virology 42:341-358.
12. Russell, W. C., C. Newman, and J. F. Williams. 1972. Characterization oftemperature-sensitive mutantsof adenovirustype5-serology. J.Gen. Virol. 17:265-279. 13. Russell, W. C., J. J. Skehel, and J. F. Williams. 1974.
Characterization oftemperature-sensitive mutantsof adenovirustype5:synthesis of polypeptides in infected cells. J. Gen. Virol. 24:247-259.
14. Salzmann, N. P., and B. Moss. 1969. Analysis of radioac-tively labeled proteins by immunodiffusion,p.327-333.
In K. Habel and N. P. Salzmann (ed.), Fundamental techniques in virology. Academic Press Inc., New York.
15. Shahrabadi, M. S., K. L. Roy, and T. Yamamoto. 1972. Macromolecular content of inclusion produced bya
canineadenovirus. J. Virol. 10:801-809.
16. Shiroki, K., H. Shimojo, and K. Yamaguchi. 1974.The viral DNA replication complex of adenovirus12.
Virol-ogy60:192-199.
17. Sundquist, B., E. Everitt, L. Philipson, and S. Hoglund. 1973. Assembly of adenoviruses. J. Virol.11:449-459. 18. Suzuki, E., H. Shimojo1 and Y. Moritsugu. 1972. Isolation
and prelimary characterization of temperature-sensi-tivemutantsof adenovirus31.Virology49:426-438. 19. Weber, J. 1972. Titration of adenovirus by counting
cells containing virus-inducedinclusion bodies. Appl. Microbiol. 23:1025-1026.
20. Weber, J., and H. F. Stich. 1969. Electron microscopy of cells infected with adenovirus type 2. J. Virol. 3:198-204.
1056 J. VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
![FIG.1.A,labeleddientThisWT["C]thymidine-labeled['H]thymidine Viral DNA synthesis. Alkaline sucrose gra-velocity sedimentationof [3H]thymidine- DNA from KB cells infected with ts mutants](https://thumb-us.123doks.com/thumbv2/123dok_us/1572293.109829/3.505.49.247.411.584/labeleddientthiswt-thymidine-thymidine-synthesis-alkaline-sedimentationof-thymidine-infected.webp)
![FIG. 4.atures.B:33equilibriummutantsthe ts C; Radioactivity profiles of cesium chloride gradients of extracts of cells infected in presence of [3H]thymidine with WT andts at the permissive and nonpermissive temper- Panel A: ts 1, 2, 6, 8, 9, 53, 74, and WT](https://thumb-us.123doks.com/thumbv2/123dok_us/1572293.109829/6.505.263.461.219.414/equilibriummutantsthe-radioactivity-gradients-extracts-presence-thymidine-permissive-nonpermissive.webp)