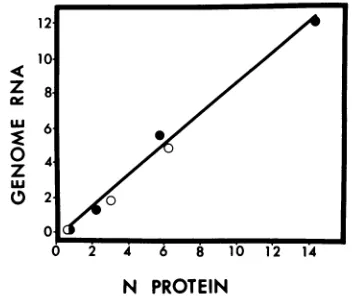
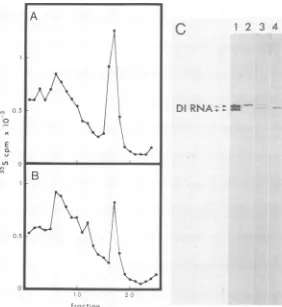
N protein alone satisfies the requirement for protein synthesis during RNA replication of vesicular stomatitis virus.
7
0
0
Full text
Figure

Related documents