Vol.65,No. 10
Identification of
a
Transactivating Function Mapping
to
the
Putative
Immediate-Early
Locus
of
Human
Herpesvirus
6t
MICHELLE E. D. MARTIN,* JOHN NICHOLAS, BRIAN J. THOMSON, CAROL NEWMAN,
AND ROBERTW. HONESS
Division of Virology, National Institute for Medical Research, London NW7]AA, United Kingdom Received19 April 1991/Accepted 18 July 1991
Sequencingstudies have indicated that the uniquecomponentof the human herpesvirus 6 (HHV-6)genome and the unique long segmentof the human cytomegalovirus genome are genetically colinear. Of particular interest is theidentification ofaregion oflocalCpG dinucleotidesuppression in thegenomeof HHV-6,afeature
conserved in thegenomes of human cytomegalovirus, murine cytomegalovirus, andsimian cytomegalovirus, and acharacteristic ofthemajor immediate-early loci oftheseviruses. Adjacenttothis region in HHV-6are approximately 30 copiesofa 103-to108-bpsequenceelement, which containsconsensusbinding sites forthe
transcriptionfactors AP2 and NFKB, in additiontoasingleKpnIrecognition site.Together,theseKpnIrepeat
units may compose an immediate-early enhancer, analogous to those found in the cytomegaloviruses. We presentthesequenceof this region of HHV-6 and demonstrate thatatransactivating functionis encoded bythis region.Wehaveused polymerase chain reactiontosynthesizefragments containing openreading framesand 5'sequenceswithorwithout theupstreamKpnIrepeatunits. Effectorplasmids containing these HHV-6 coding
and 5' sequences were able to effect activation of heterologous promoter-chloramphenicol acetyltransferase
(CAT)constructs, includingadenovirus E3-CAT and E4-CAT, human T-cell lymphotropic virus typeI long
terminalrepeat(LTR)-CAT,andhumanimmunodeficiency virus LTR-CAT, in cotransfection experiments in Verocells and peripheral blood lymphocytes. Furthermore, wehaveidentified the majoropenreadingframe
(RF4;2.3kb)asbeing essential for activation, andwehaveshownthat theNFKB,SP1, and TATA boxmotifs in the human immunodeficiency virus LTR are all required for full induction of the promoter by the
HHV-6-encoded transactivator.
Human herpesvirus 6 (HHV-6) is a recently identified herpesvirus, firstisolated following the in vitro cultivation of peripheral blood lymphocytes (PBLs) from six patients with lymphoproliferative disorders, two of whom were also in-fectedwith human immunodeficiency virus(HIV)(48). The
genome ofHHV-6consists ofalargely uniquesequence of 141 kbp with direct repeats ofapproximately 10 kbp at the termini. We haverecently constructed the restriction endo-nuclease map of the genome ofHHV-6 strain U1102 (40), andthis hasprovidedabasis forthefurthercharacterization ofthegenetic organizationoftheHHV-6genome. Previous
sequencing studies have indicated that the organization of
codingfeatures in the genome is colinear with the unique long segment of the genome of human cytomegalovirus (HCMV) (30). Furthermore, this has been confirmed and
extended by subsequent sequencing studies (lla, 39a, 57a, 57b). This sequencing has allowed us to predict the likely
location of theviral immediate-early (IE) regulatorygenes.
The IE genes are the first genes to be transcribed in
productive herpesvirus infections and are essential for the expression of delayed early genes, which are transcribed prior to viral DNA replication, and late gene expression.
Thus, the IEgeneproducts actastransactivators of
expres-sionfromhomologouspromotersand,inaddition,havebeen shown experimentally to transactivate heterologous viral and cellularpromoters (46). One such promoter isthe HIV
long terminal repeat (LTR), which has been shown to be transactivated by the IE genes of several herpesviruses, includingHCMV (9, 39, 41, 45),Epstein-Barrvirus(29, 37),
*Corresponding author.
t Wededicate this papertothememoryof BobHoness.
pseudorabiesvirus(59), and herpes simplexvirus(41, 42) by tat-independent mechanisms.
HHV-6 ishighlyprevalentin thepopulation,and virus has been isolated from the salivaofhealthyadults(4, 18, 32, 44)
andpersistsinperipheralblood ofamajorityofseropositive
individuals (15). Furthermore, HHV-6 has a predominantly CD4+ T-lymphocyte tropismboth in vivo and in vitro (35, 56) which, together with its ability to coinfect CD4+ T
lymphocytes with HIV and to transactivate the HIV en-hancer (12, 22, 34), has implicated HHV-6 as acofactor in the progression of HIV infection to AIDS. More recently
Lussoet al. (33) have shown that HHV-6 infection
upregu-lates the expressionofCD4, the major membrane receptor for HIVtype1 (HIV-1) (8),and thereforehasthepotentialto
expand therange ofHIV-1-susceptible cells. Thus, HHV-6
may actas a cofactor in HIV infection eitherby expanding therangeofsusceptiblecells and/orby regulatingthe
expres-sion of the HIV-1genes.Thegeneproduct(s)involved in this transactivation is yet to be characterized, although the
regulation has been reported tobe mediated either directly or indirectly through the NFKB sites in the HIV enhancer
region (34).
With a view to identifying the HHV-6 IE genes, our
interest has focused on a region near the right end of the
unique component of the HHV-6 genome corresponding preciselywiththelocation of themajorIEgenes inHCMV. This region shares with betaherpesviruses HCMV, murine
cytomegalovirus (MCMV), and simian cytomegalovirus (SCMV) boththe feature of local relative CpGdinucleotide
suppression (21)andacomplexarrayofrepetitivesequences
(5, 14, 26). We havesequencedtheputative IEgene locus of
HHV-6 and have identifiedopenreadingframes(ORFs)and
a putative enhancer of 3 kb, comprising KpnI repeatunits,
5381 JOURNALOF VIROLOGY, Oct. 1991,p. 5381-5390
0022-538X/91/105381-10$02.00/0
Copyright© 1991,American Society for Microbiology
on November 10, 2019 by guest
http://jvi.asm.org/
5382 MARTIN ET AL.
upstream of these ORFs. We show that effector plasmids containing this region transactivate avarietyof heterologous
promoter-chloramphenicol acetyltransferase (CAT) con-structs, including an HIV LTR-CAT target in transient expression assays, and that the major 2.3-kb
ORF,
RF4, is required for this function. Our studies using HIV LTR mutants have identified those sequences in the HIV LTR enhancer important for transactivation by the HHV-6 IE geneproduct(s).MATERIALS AND METHODS
Cells. Vero cell monolayers (Flow Laboratories, Inc., Irvine, United Kingdom) were grown at
37°C
in Dulbecco modified Eagle medium containing antibiotics (penicillin and streptomycin; 10 and 20 pLg/ml, respectively) and 10% new-born calfserum. PBLs were isolated by centrifugation onFicoll-Paquegradients (Pharmacia) and grown at
37°C
in5% CO2 in RPMI 1640 medium supplemented with 10% fetal calf serum and antibiotics (penicillin and streptomycin). PBLs, for use in transfection, were stimulated with both phytohe-magglutinin (5jLg/ml)
and human recombinant interleukin-2 (20U/ml; Boehringer Mannheim).Growth of virus and viral DNA preparation. All experi-mentsreported here employed a Ugandan isolate (U1102) of HHV-6 isolated by R. G. Downing (11). The propagation of the virus and subsequent purification of viral DNA for use in polymerase chain reactions (PCRs) were as previously de-scribed (40).
DNAsequencing. The isolation of the HHV-6 strain U1102 clonepHD12 (HindIII-C) and the PCR product from which the sequence presented in this report was obtained have been described elsewhere (40). The nucleotide sequence of sonicated DNA was determined from single-stranded tem-plate by using the dideoxynucleotide chain termination method (2, 49) or from the cloned DNA by using double-stranded sequencing methods (60). Sequence data were assembled by using the computer programs DBAUTO and DBUTIL (52, 53) and analyzed for the presence ofORFs
longer than 300 nucleotides by using the program ANALY-SEQ (54).
PCR. Oligonucleotides for use as primers in PCR were synthesized with an automated DNA synthesis machine (ABI 380B). The sequences (capital letters represent viral
sequences) of the primers used were as follows: RPT3, 5'
acacgaaTTCCAAAATTGGGAATCATGTGT
3'; RK1, 5'acacgaaTTCTCGGAAAATTAAAGGTCA
3'; RK2, 5' acacgaaTTCCTACCCAAGCGGGTTAGA
3'; RF1A, 5' acacggatccACATCTAGGTTTCATCTAGC
3'; RF1B, 5' acacaagcttTTAAACATGTGACATATAAC
3'. The positions of theprimers used in PCR, with the exception of RK2, which lies in the unique sequence at the right end of the KpnI repeat units, are annotatedin Fig. 2. PCR was performed with 200
ng ofU1102 DNA as the template and 1,uM (final
concen-tration) of each primer in a reaction mix containing 50 mM
KCl, 10 mM Tris-HCl (pH 8.8), 1.5 mM MgCI2, 0.01%
gelatin, 250 ,uM each deoxynucleoside triphosphate, and S U of Taqpolymerase (Amplitaq; Perkin Elmer Cetus) in a final volume of 100
pul.
Initial denaturation at94°C for 6 minwas followed by 25 cycles of denaturation(94°C;
1 min), anneal-ing (55°C; 1min),extension(65°C; 12, 10, or 5 min for 8.6- to 7.5-, 5.4-, or 2.6-kb products, respectively). These primersallowedthe specific synthesis of the required products with a yield ofapproximately 500 ng to 1 p.g per reaction. The products were extracted with phenol-chloroform and chlo-roform. Fragments were then digested with the appropriate
restriction enzyme, gel purified (57), and cloned into the appropriate vectors.
HHV-6 U1102effector gene constructs. Genetic
manipula-tions described below were performed as describedby
Maniatis et al. (38). Vectors used in cloningexperiments
were pUC19 and pBC12/CMV/IL-2 (7). The primer pairproducts of RPT3/RK2 (7.5 to 8.6 kb) and RPT3/RK1 (5.4
kb) were designed with terminal
EcoRI
sites andcloned into pUC19. The RPT3/RK1 product was cloned intopUC19
tocreate pK28. The products synthesized with primers RPT3 and RK2 showed some size variation. Two different
clones,
pR56
and pR47, were isolated from the RPT3/RK2 productand differ only in the number of KpnI repeat units present
(pR56,
7 copies; pR47, 28 copies), as confirmed bypartial
digestion withKpnI.
The primer pair RF1A/RF1B product(2.6 kb) was cloned as a
HindIII-BamHI
fragment into pBC/CMV/IL-2 to give pBC-ORF.pR56.ml
was created byinserting an EcoRV linker (8-mer) at the unique
BstEII
site followingBstEII
digestion ofpR56
and end repair. Linker insertion was confirmed by digestion and double-stranded sequencing with a USB Sequencing Kit (United States Biochemical Corporation, Cleveland, Ohio)according tothe manufacturer's instructions.Heterologous target and effector genes. pLTR-CAT and pTAT were gifts from A. Akrigg. The construct pLTR-CAT contains the entire LTR of HIV-1, the cat gene from
SV2-CAT (16), and the polyadenylation-termination se-quences from the HCMV major
IE
gene (1) cloned into a modified pUC18 vector. The HIV LTR-CAT plasmidscon-taining mutations in the KB, TATA, KB+TATA, or
Spl
sites (gifts from G. Nabel) have previously been described (42).pU3R-I,
which contains the LTR (nucleotides 1 to 670) ofhuman T-celllymphotropic virus type I
(HTLV-I)
linked to CAT, has been described by Sodroski et al. (51). The plasmid pE4-CAT1 contains the adenovirus early E4 pro-moter (nucleotides -224 to +33 relative to the transcription initiation site) and pE4-CAT2 is identical except for muta-tions in each of the three ATF binding sites (43). The adenovirus early E3 promoter-CAT plasmid, pE3-CAT, has been described by Weeks and Jones (58). pMLP-CATcon-tains adenovirus major late promoter sequences between -260 and +33 (relative to transcription initiationsite) cloned
upstream of the cat gene and was a gift from C. Goding. DNA transfection and transient CAT expression assays.
Plasmid DNA was purified on cesium chloride-ethidium bromide gradients (3). Monolayer cultures were transfected essentially as described by Gorman et al. (16). Vero cell monolayers were split 1 day prior to transfection. Calcium phosphate precipitates were applied for 10 h prior to chang-ing the medium, and cells were harvested 48 h after changchang-ing the medium. The cell extracts were quantitated for protein concentration, and equal amounts of protein were assayed for CAT activity as described by Gorman et al. (16).
PBLs were transfected by using the DEAE-dextran method essentially as described by
Hammarskjold
et al. (17). PBLs were taken directly from 3-day-oldphytohemaggluti-nin-interleukin-2-stimulated cultures. Briefly, PBLs were washed twice with 10 ml of TD (140 mM NaCl, 25 mM Tris,
5 mM KCI, 0.5 mM
Na2HPO4
[pH 7.5]) and resuspended in 1.5 ml of TD containing 50 p.g of DNA and DEAE-dextran (500p.g/ml/107
cells). After a40-min
incubation at37°C,
cells were suspended in 10 ml of TD, washed once with medium, and incubated at37°C
in 10 ml of growth medium until harvest. Cells were harvested 48 h posttransfection and assayed for CAT activity as described by Gorman et al. (16).J.VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
HHV-6-ENCODED TRANSACTIVATOR 5383
Kpnl
NN MrKpni IJ A B D C ILIGIKI F Jjfl Ej77TR161.5kbp
I I~~~~~~~~~~~KKH
RF3
RF4 RF2 RFI
----
7c
a
1 kb
4 6 8 10 12 kbP
FIG. 1. The KpnI restriction endonucleasemap of the genome of HHV-6 strain U1102.
Ml,
a fragment in thedirect left repeat; Mr, a fragment inthedirect right repeat. Below the genome is an expanded restriction map of the regionsequenced. B,BamHI; Bs,BstEIl; E, EcoRV; H,HindlIl; K,KpnI. KpnIrepeat units,describedpreviously (40), are indicated. Positions of fourleftwardORFs are illustrated in shadedboxes and are designated RF1, RF2, RF3, and RF4.The percentage ofchloramphenicol converted to acetylated
forms wasquantitated by liquid scintillation spectroscopy. Nucleotide sequence accession number. The sequence of theputativeIEregion of the HHV-6 genome reported in this
article willappearin EMBL, GenBank, and DDBJ nucleo-tide sequence data bases under accession number M73681.
RESULTS
Sequence analysis of the putative IE locus. The position of theregionsequenced at theright end of the unique segment
ofthe HHV-6 genome strain U1102 is illustrated in Fig. 1. Theprinciple features of the DNA sequence of 5,900
nucle-otides within this region are illustrated in Fig. 2. The translations of the ORFs RF1 (1668 to 2045), RF2 (2203 to 2574), RF3 (2547 to 2879), and RF4 (2846 to 5146) are shown above the nucleotide sequence. The deduced translation products of the ORFs shared no significant amino acid sequencesimilaritieswith the IE products of the betaherpes-viruses HCMV, MCMV, and SCMV. The IE proteins en-coded by these previously sequenced IE genes do not themselves share extensive similarity, although large
glu-tamicacid-rich domains intheircarboxy-terminalregions, a
feature not shared by RF4, have been reported (28, 55). However, this region in HHV-6 exhibited local CpG
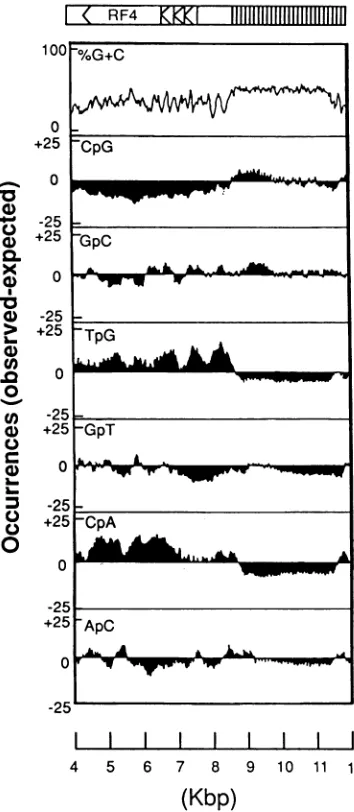
dinu-cleotide suppression (Fig. 3), which is a characteristic
fea-tureofthe betaherpesvirusmajorIE geneloci(21). TheCpG dinucleotide suppression is accompanied by an increase in
TpG and CpA dinucleotides, indicating exposure of this
regiontoDNAmethylation. Thisfeatureis not shared by the sequencesimmediately upstreamofthereadingframes (Fig. 3) orby anyothersequenced regionsof the HHV-6 genome
(30, 39a, 57b).
The positions of two overlapping putative AP1 binding
sites between nucleotide positions 593 to 603 (TGACTGA
and TGAGTCA) are indicated on the sequence (Fig. 2). Upstream ofthefirstORFare25to30
copies
ofarepeatunit containinga single KpnI site. The sequences of threecom-plete repeat units are presented (Fig. 2, nucleotides 64 to
375). Repeat units vary in length from 103 to 108
bp
andcontainaperfectconsensussequence forthe
binding
siteof thecellulartranscriptionfactor AP2(25)anda9of10match(GGGTCTTTCC) ofthe consensus sequencerecognized by
the NFKBfamily oftranscription factors
(GGGRATYYCC)
(31).There areonly minor sequence variations in eachofthecopies of the repeats. By analogy with the presence of multiply repeated transcription binding sites in the HCMV
andSCMVmajor IE enhancers (5, 24, 26, 36), it is possible thatthe HHV-6KpnIrepeatunits perform a similar function. Becauseoftheposition of ORFs colinear with HCMV IE genes, the relative local CpG dinucleotide suppression and the presence of upstream sequences with enhancerlike fea-tures, we investigated the possibility that trans-regulatory functions were encoded by this region.
Transactivation of heterologous promoters. Effector plas-mid pR56, which contains the four ORFs (Fig. 1), the 5' sequences, includingseven copies oftheKpnI repeats, and
800 bp of
3'
sequences, was used ininitial experiments. Todetermine the effects of gene products encoded by this plasmidonHIV-I LTR-directedtranscription, pR56(1to30 ,ug) wascotransfectedinto Vero cellswith 1 ,ug of the target
plasmid pLTR-CAT. The resultsoftheseexperiments
dem-onstrated that pR56 transactivated pLTR-CAT, with an
optimal induction of eightfold observed with 20 ,ig of ef-fectorplasmid (Fig. 4a).
Effector construct pR56 (40 ,Ig) was cotransfected with
pLTR-CAT (20 jig) into PBLs by using DEAE-dextran.
Expression was stimulated approximately eightfold over
basalexpression (Fig. 4b). Therefore, pR56 encodesa
trans-activator(s) which is functional in both epithelial and lym-phoid cells. Because of the relative ease ofcalcium phos-phate transfectionsand thehigherbasal levelsofexpression, furtherexperiments wereall performedin Vero cell
mono-layers.
To determine whether pR56 could activate
transcription
from other heterologous promoters, effectorplasmid pR56
was cotransfected into Vero cells with the target plasmids pLTR-CAT, pE4-CAT1, pE4-CAT2, pE3-CAT, pMLP-CAT, orpU3R-I(Fig. 5). pR56 induced levelsof
activity
of between2.5- and9.5-foldfor all theseheterologous promot-ers,andsimilar levels of activationwereobtained in numer-ous other experiments (not shown). pE4-CAT1 and pE4-CAT2, which has a mutation in each of the three ATFbinding sites, wereboth responsive topR56,
demonstrating
that the ATFsite is notrequired for induction.Identification ofthoseregions necessaryfortransactivation
incoding andnoncoding sequences. To establish whether the
largest ORFwas essential for the
transactivating
function,
wecreatedaplasmid,pR56.ml,whichcontainsaframe-shift
mutation in RF4. Thiswascreated
by
inserting
alinker in theBstEIIsite(Fig. 1)inpR56, and the mutationwasverified
by
sequencing. CATactivity
wasdramatically
reduced whenpR56.ml was cotransfected with the HIV LTR construct,
VOL. 65,1991
BH EK BS HB
O 2
on November 10, 2019 by guest
http://jvi.asm.org/
[image:3.612.131.486.77.205.2]J. VIROL. 5384 MARTIN ET AL.
Kpnl k8 Ap2
GCGA.ACGAGAACT,CTAAAAGCTGCTAACGCGGCAACCGCAGTTCCTGTTTTTCTTCATAAAG AGATCAGTGGGTACCGCGGT7OIGGGGTCTTTCCTGCACAGGCGAACGAGAACTCT
10 20 30 40 50 60 70 80 90 100 110 120
Kpn1 kB Ap2
v
AAAAGCTGCTAACGCGGCAACCGCAGTTTCTOTTTTTCT7CIGGAAA AAGGTCAGTOGGGTACCGCGGTTr.AGG7rTGTCCTACCCAGGCG)AACGAGAACTCTKACATCTGCTA.ACGCG
030 040 150 060 70 180 190 200 200 220 230 240
TKvnl kB Ap2
250 260 270.L 280 290 300 310 320 330 340 350 360
1 Ap2
370 J 380 390 400 410 420 430 440 450 460 470 480
Apl
490 500 500 520 530 540 550 560 570 580 590 600
610 620 630 640 650 660 670 680 690 700 700 720
730 740 750 760 770 780 790 800 800 820 830 840
850 860 870 880 890 900 910 920 930 940 950 960
970 980 990 1000 1010 1020 0030 1040 1050 0060 1070 0080
1090 1100 1110 1020 0130 0040 1050 0160 0070 0180 0190 1200
0210 1220 0230 0240 1250 0260 1270 0280 1290 0300 1310 1320
0330 1340 0350 1360 1370 0380 1390 1400 1400 0420 1430 1440
0450 1460 1470 0480 1490 0500 0500 1520 1530 0540 0550 0560
K Q TOIO
1570 0580 0590 1600 0610 1620 0630 1 640 1650 166 0 1670 1680
DI F F F IL 60E C KO IOYN 0 010100X TOY P s L T 8500S F 06A V GO 8 C
1690 1700 1710 0720 1730 1740 0750 1760 1770 1780 1790 1800
A Y3M L QOR R608 TOSE 00 L 60Y LW P o I 100156810I A 16818068SR
1800 1820 1830 1840 0850 1860 0870 1880 0890 0900 1910 1920
A S I F R LV F POVP 000DFF P1I8MV00 TOD V 608E L C 66800080GV
1930 1940 0950 1960 1970 1980 1990 2000 2010 2020 2030 2040
2050 2060 2070 2080 2090 2100 2000 2120 2130 2140 2150 2060
BOooHI S L L0L00L8 L 0KF N I 0Y
v
2170 2180 2090 2200 2210 2220 2230 2240 2250 2260 2270 2280
HindIII
8 P o P886M 6E0 I 66E I L E 6 S L MC 0 TOS F 00 C I P G L 6VS F 0 L I P
2290 2300 2310 2320 2330 2340 2350 2360 2370 2380 2390 2400
60088160I8E 806600006S G LOKT L AG 008801Y I C P6K1IE8N66K6
2400 2420 2430 2440 2450 2460 2470 2480 2490 2500 2510 2520
80S F 680 TO 81N L 8000S F TOS
8 TQNI F FOYIL A600608G C 10 010G 6006668606MQ
25r30- 2540 2550 2560 2570 2580 2590 2600 26010 2620 2630 2640
RFlB
Q0008606D L I K C AOG LCT 00008S F IG L O LE Q L 00Q L I 808 L L
26 50 2660 267 0 2680 2690 2700 2710 2720 2730 27 40 2750 2760
180 F A C 00K FOOGL
060 TOO 0600068SK L 8066K L D0FF N88 COEP 6 FM F L H0 I N80MG
2770 2780 2790 2800 2810 2820 2830 2840 2850 2860 2870 2880
0000606008ER L L L D L R86888801N I 060001D F 006 TOVT COD C
2890 2900 2910 2920 2930 2940 2950 2960 2970 2980 2990 3000
00K CN8006 T M LHMNACM 86 HNKE80 80 P0D 10000K F C 6018I T COO0T C
3000 3020 3030 3040 3050 3060 3070 3080 3090 3000 3110 3120
C 60686106IV TNAN608801I LOQT F 6100K MN 01I F MOO COO A F COK 3030 3140 3050 3060 3170 3180 3090 3200 3210 3220 3230 3240
on November 10, 2019 by guest
http://jvi.asm.org/
VOL. 65, 1991 HHV-6-ENCODED TRANSACTIVATOR 5385
L N V N C K N L I T A A K N L A N T I L 0 S I V I C S N E F S W 0 N L X L L R R
ACTTAATGTGAATTGTAAAAATTTAATTACTGCCGCCAAAAATCTTGCCAATACTATTCSCAGAGTASTGTTATATCCAGCAATGAA7TSATGGCAACACCTCAAACTTCTGAGACG
3250 3260 3270 3280 3290 3300 3310 3320 3330 3340 3350 3360
G F K V T M L N H I T O A C E C L E S D Y D D T G L I K P L T P L Q I N D G Y I
GGGATTTAAAGTTACCATGCTGAATATCATTACCCAGGCATGTCAATGCTTAGAA7CAGATTATrATGATACTCGTTTAATTAAACCSTrAACTCCCCTTCAAATAASr.ACGGCTACAT
3370 3380 3390 3400 3410 3420 3430 3440 3450 3460 3470 3480
N M N K N R 0 S S I C D G N T D P S D IS I L D L A D F D D H G R Y S E E S S I
TkATATGAATAAAAATAGACAGTCTTCCATCTGTGATGGAAACACAGACCCATCAGACTCAATGATATTACACTSAGC7SCATTTTGA7CACCAGGA,AGATATTCAGAGGAA7CCTTCTAT
3490 3500 3510 3520 3530 3540 3550 3560 3570 3580 3590 3600
E S I H E D D D N M YV P C T P S P E V P G K S X Y V G T F T E N S R Q S G D E
CGkATCTATCATGAAGATGATIGACAATAAkAATTATCCATGCACTCCTTCTCCAGAAGTGCCAG.GAAATCCAAATATGTAGGAACATTCACTGAAAACAGCAGACAGTCAGGGGACGA
3610 3620 3630 3640 3650 3660 3670 3680 3690 3700 3710 3720
Q T N P N C V G T A SV T D L G G P D N L N S I S G L 0 S C K N M L L E R L L D
ACAAACAAACCCTAACTGTGTAGGTACAGCCTCAGTGACAGATCTGGGCGGCCCTGATAACTTGAACTCCATCAGCGGCCTCCAGAGTSCTAAAA,ACATGSTCTAGAAAGACTACT7GA
3730 3740 3750 3760 3770 3780 3790 3800 3810 3820 3830 3840
T Q C D S V V E G T E Q D G S Y G N T L I S E N N N F G Y E T D H S A P Y E S E
CAC TIC ATIGCGATTCAGTAGTTIGAAGGGACAGAACAAGATGGTTCCTATCCCAACACTTTAATATCAGAAATGATGATCGTTTrCGTTATGAAACTGATSCACTCGGCACCCTATSGAGAGTGA
3850 3860 3870 3880 3890 3900 3910 3920 3930 3940 3950 3960
S D N N D E I D Y I A N S D S A KR T N N I H M N S T D E N T P F IS S I C S P
AAGCGATAACAATGA7GAAATTCArrACATTCCAAACTCTGATTCAGCTGCGAGAACTAATAACATTCATATCAATAGCACTGATCAGAATACACCATTTAGCAATTCTATATGSSCACC
3970 3980 3990 4000 4010 4020 4030 4040 4050 4060 4070 4080
P E V T P S K K N V K P E S MT P G S K P K 9 R V A K R K H V S S K S P K 8K K
ACCAGAGGTCACCCCTTCTAAGAAAAATGTCAAACCTAAGTICTATGACTCCG4GGAAGCAAACCTAAAAAGAGGGTAGCTAAAAGGAAACArTTTCAAGTAAATCACCCAAAAATAAAAA
4090 4100 4110 4120 4130 4140 4150 4160 4170 4180 4190 4200
1 K T D Q L P K A A D V I V I S S E S E D E E D G D N I I G N S I L I K A I K S
GATTAAGACAGATCAGTTACCTAAGGCTGCAGACGTAATTGTTATAAGTTCAGAGAGTGAAGATGAAGAGGATGGTCACAATATTATTGGTAACTCAATCTTAATAAAAGCTATTAAATC
4210 4220 4230 4240 4250 4260 4270 4280 4290 4300 4310 4320
E S D S E S S S E S N D C T S E H0 0 L H L S D Y D E V T N N G H C P S Y G F P
AGAATCAGATTCTGAATICGAGCAGCGAGTCAAATGACTGTACATCTGAACATAA,ACAGTTACATTTGAGTGATTATGATIGAAGTTACCAATAATGGTCATTGSCCAAGTTATGGSTTCC
4330 4340 4350 4360 4370 4380 4390 4400 4410 4420 4430 4440
T P V F T I P I R S Q GI T G G I S K F I P K0 N W I W Y H K K H80 V D N C
AACTCCAGTGTTTACCATCCCAATAAGGTC AATCAGGGTAC CGGTGGTATCAAATCCAA.ATTTATCCCAAAAAAAAITGATTTr.GATATGAAGAAAACTCACCAAGTAGACAATTG
4450 4460 4470 4480 4490 4500 4510 4520 4530 4540 4550 4560
P I H N S E N V D A K D D S D G T E AK H C F M N H F V P I K T D D E D Y D K N
TCCTATTCACAATTCTGAAAATGTGGCATCCAAAAGATCACAGTGATGGGACAGAAGCCAAACACTCCTTTAATCACTTTGTCC CCACAAAACGGATGATGAAGACTATGATAAGAA
4570 4580 4590 4600 4610 4620 4630 4640 4650 4660 4670 4680
N V S Y I Y N K I N S1 I D S G0D I I P T K R L I I D M V 0D N F H D L 9 D I
CAATGTCTCATATASTTATAATAAAATACAGAATAGCAAAATAGAC7CGGGAGACATAATACCAACTAAGAAACTTA7CATAGATATrGGTATGGATAATTTTATGATCTAkAAGATAT
4690 4700 4710 4720 4730 4740 4750 4760 4770 4780 4790 4800
1 0 Q G I T O H C Q D L C N K Y N V V T P T T C E D D L N9 T N S Q T F A T T A
CATTAAACAGGGCATAACCAAACATTGCCAGGA7TCrTGlCAATAAATATAATGSTCSGCACCCCTACTACATCSGAGCACGATTrAAATATGACAAATTCCCAAACSTTTCCAACAACCGC
4810 4820 4830 4840 4850 4860 4870 4880 4890 4900 4910 4920
T Q V F D P P V T G N N S S I L N I I N D T T S Q0 D 0 N R C T E GI S N S N E
ALACTCAAGTGSTTTGACCCACCGGTAACAGGAAATAATT,CCTCCATACTAAATATTATCAATr.ACACAACAAGTCAAAA7TGATGAAAACAGATGCACSC.AAGGTACAAGTAATTCTAA7TGA
4930 4940 4950 4960 4970 4980 4990 5000 5010 5020 5030 5040
OC T N I S D C N9 S D T E A F O L D G Y P S D Y D P F v E N A Q I Y
AAAATGTACCAATATAAGTGASSGCAACAGTGATGGTACAGAGGCCTAAATTAGAST.CTACCCTTCTSATTATGASCCTTTCGTAGAAAASCTCAAATCTATSAOCTAGATGAAAC
5050 5060 5070 5080 5090 5100 5110 5120 5130 5140 5150 RF1A 5160
CTAGATGTAAATTGATTAGCAATICATTAGCATACAGASGTGTATTATGTAAATATGCTrATACTGTTGTAATATAGACTTTCCCCATAr.GTr.TAACT CT,r.CTATAGATTA
5170 5180 5190 5200 5210 5220 5230 5240 5250 5260 5270 5280
Po11(A)
GAALATTACATAAGCAALATGTAAC71'TATTA7TGTAATACATCAACAAACACGTSGATTCC TCAAAr-TCAAATTTTCAATA7SCACCTTACAAATAATAAAALAGTAATATAGACATTG
5290 5300 5310 5320 5330 5340 5350 5360 5370 5380 5390 5400
CrrCTCIILLAATCCAAGAC ATGTACATICAACTGACTATA.AGTCATCATATAATATT7CCATAGTTAATTCCAGCTTTCTTAGAGACG7SATACTATAACAAST7GTATCT7
5410 5420 5430 5440 5450 5460 5470 5480 5490 5500 5510 5520
AAACCTTATAC AGAMTG AAGTATllCAGAITT AC CTCCCAACTAAACCAGTTGATTAJGAC T"AACCCCCGAGACATCTATTCTATAAATCATAGTACTGTTA
5530 5540 5550 5560 5570 5580 5590 5600 5610 5620 5630 5640
A.ATATATTACAlTAGC TATAGCTAGATCATAACACTC TACAALA/TATTT,TTlCTTTAGGTATATTAA7TCTGTTTT- GCTT-ACATCACTTATCATTATGAAXSACACATT
5650 5660 5670 5680 5690 5700 5710 5720 5730 5740 5750 5760
C AACCATTAGACTTTNGATGTCACACCAACACTThGAAGTATATAl:TCAATACCAAATICCATTTTTATTGCAACATClTATTAAiCTGATCACTTTs-GAAGGTAAGATGCTACACA
5770 5780 5790 5800 5810 5820 5830 5840 5850 5860 5870 5880
T7ArTTCCAATTT GAAAC
[image:5.612.135.478.94.661.2]RPT3 5890 5900
FIG. 2. Sequenceof5,900bpacrosstheputativeIEregion, readingfromrighttoleft
(approximately,
nucleotidepositions
9400to3500) as shown inFig. 1, withthetranslationproductsofthemajorORF RF4 and minor ORFsRF3, RF2,and RF1. The DNAsequenceincludes thefragments KpnI-J, 1.4kbofKpnI-K,and threecompletecopiesof theKpnIrepeatunits. Positions of therecognition
sites forselected restrictionendonucleases;thepolyadenylation signal; AP1, NFKB,and AP2consensus sequences;and theoligonucleotide primers
used in PCRareannotated. Thepositionof the PCRprimerRK2(notshown)isapproximately
3kbupstreamof theposition
of RK1. Therepeatunits, withinternalKpnI sites, areinbrackets.on November 10, 2019 by guest
http://jvi.asm.org/
5386 MARTIN ET AL.
a
Lane 2 3 4 5 6 7 Lane
b
luur%G+C
0
C1)
X
*-x
(n)
0
%00
C.)
C.)
C.)
0
0
25 CpG
-25
-25 GpC
0 .k& A~.. A,L, .
--25 t25
-TpG
O~~~~~~~1
-_
-25 +25 -GpT
i
o%L ""
O
n f-MNENEWW "nmm-2 +2
-2 +2
I
I
I
I
4 5 6 7 8 9 10 11 12
[image:6.612.90.267.84.490.2](Kbp)
FIG. 3. Frequencyofoccurrenceofselected dinucleotide pairs correspondingtotheregion of thegenomeof HHV-6 expanded in
Fig. 1. Themajor ORFs within this regionare indicated, together
with aplot of the mean percent G+C content. The other panels show thedifferencebetween theobserved andexpected number of
occurrences (observed-expected) of the dinucleotides CpG, GpC,
TpG, GpT, CpA, and ApC ina401-nucleotide sliding window with
datumpoints plottedevery5nucleotides and with plotsscaled from +25 to -25. A significant deficiency of CpG doublets occurs
between 4 and 9 kband isaccompanied byan excess of TpG and
CpA.
demonstratingthatthelargest ORFwasessential for efficient transactivation (Fig. 6a).
The regionencoding the transactivatorwasdefined more clearly by using pBC-ORF, which contains readingframes RF3 and RF4, under the control of the HCMV major IE
promoter/enhancer (see Materials and Methods). The amount of pBC-ORF required for optimal activation of
pLTR-CAT was determined from the results of
dose-re-sponseexperiments (1 ,ug; datanotshown). pBC-ORF (1 ,ug)
was cotransfected with pLTR-CAT (1
jig)
into Vero cells, andcellextractswere assayed for CAT activity. pBC-ORFinducedexpression of CAT significantly above basallevels
0:
*
**
* *@*
effector 0
/ug 5
10 15 20 30
°J6Ac 3.8 45 67 200 287 361 285
pUC pR56
%K.Ac 0.3 2.5
FIG. 4. (a) CAT activity induced in cotransfections of pLTR-CAT(1 jig)withincreasingamountsof the effectorplasmid, pR56 (as indicated), which contains the putativeIE sequences. Percent
acetylations (% Ac) are indicated below. Transfections were
per-formed in Vero cell monolayers as described in Materials and Methods. AmountsofDNAused in transfectionsweremadeupto
atotal of 31,ugwithpUC18. Fold inductionswere1.2, 1.8, 5.3, 7.5, 9.5, and7.5forlanes 2to 7, respectively. (b) Induction of pLTR-CAT (20 ,ug) expression by pR56 (40 jig) in PBLs. Cells were
transfectedbytheDEAE-dextranmethod,asdescribedinMaterials and Methods.
(Fig. 6a). Therefore, the sequences spanningRF3 andRF4
are sufficientfor the transactivating function.
Toexamine thepotentialeffectof theKpnIrepeatunitson theexpressionfrom the effectorplasmids,levels of
transac-tivation bypR47 (28 repeats),pR56 (7 repeats), andpK28 (0 repeats) were compared. All plasmids contain the same
sequences, including the AP1 sites, downstream of the repeats. The results shown in Fig. 6b demonstrate that at
approximately equimolaramounts of effectors pR56, pR47,
andpK28 (0.35, 0.28, and 0.28pmol, respectively),
compa-rablelevels ofexpression wereinduced in transient
expres-sionassaysin Veromonolayers, suggestingthat the minimal promoterelements are present in pK28 and that the KpnI
repeat units are notessential for expression in these cells. Since the level of activation, when equimolar amounts of effectorplasmidwereused,wasgreater withpK28than that induced by pR47, the KpnI repeats may in fact have a modest negative effecton expression. However, this repet-itiveregion containing multipleNFKB and AP2bindingsites
may actas an enhancer of HHV-6IEgene expressionin T cellspermissive for HHV-6productiveinfections.
Identification of the sequences within the HIV-1 LTR re-quired for HHV-6 transactivation. To identify functional
regionswithintheHIV LTRenhancer likelytobe important
forHHV-6transactivation, wetested thesetof site-directed mutantsthatwerepreviouslyusedtoidentifythelocation of cis-actingregulatorysequencesin the HIVLTRrequired for transactivation by HSV-1 and adenovirus ElA (42). The activities of the mutants were tested in Vero cells by cotransfection with pR56. HIV-1 encodes its own potent
transactivator, tat, and this allowed us to use an effector plasmid, pTAT, as a positive control and for comparative
purposesin theseexperiments. Figure7 illustratesthe levels
of CAT activity induced by pR56 (20 ,ug) or pTAT (5 ,ug) when cotransfected with the wild-type plasmid pLTR-CAT orconstructswithmutations in the KBsites, TATA box,Spl sites,or KB+TATA. Adecrease in theconstitutive
expres-sion ofCAT was observed with the mutants (Fig. 7). The results we obtained are clearly in agreement with those
I
<
RF4KKK[
L
,,W W.- , _
!5 -CpA
!5 ApC
O
I _,
-J. VIROL.
r
on November 10, 2019 by guest
http://jvi.asm.org/
[image:6.612.318.562.84.228.2]HHV-6-ENCODED TRANSACTIVATOR 5387
40-
35-
30-
25-0 c
20-0
15-0'I
10-
5-K
c
0
0II
K
pLTR-CAT pMLP-CAT pU3R-I pE3-CAT pE4-CAT1 pE4-CAT2
FIG. 5. Induction of the heterologous promoters pLTR-CAT (1kg),pMLP-CAT(1p.g), pU3R-1(1p.g),pE3-CAT(1rig).pE4-CAT1 (1p.g).
and pE4-CAT2 (1 ,ug) by effector plasmid pR56 (20 ,ug). Cotransfections were performed in Vero cell monolayers as described in Materials andMethods.
(a)
previously published, which demonstrated the importance of the Spl sites and TATA box motifs in tat-induced gene expression in both transient expression assays and by using
provirus constructs (13, 19, 20, 27, 47). The levels of expression induced by gene products encoded by pR56 were reduced in comparison to wild-type responses. pR56 activa-tionof the mutant KB LTR-CAT construct was reduced (4.3-to 2.7-fold), but the most dramatic effects on expression wereobserved with the other mutated target plasmids, with
activity reduced to near basal levels. The effect of the KB mutation on the relative induction of LTR-CAT by pTAT andpR56 isparticularly noteworthysince there is a specific
effect on activation by pR56, suggesting that the HHV-6 transactivator acts, at least in part, by a transcriptional mechanism.
c 0
a) C.)
0-
o-DISCUSSION
10- 9- 8- 7- 6- 5- 4- 3-
2-The conservation of the general organization of gene blocks within the HHV-6 genome has identified HHV-6 as
likely to be a member of the betaherpesvirus subfamily.
Furthermore, currentsequencedata suggest that theunique
segmentof the HHV-6 genome and theunique long
compo-nent ofthe HCMV genome arecolinear (lla, 30, 39a, 57a,
57b), and this has allowedustolocalizetheregionofHHV-6 mostlikelytoencode the IE genes. Sequencingof theregion
near the right end of the HHV-6 genome identified ORFs
corresponding precisely with the location of HCMV major
IEgenes, andthisregionisdepletedinCpGdinucleotides, a
feature sharedbytheIE genes of the
betaherpesviruses
(21). Upstreamof the ORFs isanarrayofrepetitiveelements(103to108bp) which havepotential bindingsites for the cellular
transcription factors NFKB and AP2, which are capable of
activatingtranscriptionin responsetoraisedlevels ofcyclic
Effector
(b)
12 11
10 9 8 7-6 5 4 3 2 1 0
pUCpR56pR56mlpBC-ORF pUC pK28 pR56 pR47
FIG. 6. Determinationof theregionsnecessaryfor transactivation in either coding (a) or noncoding (b) sequences. (a) Effects on LTR-CATexpressionof effectorplasr,idspR56 (20 ,ug);pUC18(20 ,ug); pR56.ml (20 ,ug), containingaframeshiftmutation in RF4; and pBC-ORF (1 ,ug),containing readingframes RF3 and RF4.(b)Effects of the number ofcopies ofKpnl repeat units on the induction of expressionfrompLTR-CAT. pK28(15 ,ug),pR56 (20 ,ug),and pR47 (20 ,ug), with0,7and 28copies, respectively,of the repeatunits, as
determined bypartialdigestion, were cotransfected into Vero cells withpLTR-CAT (1 ,ug)asdescribed in Materials and Methods. The optimalamountsof each effector shown hereweredeterminedfrom dose-responseexperiments (notshown).TotalamountsofDNAused intransfections wasmade upto21,ug withpUC18.
VOL.65, 1991
on November 10, 2019 by guest
http://jvi.asm.org/
[image:7.612.126.499.75.342.2] [image:7.612.322.562.385.617.2]5388 MARTIN ET AL.
100-
80-
60-40
c 2
0 co20
(x) 1
8-<
16- 14- 12-
10- 4- 2-
0--/
I
HIVLIwttype FoldInduction 1 4.2 24
13 12 11 10 9
7-6 5 4- 3-
2-
1-Mutant kB Mutant
1 2.738.5 1 1.4 4.3 1 1.4 7.7
[image:8.612.141.481.83.379.2]Targets
FIG. 7. Stimulation of HIV-CATpromotervariantsby pR56. pR56 (20,ug) orpTAT (5 ,ug)wascotransfected withwild-typeormutant
LTRplasmids (1 ,ug) into Vero cell monolayers, as describedin Materials and Methods. Basal levels ofexpressionweredetermined with
pUC18as acontrol. Total transfected DNAwas madeupto21 jgwithpUC18.Thefoldstimulationsobtainedareindicatedbelow.
AMP(25). TheNFKBelements identified in HHV-6 have a pyrimidinerather thanapurineatposition4of theconsensus sequence (31). The effect which this substitution has on factor binding has not yet been established, but high copy
numbermaycompensateforpotentially low-affinity binding.
It is possible that this region may constitute an enhancer, sinceit shares manyfeatures with theIE regulatory regions of HCMV, SCMV, and MCMV (5, 6, 10, 26, 36). For, althoughthespecific organization of theseregions differ, all contain multiple binding sites for cellular transcription fac-torswithinreiteratedsequences. HCMVand SCMV contain NFKB andcyclicAMPresponse elements (5, 6, 24, 36).
The usual method for detecting IE viral transcripts in herpesvirus infections employs protein synthesis inhibitors
which allow theaccumulation of IE mRNAs while blocking synthesisofdelayed-earlyand latemessages. However,our
present inability to achieve efficient cell-free infections means thatthis method is notpracticable. Herpesvirus IE
genes encode transactivators of homologous and
heterolo-gous promoters (46), and we have used this property to
provide functional evidence that the right-end sequence encodes an IEgene(s).
By usingPCRwe havesynthesized and cloned fragments containing the ORFs from this region with or without the
repeatunits. The effector plasmids encoded transactivators which enhanced expression from avarietyof heterologous
promoters. Although the adenovirus E4 and E3 promoters
and the HTLV-I LTR have ATF binding sites as common cis-acting regulatory sequences, mutations in these binding
sites in the E4 promoter did not affect induction of CAT activity. Contrarytoother findings (12), thegeneproduct(s)
described here transactivate the HTLV-I LTR.This maybe because of the different cell types used in theseexperiments
and may therefore be a consequence ofthe availability of relevant cellulartranscriptionfactors.
We have found that the carboxy-terminal region of the
largest ORFis requiredfor the transactivation of
heterolo-gous promoters and that reading frames RF3 and RF4
(containedbetween nucleotides 2547 and5146)aresufficient for the transactivating function. The region (KpnI repeats)
upstream of theseORFs containspotential binding sitesfor the cellulartranscription factors NFKB and AP2. Although
thepresenceof repeat units in the effectorconstructsdidnot
appearto influence expression from these plasmids in our
experiments in Verocells, itis still possible thatthey may
confer positive and/or negative regulatory effects on gene
expressioninacell-specificmanner.Since HHV-6replicates onlyinactivated T cells and T-cell activation induces levels of active NFKB (50),the presence ofmultiple binding sites for NFKB implies that this is probably one of the cellular
transcriptionfactors likelytoplay arole inIEgene regula-tion. The role of theKpnIrepeats asregulatory elements in a numberof cell types iscurrently beinginvestigated.
In view ofthepossiblerole of HHV-6as acofactor inthe disruption oflatency and activation of HIV replication, we have investigated the responsiveness ofthe HIV LTR en-hancer to HHV-6-encoded products. In order to identify those sites in the LTR responsive to transactivation by HHV-6, site-specific promoter mutants were used in cotransfections with the HHV-6 effector plasmid, pR56.
Transactivation occursbyatat-independent mechanism, as
has been observed with other herpesviruses (39, 41, 42).
pUC18
pR56
* pTAT
MutantkB+TATA
1 1 2
K
J. VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
HHV-6-ENCODED TRANSACTIVATOR 5389
However, although no single specific cis-acting regulatory sequence has been found to be involved in induction by
HCMV or herpes simplex virus (9, 39, 41, 42, 45), the
adenovirus transactivator, ElA, appears to require the
TATA box element(42),andSpl sites havebeen implicated
as the responsive elements for the pseudorabies virus IE protein (59).We have found that the TATA box,Spl,andKB
sitesall playaroleintheinducibilityof theHIV-1 enhancer
by the HHV-6 putative IEgeneproducts. These datasuggest
thatactivation is mediated, at least inpart, atthe
transcrip-tional level,althoughposttranscriptional effectsmayalso be
involved.
Ensolietal.(12)demonstratedthat the region-103to-48
of the HIV LTRwas critical fortransactivation by HHV-6 andthatHHV-6 induced bindingof NFKB tothe LTR. The apparentdisparity betweenourresults and those of Ensoliet
al. (12) maybe attributedto the fact that these researchers were using mock- and virus-infected cells in transfections andtherefore the possibility that other viral transactivators are playing a role cannot be excluded. We have also used
thesemutantsinsuperinfection studiesand found that
trans-activation with infected cells was NFKB independent (data
notshown). Itis alsotrue,however, that theuseof different cell lines may have a qualitative effect on the ability of HHV-6IE transactivators tofunction.
Although Horvat etal. (23) identified two regions of the
HHV-6 (strain GS) which strongly transactivate the HIV
LTR, these fragments do not map to this locus of HHV-6
(strain U1102) (40). Our studies are the first to identify
reading frames ofan HHV-6 transactivating gene, and our studies have mapped this gene to a region ofthe genome possessing noncoding features characteristic of the IE re-gions of other beta herpesviruses. Our findings linking the
local CpG dinucleotide suppression with a transactivating functionprovide further evidence that the uniquecomponent
of HHV-6 and the unique long component of HCMV are colinearand thatHHV-6 isclosely relatedtomembers of the
beta herpesvirus subfamily. We have shown that a gene product(s) from this locus transactivatestheHIV-1enhancer and therefore may play a role in the regulation of HIV replication. Lusso et al. (33) have recently shown that HHV-6 isinvolved in the upregulation of CD4 expression,
thusexpanding the range ofHIV-1-susceptible cells. It has been suggested that this function is mediated by the IE
genes. Therefore, the IE gene products may have dual functions intheir role as a cofactor in the control of HIV infection.
ACKNOWLEDGMENTS
We thank K. Cameron for technical assistance and Margaret Notleyfor all herpatience andcompetenceinthepreparation ofthe manuscript.
M. E. D.Martinwassupported by aStates ofJersey Education Department grant and B. J. Thomson by a Medical Research
Council(United Kingdom) Training Fellowship. REFERENCES
1. Akrigg, A., G. W. G. Wilkinson, and J. D. Oram. 1985. The structureof themajorimmediateearlygeneof human cytomeg-alovirus strain AD169. Virus Res. 2:107-121.
2. Bankier,A.T.,K. M.Weston,and B.G. Barrell. 1987. Random cloning and sequencing by the M13/dideoxynucleotide chain termination method. MethodsEnzymol. 155:51-93.
3. Birnboim,H. C.,andJ. Doly. 1979. Arapidalkalineextraction procedure for screening recombinant plasmid DNA. Nucleic AcidsRes.7:1513-1523.
4. Briggs,M., J. Fox, and R. S. Tedder. 1988.Ageprevalenceof
antibodytohumanherpesvirus6. Lanceti:1058-1059. 5. Chang,Y.-N., S. Crawford, J. Stall, D. R. Rawlins, K.-T. Jeang,
and G.S. Hayward. 1990. Thepalindromic series I repeatsinthe simian cytomegalovirus major immediate-early promoter be-have asbothstrongbasal enhancers and cyclic AMP response elements. J. Virol. 64:264-277.
6. Chee,M.S.,A.T.Bankier, S. Beck, R. Bohni, C. M. Brown,R. Cerny, T. Horsnell, C.A. HutchinsonIII, T. Kouzarides, J.A. Martignetti, E. Preddie, S. C. Satchwell, P. Tomlinson, K. M. Weston, and B. G. Barrell. 1990. Analysisof the proteincoding content of the sequence of human cytomegalovirus strain AD169. Curr. Top. Microbiol. Immunol. 154:125-169. 7. Cullen, B. R. 1986. Trans-activation of human
immunodefi-ciencyvirus occurs viaabimodal mechanism. Cell46:973-982. 8. Dalgleish, A. G., P. C. L. Beverley, P. R. Clapham, D. H. Crawford, M. F. Greaves, and R. A. Weiss. 1984. TheCD4(T4) antigenisanessentialcomponentof the receptor for the AIDS retrovirus. Nature(London) 312:763-767.
9. Davis, M. G., S. C. Kenney, J. Kamine, J. S. Pagano, and E.-S. Huang. 1987. Immediate-earlygene region ofhuman cytomeg-alovirus trans-activates the promoter of human immunodefi-ciencyvirus. Proc. Natl. Acad. Sci. USA84:8642-8646. 10. Dorsch-Hallser, K., G. M. Keil, F. Weber, M. Jasin, W.
Schaffner, and U. H. Koszinowski. 1985. A long and complex enhancer activates transcription of the gene coding for the highlyabundantimmediate earlymRNAinmurine cytomegalo-virus. Proc. Natl. Acad. Sci. USA82:8325-8329.
11. Downing, R. G., N.Sewankambo, D.Serwadda, R. Honess,D.
Crawford,R.Jarrett,and B.E.Griffin. 1987.Isolation ofhuman
lymphotropicherpesviruses from Uganda. Lancetii:390. 11a.Efstathiou, S. Personal communication.
12. Ensoli, B., P. Lusso, F.Schachter, S. F. Josephs, J. Rappaport, F.Negro,R.C.Gallo, andF.Wong-Staal. 1989. Human herpes-virus-6 increases HIV-1 expression in co-infected T cells via nuclear factors binding to the HlV-1 enhancer. EMBO J. 8:3019-3027.
13. Garcia,J. A., F.K.Wu,R.Mitsuyasu, and R.B.Gaynor. 1987. Interactions of cellularproteins involvedin the transcriptional
regulation of the human immunodeficiency virus. EMBO. J. 6:3761-3770.
14. Ghazal, P., H. Lubon, B. Fleckenstein, and L. Hennighausen. 1987. Binding oftranscription factors and creation ofa large nucleoprotein complex on the human cytomegalovirus en-hancer. Proc. Natl. Acad. Sci. USA 84:3658-3662.
15. Gopal, M. R.,B.J. Thomson, J. Fox, R. S. Tedder, andR. W. Honess. 1990. Detectionby PCRofHHV-6and EBV DNA in bloodandoropharynxofhealthyadults andHIV-seropositives. Lanceti:1598-1599.
16. Gorman, C. M., L. F. Moffat, and B. H. Howard. 1982. Recombinant genomes which expresschloramphenicol acetyl-transferaseinmammalian cells. Mol. Cell. Biol. 2:1044-1051.
17. Hammarskjold,M.-L., S.-C. Wang, and G. Klein. 1986. High-level expression ofthe Epstein-Barr virus EBNA1 protein in CV1 cellsand humanlymphoidcellsusinga SV40late
replace-ment vector.Gene 43:41-50.
18. Harnett, G.B.,T.J. Farr, G. R.Pietroboni,and M.R.Bucens. 1990. Frequent shedding of human herpesvirus6 in saliva. J. Med. Virol. 30:128-130.
19. Harrich,D.,J.Garcia,R.Mitsuyasu, andR.Gaynor.1990. TAR independent activation ofthehumanimmunodeficiencyvirus in phorbolesterstimulatedTlymphocytes.EMBO J.9:4417-4423. 20. Harrich, D.,J. Garcia,F.Wu,R.Mitsuyasu, J. Gonzalez,and R. Gaynor. 1989. Role ofSPl-bindingdomains in in vivo transcrip-tional regulation ofthe human immunodeficiency virus type 1
longterminal repeat. J. Virol. 63:2585-2591.
21. Honess,R.W.,U.A.Gompels,B.G.Barrell,M.Craxton,K.R.
Cameron, R. Staden, Y.-N. Chang, and G. S. Hayward. 1989. Deviations fromexpected frequenciesofCpG dinucleotidesin herpesvirusDNAsmaybediagnosticofdifferencesin thestates
of their latent genomes. J. Gen. Virol. 70:837-855.
22. Horvat, R.T., C. Wood,andN. Balachandran. 1989. Transac-tivation of human immunodeficiency viruspromoterbyhuman herpesvirus6.J. Virol.63:970-973.
VOL.65, 1991
on November 10, 2019 by guest
http://jvi.asm.org/
5390 MARTIN ET AL.
23. Horvat, R. T., C. Wood, S. F. Josephs, and N. Balachandran. 1991. Transactivation of the human immunodeficiency virus
promoterby humanherpesvirus 6 (HHV-6) strains GS and Z-29 in primary human T lymphocytes and identification of
transac-tivating HHV-6 (GS)genefragments. J. Virol. 65:2895-2902. 24. Hunninghake,G. W., M.M. Monick,B.Liu, and M. F. Stinski.
1989. The promoter-regulatory region of the major immediate-earlygeneof humancytomegalovirus respondstoT-lymphocyte stimulation and contains functional cyclic AMP-response
ele-ments.J. Virol. 63:3026-3033.
25. Imagawa,M.,R.Chiu,andM. Karin.1987.Transcription factor AP-2 mediates induction by two different signal-transduction pathways: protein kinase C and cAMP. Cell 51:251-260. 26. Jeang,K.-T.,D. R. Rawlins, P.J. Rosenfeld, J. H. Shero, T.J.
Kelly, and G. S. Hayward. 1987. Multiple tandemly repeated binding sites for cellular nuclear factor1that surround the major immediate-earlypromotersof simian andhuman
cytomegalovi-rus.J. Virol. 61:1559-1570.
27. Jones, K.A.,J. T.Kadonaga, P. A. Luciw, and R. Tjian. 1986. Activation of the AIDS retrovirus promoter by the cellular transcription factor, Spl. Science 232:755-759.
28. Keil, G. M., A. Ebeling Keil, and U. H. Koszinowski. 1987. Sequence and structural organization of murine
cytomegalovi-rus immediate-early gene1. J. Virol. 61:1901-1908.
29. Kenney,S., J. Kamine, D. Markovitz, R. Fenrick, and J. Pagano. 1988. An Epstein-Barr virus immediate-early gene product
trans-activates gene expression from the human
immunodefi-ciency virus longterminalrepeat. Proc. Natl. Acad. Sci. USA 85:1652-1656.
30. Lawrence, G. L., M. Chee, M. A. Craxton, U. A. Gompels, R.W. Honess,and B. G. Barrell. 1990. Human herpesvirus 6 is closely relatedtohumancytomegalovirus. J. Virol. 64:287-299. 31. Lenardo, M. J., and D. Baltimore. 1989. NF-KB: apleiotropic
mediator of inducible and tissue-specific gene control. Cell
58:227-229.
32. Levy, J. A., F. Ferro, D. Greenspan,andE. T. Lennette. 1990. Frequent isolation of HHV-6 from saliva and high
seropreva-lence of the virus in the population. Lancet 335:1047-1050. 33. Lusso, P., A. De Maria, M. Malnati, F.Lori,S. E. DeRocco,M.
Baseler, and R. C. Gallo. 1991.Induction of CD4 and suscepti-bility to HIV-1 infection in human CD8+ T lymphocytes by humanherpesvirus 6. Nature (London) 349:533-535.
34. Lusso, P., B. Ensoli, P. D. Markham, D. V. Ablashi, S. Z. Salahuddin, E. Tshachler, F. Wong-Staal,and R.C. Gallo.1989. Productive dual infection of human CD4+ T lymphocytes by HIV-1andHHV-6. Nature (London) 337:370-373.
35. Lusso, P., P.D. Markham, E.Tschachler,F.di Marzo Veronese, S.Z.Salahuddin, D. V. Ablashi, S. Pahwa,K. Krohn, and R. C. Gallo. 1988. Invitrocellular tropism of humanB-lymphotropic virus(human herpesvirus-6). J.Exp. Med. 167:1659-1670. 36. Mach, M., T. Stamminger,andG. Jahn.1989. Human
cytomeg-alovirus:recent aspects from molecular biology.J. Gen. Virol. 70:3117-3146.
37. Mallon, R., J. Borkowski,R.Albin,S.Pepitoni, J. Schwartz, and E. Kieff. 1990. The Epstein-Barr virus BZLFI gene product activates the human immunodeficiency virus type 1 5' long terminal repeat. J. Virol. 64:6282-6285.
38. Maniatis, T., E. F. Fritsch, and J.Sambrook. 1982. Molecular cloning: alaboratory manual. Cold Spring HarborLaboratory,
Cold Spring Harbor, N.Y.
39. Markovitz, D.M.,S.Kenney, J. Kamine, M. S. Smith, M. Davis, E.-S.Huang, C. Rosen,and J.S.Pagano.1989.Disparateeffects oftwo herpesviruses immediate-earlygene trans-activators on
the HIV-1 LTR. Virology 173:750-754. 39a.Martin,M.E.D. Unpublished data.
40. Martin, M. E.D., B. J. Thomson, R. W.Honess, M. A. Craxton, U.A.Gompels, M.-Y. Liu,E. Littler, J. R. Arrand,I.Teo, and
M.D.Jones. 1991. Thegenomeof humanherpesvirus-6: maps ofunit-length and concatemeric genomes for nine restriction endonucleases. J. Gen. Virol. 72:157-168.
41. Mosca, J.D., D. P. Bednarik, N. B. K. Raj, C.A. Rosen, J. G.
Sodroski, W. A. Haseltine, G. S. Hayward, and P. M. Pitha.
1987. Activationof human immunodeficiency virusby herpes-virusinfection:identification ofaregionwithin thelongterminal repeatthatrespondstoatrans-actingfactor encodedbyherpes simplexvirus l. Proc. Natl. Acad. Sci. USA 84:7408-7412. 42. Nabel, G.J., S. A. Rice, D. M.Knipe,and D. Baltimore. 1988.
Alternative mechanisms for activation of human immunodefi-ciencyvirus enhancer in T cells. Science 239:1299-1302. 43. Nicholas, J., andJ. R. Nevins. 1991. Distinct DNAtargets for
trans-activationby HTLV-1taxandadenovirus ElA. Virology 182:156-167.
44. Okuno, T., K. Takahashi, K. Balachandra, K. Shiraki, K. Yamanishi, M. Takahashi, and K. Baba. 1989. Seroepidemiol-ogy ofhuman herpesvirus 6 infection in normal children and adults. J. Clin. Microbiol.27:651-653.
45. Rando, R. F., A. Srinivasan, J. Feingold, E. Gonczol, and S. Plotkin. 1990. Characterization ofmultiple molecular interac-tions between human cytomegalovirus (HCMV) and human immunodeficiencyvirus type 1(HIV-1).Virology 176:87-97. 46. Roizman,B.1990. Herpesviridae:abriefintroduction,p.
1787-1794.In B. N.Fieldsand D. M. Knipe (ed.), Virology,2nd ed. Raven Press Ltd.,NewYork.
47. Rosen, C. A., J. G. Sodroski, and W. A.Haseltine. 1985. The location ofcis-actingregulatorysequencesin the human T cell lymphotropic virus type III (HTLV-III/LAV) long terminal repeat.Cell41:813-823.
48. Salahuddin,S.Z.,D. V.Ablashi,P. D.Markham,S. F.Josephs, S. Sturzenegger, M. Kaplan, G. Halligan, P. Biberfeld, F. Wong-Staal,B.Kramarsky, and R. C. Gallo.1986.Isolation ofa newvirus, HBLV, in patients withlymphoproliferative disor-ders. Science234:596-601.
49. Sanger,S.,S.Nicklen,and A. R.Coulson. 1977. DNA sequenc-ing with chain-terminating inhibitors. Proc. Natl. Acad. Sci. USA74:5463-5467.
50. Sen, R.,andD.Baltimore. 1986. InducibilityofK
immunoglob-ulin enhancer-binding protein NF-KB by a posttranslational
mechanism.Cell47:921-928.
51. Sodroski, J. G., C. A. Rosen, and W. A. Haseltine. 1984. Trans-acting transcriptional activation ofthelong terminal re-peatofhuman Tlymphotropic virusesininfected cells.Science 225:381-385.
52. Staden, R. 1980. A newcomputermethod for the storageand manipulation of DNA gel reading data. Nucleic Acids Res. 8:3673-3694.
53. Staden,R.1982.Automationofthe computerhandlingof thegel readingdataproduced bytheshotgun method ofDNA sequenc-ing. Nucleic Acids Res. 10:4731-4751.
54. Staden, R. 1986. The current status and portability of our sequencehandling software. Nucleic AcidsRes.14:217-231. 55. Stenberg, R. M., D. R. Thomsen, and M. F. Stinski. 1984.
Structuralanalysisof themajor immediate earlygeneofhuman cytomegalovirus.J. Virol.49:190-199.
56. Takahashi, K.,S.Sonoda,K.Higashi,T.Kondo,H.Takahashi, M. Takahashi, and K. Yamanishi. 1989. Predominant CD4 T-lymphocytetropismof humanherpesvirus6-related virus. J. Virol. 63:3161-3163.
57. Tautz, D., and M. Renz. 1983. An optimized freeze-squeeze methodforthe recoveryofDNAfragments fromagarosegels. Anal.Biochem. 132:14-19.
57a.Teo,I.A.,B.E.Griffin,and M. D.Jones. 1991.Characterization of the DNApolymerasegeneofhumanherpesvirus6. J. Virol. 65:4670-4680.
57b.Thomson,B.J.,andR.W. Honess. Unpublisheddata. 58. Weeks, D. L., and N. C. Jones. 1985. Adenovirus E3-early
promoter: sequences requiredfor activation by EIA. Nucleic AcidsRes. 13:5389-5402.
59. Yuan, R.,C.Bohan,F.C.H.Shiao, R. Robinson, H. J. Kaplan, and A. Srinivasan. 1989. Activation of HIV LTR-directed expression: analysis with pseudorabies virus immediate early gene.Virology 172:92-99.
60. Zhang, H., R. Scholl, J. Browse, and C. Somerville. 1988. Double-stranded DNA sequencing as a choice for DNA se-quencing. Nucleic AcidsRes. 16:1220.
J. VIROL.