Cytokine Response in Multiple Lymphoid Tissues during the Primary Phase of Feline Immunodeficiency Virus Infection
5
0
0
Full text
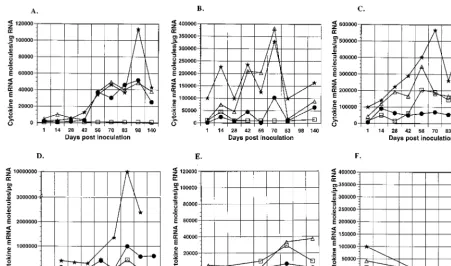
Figure

Related documents