0022-538X/88/114270-10$02.00/0
Copyright C) 1988, American Society for Microbiology
Synthesis and Processing of the Marek's Disease Herpesvirus
B
Antigen Glycoprotein
Complext
I. SITHOLE,' L. F. LEE,2 AND L. F. VELICER1*
DepartmentofMicrobiology and Public Health, Michigan State University, East Lansing, Michigan48824-1101,' and
Regional Poultry Research Laboratory, Agricultural Research Service, U.S. Department of Agriculture, EastLansing, Michigan 48823-53382
Received 5 April1988/Accepted 1 August 1988
TheMarek's diseaseherpesvirus Bantigen(MDHV-B) complexwaspreviously immunologically identified andmolecularly characterizedas asetof three glycoproteinsdesignated gplOO,gp6O, andgp49onthebasis of
apparent molecular weight and immunoprecipitation with both polyclonal and monoclonal antibodies. Immunoprecipitation analysis, previously withpolyclonaland more recently with monoclonal antibodies, of infected cell lysates labeled with [35S]methionine in the presence of tunicamycin, an inhibitor ofN-linked glycosylation, revealedtwo putativeprecursormoleculesof88,000 daltons (pr88) and44,000 daltons (pr44). High-resolution pulse-chase studies revealedthatgplOOwas aglycosylated intermediate whichwasprocessed
toyield gp6O and gp49. This cleavage was inhibited by monensin, an inhibitor of glycoprotein processing.
Endo-o-N-acetylglucosaminidases
F and H(endo-F, endo-H) reduced gplOOtopr88,indicating that the latter isanintermediatein thebiosynthetic pathway. Thesesameenzymesreducedgp49,andtoalesserextentgp6O, topr44, suggestingthatpr44istheirpolypeptidebackbone.Significantsupportfor thisconceptis thefactthat the same monoclonal antibody recognized all three molecules, gp60, gp49, and pr44. In the presence of monensin, terminal addition ofcomplexsugars wasalso prevented, since gp6Owasreplacedbyaslightly faster migratingcomponent which wasinsensitivetoboth endo-F andendo-H. Cell-free translation of infected-cell mRNA, followed byimmunoprecipitation analysiswitheitherpolyclonalormonoclonalantibody, resultedin detection ofaputative unglycosylatedprecursorpolypeptideof44,000daltons. Sincepr88wasnotthe initialprecursorpolypeptide ofthe MDHV-Bcomplex, itsexistencemay have resultedfrom dimerization of pr44.
Again, detection of both pr88 and pr44 with the same monoclonal antibody is consistent with this interpretation. These collective data obtained from the cell-free and in vivo studies with polyclonal and monoclonal antibodies reactive with MDHV-B are consistent with the concept that pr44, the initial gene product, dimerizestoformpr88anddemonstrate thatpr88isactuallyaprocessing intermediate glycosylated togplOO, anotherprocessing intermediate, which isthenprocessedtogp6Oandgp49.
Marek's disease (MD) is a
lymphoproliferative
diseaseof chickensthatwasfirstdescribedbyMarek in 1907 (29,37).The disease is caused by the Marek's disease herpesvirus
(MDHV) and represents the first neoplastic disease for which an effective vaccine has been developed (33, 35, 37). The mechanism of this immunity is not fully understood; however, it appears to be a two-step processinvolvingboth antiviral and anti-tumor-cell immunity (32, 33). Experimen-tal evidence suggests that antiviral immunity is superiorto
anti-tumor-cell immunity and that if the former occurs the
latter is not necessary (32).
Thevaccinevirus, herpesvirusofturkeys(HVT) (35), and MDHVareextensivelyrelatedimmunologically (35, 45,46),
sharing a large number ofpolypeptides that are apparently
common antigens (15, 41, 45, 46), including the two most prominent common antigens, Aand B (33, 34, 37).
In anefforttodetermine the mechanism of this protective
immunity at a molecular level, thesetwo common antigens have been the focus of recent immunologic and molecular
biologicstudies in thislaboratory (11,16-18).TheMDHV A
antigen (MDHV-A), a secretory glycoprotein of 57 to 65
(gp57-65) kilodaltons (kDa), has been extensively studied with regard toits synthesis, processing, and secretion (18). The gene encoding MDHV
gpS7-65
(i.e., MDHV-A) was* Correspondingauthor.
tJournal article 12564 from the Michigan Agricultural Experi-mentStation.
identified(16) andsequenced (8), and similar work is almost complete for HVTgp57-65 (i.e., HVT A antigen [HVT-A]) (P. M. Coussens and L. F. Velicer, manuscript in
prepara-tion). Theprocessing studies with MDHV-A(18) serve asa useful background for designing similar studies with other antigens in this system.
The B antigens of MDHV(MDHV-B) and HVT (HVT-B) weremolecularly characterized as a complex of three immu-nologically relatedglycoproteins,gplOO,gp6O,andgp49, one of which may be a membrane component, on the basis of studies withpolyclonal seracontaininganti-MDHV-B
activ-ity (17). Tunicamycin inhibition of N-linked glycosylation resulted in either nonglycosylatedor 0-linked glycosylated putative precursor polypeptides of MDHV-B and HVT-B, called pr88 and pr44, being immunoprecipitated with the same polyclonalsera(17). Inthatsame studythe
identifica-tion, as MDHV-B, of similar-size polypeptides, previously detected by others usingtheir(15, 41) monoclonal antibod-ies, was alsoaccomplished (17), thus making it possibleto relate their subsequent virus neutralization (15, 42),
immu-noprotection (36), and pulse-chase (14, 42) studies to MDHV-B.
Silva and Lee (42) have carried out pulse-chase studies with 10-min pulses and even longer chase intervals, using
HVT-infected cells and their monoclonal antibody reactive with the three MDHV-B glycoproteins. They reported that thedisappearanceofgplOO coincided with the appearance of
gp6Oandgp49, suggestingthat the former molecule may be
4270
on November 10, 2019 by guest
http://jvi.asm.org/
MDHV-B ANTIGEN GLYCOPROTEIN COMPLEX 4271
cleavedto generatethe lattertwomolecules. However, they
state that confirmation of therelationship of gpl00 to gp6O
and gp49 must await further experiments. Ikuta et al. (14), using HVT-infected cells, have also performed pulse-chase
studieswith similar time intervals, usingboth a monoclonal antibody and rabbit anti-MDHV-gB antiserum. They found essentially the same sequence of events as Silva and Lee (see above) and identified the larger glycoprotein as the precursorform of B antigen which is processed to the two smaller glycoproteins (14). However, in both pulse-chase studies no smaller unglycosylated precursor forms were detected, supporting the suggestion that cells may need to be pulsed foreven shorter periods of timeto visualize rapidly occurring early events (42). This may be especially true in view of the very rapid processing of MDHV-A, in which most ofthe glycosylationoccurswithin 1 min (18).
Since their pulse-chase experiments did not detect an
unglycosylated precursor polypeptide, Silva and Lee (42)
used lysates from tunicamycin-treated infected cells to re-solve this point. Their monoclonal antibody detected a 44-kDa molecule they called pr44 on the basis of their
interpretation that it was a precursor polypeptide. In their studyapr88 molecule was notreported, although reexami-nation of their data (42) (see Fig. 4) suggests that there may
have beensomeprotein ofpr88 sizepresent, but the amount was clearly less than that of pr44 (R. F. Silva, personal
communication). On the other hand, Ikuta et al. (14) de-tected what appears to be their equivalentof pr88, but no pr44, intunicamycin-treatedinfectedcells, usingtheir
mono-clonal antibody. While our laboratory has more recently found bothputative precursorpolypeptides after
tunicamy-cin treatment (summarized above), full analysis of their
possiblerole in the MDHV-Bprecursor-processing relation-ships wasbeyond the scope of the previous study(17).
The purpose of this study was to examine the entire MDHV-Bprecursor-processing questionin more depthand answer the existing questions by using approaches not
previouslytried. Thus, the analyses ofMDHV-B pr44
syn-thesis and processing reported here were performed with a
combination of higher-resolution pulse-chase studies, two
glycosylation inhibitors (tunicamycin and monensin), two
endoglycosidases (F and H), protease inhibitors, and cell-free translation and serve to provide a more complete analysis of processing events than previously existed. Of particular significance was the cell-free translation analysis followed by immunoprecipitation, whichrevealed that pr44 is the initial unglycosylated precursor polypeptide of MDHV-B. Ofequal significancewas theuseof monoclonal
antibody, which establishedthatpr44 was a component of all
larger moleculeswith MDHV-B reactivity.Thesequenceof
events elucidated by this combination of approaches and reagentsinvolvesthe apparentdimerizationof pr44 to pr88,
followed by glycosylation ofthisintermediate togpl00and
subsequent processing ofgpl00 to yieldpg6Oand gp49.
MATERIALS ANDMETHODS
Cells and viruses. Thepreparation, propagation,and infec-tion of small-scale duckembryo fibroblast cell cultures with MDHV was generally performed as first reported by
Glau-bigeret al. (11) and more recently by Isfort et al. (17). One
exception was that the MDHV strain GA used was at passage level 6, afterisolation ofcell-free virus from feather
tipsobtained from infected birds (39).
Radiolabeling of proteins. Infected and noninfected cells werelabeled with
[35S]methionine
at 48 h postinfection for 4hby standard methods,aspreviouslydescribed (17).
Pulse-chaselabeling was by methods already established for this system (18), with the following minor modifications. Prein-cubation without methionine was for 40 min (18); pulse labelingwasby incubation with 250
,uCi
of[35S]methionine
(specific activity, 1,000
,uCi/mmol;
AmershamCorp.)
perml for 1, 4, or 15 min; and chases were for the various timesindicated in the figure legends. For
labeling
in the presence oftunicamycin (Sigma
ChemicalCo.)
ormonensin(Calbio-chem-Behring), cellswerepreincubatedwith thedrugsfor 1 hasreportedelsewhere(18)or atvarious timesasindicated in the figure legends. Both
drugs
were presentthroughout
the
remaining
courseof eachexperiment.
Antisera.
Preparation
andcharacterizationof thepolyclo-nalantiserarabbitanti-MDHV-A(RoA), rabbit anti-MDHV-B(RaB), rabbit anti-MDHVinfected-cell
plasma
membrane (RaPM), and immune chicken serum have been describedpreviously
(17, 18). Themonoclonalantibody 1AN86.17,
animmunoglobulin Gl
isotype,
is a cloned derivative from hybridoma IAN86, and itimmunoprecipitated
gplOO,gp6O,
andgp49(datanotshown),as
previously reported
for 1AN86(41, 42). However, while this cloned monoclonal
antibody
reactedpoorlyinimmunofluorescencewith MDHV-infected
monolayer cell cultures, it exhibited a strong reaction with the surface antigen of infected cells in
suspension. Also,
itneutralized cell-freeHVT,butnotMDHV
(data
notshown).
This monoclonal antibodywas used becauseofits
immuno-precipitationproperties andespeciallybecause of its
ability
to
recognize
anepitope
presentinpr44 (42).
Immunoprecipitation and SDS-PAGE.
Immunoprecipita-tionwascarriedoutas
previously
described(17),
exceptthatlysate
preparation throughout
was withdetergent
buffercontaining
300 ,ug ofphenylmethylsulfonyl
fluoride perml,
and a second
antibody
was added as needed with mousemonoclonal
antibody
(asdescribedbelow).
Theimmunopre-cipitates
werewashed, suspended
insample
buffer,
andanalyzed bysodium
dodecyl
sulfate-polyacrylamide gel
elec-trophoresis
(SDS-PAGE)
(24).Acrylamide
concentrationswere 7.5% for all gels presented. Molecular sizes were calculated
by
interpolation
betweenprotein
standardsby
the method of Weber and Osborn(47). Fluorography wasper-formed
by
the methodofBonnerandLaskey (4).
Autoradi-ography
ofthe driedgels
was carried out at-70°C
for the times indicated in thefigure legends.
Enzyme digestions. All
digestions
wereperformed
withproteins
first bound toStaphylococcus
aureusprotein
Aduring
immunoprecipitation
as described above. Fordiges-tion with
endo-,-N-acetylglucosaminidase
H, also calledendoglycosidase
H(endo-H) (Miles Laboratories,
Inc.), immunoprecipitatedproteins
were eluted by incubating the bacterialpelletsfor 20 minat37°C
in 50 ,ulof sodium citrate buffer(50 mM, pH 5.5)containing
0.2%SDS,
thenboiling
for 2 min and
centrifuging
for 3 min. The supernatant wascollected,
10 U ofendo-H wasadded,
and the mixturewas incubated at37°C
for 16 to 24 h (1). Fordigestion
withendo-3-N-acetylglucosaminidase
F,
also calledendoglycosi-dase F (endo-F) (Dupont, NEN Research Products), the
immunoprecipitated proteinswereeluted in 100 ,ulof sodium
phosphate (100 mM, pH 6.1)
containing
0.1%SDS,
0.5% NonidetP-40,and 50mMEDTA. Endo-F(1/4
U)wasadded to thesample
and incubated as above. Controlsamples
without endo-H orendo-F were treated the same way, and afterincubation all sampleswere
precipitated
with ice-cold acetone for 1 h at4°C
andcentrifuged
in anEppendorf
microfuge
(model 5414)at topspeed
for 3 min. Theprecip-VOL.62, 1988
on November 10, 2019 by guest
http://jvi.asm.org/
4272 SITHOLE ET AL.
itateswerewashedwith100%ethanol, vacuumdriedbriefly, and suspended in sample buffer for electrophoresis.
Isolation of total cellular RNA. Total cellular RNA was isolated from MDHV-infected duck embryo fibroblast cells at 48 h postinfection by the guanidinium isothiocyanate procedure described by Chirgwin et al. (6). Cells were harvested, pelleted, and suspended in 4 ml of guanidinium isothiocyanate buffer (4 M guanidinium isothiocyanate, 1% sodium sarcosyl, and 0.1 M2-mercaptoethanol) per 108 cells. A portion (8 ml) of the resulting lysate was layered on 3 ml of 5.7 M cesium chloride in a Tris-EDTA solution (TE) consisting of 10 mM Tris hydrochloride (27) and 1 mM EDTA (pH 7.5). The RNA in the lysate was pelleted at 25,000 rpm for 20 h at 22°C in a Beckman SW41 rotor. The pelleted RNA was solubilized in TE with 0.1% SDS, ex-tracted with an equal volume of phenol-chloroform, ethanol precipitated, and solubilized in H20 (27).
Cell-free translation. Total cellular RNA from MDHV-infected cells was translated by use ofa rabbit reticulocyte lysate system from Bethesda Research Laboratories, Inc. The RNA (10 ,ug) was translated in a
30-pI
reaction volume as recommended by the supplier. An equal volume of detergent buffer was added to stop the reaction. A 10-jl portion of this diluted translation mixture was analyzed directly by SDS-PAGE, and the rest was immunoprecipi-tated, as described above, with undiluted antiserum or an ascitesfluid stock solution (ascites fluid previously diluted 1: 50 in phosphate-buffered saline) at 1/20 volume (i.e., for 50[L1
of translation mixture, 2.51l
ofantiserum or ascites fluid stock solution was used). To ensure successful immunopre-cipitation (see above), rabbit anti-mouse immunoglobulin G (Sigma) was added along with mouse ascites fluid stock solution at a ratio of 1:25, and rabbit anti-chicken immuno-globulin G was added along with immune chicken serum as previously described (17).RESULTS
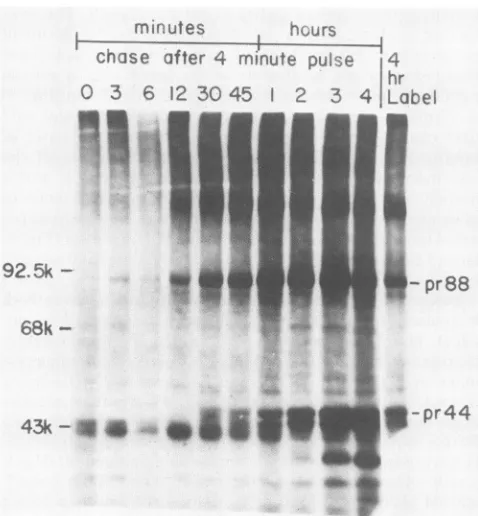
Kinetics ofMDHV-Bprocessing. Initially pulse-chase anal-yses were performed with 1-min pulses, as previously done for MDHV-A processing studies (18). The intent was to identify and analyze early rapid processing events, should they exist for MDHV-B, as in the case for MDHV-A. However, during preliminary experiments it became appar-ent that the appearance of molecules of interest was suffi-ciently slow(unpublished data) that a somewhat longer pulse interval could be used to facilitate isotope conservation and still critically analyze early MDHV-B processing events. Thus, for most subsequent pulse-chase analyses, infected cells were pulsed 4min and chased at various times (Fig. 1). The first molecule with MDHV-B immunological reactivity that could be detected, gplOO, was easily seen at the 12-min chase, although as early as 0-min chase trace amounts appeared (longer exposure of same gel, data not shown) as a heterogeneous slightly smaller putative glycosylation inter-mediate which acquired gplOO properties as glycosylation proceeded. The intensity of the gplOO band increased to a maximum at 45 min ofchase and declined with increasing chase periods. In contrast, gp6O and gp49 were first seen after a30-min chase and subsequently increased in amount, with the majority of the processing completed by 120min. It is important to note that the above events coincide in a manner consistent with a precursor-product relationship. The results of these pulse-chase studies with short pulses indicate that gplOO is processed relatively slowly to yield gp6O and gp49, confirming the earlier observations by Ikuta et al. (14) and Silva and Lee (42).
minutes
hours
92.5k
-
68k-43k
-chase ofter 4 minute
pulse
4hr
0 3
6
12 30 45
2
3 4 Label
gploo
...
*,it..::,.e,ls'W,S,.>l'; W -
g
6.
'
'
'5~~~~i15)49>g*gp
FIG. 1. KineticsofMDHV-Bprocessingasdeterminedby pulse-chase analysis in the absence oftunicamycin. A4-min pulse with
[5S]methionine was followed by chases of various times shown above each lane. Immunoprecipitationanalysis of1 mlof celllysate was done with RaPM. Fluorographic exposure was for 8 days. Molecularsizes areindicated inkilodaltons (k).
A careful examination of the data in Fig. 1 reveals two lines ofevidence that could beinterpreted tomeanthatsome gp49 forms directly by glycosylation ofasmaller precursor polypeptide, such as the pr44 described below. One obser-vation is theappearance of animmunoprecipitable
polypep-tideslightly smaller than gp49, but increasingin size, atthe 45-min and 1-h pulseintervals(Fig. 1and unpublished data).
The second observation is the accumulation ofmore radio-activity in the gp49 band than in the gp6Oband atthe 2-, 3-,
and 4-h chase intervals. Densitometric analysis (data not shown) confirms this visual observation. If gplOO is being
processed to yield gp6O and gp49, the latter two should be present in equimolaramounts. The greater accumulation of gp49could beexplainedifsome wasformedbyamoredirect route. While it is tempting to speculate about an alternate processing route, another explanation could be differential release ofgp6O and gp49 from membranes by the detergent
bufferused inthislaboratory (17), and/or
varying degrees
of susceptibility to protease degradation. In any event, if an alternate route does exist it would have to be a minor pathway and would not change the obvious existence of gplOO as aprocessingintermediate. Resolutionofthis ques-tion is beyond the scope of this study.Processing in the presence of monensin. Monensin, a mo-novalentionophore,inhibits transport of
glycoproteins
spec-ified by vesicular stomatitis virus, Sindbisvirus,
andherpes
simplex virus (HSV) (9, 19-21). In
addition,
thisionophore
blocks both theprocessing of N-linked
oligosaccharides
and the addition of 0-linked oligosaccharides to HSV type 1 glycoproteins (20, 21) and varicella-zoster virus gpll8 and gp98-67 (30, 31). To further examine theprocessing
of MDHV-B, lysatesfrominfected cells labeled in the presence and absence of monensin wereimmunoprecipitated
with RotPM and analyzed by SDS-PAGE. At apreviously
deter-mined optimal monensin concentration of 0.5 ,uM, and at various times of pretreatment, theamountsofgp6Oandgp49were reduced relative tothe amountofgplOO, and
gp6O
was J. VIROL.on November 10, 2019 by guest
http://jvi.asm.org/
MDHV-B ANTIGEN GLYCOPROTEIN COMPLEX 4273
minutes
chase after
15
minute
pulse 4 hr53045615
304660
Label_-
_ _ _
+ + ++
-_Monensin
92.5k-
68k-
43k-
25.7k-FIG. 2. The effect of monensin on MDHV-B processing as determined by pulse-chase analysis. A 15-min pulse with [35S]methionine was followed by chases of various times shown above each lane. Immunoprecipitationanalysis of the celllysate(1 ml) with (+) and without (-) monensin (0.5 ,uM) was done with
RoxPM.Fluorographicexposure was for 8 days. Molecular sizes are indicatedinkilodaltons(k).
replacedby aslightly faster migrating componentof about57 to58 kDa(unpublisheddata), suggesting inhibition of either
0-linked glycosylation or addition of terminal sialic acid
residues. Thisprelimninary finding suggested thatmonensin
inhibited processing of gplOO to gp6O and gp49. This was further confirmed by a modified pulse-chase experiment
performed in the presence of monensin. Since only later
processing events needed to be analyzed over a critical
period,thepulseinterval wasincreased from 4 min (Fig. 1) to 15 min to facilitate radioisotope incorporation, and only
four chase intervals were used (Fig. 2) on the basis of
information shown in Fig. 1. The appropriateness of the
conditions is shown in Fig. 2, lanes 1 through 4, where
normal processing (Fig. 1) can be seen in the absence of
monensin, with gplOO first appearing and becoming proc-essed to gp6Oand gp49.Inthe presence of monensin (Fig. 2, lanes 5through 8), however,gplOO accumulatesand no gp6O orgp49can befound.
Digestion with endo-F and endo-H. Endo-F cleaves both high-mannose and complex oligosaccharides but does not
affect 0-linked sugars (1, 23). Endo-H predominantly
cleaves between the twoproximalN-acetylglucosamine res-idues of high-mannose oligosaccharides (1, 23). Complex
oligosaccharides are resistant to endo-H, whereas
interme-diate oligosaccharides exhibit intermediate sensitivity. To
further evaluateglycoprotein processing, endo-Fand endo-HwereusedtodeglycosylateimmunoprecipitatedMDHV-B
glycoproteins (Fig. 3A and B, respectively). Both endo-F
and -H cleaved gplOO to pr88. Also, gp49, and to a lesser extentgp6O, were cleavedto pr44 by the same enzymes.
Analysis of the relationship between pr44 and pr88. The
consistent appearance of the two putative precursor
poly-peptides, pr44 and pr88, after metabolic labeling for 4 h in the presence of tunicamycin (17) (Fig. 4) suggested that
pulse-chase studies in the presence of the inhibitor might
provide information concerning the relationship between these two molecules. However, when such an experiment
was performed in parallel with, and identical to, the one
presented in Fig. 1, except in a second set of cell cultures
A
-+
-
+
ENDO-F
- -
+ +
TM
92.5k-h
---gplO0
0..
*-pr88
68k
-_b
--gp6O
43k-
VW25.7k-B
-+
-+
-+
Endo-
H
--
---+
+
Monensin
- -
+
+
- -TM
92.5k
--
-
i
gpIOO
ow
NEW40-prBB
68k--a
-gp49
-pr44
-
gp6O
.-.'-w W
_5rgp49
43k
-W
-pr44
25.7k
-FIG. 3. Deglycosylation of MDHV-B with endo-F and endo-H. (A) SDS-PAGE analysis of MDHV-B polypeptides labeled with
[35S]methionine
for4h in thepresence (+) andabsence(-) oftunicamycin(2jj.g/ml)immunoprecipitatedwithRaPM,eluted, and incubated in thepresence (+) and absence (-) of endo F. (B)SDS-PAGEanalysis ofMDH4V-Bpolypeptideslabeled with[35S]methioninefor 4 h in the presence(+)andabsence(-) oftunicamycin(2,ug/ml) or monensin (0.05R,M),immunoprecipitatedwithRaPM, eluted, and incubated in the presence (+) and absence (-) of endo-H. For both panels, fluorographic exposure was for 1 day and molecular sizes are indicated in kilodaltons(k).VOL.62,1988
on November 10, 2019 by guest
http://jvi.asm.org/
[image:4.612.62.280.74.301.2] [image:4.612.117.487.451.670.2]mrnuutes I m,,urs
cnhse
after 4' rfninute put e-i'hro 3 6 123045 2
I,w
i-
-4
La ce92.5k
-68k
-43k
-FIG. 4. Kinetics of MDHV-B processingasdetermined by
pulse-chase analysis in thepresenceoftunicamycin.Thisexperimentwas
performed in parallel with, and in an identical manner to, the
experiment presented in Fig. 1, exceptthat tunicamycin (2 pLg/ml)
was added to these cell cultures. Also, the fluorographic exposure
wasfor64days.
treated with tunicamycin, the hoped-fordirect evidence ofa
precursor-product relationship between pr44 and pr88 (in either direction) was not seen. The first molecule with
MDHV-B immunological reactivity that could be detected, pr88, was easily seen atthe 12-minchase and was apparent in trace amounts after the3-min chase, whena long
fluoro-graphic exposure was used (Fig. 4). The lower amount of pr88 inthe6-minchaselaneis duetolossof about50% ofthe sample,but thisdoesnot interferewith interpretation of the data. The amount of pr88 increased throughout the first3 h of chase (Fig. 4) and remained constant for hour 4 (more
apparent on a shorter exposure of the same gel, data not shown). pr44wasfirst detectedafter30min of chase (Fig. 4),
increased in amountthroughhour 3, and remained constant for hour 4 (more apparenton ashorterexposure of thesame gel, datanotshown). While pr88 increased inamountbefore pr44 did, the absence of any reduction in pr88 as pr44 increased is inconsistent with a precursor-product
relation-ship.
Withtwoputativeprecursorpolypeptides detected invivo
after labeling in the presence oftunicamycin, and only one
seen invitro (preliminary cell-free translation dataobtained
with polyclonal sera, not shown), there was concern that
pr44 might be an artifact of pr88 degradation by cellular proteasesin theformer situation. We examined the effect of four classes ofprotease inhibitors (2, 40)on theappearance ofpr44invivo, synthesized in thepresence oftunicamycin.
In all cases the ratio of pr44 to pr88 remained very similar (unpublished data), as would be expected if pr44 is a precursorpolypeptide andnotadegradation productof pr88.
Cell-free translation. Interpretation of data obtained from cell-free translation and immunoprecipitation analysis of the translation product is aided by careful choiceofantiseraand
ideally some prior informationconcerning the putative pre-cursorpolypeptides. While the RcxB and RoLPM sera gener-atedin thislaboratoryhavebeen useful in MDHV-B studies and detect pr44 made intunicamycin-treated cells, they are
of low titer(17). Thus, even the higher-titered RaPM must
be used in such quantities that the amountof
immunoglob-ulin G heavy chain present causes a distortion of protein
migration in that region ofthe gel, which in turn interferes with normal migration of MDHV-B pr44. Fortunately, a
panel ofmonoclonal antibodies that neutralize MDHV and
immunoprecipitate the MDHV-B polypeptides gplOO, gp6O, andgp49 has been prepared (41). The monoclonal antibody
1AN86.17 was ofparticular interest because the antibody
from the original hybridoma, 1AN86, was one of the more
extensively used and better-characterized members of the panel (41) andbecause itcouldimmunoprecipitate pr44 from
tunicamycin-treated cells (42), implying that it should be
possibleto useitsderivative (lAN86.17) to do the same from cell-free translation products. This antibody was first com-pared directly with RaPM (analysis in the same gel) with respect to their ability to immunoprecipitate the known MDHV-B polypeptides. As would be predicted from the
independent analyses (17, 41), both antibodies immunopre-cipitated gplOO, gp6O, and pg49 from MDHV-infected cells labeled in the absence of tunicamycin (data not shown). Both RoxPM and 1AN86.17 alsoimmunoprecipitated pr88 andpr44 formedin the presence of tunicamycin (data not shown). As predicted, there was no distortion of protein migration with 1AN86.17.This feature, and itsability toimmunoprecipitate
both pr88 and pr44, made 1AN86.17 the reagent ofchoice for identification of the MDHV-B precursor polypeptide made
bycell-free translation.
Once work shifted to analysis of the products resulting from cell-free translation of MDHV-infected-cell mRNA,
another point required clarification. The MDHV-A precur-sor polypeptide is very nearly the size (16, 18) of MDHV-B pr44 (17). In fact, its previous size estimate as pr47 was based on the previous estimate of the size of the ovalbumin marker as 46 kDa. Now that the supplier has changed this marker size to 43 kDa (17), on the basis of amino acid sequence data, it might be predicted that the MDHV-A precursorwill migrate close to MDHV-B pr44 in SDS-PAGE and that confusion could exist if both were present in the same gel because of either coprecipitation or nonspecific trapping. On the basis of our previous cell-free translation studies (16, 18) the MDHV-A precursor will definitely be present in any translation product resulting from translation of unselected MDHV-infected-cell mRNA. Furthermore, it will be immunoprecipitated with immune chicken serum, which has both MDHV-A and MDHV-B reactivity. There-fore, intentional coimmunoprecipitation of the two precursor polypeptides was done with immune chicken serum, with and without priorimmunoprecipitation by RotA, to differen-tiate the MDHV-B pr44 from the MDHV-A precursor. The results (data notshown) indicate that the apparent molecular weight of the MDHV-A precursor polypeptide (16, 18) is more appropriately presented as approximately 43 kDa (if ovalbumin marker is reported as 43 kDa), since it migrates veryclose toovalbumin and just below MDHV-B pr44. Now that the actualdifferences in the apparent molecularweights
of these two precursor polypeptides have been shown in the same gel lane, it is clear that MDHV-B pr44canbe identified without confusion.
Immunoprecipitation analysis of the product of cell-free translation of MDHV-infected-cell mRNA, with the mono-clonalantibody 1AN86.17, revealed a precursorpolypeptide
-
pr88
on November 10, 2019 by guest
http://jvi.asm.org/
[image:5.612.70.309.71.329.2]MDHV-B ANTIGEN GLYCOPROTEIN COMPLEX 4275
_1., ,~,1, ,~
L
1
Q Y
N
N
S
F
pr88-*:@
-
92.5k
-
68k
pr44-
-43k
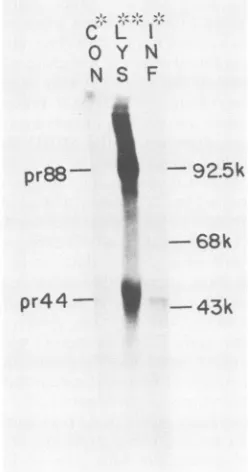
FIG. 5. Identification of the [35S]methionine-labeled MDHV-B precursorpolypeptidepr44 bycell-free translation and immunopre-cipitation. Immunoprecipitation and SDS-PAGE analysis of cell-free translation products (*) from infected (INF*) and uninfected (CON*) cellmRNAandoflysates (LYS**) of infectedcells treated with tunicamycin (2 ,jg/ml) by using monoclonal antibody 1AN86.17. Fluorographic exposure was for 8 days. Molecular sizes are asindicated in kilodaltons(k).
of 44 kDa(Fig. 5, lane INF) which comigrated with pr44 seen after immunoprecipitation of lysates labeled with [35S]meth-ioninein the presence of tunicamycin (Fig. 5, lane LYS). In contrast, the larger putative precursor molecule, pr88, also detected by immunoprecipitation analysis of these lysates
from tunicamycin-treated cells (Fig. 5, lane LYS), was not seen after cell-free translation and immunoprecipitation
analysis(Fig. 5, lane INF). This observation establishes pr44 astheprimary product of the MDHV-B gene.
pr88
*_
Formed in
presence
ofTucmcn
DISCUSSION
Processing of the MDHV-B glycoproteins. The already published detailed study of MDHV-A antigen synthesisand processing (18), which was done with the polyclonal antise-rumRoA,serves asprecedence for similar analyses involv-ing other MDHV-encoded antigens. In the current study, MDHV-B synthesis andprocessing events were elucidated by using similarhigh-resolution pulse-chase studies, a vari-etyofother methodsnew tothe system, andacombination of the polyclonal antibody RaPM and the monoclonal anti-body 1AN86.17. These two sera were useful because they were both known to immunoprecipitate the MDHV-B gly-coproteins gp 100, gp6O, and gp49 and theMDHV-Bputative precursorpolypeptides pr88 and pr44. The results obtained with this combination of approaches and reagents are sum-marized in Fig. 6.
While the pulse-chase studies were set up in a manner verysimilar to that ofstudiesused for MDHV-A (18), some results obtained during MDHV-B processing analysis were very different. First, no immunoprecipitable MDHV-B pu-tative precursorpolypeptides(either pr44orpr88)were seen in the absence oftunicamycin (Fig. 1), apparently because
glycosylationis cotranslational. In contrast, some MDHV-A precursor polypeptide was detected after a 1-min pulse, despite rapid glycosylation kinetics consistentwith cotrans-lationalglycosylation(18). The first MDHV-B moleculeseen was an apparent glycosylation intermediate slightly smaller than gplOO, and eventhen it was present in an immunopre-cipitable form in only trace amounts after a 4-min pulse (eightfold-longer fluorographic exposure of same gel used in Fig. 1). Labeled precursor molecules must have been present in normal amounts from the start, since MDHV-B intermediates and products appeared in expected amounts
later,butthey apparentlyare notaccessible to
immunopre-cipitation at theseearly times.
Presumablyduringcotranslationalglycosylation, the pre-cursor molecules were membrane bound and were not
solubilized efficiently by our detergent buffer; hence the failuretodetectexpected levels ofsignal,aswasfoundtobe thecase with HSV type 1 precursor to gB (43) and withthe latent membraneprotein, Imp63,of Epstein-Barr virus (28). The presumption that MDHV-B precursor polypeptides
N-LINKED GLYCOSYLATlON
Endoglycosidce
F and HHYPOTHESIZED
DMERIZATION STEP
*gp)OO
Mon
Tr
hhtion
fI2~~~~~~~~~~~~~~~W
pr44
g4Endoglyc
ses F and Hg49
Alsoformed by
Cell-free
tromotin
FIG. 6. MDHV-Bprocessing sequence summary. Heavy arrows denote biological pathway. VOL.62, 1988
on November 10, 2019 by guest
http://jvi.asm.org/
[image:6.612.122.246.77.313.2] [image:6.612.155.484.526.718.2]were present but undetectable at these early times was
supported by pulse-chasestudiesperformed in thepresence
oftunicamycin (Fig. 4), in which pr88 wasfirstdetected at the 3-min chase after a 4-min pulse, 3-min after a partly
glycosylated form of gplOO appeared in the absence of tunicamycin (Fig. 1). pr44 was first detectedafter a30-min
chase,after which it increased in amountin theabsence of
anyreduction in pr88. This is notthe resultexpectedif pr88 cleavage (degradation) resulted in pr44 formation. Appar-ently pr44 and pr88 are also membrane bound and not solubilized efficiently in our detergent buffer. While this
pulse-chase experiment in the presence oftunicamycin did
notdemonstrateaprecursor-processing relationship
consis-tent with dimerization of pr44 to pr88, as hypothesized
below, that fact cannot be interpreted as evidence against
such asequenceofevents. Insteadit is nowapparent, both in thepresence(Fig. 4) andabsence(Fig. 1)oftunicamycin,
that the earliest appearance ofeach MDHV-B molecule in
the gel is determined by when it becomes available for immunoprecipitation (by a mechanism yet to be deter-mined), notby the actual sequenceof processingevents.
The comparison of early processing events of MDHV-A and -Bthatinvolvedprimarilyglycosylationwarrantsfurther discussion. The heterogeneousMDHV-B molecules slightly smallerthan gplOO that were seen first (Fig. 1) were most probably glycosylation intermediates. While some details differed, suchdiffuse glycosylation intermediates were also
seenwith MDHV-A (18). MDHV-B wasglycosylated to its full-sizedgplOOform aftera3-min chase aftera4-min pulse. Similarly,MDHV-A reached its full-size form aftera6-min chase following a 1-min pulse. Possibly when similarities
occur,suchasglycosylationkinetics, theirexistence reflects
the involvement of the same cellular (duck embryo
fibro-blast) glycosylation mechanism. On the other hand, when differencesoccur,suchasfailuretodetect primaryprecursor
polypeptides (see above), fundamental polypeptide
differ-ences may be involved, since MDHV-A is predominantly
secreted (18), whereas MDHV-B appears to be membrane associated (17).
The pulse-chase studies reported here suggest gplOO is processed to form gp6O and gp49. The amount of gplOO molecule increases to a maximum at 45 min ofchase and then decreases. This decrease coincides very closely with the appearance of gp6O and gp49, in a manner consistent with a precursor-product relationship, thus confirming the
earlier observations ofSilva and Lee (42) and Ikutaet al. (14).Thisinterpretation isfurther substantiated in thisstudy by the observation that monensin inhibited processing of gplOO to gp6O and gp49 (Fig. 2). Use ofthis inhibitor is a majorextension of thepublished work referredtoabove and leadstoa veryimportant conclusionthatgplOOisactuallyan intermediate intheoverall biosynthesis ofMDHV-B. Since processing isslow,and gplOOisanintermediate, someof the moleculeis alwayspresentinthe cell. This conclusionraises
animportantnewquestion. Up untilnow we(17)and others (15, 41) have referred to a complex ofthe three glycopro-teins, designated gplOO, gp6O, and gp49 (17, 41), that we
identifiedasMDHV-B(18). Thesenewdata(summarized in Fig. 6) suggestthat themature forms ofMDHV-B are only gp6O and gp49, since gplOO is apparently an intermediate. Thepreviousinterpretationthatit ispartof thefinalcomplex
canbe understoodwhenoneconsiders the slowprocessing kinetics of gplOO and the long (4-h) labeling periods used previously (17). Possibly the profile of three polypeptides seenpreviously simply reflects the presence ofasignificant
amountofradiolabeled intermediate gplOOunder the
condi-tions
previously
used. On the otherhand,
when chase intervals aslong
as 20 h were used(data
notshown),
the amountofgplOO
was notreducedtozero,although
lesswas detected thanwasfound after the 4-h chasepresented
inFig.
1. Final resolutionof this
question
mayrequire
determina-tion of the arrangement of MDHV-Bpolypeptides
in the infected-cell nuclear andplasma
membranes.Endoglycosidase digestion of the MDHV-B
glycoproteins.
The useof
endoglycosidase
digestion
alone and in combina-tion with monensin was also a newapproach
tostudying
MDHV-Bprocessing
and alsogenerated
additional informa-tion that contributes to our overallunderstanding
of thissystem. Thefacts that
gplOO,
andnotpr88,
wassensitivetoendo-H and endo-F
(Fig. 3)
and that the formeryields
the latter upondigestion
suggest thatpr88
is the immediate precursor ofgplOO
and thus is an intermediate in thebiosynthetic
pathway.
The fact thatgp6O
exhibitedpartial
sensitivity
to both endo-F andespecially
endo-H indicatedthat it had
undergone
some modifications such as the addi-tion ofcomplex
sugarsor0-linkedglycosylation (9, 20, 21).
At lowerconcentrations of monensin
(.0.5
pLM)
(data
notshown)andatincreasedtimesofits pretreatment,
gp6O
wasreplaced by
afaster-migrating
molecule(data
notshown)
which does notcomigrate
with any of the endo-H- orendo-F-resistant
gp6O
species
(Fig.
3A andB).
However,
gp49 was
completely
sensitiveto both endo-F andendo-H,
indicating
thatonly
N-linked sugarswereaddedtothegp49
portion
of the moleculebeforecleavage.
On thebasis ofdatadescribed
above,
takentogether,
itappearsthatpr88
under-goescotranslationaladdition of N-linkedhigh-mannose
sug-ars toyield
gplOO,
which in turn isprocessed
togp6O
andgp49.
Apparently
gp6O
andgp49
aredifferentially
glycosyla-ted,
withgp49retaining
N-linkedhigh-mannose
sugars(en-tirely
endo-H and endo-Fsensitive)
and withgp6O
presum-ably
undergoing trimming
andpossibly
addition of0-linked andcomplex
sugars, assuggested
by
both the effect ofmonensin and
partial
sensitivity
to endo-H andendo-F.Identification of
pr44
as theprimary
MDHV-B precursorpolypeptide.
The role ofpr44
as theprimary
precursorpolypeptide
becomes clear when the full range ofdatanowavailable is considered.
First,
the cell-free-translation data(Fig. 5) clearly
indicate thatonly pr44,
andnotpr88,
is made invitro. This is not dueto premature termination oftrans-lation,
because othermoleculeslarger
thanpr88
aremade in our cell-free translation system.Also,
thepr44
comigrates
with thepr44found in cells after
tunicamycin
treatment(Fig.
5).
Second,
studies on the effects ofprotease inhibitors onpr44and
pr88
(data
notshown)
indicate thatboth molecules are made invivo,
since the former is not adegradation
product ofthe latter.
Third,
preliminary
partial
proteolytic
cleavage
analysis
ofpr44
andpr88
(data
notshown)
suggests
that the two molecules are related.
Fourth,
the results obtainedby
deglycosylation
ofgp6O
andgp49
by
endo-Fand -H(Fig.
3) and theimmunoprecipitation
ofall three mole-cules with the same monoclonalantibody
(41, 42;
data notshown)
indicate thatpr44
is thepolypeptide
backbone of eachglycoprotein.
Fifth,
the geneencoding
MDHV-B has beenidentified,
andanalysis by
hybrid
selection,
cell-freetranslation,
andimmunoprecipitation
(I.
Sithole,
P. M.Coussens,
and L. F.Velicer,
manuscript
inpreparation)
revealed a 44-kDapolypeptide
thatcomigrates
withpr44
seen after
tunicamycin
treatment.Sixth,
the fact thatpr44
from both the in vitro and in vivo sources areimmunopre-cipitated
by
thesamemonoclonalantibody
(Fig.
5)
indicates thatthey
areidentical.Finally,
Northern blotanalysis
witha MDHV-B geneprobe (Sithole
etal.,
inpreparation)
detectedon November 10, 2019 by guest
http://jvi.asm.org/
MDHV-B ANTIGEN GLYCOPROTEIN COMPLEX 4277
atranscript of 1.8 kilobases, a size appropriate toencode a
polypeptide of 44 kDa, as is the case for the MDHV-A
precursor polypeptide (16). The conclusion based on these
data is that pr44 is the MDHV-B primary gene product.
Thus, it is tempting to speculate (Fig. 6) that the initial processingstepinvolvesformation of pr88asadimer oftwo pr44 molecules. The finding that only pr44, and not pr88, forms in a cell-free-translation system suggests that this hypothesized dimerization may occur on the rough
endo-plasmic reticulum.
Theconceptof glycoprotein oligomerizationisnotwithout precedent in biology. Malek-Hedayat et al. (26) have char-acterized an endogenous lectinfrom cultured soybean cells
whichcanbe found asbotha30-kDamonomeranda60-kDa
dimer. Surprisingly, the dimerwas stable underconditions
such as heating in buffers containing SDS,
,-mercapto-ethanol, andurea. The two forms are interconvertible, with
theextentofinterconversion dependentontheconditionsof
prolonged incubation. The interconversion establishes that the dimer forms by noncovalent association (26). The fact that thedimercanbefoundinrootextractsisconsistentwith the idea that it may be more than an artifact that occurs duringstorage.Glycoprotein B (gB) encoded byHSVcanbe
found in both virions and infected cells in the form of detergent-stable oligomers (7)that have certain similarityto
MDHV-Bwithrespect tothe processing datareported here. Specifically, for both HSV gB (7) and MDHV gplOO, dimer-izationoccurred withinminutesofpolypeptidesynthesis and did not depend on N-linked glycosylation, since their pre-cursors both formed in the presence oftunicamycin. How-ever, HSV gB differs in one important way: it can be
disassociated by heat (7), whereas MDHV-Bpr88andgplOO
canwithstand boiling in SDS sample buffer (17).
Despite this difference it is tempting to speculate that dimerization is a step that is needed before glycosylation, since MDHV-B pr88 forms in the presence oftunicamycin
(Fig. 4). Experiments to resolve questions concerning the detailed nature of the dimerization process, such as the
nature ofthe bonds,the partof the peptideinvolved, andin the case of MDHV-B, the mechanism of separation of the
dimer(i.e., the processingofgplOOto yield gp6O and gp49),
are beyond the scope of the current work on MDHV-B.
Possibly dimerization of pr44 to pr88 and subsequent
proc-essing ofgplOO occur as the molecules move through
dif-ferent environments within the cell, and the existence of MDHV-Bdimersresistanttodissociation by heatrepresents
some difference between the properties of the polypeptides
of MDHV-B and HSV gB unrelatedtosomesortofcommon processing mechanism. However, it must be emphasized thatthenamesofthesetwoherpesvirus polypeptidesinclude the letter designation B for entirely unrelated reasons, not because they are homologs. The comparison ofthese two herpesvirus polypeptidesis thereforeinrelationtoapossible
common glycoprotein-processing mechanism, not because theyare genesconserved between the twoherpesviruses. In fact,werecentlylearned (T. M. O'Tal,P. M. Coussens, and
L. F.Velicer, unpublished data)thattheMDHVhomologof HSVtype 1 gB is located inadifferentregion of the MDHV genome than the geneencoding MDHV-B (Sithole et al., in preparation).
Possiblerole of MDHV-Bin MDHVmembrane biology and immunobiology. Our pulse-chase data (Fig. 1) indicate that
mature MDHV-B may not be the three polypeptides as we
have recently reported (17). Theresults summarized in Fig. 6indicatethat gp49 and gp6Oarethe mature fully processed final form of MDHV-B, whereas gplOO is an intermediate
that is processed slowly, and therefore small amounts are present atall times. Now that we know that gplOO is really an intermediate, and that a slow processing sequence is involved, we can moreaccuratelyplan future experiments to monitor the processing of MDHV-B polypeptides into the nuclear and plasma membranes. We predict thatgp6O and gp49 will both be in the membrane as mature forms of MDHV-B and that they will appear slowly accordingto the kinetics seenfor their processing. This leads to an interesting question: doesgplOO move tothe membrane, where it is then processed, or doesprocessing occurfirst, and then do gp49 and gp6O move to the membrane? On the basis of the monensin data, processing appears to occur in the Golgi apparatus, which would suggest that gplOO is processed to gp60and gp49, which then move to the membrane. Alterna-tively, disruption of the Golgi apparatus (20, 21) by
monen-sincould prevent transport of gpl00 to a site where process-ing can occur. We do not know whether processing is brought about by a cellular or virus-specific protease, or even if proteolysis is involved. Processing ofa larger mole-cule to two smaller molemole-cules has been observed for Rous sarcoma virus (22) and for at least four members of the herpesvirus family, pseudorabies virus (13, 25), bovine her-pesvirus (44), varicella-zoster virus (12), and cytomegalovi-rus (3, 5, 10). In some cases the processing involves
poly-peptide cleavage andformation ofdisulfidelinkages(3, 5, 10,
12, 13, 44). However, gp6O and gp49 are not linked by disulfide bridges (17). In vitro artifactual cleavage has been documented to be cell-line specific for HSV glycoproteins (38, 48). Althoughartifactual cleavage has not been demon-strated in human fibroblasts, gpl60ofhuman
cytomegalovi-rus is cleaved togpll6 and gp55 (5). Inthis MDHV study,all steps involving cell lysis and immunoprecipitation were carried out at4°C inbuffer containing the protease inhibitor phenylmethylsulfonylfluoride to preventnonspecific proteo-lytic cleavage.
The evidence that virus neutralization could be achieved with monoclonal antibody that also immunoprecipitated the three glycoproteins previously identified as the MDHV-B complex (17), as originally presented by Silva and Lee (41)
andIkuta et al. (15), suggests that someform of MDHV-B is the primaryantigen responsible for generating virus-neutral-izing antibody in vivo. Ono et al. (36) reported that
glyco-proteins purified by affinity chromatography with monoclo-nal antibody to gB (their terminology for the B antigen
complex) elicited partial protection against MD. While the relationship between viralneutralization and
immunoprotec-tion remains to be elucidated, it is assumed to be highly
significant (36). On the other hand, cell-mediated immunity
may be a factor as well. In either case the probable mem-brane location of these glycoproteins becomes important.
Determining the relationship of the MDHV-B polypeptides
to the cell membranes is part of the planned future
experi-ments on the kinetics of their insertion into the plasma and nuclearmembranes. Regardlessof the relativeimportance of virus neutralization and cell-mediated immunity, the ques-tion of whether all three glycoproteins are involved in protective immunity or whether just asingle one is involved must beasked. Since they all seem to contain thesamepr44 polypeptide backbone,eitherindividually(gp6Oandgp49)or in a dimer form (gplOO), they should all have the same epitopes that are determined by the polypeptide backbone. Possibly their role willbe determined more by their location inrespect to the surface of relevant membranes. The
signif-icance of the ideas presented above become more apparent when the observation (L. F. Lee, data not shown) that the VOL.62, 1988
on November 10, 2019 by guest
http://jvi.asm.org/
monoclonal antibody used in this study, 1AN86.17, reacts well with the surface of MDHV-infected cells in a membrane immunofluorescence assay is taken into consideration. Clearly the epitope recognized by this antibody is exposed on the cell surface. However, presenting a more complete answer to these questions concerning possible membrane locations and possible epitopes involved inprotective immu-nity is beyond the scope of this article.
ACKNOWLEDGMENTS
We thank R. A. Stringer for technical supportof the project. We also thank R. F. Silva of the U.S. Department of Agriculture Regional Poultry Research Laboratoryforevaluationof thiswork in relation to his own and Paul M. Coussens of the Department of Animal Science formanuscript review.
This research was supported primarilyby a grant from theElsa U. Pardee Foundation and partly by grant 86-CRSR-2-2870 under the Special Research Grants programadministered by theCooperative State Research Services, Sciences and Education, of the U.S. DepartmentofAgriculture.
LITERATURE CITED
1. Balachandran, N., and L. M.Hutt-Fletcher. 1985.Synthesisand processingofglycoproteingG ofherpes simplex virus type 2. J. Virol. 54:825-832.
2. Bartles, J. R., L. T. Braiterman, and A. L. Hubbard. 1985. Biochemical characterization of domain-specific glycoproteins of the rat hepatocyte plasma membrane. J. Biol. Chem. 260: 12792-12807.
3. Benko, D. M., and W. Gibson. 1986. Primate cytomegalovirus glycoproteins: lectin-binding propertiesand sensitivitiesto gly-cosidases. J. Virol. 59:703-713.
4. Bonner, W. M., and R. A. Laskey. 1974. A film detection method for triton labeled proteins and nucleic acids in poly-acrylamidegels. Eur. J. Biochem. 46:83-88.
5. Britt, W. J., and D. Auger. 1986. Synthesis and processingof theenvelope gp55-116 complex ofhuman cytomegalovirus. J. Virol. 58:185-191.
6. Chirgwin, J. M., A. G. Przybyla, R. J. MacDonald, and W. J. Rutter. 1979. Isolation ofbiologically active ribonucleic acid from sources enriched inribonuclease. Biochemistry 18:5294-5299.
7. Claesson-Welsh, L., and P. G. Spear. 1986. Oligomerization of herpes simplex virus glycoproteinB. J. Virol.60:803-806. 8. Coussens,P. M.,andL. F.Velicer.1988.Structureandcomplete
nucleotide sequenceoftheMarek's diseaseherpesvirus gp57-65 gene.J. Virol. 62:2373-2379.
9. Dall'olio, F., N. Malagolini, V. Speziali, G. Campadelli-Fiume, and F. Serafina-Cessi. 1985. Sialylated oligosaccharides 0-gly-cosidically linkedto glycoprotein Cfrom herpes simplex virus type 1. J. Virol. 56:127-134.
10. Farrar,G. H., and P. J. Greenaway. 1986. Characterization of glycoprotein complexes present in human cytomegalovirus en-velopes.J. Gen. Virol. 67:1469-1473.
11. Glaubiger, C., K. Nazerian, and L. F. Velicer. 1983. Marek's disease herpesviruses. IV. Molecular characterization of Ma-rek's disease herpesvirus Aantigen. J. Virol. 45:1228-1234. 12. Grose, C., D. P. Edwards, K. A.Weigle, W. E. Friedrichs, and
W. L. McGuire. 1984. Varicella-zoster virus-specific gpl40: a highly immunogenic and disulfide-linked structural glycopro-tein. Virology 132:138-146.
13. Hampl, H., T. Ben-Porat, L. Ehrlicher, K.-O. Habermehl, and A. S.Kaplan. 1984.Characterizationof theenvelope proteinsof pseudorabies virus. J. Virol. 52:583-590.
14. Ikuta, K., S. Ueda, S. Kato,and K. Hirai. 1984. Processing of glycoprotein gB related to neutralization of Marek's disease virus andherpesvirus ofturkeys. Microbiol. Immunol. 28:923-933.
15. Ikuta, K., S. Ueda, S. Kato, and K. Hirai. 1984. Identification withmonoclonalantibodiesofglycoproteinsofMarek's disease virus andherpesvirusofturkeys related to virusneutralization.
J. Virol. 49:1014-1017.
16. Isfort,R.J., H.-J. Kung,and L. F.Velicer.1987. Identification of the geneencodingMarek's diseaseherpesvirusAantigen.J. Virol. 61:2614-2620.
17. Isfort, R. J., I. Sithole, H.-J. Kung, and L. F. Velicer. 1986. Molecular characterization of Marek's disease herpesvirus B antigen. J. Virol. 59:411-419.
18. Isfort,R.J.,R.A.Stringer,H.-J. Kung,andL. F. Velicer.1986. Synthesis, processing, and secretion of the Marek's disease herpesvirusAantigen glycoprotein. J. Virol. 57:464-474. 19. Johnson,D.C.,and M.J.Schlesinger. 1980.Vesicular stomatitis
virusand Sindbis virus glycoprotein transportto the cellsurface isinhibited by ionophores. Virology 103:407-424.
20. Johnson, D. C., and P. G. Spear. 1982. Monensin inhibits the processing of herpes simplex virus glycoproteins, their trans-portto the cell surface,andthe egress of virions frominfected cells.J. Virol. 43:1102-1112.
21. Johnson, D. C., and P. G. Spear. 1983. 0-linked oligosaccha-rides areacquired by herpes simplexvirus glycoproteins in the Golgi apparatus. Cell 32:987-997.
22. Klemenz, R., and H. Diggelmann. 1979. Extracellular cleavage oftheglycoprotein precursor of Rous sarcoma virus. J. Virol. 29:285-292.
23. Kornfeld, R., and S. Kornfeld. 1985. Assembly of asparagine-linkedoligosaccharides. Annu. Rev. Biochem. 54:631-664. 24. Laemmli, U. K. 1970.Cleavage of structural proteins duringthe
assembly of the head ofbacteriophage T4. Nature (London) 227:680-685.
25. Lukacs, N., H.-J. Thiel, T. C. Mettenleiter, and H.-J. Rziha. 1985. Demonstration of three major species of pseudorabies virus glycoproteins and identification of a disulfide-linked gly-coprotein complex.J. Virol. 53:166-173.
26. Malek-Hedayat,S., S. A.Meiners, T. N. MetcalfIII,M. Schind-ler, J. L. Want, and S.-C. Ho. 1987. Endogenous lectin from cultured soybean cells. J. Biol. Chem. 262:7825-7830.
27. Maniatis, T., E. F. Fritsch, and J. Sambrook. 1982. Molecular cloning: alaboratory manual. Cold SpringHarborLaboratory, Cold Spring Harbor, N.Y.
28. Mann, K. P., and D. Thorley-Lawson. 1987. Posttranslational processing of the Epstein-Barr virus-encodedp63/LMPprotein. J. Virol. 61:2100-2108.
29. Marek, J. 1907. Multiple Nerventzuendung (Polyneuritis) bei Huchnern. Dtsch. Tieraerztl. Wochenschr. 15:417-421. 30. Montalvo,E.A., andC. Grose. 1986. Neutralization epitopeof
varicella zoster virus on native viral glycoprotein gpll8 (VZV glycoproteingpll8). Virology 149:230-241.
31. Montalvo,E. A.,R. T. Parmley,and C. Grose. 1985. Structural analysis of the varicella-zoster virus gp98-gp62 complex: post-translational addition of N-linked and0-linked oligosaccharide moieties. J.Virol. 53:761-770.
32. Murthy, K. K., and B. W. Calnek. 1979. Pathogenesis of Marek's disease: effect of immunization with inactivated viral and tumor-associated antigens. Infect. Immun. 26:547-553. 33. Nazerian, K. 1980. Marek's disease: a herpesvirus-induced
malignant lymphoma of the chicken, p. 665-682. In G. Klein (ed.), Viral oncology. Raven Press, New York.
34. Nonoyama,M. 1982. The molecularbiology ofMarek's disease herpesvirus, p. 333-346. In B. Roizman (ed.), The herpesvi-ruses, vol. 1. Plenum Publishing Corp., NewYork.
35. Okazaki, W., H. G. Purchase, and B. R. Burmester. 1970. Protection against Marek's disease by vaccination with a her-pesvirusofturkeys (HVT). Avian Dis. 14:413-429.
36. Ono, K.,M.Takashima,T.Ishikawa,M.Hayashi, I.Yoshida,T. Konobe, K.Ikuta,K.Nakajima, S. Ueda, S. Kato, and K. Hirai. 1984. Partial protection against Marek's disease in chickens immunized with glycoproteins gBpurified from turkey-herpes-virus-infected cells by affinity chromatography coupled with monoclonal antibodies. Avian Dis. 29:533-539.
37. Payne, L. N. 1982. Biology of Marek's disease virus and the herpesvirus ofturkeys, p. 347-431. In B. Roizman (ed.), The herpesviruses, vol. 1. PlenumPublishing Corp., New York. 38. Pereira, L., D. Dondero, and B. Roizman. 1982. Herpessimplex
virus glycoprotein gA/B: evidence that the infected Vero cell
on November 10, 2019 by guest
http://jvi.asm.org/
MDHV-B ANTIGEN GLYCOPROTEIN COMPLEX products comapand arise byproteolysis. J.Virol. 44:88-97.
39. Sharma, J. M., and H. A. Stone. 1972. Genetic resistance to Marek's disease. Delineation of the response of genetically
resistant chickenstoMarek'sdiseasevirus infection.Avian Dis.
16:894-906.
40. Shin, J., and T. H. Ji. 1985. Composition of cross-linked 1251-follitropin-receptor complexes. J. Biol. Chem.
260:12822-12827.
41. Silva, R. F.,and L. F. Lee.1984.Monoclonal antibody-mediated immunoprecipitation of proteins from cells infected with Ma-rek's disease virusorturkey herpesvirus.Virology 136:307-320.
42. Silva, R. F., and L. F. Lee. 1985. Isolation and partial charac-terizationof three glycoproteinscommon toMarek's's disease virusand turkey herpesvirus-infected cells,p.101-110. In B. W.
Calnek andJ.L.Spencer (ed.),Proceedings of the International Symposium on Marek's Disease. American Association of
AvianPathologists, Inc., KennettSquare,Pa.
43. Spear, P. G. 1976. Membrane proteins specified by herpes simplexviruses.I.Identification of fourglycoproteinprecursors
andtheirproducts intype1-infected cells. J.Virol. 17:991-1008.
44. Van DrunenLittel-Van Den Hurk,S., J. V. Van Den Hurk, J. E. Gilchrist, V. Misra, and L. A. Babiuk. 1984. Interactions of monoclonal antibodies and bovineherpesvirustype1(BVH-1) glycoproteins: characterizationof their biochemical and immu-nological properties. Virology 135:466-479.
45. Van Zaane, D., J. M. A.Brinkhof, and A. L. J.Gielkens.1982. Molecular-biologicalcharacterization ofMarek's diseasevirus. II.Differentiation of various MDV and HVT strains. Virology
121:133-146.
46. VanZaane, D., J. M. A. Brinkhof, F.Westenbrink, and A. L.J. Gielkens. 1982. Molecular-biological characterization of Ma-rek's disease virus. I. Identification of virus-specific
polypep-tidesininfected cells. Virology 121:116-132.
47. Weber, K., and M. Osborn. 1969. The reliabilityof molecular weight determination by dodecyl-sulfate-polyacrylamide gel electrophoresis. J. Biol. Chem. 244:4405-4412.
48. Zezulak, K.M., and P.G. Spear. 1984. Limitedproteolysisof herpes simplex virus glycoproteins that occurs during their extraction fromVero cells.J. Virol.50:258-262.
VOL.62, 1988 4279
![FIG. 2.determinedaboveRoxPM.ml)indicated[35S]methionine The effect of monensin on MDHV-B processing asbypulse-chaseanalysis.A15-minpulsewith was followed by chases of various times shown each lane](https://thumb-us.123doks.com/thumbv2/123dok_us/1330561.86925/4.612.117.487.451.670/determinedaboveroxpm-indicated-methionine-processing-asbypulse-chaseanalysis-minpulsewith-followed.webp)