Copyright© 1976 American SocietyforMicrobiology PrintedVol.in19,U.S.A.No.3
Isolation
of a
Transcriptive
Complex
from
Newcastle Disease
Virions
RICHARD J. COLONNO' AND HENRY 0. STONE*
DepartmentofMicrobiology, University of Kansas, Lawrence, Kansas66045
Received for publication 13August 1975
An activetranscriptive complex was isolatedfrompurifiedvirions of Newcas-tledisease virus. Afterdisruption withTritonX-100andhigh salt, soluble and
particulate fractions were separated by density gradient centrifugation. The transcriptive complex, recovered at a density of 1.275 g/cm:, appeared as a
nucleocapsid structure by electron microscopy. When analyzed by polyacryl-amide gel electrophoresis, the nucleocapsids consistedofthe nucleocapsid pro-tein, a minor protein of 53,000 molecular weight, and the large L protein.
Nucleocapsids possessed less than 1% ofthehemagglutinating and neuramini-dase activities originally associated with virions. The active complex synthe-sizedpredominantly11to20S RNAinvitroandapproximatelyone-fourthofthe
RNA molecules contained polyadenylic acid segments. In the presence of
S-adenosyl-L-methionine,
the RNAmolecules werecapped andmethylatedatthe 5' termini. The transcriptive complex was also capable ofmethylating exoge-nousEscherichia coli RNAinthe absence ofviral RNAsynthesis.Newcastle disease virus (NDV) is a large, enveloped RNA virus (16) that contains a
sin-gle-stranded 50S genome (8, 16, 19) encapsi-dated bynucleocapsid protein (3, 13, 15, 21, 27, 31),designated NP protein (30). The virion
con-tains two other major proteins, a membrane protein (M) and aglycoprotein (HN), shownto
contain both neuraminidase and hemaggluti-nating activities (29). Several minor proteins havealso been detected (3, 13, 15, 27, 28).
Purifiedvirions contain a transcriptase that
transcribes the50Sgenome RNA into 18Sviral
RNA, which is complementary in base
se-quence to the genome and contains
polyade-nylic acid lpoly(A)] (18, 33). Marshall and
Gil-lespie (23) have shown that NDV genomes do
not contain stretches of polyuridylic acid
Ipoly(U)] to code for the poly(A),
suggesting
a post-transcriptional addition of poly(A) by a virion-associated enzyme.In addition to poly(A) synthesis, we have
recently demonstrated that purified virions of NDV are capable ofcapping and methylating
the 5' terminus ofviral mRNAsynthesized in
vitro (6; R.Colonno andH. Stone, Nature [Lon-don], in press), indicating the presence of a
methyl transferase and a capping enzyme.
Thus the virion may contain as many as four
enzymes that arerelated to RNAsynthesis. Scheid and Choppin (30) have been able to
solubilize the M and the HNprotein free from
Present address: Department of Cell Biology, Roche InstituteofMolecularBiology,Nutley, NJ 07110.
the virion
by disrupting
purified
NDV withTriton X-100
detergent
andhigh
salt.Meager
and Burke (26)
disrupted
NDV with TritonN101
alone,
but when viewedby
electron mi-croscopy the subviralparticles produced
ap-peared
tobespikeless
virions. Intheabsenceofhigh
salt,
thedetergent
releasedonly
the HNglycoprotein
from the virion.Nucleocapsids
prepared
by
this and othertechniques
did notretainRNA
polymerase
activity
(26).Using
theprocedure
ofScheid andChoppin (30),
Marxetal. (24)isolateda
nucleocapsid
structure(tran-scriptive
complex)
from Sendai virus thatre-tained the
ability
tosynthesize
RNAinvitro. We report in this communication that anactive
transcriptive complex
can also be iso-lated from NDVby
Triton X-100andhigh salt,
which
synthesizes
complementary RNA andalso retains the
ability
topolyadenylate,
cap,and
methylate
viral RNAin vitro.MATERIALS AND METHODS
Virus purification. NDV wasgrown in
embryo-nated hen eggs (14) andpurified as previously
de-scribed (6). Virus wasthen resuspended in 0.01 M
Tris (pH 7.3)-0.03 M NaCl and stored at -70°C. Protein concentrations were determined according totheprocedureofLowryetal.(22).Purified virions were labeled in vitro using ['4C]formaldehyde and
sodium borohydride asdescribed byMcMillen and
Consigli (25).
Isolation of transcriptive complex. Virus at a
finalproteinconcentration of 1mg/mlwasdisrupted in 5ml of1% (vol/vol) Triton X-100 (NewEngland 1080
on November 10, 2019 by guest
http://jvi.asm.org/
Nuclear Corp.)-0.75 M KCl-0.01 M Tris-hydrochlo-ride (pH 7.3) for 20 minat20°C. The nucleocapsid
structure (transcriptive complex) waspurified free
ofsolubilizedproteinsbyisopycnicbandingina7-ml
linear gradientofD20glycerol (1.17 g/cm3)-D20
su-crose (1.37 g/cm3)buffered with 0.01 M
Tris-hydro-chloride (pH 7.3)-0.03 M NaCl. The gradientswere
centrifuged inaBeckmanSW41rotor(8°C, 90 min, 37,000rpm). Fractionationwasperformed either by an ISCO gradient fractionator (Instrumental
Spe-cialities Co.) ormanually. In the latter case,
pro-teinsthatwere solubilized by Triton and high salt wereobtained byremovingthetop5mlof the
gra-dient. The transcriptive complex, visible as a very
sharpband,waseasilyrecovered ina0.3-to0.4-ml volume with a 1-ml syringe containinga 20-gauge
needle. Solubilizedproteinswerelaterisolated free
ofthe detergent solution by butanol ether precipita-tion asdescribed byScheid and Choppin (30).
Electron microscopy. Virionsorthe transcriptive
complexataprotein concentrationof 0.1to0.5mg/ ml were appliedto a Parlodion-carbon coated grid
for 1 min andnegatively stained with 2% (wt/vol) uranyl acetatefor 30s as described by Home (17).
The negatively stained preparationswereexamined
inaPhillips 300 electron microscope.
Slab gel electrophoresis. A 10% (wt/vol) poly-acrylamide gel containing 0.13%(wt/vol) N,N'-bis-methylene acrylamide, 0.4 M Tris-hydrochloride (pH 8.7),and1%(wt/vol) SDSwaspolymerized with
0.03% (wt/vol) ammonium persulfate-0.03% (vol/ vol)tetramethylethylenediamine, and usedtopour aseparating gel (0.15 by10by16cm).Astacking gel composed of 5% (wt/vol)polyacrylamide,0.26% (wt/ vol) N,N-bis-methylene acrylamide, 0.06 M Tris-hydrochloride (pH 6.8), and 1% (wt/vol) SDS was
prepared, polymerized in the same manner, and
usedtoform 0.5-cmstacking gel.
Gelelectrophoresis wascarriedoutusing aslab gel box (Hoffer model SE 500) anda powersupply
(Buchlermodel3-1014A). Running buffer contained 0.05MTris-hydrochloride (pH 8.3), 0.38M glycine, and0.1%(wt/vol) SDS.Proteinsamples, containing
10to25,g of proteinper5to25 pl,2%(vol/vol)
f-mercaptoethanol, 1% (wt/vol) SDS, and 20% (vol/ vol)glycerol,wereplaced inaboilingwaterbathfor 1minbeforeloadingontheslabgel. Sampleswere
subjected to electrophoresis at 15 mA through the 5% stacking gel and at 30 mA through the 10%
separating gel until the bromophenol bluemarker
reached the bottom of the gel (about4h). The gels
werethenstained for 3 h with0.1%Coomassie
bril-liant blue-7% (vol/vol) acetic acid-25% (vol/vol) methanol and destained for16h withseveral
wash-ingsof 14%(vol/vol)aceticacid-50%(vol/vol) metha-nolbeforedrying.
RNA synthesis and purification. Standard
tran-scriptaseassays (0.1 ml) contained50 mM Tris-hy-drochloride (pH7.3), 120 mMNaCl,0.4mMMnCl,,
0.015% (vol/vol) Triton N101, 3 mM dithiothreitol,
0.7mMCTP,0.7 mMGTP,0.7mMUTP(reducedto 0.09 mM when [3H]UTP [21 Ci/mM] was used as
label), 0.7 mM ATP (reduced to 0.9 mM when [a-32P]ATP [15.3 Ci/mM] was used as label), and 1.5
ACi of[3H]UTPor 1 ,Ci of [a-32P]ATP. When the
transcriptive complex was assayed, the NaCl was reducedto 30 mM and Triton N101waseliminated. Methylation assay conditions were identical except that the 50 mMTris-hydrochloride (pH 7 .v-asused in place ofTris-hydrochloride (pH7.2') X - al pH in
each case was the same), and 0.78 A ,Ci) S-adenosyl-L-[methyl-3H]methionine (1 -C. Ci/mmol) was added. Allreaction mixtures contained 25 to 45
,Agofprotein (purified virions ortranscriptive com-plex) and wereincubated at 32°C for 6 h. Labeled product RNA was phenol-SDS extracted and puri-fied as previously described (6).
Analysis of in vitroproduct RNA. Procedures for rate zonal centrifugation, RNase digestions, near-est-neighbor analysis, DEAE chromatography, poly(U)-cellulose chromatography, and hybridiza-tions have been described previously (6, 7).
Hemagglutination titration. Tritrations, using 2.5 ,ug of protein (intact virions,solubilized protein, ortranscriptive complex), were performed at4°Cfor 2 h asdescribed by Choppin (5).
Neuraminidase assay. Samples containing 2.5 ,ug of protein (solubilized protein, intact virions, or transcriptive complex) were assayed, using neura-minlactose (Sigma) as a substrate, for 50min at 37°C asdescribed by Scheid and Choppin (30) and Ami-noff (2). Oneneuraminidaseunit was defined as the amount ofenzyme that releases 1 nmol of N-acetyl-neuraminic acid in 1 min at 37°C.
RESULTS
Isolation of transcriptive complex.
Experi-ments were performed to identify the virion
proteins and the structural components of the virionthatare involved in RNA synthesis. To
provide a sensitive and convenient assay for
viral protein,purified NDVwaslabeledin vitro
with
['4C]formaldehyde
(25). Purified virionswerethendisrupted with Triton and high salt, and the solubilized and particulate fractions
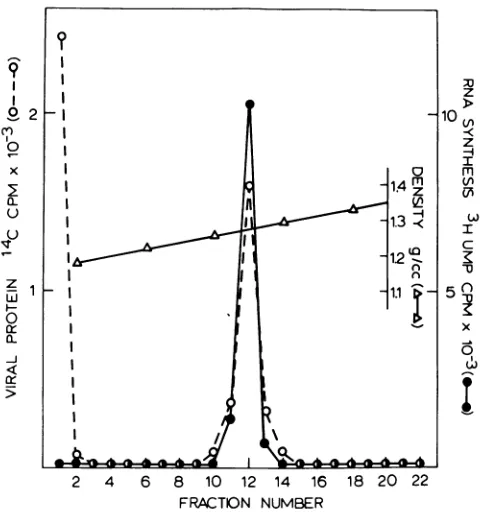
wereseparated by centrifugationon aD.O glyc-erol-sucrose gradient (Fig. 1). After fractiona-tion,all of the '4Cradioactivelabelwaslocated either in the solubilized fraction at the top of the gradient or ina sharpband having a den-sity of1.275 g/cm:1. To determineifthe
virion-associated transcriptase resided in either of
these peaks, portions of each gradient fraction
were assayedfor [H]UITPincorporation in an invitroRNA polymeraseassay.Asseen inFig.
1, all of therecovered polymerase activity was
associated with the structure havingadensity of1.275g/cm3,which will bereferred toasthe transcriptive complex. The specific activity of
thetranscriptivecomplexvaried between prep-arations, but always exhibited a 2- to 20-fold
increase in specific activity as compared with virions(datanotshown).
Since the transcriptive complex appeared to have the same densityaspurified nucleocapsid
(24, 29, 33), it was of interest to confirm this
on November 10, 2019 by guest
http://jvi.asm.org/
a-1
9
U z w
x
F-0 :
a-z
10LA
z
I m
(0
I C
K
u 50
i
xA-2 4 6 8 10 12 14 16 18 20 22 FRACTON NUMBER
FIG. 1. Isolation ofa transcriptivecomplex fromNDV.Purifiedvirions,disruptedwith1%(vollvol)Triton X-100and 0.75 MKC1, wereisopycnically banded in aD/Oglycerol-sucrose gradient. After fractionation, samples (20pi)of eachfraction (0.4 ml) were assayed for [3H]UTP incorporation (*) in an RNA polymerase assay. Aftertrichloroacetic acid precipitation, the samples were counted under double label conditions to simultaneously determine the amountof RNAsynthesizedand the amountof14C-labeled proteins (0) in the sample.
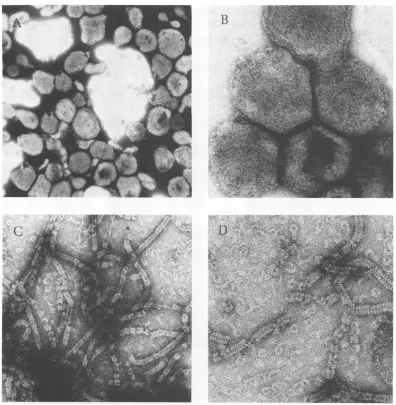
observation using electron microscopy.
Elec-tron micrographsofintact virions (Fig. 2A, B)
andisolated transcriptive complex (Fig. 2C, D) indicated that theactivetranscriptive complex
was indeed a nucleocapsid structure.
Treat-mentwithTriton-high salt completelyremoved theoutermembrane of the virion, and the
nu-cleocapsid was no longer confined within a tightlypacked corestructure.
Polypeptides in the transcriptive complex. Virions, transcriptive complex,and solubilized
proteinswerenextanalyzedby gel electropho-resis (Fig. 3). Slab gel electrophoresis was
foundtogive better resolution of proteins than
standard disc gels, revealingasmany aseight bandsinintact virions.Besidesthe threemajor proteins, HN, NP, and M, several minor pro-teinswerefoundincludingthelarge L protein.
After Triton-high salt disruption, most ofthe
HNandMproteins,alongwithpartofthelarge
L protein and a minor protein having an
ap-proximate molecular weight of 50,000, were
found in the solubilized fraction. The isolated
transcriptive complex, which retained the abilty to synthesize RNA, contained
predomi-nantlythe NP, a minor protein having an
ap-proximate molecular weight of 53,000, and a
reduced amount of the large L protein. Trace
amounts of HN andM proteinwere also pres-ent.
Enzymatic activities of solubilized
pro-teins. Bothneuraminidaseand
hemagglutinat-ing activitieshave been showntobeassociated
with theglycoprotein HN (30).Todemonstrate that the transcriptive complexwas free of the HNprotein,solubilizedproteinsand transcrip-tivecomplexwereextracted withbutanol-ether
andresuspendedatidenticalprotein
concentra-tions.Equalamountsofeach,along with intact
virions, were assayed for both neuraminidase and hemagglutinating activities. Results (Ta-ble1)indicate that thesolubilized fraction
con-tained96Uofneuraminidaseper mgofprotein
compared with 0.1 for the transcriptive
com-plex, and1.6 x 106hemagglutinatingunitsper mg ofprotein compared with 2.4 x 103 forthe
isolated complex. Thus,treatmentwith Triton-high salt left the transcriptive complex
vir-tually devoidof any enzymatic activity
associ-ated with the HNprotein.
Size of RNA product synthesized by
tran-scriptive complex. RNAwassynthesizedinthe presence of [a-32P]ATP to determine the
sedi-mentation properties of RNA synthesized in
on November 10, 2019 by guest
http://jvi.asm.org/
[image:3.505.145.385.75.331.2]FIG. 2. Electronmicrographs of (A,B) NDV virions and(C, D)thetranscriptivecomplex.Allspecimens
werenegatively stained with uranyl acetate. (A, x42,000; B, x132,000; C, x73,000; D, x127,000)
vitroby the transcriptivecomplex. The result-ingproduct RNAwaspurified byphenol
extrac-tionand Sephadex chromatography before
sedi-mentation on a 15 to 30% (wt/wt) sucrose
gra-dient (Fig. 4). Results indicate that the RNA synthesized by the transcriptive complexis
het-erogenous insize, rangingfrom2to28S,with a majority ofthe RNA sedimenting in the 11 to 20S region of sucrose gradients. The smaller
peakofRNAsedimenting at6S may be due to
fragmentation of the nucleocapsid structure
(Fig. 2C and D), resultinginapretermination of nascent RNA chains duringRNA synthesis.
Thus, the majority of RNA synthesized by the transcriptive complex is slightly smaller than
the predominant 188 mRNA species synthe-sized by purified virions (6) or isolated from NDV-infected cells (18).
Presence of poly(A) on viral mRNA. Since virions of NDV synthesize complementary
RNA-containing poly(A) segments (33), we nextdetermined whether theabilityto
synthe-size poly(A) was retained in the transcriptive complex. RNA wassynthesizedinvitroby viri-ons and by the transcriptive complex in the presence of either [3H]UTP or [a-32P]ATP.
After purification, the labeled product RNA
wasassayedfor the presence ofpoly(A) by enzy-maticdigestionwithRNaseA + T1, using 14C-labeled synthetic poly(A) and 28S ribosomal
on November 10, 2019 by guest
http://jvi.asm.org/
[image:4.505.54.450.75.480.2]WHOLE
SOLUBLE
VIRON
PROTEINS
ACTIVE COMPLEX
FIG. 3. Polyacrylamide slabgelelectrophoresisofNDV polypeptides. Virionsweredisruptedwith
Triton-high salt, andthe resultingtranscriptive complex was isolatedby isopycnic banding.Solubilizedproteins wereisolated free ofTritondetergent bybutanol-etherprecipitation beforegelelectrophoresis.Protein samples
(10to25ugin5to20pi)wereheatdenaturedin1%(wtlvol)SDS,2%(vollvol),3-mercaptoethanol, and20%
(vollvol) glycerol,and werethensubjectedtoelectrophoresis on a10%SDS-polyacrylamideslab gel.
TABLE 1. Assay forneuraminidase and hemagglutinating activities
Samplea Neuraminidase
(UlXa10t2]/mg
(U/mgpotein) protein)Intact virions 52 324
Solubilized pro- 96 16,000
teins
Transcriptive <0.1 24
complex
" Proteinsamples contained 2.5ggof protein in25
,A of 10 mM Tris-hydrochloride (pH 7.3)-30 mM NaCl.
RNA as controls. Results (Table 2) show that the AMP-labeled product RNA of both intact
virions and the transcriptive complex contain
8% and 11% RNase resistance, respectively, compared with the 100% RNase resistance of
synthetic poly(A). Both UMP-labeled product
RNAand28SribosomalRNA showedlessthan
1% RNase resistance. All the RNA
samples
were found to be totally sensitive to digestion withRNaseT2, indicatingthat eachradioactive label residedin RNA.
A further indication of the presence of poly(A)onviral RNAtranscriptswasobserved upon passage of the labeled RNA through a
poly(U)-cellulose column (Table 2). Controls showedthat 100% of the synthetic poly(A)and 0.5% of the 28S ribosomal RNAwereretained by the poly(U)-cellulose column. When [32P]AMP-labeled product RNAwasdenatured
andchromatographed, 17% of the RNA synthe-sized by virions and 24% of the RNA
synthe-sized by the transcriptive complex were
re-tained by the column. Proofthat the poly(U)
column was separating moleculeson thebasis
of the presence of poly(A) was obtained after
digestion of bound and unbound RNA with
RNase A + T,. More than 96% of the RNase
resistance resided in RNA that bound to the
NP-53K
50K
J. VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:5.505.151.374.75.392.2] [image:5.505.62.254.458.559.2]column (data not shown). These results indi-catethat thetranscriptive complex retains the ability to synthesize viral RNA-containing poly(A).
Size of in vitro poly(A). The mechanisms involved in controlling the size of viral poly(A) segmentshave yetto be determined. Itwas of
interest to determine what effect, ifany,
Tri-ton-high salt disruption haduponthese
control-ling mechanisms. Product RNA labeled with [32P]AMP was digested with RNase A + T1,
chromatographed free ofmononucleotides, and
18S 28S
6
0
x
E4
C.
Ca4
0 10 20 30
FRACTION NUMBER
FIG. 4. Size determination ofRNAsynthesized by transcriptive complex. RNA synthesized in vitro in thepresenceof [a-32P]ATP by the isolated
transcrip-tive complex wasphenol-SDS extracted,
chromato-graphed on a Sephadex G-50 column, and
sedi-mentedon a15to30%linearsucrosegradient. After
fractionation, each gradient fraction (0.4 ml) was
assayed for[32P]AMP-labeled RNA (0) using PCS (AmershamlSearle). Thepositionsofunlabeled18S
and28Smarkersareindicated byarrows.
sedimentedonsucrosegradients. Unlabeled
N-formylmethionyl tRNA from Escherichia coli
andlabeled chickenembryofibroblasts4SRNA
were sedimented in a parallel gradient. After
fractionation, each fractionwasassayedfor
ra-dioactivity (Fig. 5). Nearly identical poly(A)
segmentsweresynthesized by both virions and
the transcriptive complex. The poly(A) seg-mentswereveryheterogeneous, ranging in size
from 4to 11S,with amajorpeakof
radioactiv-ity sedimenting at 4.4S. No poly(A) segments smaller than 4S weredetected.
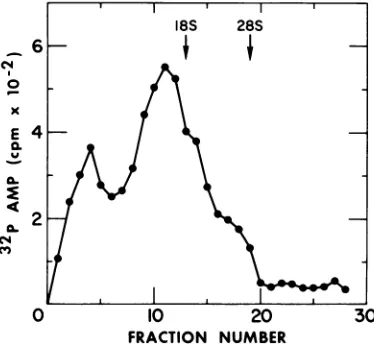
Nearest-neighbor analysis. As afinal proof
ofthe presence ofpoly(A), the nearest neigh-bors of the[32P]AMP residuesweredetermined.
The RNase-resistant RNA synthesized by the transcriptive complexwaspooled from the4 to
11S regions ofsucrose gradients (Fig. 5) and
hydrolyzed with KOH. Results (Table 3) show
that 95.5% of the RNase-resistant RNA
con-tainsAMPas anearestneighbor, proving that it is indeed poly(A). Approximately 2% ofthe
radioactivity migrated asGMP, perhaps
repre-sentingthe baseadjacenttopoly(A).
Assay for methyl transferase activity. We previously (6)demonstratedthatpurified NDV contains anenzyme that transfers the methyl group from S-adenosyl-L-methionine to viral RNAinvitro. To assayfor methyl transferase
activity,theisolatedtranscriptivecomplexwas
incubated in an in vitro transcriptase reaction
containing S-adenosyl-L-[methyl-3H]methio-nine and examined for 3H incorporation into viral RNA. Results (Table4) indicate that the transcriptive complex retained the ability to
methylate the RNA synthesized in vitro.
Addi-tionofS-adenosyl-L-homocysteine inhibited 3H incorporation by 89%, indicating that only the methyl group of
S-adenosyl-L-[methyl-3H]methionine was incorporated into viral
TABLE 2. RNasedigestionandpoly(U)-cellulosechromatography ofin vitroproductRNA
RNA sample % Resistance
%
Bound topoly
(U)-cel-RNaseA+T,a RNaseT2b lulos
Controls
'4C-labeled poly(A) 100 0.5 100
'4C-labeledchickenembryofibroblast 0 0.8 0.5
2&S rRNA Virion
[32P]AMP productRNA 8.0 0.1 17.3
Transcriptive complex
[32P]AMP productRNA 11.3 0.1 24.5
[3H]UMPproduct RNA 0.7 0.3
aRNAsampleswereincubated with50,ug of RNase A permland2,ug of RNaseT,per ml for30minat
37°C.
bRNAsamplesweredigestedwith 20 UofRNaseT2perml for 5 hat
37aC.
' More than97%of thelabeled RNAloaded onthecolumnwasrecovered.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:6.505.50.237.212.384.2] [image:6.505.44.448.501.629.2]1086 COLONNO AND STONE
1-1
3
x
1
0\j
b 2
(-A
3
x
0
10 20
FRACTION NUMBER
FIG. 5. Sizedeterminationofin vitropoly(A). RNA synthesized invitroin thepresenceof[a-32P]ATP by
virionsorby the isolated transcriptive complexwaspurified and treated withacombinationofRNase A and
T,. The RNase digest was phenol-SDS extracted, chromatographed on a Sephadex G-50 column, and
sedimentedon a15to30%linearsucrosegradient. Samples (100pi1) ofeachgradient fraction (0.4 ml)were
assayed for [:'2P]AMP-labeled RNase-resistant RNA synthesized byvirions(O)orthetranscriptivecomplex
(0).As4S markers, unlabeledN-formylmethionyl tRNA from E.coli(arrow)and 14C-labeled 4StRNA from
[image:7.505.151.387.73.340.2]chickenembryo fibroblasts (---)weresedimentedinaparallel gradient.
TABLE 3. Nearest-neighbor analysisof Poly(A) synthesizedinvitroby NDV transcriptivecomplex
Migration cpm recovereda %Totalcpm
re-covered
AMP 674 95.9
CMP 6 0.8
GMP 16 2.3
UMP 7 0.9
"In each experiment,morethan93% of the radio-activity loaded on the thin-layer plate was
re-covered.
RNA. Todetermine if the methyl transferase
wasabletomethylate other RNAtemplatesin
the absenceofviralRNAsynthesis,the ribonu-cleosidetriphosphateswerereplacedbyvarious RNA templates. Results (Table 4) show that
there was little or no 3H incorporation with RNA templates such as poly(A), poly(G), and yeast carrier RNA, but incorporation was
ob-served usingstripped tRNA fromE. coli W. Location of methyl groups in RNA
prod-ucts. The methyl-1H-labeled RNAs were next examined for the presence ofmethyl label ina
TABLE 4. Assayfor methyl transferase activityin
transcriptivecomplex
cpm[3H]SAM
Additionw incorporated/mg
protein
None 250
0.7mMATP,CTP, GTP, UTP 13,450
0.7 mMATP, CTP, GTP, UTP+ 1 1,550
mMSAM
0.1mgPoly(A) (MILES) 350
0.1mgPoly(G) (MILES) 950
0.04 mg Yeast carrier RNA 1,400
(MILES)
0.1mgStripped tRNA fromE.coli 1]8,650
W (GIBCO)
't Beforeadditions, reaction mixtures contained 50
mM Tris-hydrochloride (pH 7.8), 30 mM NaCl, 0.4
mM MnCl2, 3 mM dithiothreitol, and 0.78 ,uM
[3H]SAM (S-adenosyl-L-[methyl-3H]methionine).
5'-terminal "cap" structure. Methyl-labeled RNA was phenol extracted, hydrolyzed with
KOH, andchromatographedon a DEAE-cellu-lose column. Results demonstrated that all of themethyl-3Hlabel in RNAsynthesized by
on November 10, 2019 by guest
http://jvi.asm.org/
[image:7.505.272.465.441.567.2] [image:7.505.70.260.444.505.2]15
10
5
15
CY
20 40 60 duO
FRACTON &JM8ER
FIG. 6. DEAE-cellulosechromatography of kaline digest of RNA synthesized in vitro. RP beledinvitrointhepresenceofS-adenosyl-L-[m
3H]methionine by either virions orthetransci
complex was hydrolyzed with KOH and chrc
graphedon aDEAE-cellulose column (0.6by1
aspreviously described (6). The elutionpattern
oligonucleotide markers resulting from RNase tionof wheatgermrRNA isindicated by thea andthesodium chloride concentration(---) w
tained fromastandardcurve.Column fraction
assayed for radioactivity (0)aspreviously desi
(6). (A)Methyl-3H-labeled viral RNA synthesi,
virions; (B) methyl-3H-labeled viral RNA s*
sized by the transcriptive complex; (C) meth labeled RNA from E. coli methylated by the scriptivecomplex.
ons (Fig. 6A), and the transcriptive con
(Fig. 6B) elutedas asingle peak havinga
charge, and has been previously identifi
7MeGpppGp (Colonno and Stone, in p
DEAE chromatography of the methyl-3H-la-beledtRNA (Fig. 6C), methylated by the
tran-scriptive complex, demonstrated that the
methyl label was not incorporated into a
5'-terminal structure, but instead eluted as an
internal methylated base (-2 charge), which
02 lateridentifiedas 7MeG (datanotshown).
Hybridization of in vitro-labeled RNA to
50S genomes. To demonstrate that the RNA
QI productssynthesized byvirionsandby the
iso-latedtranscriptivecomplexwere faithful
tran-scripts of the genome, in vitro RNA products were hybridized to purified NDV 50S genome
RNA. These hybridization experiments (Table
5) show that 97% of the radioactive label was
x incorporated intovirus-specificRNAexceptfor
methyl-3H-labeledtRNA. In thecaseof
methyl-Q2}3H-labeled
tRNA,
more than90%
of the zHlabel was not incorporated into virus-specific
RNA, proving that the virion-associated
methyltransferasewasabletomethylate
exog-01 enousRNA.
DISCUSSION
Purifiedvirionsof NDV contain atleastfour
enzymatic activities relatedto RNA synthesis:
RNA transcriptase (18), methyltransferase (6),
capping (Colonno and Stone, in press), and
02 poly(A) polymerizing activities (33). To date,
the specific polypeptides involved in each of these enzymatic activities are not known. The ol membrane proteins HN and M, along with the
lipidenvelope of thevirion, canberemovedby
disrupting purified virions with Triton X-100 detergentand high salt(30).Afterdisruption,a
nucleocapsid structure remains (Fig. 2) which retainstheabilitytosynthesize complementary
RNA in vitro(Table5)and is therefore calleda anal- transcriptive complex. In addition to genome
VA la- RNA,theisolatedcomplexwascomposedofthe
wethyl- nucleocapsid protein N, a minor protein with
rmpato
anapproximate
molecularweight
of53,000,
t7cm)
Iofthe TABLE 5. Hybridization of invitrolabeled RNA to
diges- 50S Genome RNA
irrows
was
ob-swere
,cribed 'zedby ynthe- lyl-3H-
tran-nplex ,-4.5 ed as
ress).
%RNaseresistance RNAsample
Notannealed& Annealed' Virion
[32PIAMPproduct RNA 8.4 96.3 Methyl-3HproductRNA 1.5 100 TranscriptiveComplex
[32P]AMPproductRNA 10.8 100 13H]UMPproductRNA 1.3 97.0
Methyl-3H product RNA 6.5 97.5
Methyl-3H tRNA 0.1 9.5
a ProductRNAwastreateddirectlywith RNase A +T,. b Product RNAwasfirstannealedto33,ug of 50Sgenome
RNApermland thentreatedwithRNase A+T,.
A -2 -3 *4 -5 -7-8
o
*;*+
B -2 -3 -4 -5 -e 7 X
5_ ~~~~on
I
I/I
I I
c--3 -4 -5 6 - A
* *
*
* + i
6_Xj '~~~A
2<j; oA
f1,'A
A con November 10, 2019 by guest
http://jvi.asm.org/
[image:8.505.52.241.66.452.2] [image:8.505.256.448.533.640.2]1088 COLONNO AND STONE
andthe large 150,000-molecular-weight L pro-tein, with traces of HN and M proteins (Fig. 3).
Investigations by Bishop and Roy (4) with vesicular stomatitis virus (VSV), a
rhabdovi-rus, havealsoshown that neither the G nor M proteins are required for RNA transcription.
Emerson and Wagner (10, 11)demonstratedan L protein requirement for RNA synthesis by VSV, and Emerson and Yu (12)have recently shown a requirement for the NS protein.
Per-haps the L, N, and 53,000-molecular-weight
proteinsof NDVareanalogoustotheL,N,and NSproteinsof VSV.
The transcriptive complex isolated from
NDV synthesized an RNA in vitro that was
heterogenous andslightly smallerinsize (Fig. 4)thanthat synthesized by purifiedvirions (6).
This may be due to the isolation procedures,
sincethe transcriptivecomplex didnot appear as an intact nucleocapsid structure (Fig. 2). However, the isolated transcriptive complex didretaintheabilitytosynthesizepoly(A) seg-ments on viral RNA in vitro (Tables 2 and 3; Fig. 5). Attempts to synthesize poly(A) using
several knownprimers inthe absence ofRNA
synthesis have been unsuccessful. Thisresult wouldseem to indicate that the RNA transcip-taseand poly(A) polymerase activities are
cou-pled. Removal of the outer membrane of NDV
had no apparent effect on the size of poly(A)
segments attachedtomRNA (Fig. 5), suggest-ing that the secondary structure of
nucleocap-sids doesnotplayarole indeterminingthesize
ofpoly(A) segments.
Wehave recently identified the 5' terminus
of NDVviral RNA synthesized invitro in the presence ofamethyl donoras 7MeGpppGpPyp
... (Colonno and Stone,inpress).Similar
stud-ies using the isolated transcriptive complex demonstrated that the methyl transferase and
capping activities (Table 4, Fig. 5) are also retained. Both the virions and thetranscriptive complex were able to synthesize methylated viralRNA in vitro, whichcontains a
5'-termi-nal structure, 7MeGpppGp (data not shown).
Unlike the poly(A) polymerase, the methyl
transferase may be a separate enzyme activity
fromthe RNAtranscriptase. Thisis supported
by the findingthatthevirion-associatedmethyl
transferase can methylate exogenous RNA such as E. coli tRNA. Analysisofthe
methyl-labeled E. coli RNA showed that it was not virus specific (Table 5) and that the methyl
label did not reside in a capped 5'-terminal
structure. Instead, the methyl label appeared
to exist as a single base methylation (Fig. 6), which has been further identified as 7MeG
(datanotshown).
Similar experiments by Abraham and Baner-jee (1) have demonstrated that nucleocapsid structuresisolated from VSVby high salt and
Triton disruption alsoretainthe abilityto syn-thesize viral mRNA that is capped, methyl-ated, and contains poly(A) segments identical
tothosesynthesizedbywhole virions. Analysis
of the active nucleocapsid structure by poly-acrylamide gels demonstrated the presence of L, N, and NS with a traceofM protein. Thus, VSV, arhabdovirus, andNDV, a paramyxovi-rus, mayutilizesimilarenzymaticmechanisms totranscribe andmodifyviral mRNA.
ACKNOWLEDGMENTS
Wewish to express our gratitude to David Wright and Jeanette Robbins who provided skilled technical assistance. Theexcellenttechnical assistance of Lorraine Hammer and the use ofMcCollumLaboratory's electron microscope facil-ity is gratefully acknowledged. We wish to thank Janis McMillen and Richard Consigli for the gift of ['4C]formaldehyde-labeledNDV virions.
This research wassupportedby Public Health Service grantAI-11127 from the National Institute ofAllergy and Infectious Diseases.
LITERATURE CITED
1. Abraham, G., and A. K.Banerijee.1976. The nature of the RNAproducts synthesized in vitro by subviral components of vesicular stomatitis virus. Virology 71:230-241.
2. Aminoff, D. 1961. Methods for the quantitative estima-ti9n ofN-acetylneuraminic acid and their application tohydrolysates of sialomucoids. Biochem. J. 81:384-392.
3. Bikel, I., and P. H.Duesberg.1969. Proteinsof New-castledisease virus and of the viral nucleocapsid. J. Virol. 4:388-393.
4. Bishop,D. H.L., and P.Roy.1972.Dissociation of VSV and relation of the virionproteins to the viral tran-scriptase. J. Virol.10:234-243.
5. Choppin, P. W. 1964. Multiplication of a myxovirus (SV5) with minimal cytopathiceffects and without interference.Virology23:224-233.
6. Colonno, R.J.,andH.0.Stone.1975. Methylationof messengerRNA ofNewcastle disease virus in vitroby a virion-associated enzyme. Proc. Natl. Acad. Sci. U.S.A. 72:2611-2615.
7. Colonno, R. J., and H. 0. Stone. 1976. Synthesis of complementaryRNAcontainingpolyadenylicacidby Sendai virions in vitro. J. Virol.17:737-744. 8. Duesberg,P.H.,and W. S. Robinson. 1965. Isolation of
the nucleic acid ofNewcastle diseasevirus (NDV). Proc.Natl.Acad. Sci. U.S.A. 54:794-800.
9. East, J. L. 1968. Nucleotide composition of ribonucleic acidbyspectralanalysisandthin-layer chromatogra-phy. Anal. Biochem. 24:409-418.
10. Emerson, S. U., and R. R. Wagner. 1972. Dissociation andreconstitutionofthetranscriptase andtemplate activitiesofvesicular stomatitis B and T virions. J. Virol. 10:297-309.
11. Emerson, S. U., and R. R. Wagner. 1973. L-protein requirement for in vitro RNA synthesisby VSV. J. Virol. 12:1325-1335.
12. Emerson, S. U., and Yu-Hwa Yu. 1975. Both NS and L proteinsarerequiredforinvitroRNAsynthesis by VSV.J. Virol.15:1348-1356.
J. VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
13. Evans, M.J., and D. W.Kingsbury. 1969. Separation of Newcastle disease virus proteins by polyacrylamide gel electrophoresis. Virology 37:597-604.
14. Granoff,A. 1959.Studiesonmixed infectionwith New-castle disease virus. I. Isolation of NDV mutants and tests for geneticrecombination between them. Virol-ogy9:636-648.
15. Haslam, E. A., I. M.Cheyne, and D.0. White. 1969. Thestructural proteins of Newcastle disease virus. Virology 39:118-129.
16. Home, R. W., A. P.Waterson, P. Wildy, and A. E. Farnham. 1960. The structure and composition of the myxoviruses. Virology 11:79-98.
17. Home,R. W. 1965. Negative stainingmethods,p. 328. In D. H. Kay (ed.), Techniques for electron micros-copy,2nded. F. A. David, Philadelphia.
18. Huang, A.S., D. Baltimore, and M. A.Bratt.1971.An RNApolymeraseinvirions ofNewcastledisease vi-rus: comparison with thevesicular stomatitis virus polymerase.J.Virol. 7:349-389.
19. Kingsbury, D. W. 1966. Newcastle disease virus RNA. I.Isolation and preliminary characterization of RNA from virusparticles. J. Mol. Biol. 18:195-203. 20. Kingsbury, D. W. 1966. Newcastle disease virus RNA.
II. Preferential synthesis of RNA complementary to parental viral RNA by chick embryo cells. J. Mol. Biol. 18:204-214.
21. Kingsbury, D. W., and R. W. Darlington. 1968. Isola-tionandproperties of Newcastle disease virus nucleo-capsid.J.Virol. 2:248-255.
22. Lowry,0.H.,N. J. Rosebrough, A. L.Farr,and R. J. Randall. 1951. Protein measurementwith the Folin phenol reagent. J. Biol. Chem. 193:265-275. 23. Marshall, S., and D. Gillespie. 1972. Poly(U) tracts
absent fromviral RNA. Nature(London) New Biol. 240:43-45.
24. Marx, P. A., A. Portner, and D.Kingsbury. 1973. Sen-dai virion transcriptase complex:polypeptide compo-sitionand inhibition byvirionenvelopeproteins. J. Virol. 13:107-112.
25. McMillen, J., and R. A.Consigli.1974. In vitro radiois-otopic labelingofproteins associated with purified polyomavirions. J.Virol. 14:1627-1629.
26. Meager, A., and D. Burke. 1973.Studies on the struc-tural basis of the RNApolymerase activity of New-castlediseasevirusparticles. J. Gen. Virol. 18:305-317.
27. Mountcastle, W. E., R. W. Compans, L. A. Caliguiri, andP. W.Choppin.1970. Nucleocapsid protein sub-units of simian virus 5, Newcastle disease virus, andSendaivirus.J.Virol. 6:677-684.
28. Mountcastle, W. E., R. W. Compans, and P. W. Chop-pin. 1971. Proteinsand glycoproteins of paramyxovi-ruses: a comparison of simian virus 5, Newcastle disease virus, and Sendai virus. J. Virol. 7:47-52. 29. Robinson, W. S.1971. Ribonucleic acid polymerase
ac-tivity inSendai virions and nucleocapsid. J. Virol. 8:81-86.
30. Scheid, A. and P. W. Choppin. 1973. Isolation and purification of the envelope proteins ofNewcastle disease virus. J. Virol. 11:263-271.
31. Scheid, A. and P. W. Choppin. 1974. Identification of biological activities of paramyxovirusglycoproteins. Activation ofcell fusion, hemolysis, and infectivity by proteolyticcleavage of an inactive precursor pro-teinofSendai virus. Virology 57:475-490.
32. Sheldon, R., C.Jurale,and J. Kates. 1972.Detection of polyadenylic acid sequences in viral and eukaryotic RNA. Proc. Natl. Acad.Sci. U.S.A.69:417-421. 33. Weiss, S. R., and M. A. Bratt. 1974. Polyadenylate
sequences onNewcastle disease virus mRNA synthe-sizedinvivoand in vitro. J. Virol. 13:1220-1230.
on November 10, 2019 by guest
http://jvi.asm.org/

![FIG. 5.sedimentedchickenT,.virionsassayed(0). Size determination of in vitro poly(A). RNA synthesized in vitro in the presence of[a-32P]ATP by or by the isolated transcriptive complex was purified and treated with a combination ofRNase A and The RNase dige](https://thumb-us.123doks.com/thumbv2/123dok_us/1554841.108078/7.505.70.260.444.505/sedimentedchickent-virionsassayed-determination-synthesized-presence-isolated-transcriptive-combination.webp)
