0022-538X/80/08-0414/06$02.00/0
Picornaviral
VPg Sequences
Are Contained in the
Replicase
Precursor
MARK A.PALLANSCH,'* OLEN M.
KEW,'t
ANN C. PALMENBERG,' FREDGOLINI,2t ECKARD WIMMER,2ANDROLAND R. RUECKERT'BiophysicsLaboratory ofthe Graduate School andDepartment ofBiochemistry oftheCollege of AgriculturalandLifeSciences, University of Wisconsin, Madison, Wisconsin53706,1andDepartment of
Microbiology,StateUniversity ofNew York atStony Brook, Stony Brook,New York117942 Ithas
previously
been shown that the RNAreplicaseofencephalomyocarditis
virus contains two virus-coded proteins, D and E, which are produced in twosuccessiveproteolytic steps: (i)C-- D+?;and(ii)D -*p22+E. It is here shown
(i)
that virus protein H (molecular weight, 12,000) is the previouslyunidentified
product of the firststepand(ii) thatVPg,aprotein linkedcovalentlyto thevirionRNA,
yields twotryptic peptides foundinprotein C but not in proteinD.The resultssuggestthatVPg
isderivedby
cleavage
ofproteinC and thatproteinHmaybeanintermediate.
Preliminary
experimentswith VPgsequencesin polio-viralnoncapsid protein lb, the counterpartofencephalomyocarditisviralprotein C,wereinconclusive.Inall
picornaviruses
examined there isasmall protein (VPg) covalentlyattached to the 5'-ter-minal residueof thegenomeRNA(8,
10, 13,
21). Circumstantial evidence suggests thatVPg
isencoded bythe viral genome (8, 13);
however,
its function ispresently
unknown.BecauseVPg has been found linked to nascent RNA strands of thereplicative
intermediate(7,
16),
it hasbeen
proposed
thatit isinvolved in RNArepli-cation,
possibly
attheinitiation
step(7,
13, 16,18).
Anunexpected result,
however,
was theobservation that
freeVPg
cannotbe detected
in extracts ofpoliovirus-infected
cells(Golini
andWimmer, unpublished
data).
This led
to thesuggestion
thatVPg
isgenerated
by proteolytic
cleavage
from alarger
precursorprotein during
initiation
ofRNA synthesis
(16,25), possibly
fromnoncapsid
viralprotein
2(NCVP
2; refer-ence6).
(Nomenclature for polioviral protein
is that ofSummers etal.[24]
asmodified by
But-terworth[1],
and that forencephalomyocarditis
[EMC] viral proteins
isthat of
Butterworth
et al.[2].)Alternatively,
it has beensuggested that VPg may represent theproduct
of a lessfre-quently
translated second cistron(11). There is,however,
nodirectevidencesupporting
either of theseproposals.We now report
evidence,
based on trypticmapping,
thatVPg isindeed
avirus-coded
pro-tein and that it is derived fromprotein
C of EMCvirus,possibly
via anintermediate protein,
H.tPresent address:DivisionofVirology, CenterforDisease Control,Atlanta, GA 30333.
fPresentaddress: Department of Cellular Biology, Scripps Clinic and Research Foundation,La Jolla, CA 92037.
MATERIALS AND METHODS Virus and cells. The EMC virus and thepoliovirus
type 1(Mahoney) used have been describedpreviously
(9, 19). Virus was propagated in H-HeLa cells which were grown in medium B (15) containing 10% calf serum (K. C. Biologicals, Inc., Lenexa, Kans.) except whereotherwise noted.
Preparation of radiolabeled EMC VPg. Infec-tion ofcells insuspension culture followed the proce-dure ofMcGregoretal.(14), with the exceptionofa 30-minvirus attachmentperiod. Virionswerelabeled by addition ofeitherL-[4,5-3H]lysine (5.0 mCi to 4 x 107 cells) or L-[4,5-3H]leucine (10.0 mCito 4 x 105 cells) at2.5 hpostinfection. Incubation (37°C) in the presence of label continued until 6 h postinfection,at which time theinfected cell culture was subjected to threefreeze-thaw cycles. The released virus was puri-fied by pelleting,followed by banding in an isopycnic cesiumchloride gradient as previously described (15). The virus was removedfrom cesium chloride by pel-leting through a sucrose cushion. The RNA was ex-tracted from virions and precipitated as described previously (22). The VPg-containing RNA was di-gested for 60 minat37°C with a mixture of pancreatic RNase A (10,ug/ml),RNaseT,(10
jg/ml),
andRNase T2 (5U/ml) in 0.05Msodium acetate (pH 4.5). VPg was dissolved in 0.01% sodium dodecyl sulfate by heating for5min at100°C.Preparation ofradiolabeled pollovirus VPg. Poliovirus VPg labeled withL-[4,5-3H]lysine was pre-paredasdescribed previously (19).
Synthesis of protein in cell-free extracts. In vitro EMCRNA-dependent protein synthesis was car-ried outexactly aspreviously described (22, 23). Trans-lation
(90-pl
reaction mixture) was in the presenceofL-[4,5-3H]leucine (24,LCi) for 60 min, at which time 5
,ul
of pancreatic RNase A (1 mg/ml) and 10,Ld of cycloheximide (1mg/ml) wereadded. After an addi-tional 60min ofincubation, the sample was acetone 414on November 10, 2019 by guest
http://jvi.asm.org/
VPg IN REPLICASE PRECURSOR
415
precipitated as previously described (17), dissolved in
electrophoresissample buffer, and dialyzed as previ-ously described (2).
Synthesis of protein in infected cells. Lysates wereprepared from 2x 107cellsinfected at a multi-plicityof 100 PFU/cell. To obtain proteins D or NCVP 2,cells were exposed at 240 min postinfection to either
L-[4,5-3H]lysine or L-[4,5-3H]leucine for 15 min. To obtainproteins C or NCVP lb, cells were infected in the presence of0.05%fetalcalf serum and treated with 0.91 mM (EMC virus) or 0.70 mM(poliovirus) ZnCl2
at204minpostinfection as described (3). Either
L-[U-'4C]lysine
orL-[U-'4C]leucine
wasadded from 210min to 310 min postinfection. In both cases, cells were lysed at the end of the labeling period without washing, using hot sodium dodecyl sulfate, and the samples weredialyzed (2).Preparative electrophoresis. Preparation of samples for electrophoresis has been described (15). Preparativeelectrophoresis for infected cell lysates to isolate proteinsC, D, NCVP lb, or NCVP2was the sameaspreviously described (22) except that the gel composition was 4.9% acrylamide, 0.2%
N,N'-methyl-enebisacrylamide, 0.1% sodium dodecyl sulfate, 0.1 M sodium phosphate at pH 7.2, 0.1%
N,N,N',N'-tetra-methylenediamine, 0.5M urea, andammonium per-sulfateat afinalconcentration of 0.05%. Preparative electrophoresis of in vitro reaction mixtures to isolate protein H wasperformed on 10% acrylamide gels con-taining8Murea(12).
Tryptic mapping. Proteinsweremixed,reduced,
alkylated,acidprecipitated,digested with trypsin, and analyzed by cation-exchange chromatographyexactly as
previously
described(12).RESULTS
Evidence that
VPg
sequencesof EMC
vi-rusreside in
protein
C. Evidence that
VPg
sequences arecontained
inprotein C
wasob-tained
by
digesting
amixture of the
proteins
which had been
differentially
labeled
by
incor-poration of radiolabeled leucine.
Toobtain
la-beled
VPg,
purified
RNA,
extracted fromvirions
grownin
the
presenceof
[3H]leucine,
wasdi-gested
for
1h
with
amixture of
RNasesusing
conditions which had
previously
been
showntorelease the
covalently
attached
protein
of
polio-virus
in the form
VPg-pUp (7, 13).
ProteinC
wasisolated
by
preparative electrophoresis
of
lysates from cells which had been
exposed
to['4C]leucine
from
3.5to 4.5hpostinfection. Zinc
chloride
(0.91
mM)
wasincluded
during
the
incorporation period
to enhance theyield
of EMC viralprotein C (3),
which is otherwise present inonly
small amounts. The mixtureof
VPg
andprotein C
wasalkylated,
digested
withtrypsin,
andchromatographed
on acation-ex-change
column
toresolve
thepeptides (Fig. 1A).
VPg
yielded
asingle
leucine-containing
peak
which
emerged
atfraction 11(Fig. 1A,
arrow)
just behind
theflow-through
peak (fraction
5).
The
digest
fromprotein
C
alsoexhibited
apeak
in this position which
was,
however, somewhat
broader
onthe
right
shoulder.
That this broaderpeak is
acompound
one isshown
bycomparing
thetryptic profile
ofprotein C with that of itscleavage product
(2), protein D (Fig.1B).
Thisprofile shows that
thetryptic
peak from VPg is missing fromprotein
D. Thereremains,
how-ever, asmaller peak
in Dwhich accounts for thenoncoincidence
of the peaks fromprotein
C andVPg
(seenin
Fig. 1A, arrow). Thedifference
between the profiles of
proteins C and D was,however,
notcompletely accounted
for by thetryptic peptide of VPg.
Thus one sees in theprofile
of protein C noncoincidences
atfractions 27,62, 93,and
98.As
seenin
Fig. 1C, the
tryptic profile of protein Haccounts
forall
of thesedifferences
betweenC and
D,including
the tryptic peakemerging
with that in
the VPg profile (arrow). These dataindicate that
H is thepreviously unidentified
cleavage
complement
of
D(i.e.,
C -. H
+D).
The data also
strongly
suggest that VPg is partof the H sequences
contained
inprotein
C but notin
protein D.
The
latter conclusion
isgreatlystrengthened
by
asecond experiment modified by substituting
radiolabeled
lysine for radiolabeled
leucine (Fig.2A).
The
lysine-labeled
peak coincides with
onefound in
protein C but
notin
protein
D
(Fig.
2B), and this
peak
is
different from the
leucine-labeled
peak
inFig.
1. ThusVPg exhibits
twotryptic
peaks,
onecontaining
lysine
but
noleu-cine
and
asecond that contains leucine but
nolysine, which coincide with
tryptic
peaks
from
protein
C but
notfrom
protein
D.
VPg in
polioviral
proteins.
The
polioviral
homolog of
protein
C is NCVP
lb,
which is
cleaved
toNCVP
2plus
astill
unidentified
pro-tein.Based
uponanamino acid
analysis
of
VPg,
the
RNA
sequencecoding
for
VPg
has been
determined (N.
Kitamura, C.
Adler,
J.
Martinko,
S. G.
Nathenson,
and E.
Wimmer,
submitted for
publication).
This RNA
sequence islocated
ap-proximately
2,000
nucleotides from the 3' end of
the
poliovirus
genome,that
is,
within
asegment
of RNA
coding
forNCVP lb.
Moreover,
the
amino
acid sequence of
VPg,
asdeduced
from theRNA sequence,
predicts
that
digestion
of
VPg with
trypsin
should
yield
three
peptides.
Thesethree
peptides
haveindeed
beenobserved
(Fig.
3A).To
determine
whether theVPg
sequences of
poliovirus could
bedemonstrated
inNCVPlb,
the
proteins,
differentially
labeled with
lysine,
were
compared
asbefore.
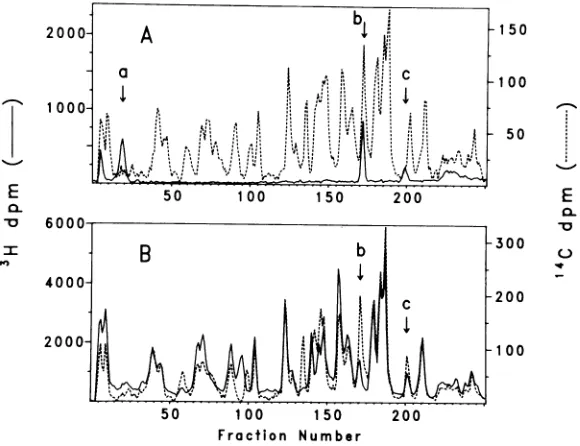
Inthis
casepolioviral
VPg
exhibited
threetryptic
peaks,
a,b,
and
c(Fig. 3A).
Nonecoincided
exactly
withthemajor
peaks
inNCVP
lb;
moreover,peak
afromVPg
appeared
tobeentirely
absentfromNCVP
lb.VOL. 35, 1980
on November 10, 2019 by guest
http://jvi.asm.org/
416 PALLANSCH ET AL.
4000-
A
2000- N
E
la
CL
50 1 00 150 200
Fraction Number
FIG. 1. Tryptic profiles ofleucine-labeled EMCproteinsC, D,H, andVPg.Differentially labeled proteins were mixed, digestedwithtrypsin, chromatographedon acation-exchange column, andassayed for radio-activityasdescribed in thetext.(A)3H-labeledVPg( )versus
"4C-labeled
proteinC(---). (B)3H-labeled proteinD( )versus"C-labeledproteinC(---). (C)3H-labeledproteinH( )versus"C-labeledprotein C(---).ProteinHwasisolatedfromacell-freetranslationproduct (seethetext).Scaleswereadjustedsuch that theratiooftheirheightsequaledthespecificactivityoftheproteins (disintegrationsperminute[dpm]perdalton).
These results do not exclude the possibility,
however, thatVPg peaksb andc aremembers
ofcompound tryptic peaksfromNCVP lb,and that VPg peak a represents the new tryptic peptideproducedwhen NCVP lb iscleavedby
the viralprocessingenzyme.Alternatively, peak
acould beatryptic peptidelinkedto-pUp.This
nucleotidyl peptidewould have no counterpart inNCVP lb.
The possibility of compoundpeaks in NCVP lbwasfurtherexaminedbycomparingthe tryp-ticprofilesofNCVP lband its cleavage product
NCVP2 (Fig.3B). The resultsobtained in this
experimentwereinconclusive. Thiswas due in part to inability to resolve the VPg peptides
definitively withinthe largenumber of peptides found in NCVP lb and 2. A second difficulty was the presence of an unrelated protein, de-tectedbyanalytical electrophoresisinslab gels, in ourpurified preparations of NCVP lb. This
contaminantwasalsomanifestedbytryptic
pro-files which became significantly simpler when
NCVPlbwasenriched by treating infectedcells
with zinc (results not shown). When VPg and NCVP 2 were compared directly, two of the three VPgpeptides were clearly resolved from
anyinNCVP 2,suggesting the absence ofVPg
sequences in this protein (not shown). Other
experiments with lysine-labeled VPg excluded thepresenceofVPgsequencesinpolioviral
cap-sid precursor NCVP la, in NCVP5b, which is relatedtostableproteinX(20),and in thesmall unmapped proteins 9a, 9b,and 10.These
exper-imentswererepeated usingleucineasthe
radi-olabel, but the results again failed to establish conclusive evidence forVPgsequencesin lb.
DISCUSSION
Thedatapresented hereindicate (i) that pro-tein H is the previously unidentified product
50 100 150 200
-6 0 0
-400
-20 0
E
C)
)00
-600
-300
I
on November 10, 2019 by guest
http://jvi.asm.org/
[image:3.504.119.407.62.392.2]VPg
IN REPLICASE PRECURSOR 417
2000-1
000-E 0Q
2:
A
4
:4
.4.': ::
1: :*
-800
-400
E
0-p400
t1*
-20 0
50 100 1 50 200
Fraction Number
FIG. 2. Trypticprofiles oflysine-labeled EMC viral proteins C, D, and VPg. (A) 'H-labeled VPg ( )
versus
"C-labeled
protein C (---). (B)3H-labeledprotein D ( )versus"C-labeledprotein C(---)-
2000-1
000-E
0--o
-1 50
-100
50
E
-o
[image:4.504.103.398.68.291.2](-Fraction Number
FIG. 3. Tryptic profiles oflysine-labeled poliovirusproteinsNCVPlb, NCVP2, andVPg. (A)3H-labeled VPg( ) versus '4C-labeled NCVP lb (---). (B) 3H-labeled NCVP2 ( ) versus"C-labeledNCVP lb
(-- ).
obtained when EMC viralprotein Cis cleaved to formprotein D, and (ii) thatVPgis partof
protein H. These experiments are consistent
with models ofpicornavirus replication proposed previously (7, 13, 16, 18, 25) which suggest in-volvement ofVPgininitiation of RNAsynthesis.
They arealsoconsistentwith theproposal (17)
that thepolymeraseprecursorC donatesVPgto the nascent RNA strand during initiation of
synthesis.ThefindingofVPgsequencesin
pro-tein H suggests that generation of VPg from
protein C involves more than one proteolytic
50 1 00 1 50 200
A
b4
a C
4 *:A:
4
A ::
.i1
hAI
50 100 1 50 200
u
on November 10, 2019 by guest
http://jvi.asm.org/
[image:4.504.102.392.329.552.2]CAPSID
.I
NON -CAPSID
C(83)
A(loo) [ H(12) D (75) ]
B(90)
VPl8)([p22(20)
E(56) ]DI (65) a(34)
C(40) Y(23)
G016) 1(11)
i(9)()
FIG. 4. ProcessingmapofEMC viralproteins re-visedto includeproteinsH andVPg. Brackets en-closeproteins whose order is still unclear. Box en-closesunmapped proteins.Electrophoretically deter-mined molecular masses(parentheses) aregiven in kilodaltons (17). Theelectrophoretically determined molecularmassof VPgis8,000to10,000daltons(8); however, sequencing studiesonpolioviral- VPg sug-gestamasslessthan3,000.
step.It remainstobeclarified,however,whether
H is an intermediate in the pathway of VPg additiontoRNA.
Thenewdata have beenincorporated into the proteolytic processing mapof
EMC
virus (Fig. 4). The position of protein H on the amino-terminal side ofprotein D is basedon pactamy-cin mapping (4); however, noconclusioncanbe drawn from the tryptic dataon the position of VPg within protein H.IfEMC andpoliovirus follow thesameprotein cleavage pattern, then the presence ofVPg in protein H could explain why polio VPg is not
found free in the infectedcell(25). Because it is known that the number of translations of the viral genome significantly exceeds the number ofnewviral RNA strands made inaninfection (reviewed in N. Kitamura, C. Adler, and E. Wimmer, Ann. N. Y. Acad. Sci., in press, and
reference5),there should beasignificantexcess ofVPgsequences.If thesequencescontainedin
proteins C and H areonly released when RNA synthesisoccurs,freeVPgmaynotbe detectable ininfected cells. In otherwords, protein Hmay
representtheexcesssynthesis of
VPg,
although its role in RNAreplication ispresently unclear. Themajor cleavagestepsin the processing of replicase-related proteins of poliovirus and EMC virusareanalogous (lb-- 2+? is analogoustoC
-- D + H; reference 20). Accordingly, weanticipate that VPg of poliovirus is containedin
NCVP lb andnotin NCVP2.Theresults
pre-sentedherearecompatible with this model,but inconclusive.
ACKNOWLEDGMENTS
WethankDaniel Omilianowski forexcellenttechnical
as-sistance in running the tryptic peptide column and Jane Hubertz for assistance inthepreparationofVPg.
This workwassupported bygrantMV-33Ifrom the
Amer-icanCancerSociety and by Public Health Service grants CA-08662,CA-16879, and AI-15122 from the National Institutes of Health. M.A.P. is a National Science Foundation Predoc-toralFellow.
LITERATURE CiTED
1. Butterworth, B. E. 1973. A comparison of the virus-specific polypeptides ofencephalomyocarditis virus, hu-manrhinovirus-lA, andpoliovirus. Virology 56:439-453.
2. Butterworth,B.E., L.Hall,C. M.Stoltzfus,and R. R. Rueckert.1971.Virus-specific proteinssynthesizedin encephalomyocarditis virus-infected HeLa cells.Proc. Natl. Acad. Sci. U.S.A. 68:3083-3087.
3. Butterworth, B. E., and B. D. Korant.1974. Charac-terization of the large picornaviral polypeptides pro-ducedin thepresence of zinc ion. J.Virol. 14:282-291. 4. Butterworth, B.E., and R. R. Rueckert. 1972.Gene
order ofencephalomyocarditis virusasdeterminedby studies withpactamycin.J.Virol. 9:823-828.
5. Cooper,P.D.,A. Steinor-Pryor,and P. J.Wright. 1973.Aproposed regulator for poliovirus: the equestron. Intervirology1:1-10.
6. Dasgupta,A.,M.H.Baron,and D. Baltimore.1979. Poliovirusreplicase: asoluble enzyme able toinitiate copying of poliovirus RNA. Proc. Natl. Acad. Sci. U.S.A. 76:2679-2683.
7. Flanegan, J. B., R. F. Pettersson, V.Ambros,M.J. Hewlett, and D. Baltimore.1977.Covalent linkageof aproteinto adefined nucleotide sequenceatthe 5'-terminus of virionandreplicative intermediate RNAs ofpoliovirus. Proc. Natl. Acad. Sci. U.S.A.74:961-965. 8. Golini, F.,A. Nomoto, and E. Wimmer. 1978. The genome-linked protein of picomaviruses. IV. Difference in theVPg's ofencephalomyocarditis virus and polio-virus as evidence that the genome-linkedproteinsare virus-coded.Virology 89:112-118.
9. Hall, L., and R. R. Rueckert. 1971. Infection ofmouse fibroblastsbycardioviruses: prematureuncoating and itsprevention by elevated pH and magnesium chloride. Virology 43:152-165.
10. Hruby,D.E.,and W. K. Roberts.1978. Encephalomy-ocarditis virus RNA. III. Presence ofa genome-associ-atedprotein. J. Virol.25:413-415.
11. Humphries, S.,F.Knauert, and E. Ehrenfeld. 1979. Capsidprotein precursor is one of two initiated products oftranslation ofpoliovirus RNA in vitro. J. Virol. 30: 481-488.
12.Kew,0.M., M. A.Pallansch, D. R.Omilianowski, and R. R. Rueckert.1980.Changes in three of the four coatproteinsof oralpoliovaccine strainderived from type1poliovirus.J.Virol.33:256-263.
13. Lee,Y.F.,A.Nomoto,B. M.Detjen,and E. Wimmer. 1977.Aprotein covalently linkedtopoliovirus genome RNA. Proc. Natl. Acad. Sci. U.S.A. 74:59-63. 14.McGregor, S., L. Hall, and R. R. Rueckert. 1975.
Evidence for theexistence ofprotomersinthe assembly ofencephalomyocarditisvirus. J. Virol. 15:1107-1120. 15.Medappa,K.C.,C.McLean,and R. R. Rueckert.1971.
Onthe structureof rhinovirusIA.Virology 44:259-270. 16.Nomoto, A.,B.Detjen,R.Pozzatti, and E. Wimmer. 1977.The location of thepolio genome protein in viral RNAs and itsimplication for RNA synthesis. Nature (London) 268:208-213.
17. Palmenberg,A.C.,M. A.Pallansch, and R.R. Rueck-ert. 1979.Protease required forprocessing of picorna-viral coatprotein residesinthe viralreplicase gene. J. Virol.32:770-778.
18. Pettersson, R. F., J. B. Flanegan, J.K. Rose, and D. Baltimore. 1977.5'-Terminal nucleotide sequences of poliovirus polyribosomal RNA and virion RNA are identical.Nature(London)268:270-272.
19.Rothberg,P.G.,T. J. R.Harris, A. Nomoto, and E.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:5.504.68.259.47.166.2]Wimmer. 1978. Genome-linked protein of
picornavi-ruses.5.04-(5'-uridylyl) tyrosine isbond between
ge-nome-linkedprotein and RNA of poliovirus. Proc. Natl. Acad.Sci. U.S.A.75:4868-4872.
20. Rueckert, R. R., T. J. Matthews, 0. M. Kew, M.
Pallansch, C. McLean, andD.Omilianowski.1979. Synthesis and processing of picornaviralpolyprotein,p.
113-125.InR.Perez-Bercoff (ed.), The molecular
biol-ogyofpicornaviruses. Plenum Press, New York. 21. Sangar, D. V.,D. J.Rowlands,T.J.R.Harris, andF.
Brown. 1977. Protein covalently linked to foot-and-mouth diseasevirus RNA. Nature (London) 268:648-650.
22. Shih,D.S.,C. T.Shih,0.Kew,M.Pallansch,R.R.
Rueckert,and P.Kaesberg.1978.Cell-freesynthesis
andprocessingoftheproteinsofpoliovirus.Proc.Natl. Acad.Sci. U.S.A. 75:5807-5811.
23. Shih, D.S., C. T. Shih, D. Zimmern, R. R. Rueckert, and P.Kaesberg. 1979. Translation of encephalomy-ocarditis virus RNA inreticulocyte lysates: kinetic anal-ysis of the formation of virion proteins andaprotein
required for processing. J.Virol.30:472-480. 24. Summers, D. F., J. V. Maizel, Jr., and J. E. Darnell,
Jr. 1965.Evidence forvirus-specific noncapsid proteins inpoliovirus-infected HeLacells.Proc.Natl. Acad. Sci.
U.S.A. 54:505-513.
25. Wimmer. E. 1979.Thegenome-linkedproteinof picor-naviruses:discovery, properties and possible functions,
p. 175-190. InR. Perez-Bercoff (ed.), The molecular biology of picornaviruses. Plenum Press,NewYork.