Structural and
Functional
Characterization of rev-Like
Transcripts of
Equine Infectious Anemia Virus
RINA
ROSIN-ARBESFELD,l
MICHALRIVLIN,l
SILVIANOIMAN,1
PNINAMASHIAH,'
ABRAHAMYANIV,' TORU MIKI,2 STEVEN R. TRONICK,2* ANDARNONA GAZIT' Department of Human Microbiology, Sackler School of Medicine, Tel Aviv University,
Tel Aviv 69978, Israel,1andLaboratory ofCellular and Molecular Biology, Building 37, Room 1E24, National Cancer Institute,
Bethesda, Maryland 208922
Received 1 March 1993/Accepted 18 May 1993
Three cDNA clones representing
structurally
distinct transcripts were isolated from a cDNA librarypreparedfrom cells infected withequineinfectiousanemia virus(EIAV) byusingaprobe representing the S3 openreading frame,which isthoughttoencode Rev.Onespecies, designated p2/2, contained fourexonsand wasidenticaltoa
previously
described polycistronicmRNAthat encodes Tat. This transcriptwaspredictedto alsodirect the synthesisofatruncated form ofthetransmembrane protein andaputative Rev protein whose N-terminal 29 amino acids, derived from env, are linked to S3 sequences. The second cDNA, pl76, also consisted of fourexonswhichweregenerated bytwoof three of thesamesplicingeventsthatoccurwithp2/2 but notwith the Tat mRNA. The alternative splice sitegiving rise tothe second exon ofp176 results in abicistronicmessagethat would encode thesametransmembraneand Revproteinsasp2/2.The firstexonofthe thirdtranscript, p20,wasidenticaltothose ofp2/2andp176butwasspliced
directly
toS3.This monocistronic messagecould encodeasecondform ofRev that lacksenvsequences,providedthatRevsynthesiswould initiate atanon-AUG codon.Thecoding capacityofeachcDNAwasassessed inaeukaryoticsystemusingS3antisera. TwoputativeRevproteinswith apparentmolecularmassesof 18 and 16kDawereexpressedby p2/2andp176,while p20 expressed only a 16-kDa species. Analysis of EIAV-infected cells with S3 antisera revealed the presence ofan 18-kDa protein.
Surprisingly,
the sameproteinwas detected in purified virions. By usinga reporterconstruct,thechloramphenicolacetyltransferasegenelinkedtoEIAVenvsequences,we wereableto demonstrate greatly enhanced chloramphenicolacetyltransferase
activity
in cells cotransfected with this constructandanyof the threecDNAs.Thegenomeofequineinfectious anemia virus(EIAV)(20, 42),alentivirus,isstructurallylesscomplexthan those of all other members ofthis virussubfamily (14).Inadditiontothe threemajoropenreadingframes(ORFs),gag,pol,andenv, threesmallORFs, designated Si, S2,andS3,arepresent.Si encodessequencesrequiredfor the function of the EIAVtat
gene (11, 33, 45). There ispersuasive but indirect evidence thatS3 sequencesrepresentrev(35, 47),whereas the func-tion ofS2 is unknown. The pattern of EIAVgeneexpression
in infected cells belies its simple genomic structure and resembles that of its muchmore complexcounterparts (8).
Thus, at leastfivespeciesofvirus-specific transcriptswere observed (34), and analysis of cDNA clones has revealed that the smaller bands present in Northern (RNA) blots
actually represent multiple mRNA species (33, 35). In our
analysis of cDNAlibraries ofan EIAV-infected canine cell line (33, 35),wedemonstrated that theEIAV Tatproteinis encoded by at least three alternatively spliced transcripts.
One mRNAwasshown tobepolycistronic, encodingTat, a
putative Rev, and/or a truncated transmembrane (ATM)
protein. Another bicistronic tat message that could also potentially direct the synthesis of a truncated TM protein was isolated. A monocistronic tat mRNA was also de-scribed. We found that the relative abundance of these transcripts differed in infected cells, and trans-activation assaysshowedthat thetatactivityof themonocistronicform
*Correspondingauthor.
was significantly higher than that of the more complex
species (35).
Here we present the characterization of putative EIAV Revtranscripts isolated from cDNA librariespreparedfrom EIAV-infected cells by screening for recombinant phages containing S3 sequences. Although one clone, designated
p2/2, was found to be identical to the previously reported polycistronictat clonep105 (33), two other distinct cDNA
clones, designated p176andp20,werefound. These cDNAs were isolated by using pCEV vectors which allow highly
efficient directionalcloningof functional full-length cDNAs whicharereadilyconvertedintoaplasmidform forrescuein bacterial cells(30).Forsequenceanalysis,the cDNA inserts were subcloned into pBluescript vectors (Stratagene) and
analyzedwith the Sequenase DNA sequencingkit (United
States Biochemical Corp.), as previously described (35). Clonep176 (Fig. 1)was1,361 bplong,and its first nucleotide
correspondedtoposition223 of the EIAVgenomicsequence (20),which is 16 bpdownstream of the transcription start site. Comparison of the nucleotide sequenceof this cDNA with thegenomicsequencerevealed that itwascomposedof fourexons(Fig.1 and2).Exon 1 extendedtotheconsensus
splicedonor siteatposition459. The secondexonstartedat base 5210, located approximately in the middle ofS1 and upstream of env, and terminated at the splice donor site
previouslyshowntobe used togeneratetatmRNAs
(posi-tion5276) (33, 35). The thirdexonstarted within S2atbase 5437 and extendedtobase 5534. The fourthexonstartedat
the firstbase of S3(position 7232) and endedatthepoly(A)
site within the long terminal repeat. Thus, this cDNAwas
5640
on November 9, 2019 by guest
http://jvi.asm.org/
10 20 30 40 50 60 70 80 90 100 CGGTCTGAGTCCCTTCTCTGCTGGGCTGAAAAGGCCTTTTAATAAATATAATTCTCTACTCAGTCCCTGTCTCTAGTTTGTCTGTTCGAGATCCTACAG
110 120 130 140 150 160 170 ISO 190 200
TTGGCGCCCGAACAGGGACCTGAGAGGGGCGAGACTACCTGTTGAACCTGGCTGATCGTAGGATCCCCGGGACAGCAGAGGAGAACTTACAGAAGTCTTC
Exon 1 4591 4S Exon 2 [52101
210 220 230 240 250 260 270 280 290 300
TGGAGGTGTTCCTGGCCAGAACACAGGAGGACAGAGGAAGAAGAATA^A AAAGACTGAAGGCAATCCAACAA(GGAAGACAACC-TC.AATA TT TGTTAIAA
II K11 e G N
Tm
P TI1)VH~
R K S TExon 2
[5276
Sl)
T (altrnate frame in Exon 4(72321
Exon 3 [5
41
7 Exon 3[55341j
'
4-
-310 320 330 340 350 360 370 380 390 400
t
(SU
codon 43)
'.'.'.''
.'''...'
K.'
''.K
'.'.-.K-.
K'''"''
K-'V'
K ''I''''.'#.'-'"t (alternate frame in SU)
410 420 430 440 450 460 470 480 490 500
D P Q G P LWSE S W C R V P E E K I P S Q T C I A R
...* .. ,.
t (TM codon 197)
S10 520 530 540 550 560 570 580 590
600
IHF L A P G. P T 0 T P s R R D R W T R a 0 T L a T E V L F R
LF.I
...-- ---WK
610 620 630 640 650 660 670 680 690
700
rAr,AATrR-ArAr. r. ,TArlAAAeRlRlRRrIMrs AAAr,Ar,r.r.r^RTT Arer r.T* Arr,CAT TTR rARAR^r,Ar, TT T^n^R
ARnn
*,
*RAr TI RGB V QQAAKFn K rFA P E L Y F R E R G D I9
710 720 730 740 750 760 770 780 790 800
~ .6.
A*F
p AL-P
R V L s810 820 830 840 850 860 870 880 890 900
r,-TTT,*TGACTGTTGCATTAAAGCCCAAGAAGGAACTCTCGCTATCCCTTGCTGTGGATTTCCCTTATGGCTATTTTGGGGACTAGTAATTATAGTAGGA
...--..W...
910 920 930 940 950 960 970 980 990 1000
CGCATAGCAGGCTATGGATTACGTGGACTCGCTGTTATAATAAGGATTTGTATTAGAGGCTTAAATTTGATATTTGAAATAATCAGAAAAATGCTTGATT
5.14.0V"~~~~'L'* * 4. A ~~~~ V ~~1 ft '1 0 1 ft S N.. LI. ..I....I .~~..-...~~~~~~~.I...5 1 .1.
1010 1020 1030 1040 1050 1060 1070 1080 1090
1100
ATATTGGAAGAGCTTTAAATCCTCGCACATCTCATGTATCAATGCCTCAGTATGTTTAGAAAAACAAGGGGGGAACTGTGGGGTTTTTATGAGGGGTTTT
lio 1120 1130 1140 1150 1160 1170 1180 1190
1200
ATAAATGATTATAAGAGTAAAAAGAAAGTTGCTGATGCTCTCATAACCTTGTATAACCCAAAGGACTAGCTCATGTTGCTAGGCAACTAAACCGCAATAA
1210 1220 1230 1240 1250 1260 1270 1280 1290 1300
CCACATTTGTGACGCGAGTTCCGCATTTGTGACGCGTTAAGTTCCTGTTTTTACAGTATATAAGTGCTTGTATTCTGACAATTGGCACTCAGATTCTGCG
1310 1320 1330 1340 1350 1360
GTCTGAGTCCCTTCTCTGCTGGGCTGAAAAGGCCTTTGTAATAAATATAATCCTCTACTCA
FIG. 1. Nucleotidesequence and deduced amino acidsequenceofcDNA clonep176.Nucleotide positions of the cDNA are shown above thesequence, and thecorrespondingpositions intheEIAVgenomic sequence, indicating exon borders (20), are bracketed. The REV ORF
(nt249to806) (openbox), sequences derived fromalternatephases of Si (nt 249 to 300), SU (nt 301 to 400), and S3 (nt 401 to 806), a potential initiationcodon (nt 312) (boxed), theATMORF(nt355to1056)(shaded), and sequences derived fromalternatephases in SU (nt 355 to 399) and TM(nt 400 to 1056) areindicated. The figure waspreparedwith the aid of the computer program DNAdraw, written by M. Shapiro, National Institutesof Health.
similarto
p2/2
but could not encode a Tat proteinbecause oftheuseofa
splice
site well within S1 (Fig. 2). The first exonof
p20
wasidentical to that ofp176 but was spliced directly to the second codon of S3 (position 7235) (Fig. 2).The sequence ofp2/2 (data not shown)is identical to that
of clone
p105
(33, 35),
which encodes Tat and is predictedtoexpress Revand atruncated TM protein. Clonep176 (Fig. 1
and
2)
containedonly
the ORFs REV and ATM. In theREVORF
(nucleotides
[nt]
249 to 806), there is an AUG codon(residue
22)
in a favorable context (22, 23) for an initiation codon. Iftranslation
initiates here, the product would bepredicted
toconsist of 165 residues (19,760 Da),with its first29
amino
acids derived from env and fused to 136 codons encodedby
S3. The second ORF, ATM (nt 355 to 1056),containsa
potential
AUG initiation codon at position11, butits context is suboptimal (23). If this initiation codon is
functional,
aprotein
of 224 amino acids (24,995 Da) would result. Its N-terminal five amino acids are derived from analternate
phase
of the envsequence and are linked directlytocodon 197 of the TM ORF. Rice et al. (40) found that the EIAV TM
protein
is cleaved at residue 240 into N-terminalglycosylated
and C-terminalnonglycosylated
productsdes-ignated
gp32
andp20,
respectively. These proteins wereEIAVGENOME
LTR ga
Upo pot
S2 LTR
U
p2/2
-evRevA
Tat
---;E1---p
Rv2p20
FIG. 2. Structuresof putative Rev cDNAs. The organization of theEIAVgenome, the surface glycoprotein (SU) and TM coding regionswithinenv, and the exons (black boxes) and predictedORFs
(patterned boxes) of the Rev cDNAs are illustrated. Patterns on boxescorrespondtothepatterns used for the EIAV genomic coding regionsatthetop.The white boxes that start the ATM ORF indicate analternate phase inenv. LTR, long terminal repeat.
F37277777,7777773..,/,
Si EE]
on November 9, 2019 by guest
http://jvi.asm.org/
[image:2.612.68.563.98.412.2] [image:2.612.324.568.511.661.2]10 20 30 40 50 60 70 80 90 100
110
120
130
140
150
160
170
180
190
200CGCCCGAACAGGOACGAGGQAATACCTGATTGAACCTGGCTGATCGTAGGA.LCCCCGGGA.CAGCAGAGGAG
AACTTACAGAAGTCT TCTGGA
I0PR
0S
RG
E L TEVFW1
21
1Ext
2
30
E 2o242250
260
2
70
2
80
2 90
300
GGTGT
TCCTGGC,CA.G.AA
A AG
A.&A,,A.T
TQTAGCTTG
QA.C.C.A.G.TGGT.GCA.G.G.T.CC.T.C.C.G.GCAG.TC.GT.T.A.C.CTGAA.G.A.A.A.AAAT
t (TU flln lAfl
t (S3 codonf 1)
3 10
32
3
30
340
3
50
3
60
3 70
3
80
3 90
40 0410 4
20
430
440
4504860
470
480
490
50 0
[image:3.612.56.551.73.361.2]ACAGAAGTAC
TCCAG.G.A
C
ATGGikA.TGGAG.AAT.CAGA
GGAGTACAACA
GGCG%GC.CA
A.AGA
GCT
GGGTGA.AGT.C.A.A.T.C.G.AGGC.A.T.T.T.GG.A.G.AG.A.GC.T.AT
... ...44 t t t t W V X ...A
510
529
530
540
550
560
570
580
590600
A....C-.A...CCAA Q..
TTT.ACTAG0.G...CAACOACACAAGA....CTCT...06..GA.c....CCT.ACCAAGGGTCCT..
7A~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~...
FRA-~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~... ...
610 egb. 630 640 650 660 670 680 690 700
AC.C.TQGA.GA.T.T. TTTATOIIAC
CO A.
A,2l..QJQ...&&Qj.AGRA.A.
6.A.C.A ... .... .... lix,S. F.
P G D S K R R R K H L
710 72
0
730 740 750 760 770 780 790 800A.T.TT.T.G.GQG.A.C.T.A.G.TA.A,T.T-4.1.A.G.T.A.G.GAC.G.CA.T.A..G.C..A.G.GC.T.A.TGG.A.T.T.A.C.G.T.O.G.A.C.T.C.G.C.T.G.TT.A.T.AA.T.A.AG.G.A.T..T.T.G.TAT.T.AGA.G.GCT.T.AA.A.T.TT.G.A.TA...V... ..
... ... ...a... ...L... ... ... .. ... ... ...v... . V A %: %
810 820 830 840 850 860 870 880 890 900
T.T.TQA.A.A.T.A.A.TCA.G.AA.A.A,... X A-TOCTT.G.A.TT.A.T.AT.T.G.GA.A.G.A.GC.T.TT.A.A.A.T.C.C.TGG.C.A.CA.T.C.T.C.A.T.G..T.A.T.C.A.A.T.GC.C.T..C.A.G.TA..T.G.T.T.TAGAAAAACAAGGGGG... .3L ...:x, ..T. -Xm... y
V--910
920
930
940950
960 970980
9901000
1010
1Q20-
1030
1040
1050
1060
1070
1080 1090 11001110
1.12
1130
1140
11501160
1170
1180TTCTGACAATTGGGCACTCAGATTCTGCGGTCTGAGTCCCTCTCTGCTGGGCTGAAAAGGCCTTTGTAATAAATATAATTCTCTACTCA
FIG. 3. Nucleotide
sequence
and deduced amino acidsequence of cDNA clonep20.
The REV ORF(nt
158to634) (open
box),
sequences derivedfromexon1(nt
158to230)
and S3(nt
231to634),
the ATM ORF(nt
120to884) (shaded),
sequences derived fromexon1(nt
120to230)
and from TM(nt
231 to884),
and the codon used toinitiate the Tatprotein
(nt
159)
(boxed)
areindicated.found in virions.
Whethier
theputative
truncatedenvelope
proteins
encodedby
the. EIAV cDNAs described here have afunctionanalogous
tpthiat
ofp20
remainstobedetermined. A truncatedenvelope
protein
was alsosuggested
to be encodedby
therevtranscripts
of visnavirus(29)
andcaprine
arthritis
encephalitis
viru's
(19).
Itis ofinterest that C-termi-nal sequencespresent -in
EIAV and other lentiviral TMproteins
possessstructuires
similar to those of afamily
ofcytolytic
peptides
andcouldplay
arole incytopathic
effects inducedby
lentiviral-infection
(31).
Clone
p20
ispredicted
tocontaintwoORFs(Fig.
2and3).
The first
ORF,
designgated
ATM(nt
120 to884),
lacks an obvious translation initiationcodon,
but it isintriguing
that its first 38 codons are identical to those of the tat geneproduct
andarespliced,
tocodon 198 of the TM ORF.Thus,
atruncated TMprotein
of
26,576
Da could besynthesized
by
using
the Tatprotein's
initiation
codon(isoleucine,
AUC)
atposition
14(35). Interestingly,
Beisel et al.(3)
detected atranscript analogous
toc~DNA
p20
in horsesacutely
infected with EIAV. The secondORF,
designated
REV,
(nt
158 to634),
starts 66bp
downStream
from the 3' end of thelong
terminalrepeat.
Thisstre'tch,
whichlacks anAUGcodon,
isspliced
tothe second co'don ofS3.
Thus,
theproduct
ofthis ORF wouldpotentially,represent
a form of Rev with an amino terminusdifferentfrom
that encodedby p176
(assum-ing
that initiationoccurs at anon-AUGcodon).
The
coding capacity
of~
each cDNAwas assessedby
the use of acoupled
in vitrotranscription-translation
system.
Each
cDNA,
subclorned
into aBluescript
vector,wastran-scribed in vitro with a
Stratagene
kit,
and theresulting
capped
RNAswere usedtoprogram areticulocyte
transla-tionextractinthe presence of
[35S]methionine
and[35S]cys-teine
(35).
Twoproteins
withapparent
molecularmassesof 20 and 17 kDaweresynthesized
in response to clonep2/2
DNA
(Fig.
4A,
lanea).
The differences in the sizes of the bands(20
and 17 kDa in vitroversus 18 and16 kDa invivo)
are
likely
duetogel
andsample
composition variability.
The 8-kDa Tatprotein
(35)
was notobserved,
presumably
be-cause ofthe inefficient use of non-AUG codons in in vitro translationsystems
(23).
The sameresultwasobtained with clonep176
(Fig.
4A,
laneb).
NocDNA-specific
bandswere observedinextractsprogrammed by
clonep20
(Fig.
4A,
lanec).
This result may also beexplained
by
thelack ofanAUG codon in thepredicted
ORFs ofp20.
To further characterize the
products expressed by
clonep176,
apool
of anti-S3sera(antiserum
177,
whichwasraisedagainst oligopeptide
177[LRQSLPEEKIPSQTY],
and anti-serum181,
raisedagainst oligopeptide
181[DFSAWGGY
QRAQERL])
wasused.Thesametwoproteins
werespecif-ically immunoprecipitated by
anti-S3 sera(Fig.
4A,
lanee),
suggesting
that bothproteins
weretranslated from the REV ORF. Their sizes were most consistent with initiation atpositions
22and 33 of the REV ORF.Among
otherpossi-bilities,
bothmight
start atcodon 22 and the differences inapparent
molecularweights
might
be due either to RNAediting
(18)
ortodifferentialmodificationby
phosphorylation
(9)Next,
weinvestigated
thecoding capacities
of the three cDNAspecies
in aeukaryotic expression
system.
The cDNAs were cloned into amammalianexpression
vector,pMAMneo
(Clontech),
downstream of the dexamethasone-induciblelong
terminalrepeat
of mammary mouse tumoron November 9, 2019 by guest
http://jvi.asm.org/
B
a
b
e
"i
.....:.. ..:. ...
d
<201
4 17
a
b
cd
a18
4 16
C
9
ab
ckmrn
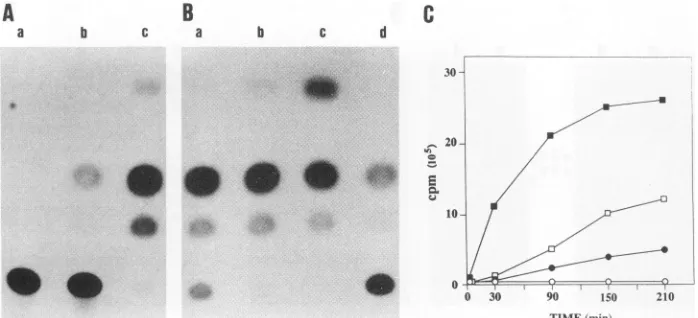
_Ial
FIG. 4. Coding capacity of the Rev transcripts. (A) Analysis of in vitro translation products of cDNAs. [35S]methionine- and [35S]cysteine-labelledpolypeptidesweresynthesizedinanin vitrotranscription-translation reaction mixturecontaining5 ,gof DNA from pKS176 (lane a), pKS2/2 (lane b),orpKS20 (lane c)orfrompKS2/2in theantisenseorientation(lane d),andtheproductswereseparated bysodiumdodecyl sulfate (SDS)-polyacrylamide gel electrophoresis (15%
[wt/vol]
polyacrylamide gel). In asecondgel, products of the reactionprogrammed by pKS176DNAwerepreadsorbed with preimmuneserum(1hat0°C)and thentreated withprotein A-Sepharose(30 minat0WC). Supernatantswerethenimmunoprecipitatedwithapoolofanti-S3sera(no.177and181) (lane e)oranti-S3serapreadsorbedwith theappropriate oligopeptides (lane f).Arrowsindicate thepositions ofthe17- and 20-kDaproducts. (B) Analysis of cDNA expressioninCOS cells. COScellsweretransfected withpMAM/176 (lane a), pMAM/2/2 (lane b), pMAM/2O (lane c),orpMAM (lane d). Cellsweregrownfor 32daysin thepresenceofG418 (900pg/ml) and then treated for48 h withdexamethasone(10-6 M). Lysatesfrom 6x106 cellswereincubatedfirst with preimmune serum (1 h, 0°C) and then with protein A-Sepharose (30 min, 0°C) before immunoprecipitation of the resultant
supernatantwithapoolofanti-S3sera(no. 177 and 181). Inasecond gel, celllysatesofpMAM/2/2-transfectedcellsincubatedinthepresence (laneseandg)orabsence(lane f)ofdexamethasonewereadsorbed withpreimmuneserum asdescribed aboveand thenimmunoprecipitated with anti-S3sera(laneseand f)oranti-S3serapreadsorbed with the appropriate oligopeptides(lane g). Proteinsweredetectedby using the poolofanti-S3seraand
"2I-labelled
proteinA.Arrowheadsindicate thepositionsof the16- and 18-kDaS3-encodedproducts. (C)Analysisof EIAV-infected cells and virions. Totalcelllysatesprepared from5 x 106persistentlyinfected(lane a)oruninfected (lane b)canine cells andSDS-lysed, gradient-purified EIAVvirions(lane c)were treated with preimmune serum,immunoprecipitatedwith anti-S3 sera,and immunoblotted asdescribedabove.
virus. The three cDNA expression vectors pMAM/2/2, pMAM/176, andpMAM/20weretransfected into COScells.
Following 32 days ofgrowth in the presenceof G418 (900 ,ug/ml),cellsweretreated withdexamethasone(10-6M) and
48 hlater, cell extractswere immunoprecipitated and then
probedwith apoolof thetwo S3antisera. Twoproteins of approximately16 and 18kDawereimmunoprecipitated from cells transfected with pMAM/2/2 or pMAM/176 (Fig. 4B, lanes a and b), while only one protein of 16 kDa was
immunoprecipitated from cells transfected with pMAM/20 (lane c). In contrast,noequivalentbandwasobserved with cells transfected with the vectoritself, nor was there any S3-specific signal when the S3 antisera were preadsorbed
with the immunizing peptides. The S3 antisera were next usedtoanalyzeS3-encodedproteinsinEIAV-infected cells.
Figure 4C (lane a) shows that an 18-kDa protein, which comigratedwith the S3-encoded 18-kDaprotein of pMAM/ 2/2 (data not shown),was specifically immunoprecipitated
from extracts of infected cells. Stephens et al. (47) previ-ously reported an S3-encoded 18-kDa protein in EIAV-infectedequine fetalkidney cells.
Since itwasreportedthat the Revproteinof visnavirus is presentinmaturevirions(29),itwasofinteresttodetermine
whetherthiswasalsotruefor EIAV.Supernatantsof canine
cellspersistentlyinfected with EIAVwerecollectedat24-h
intervals.Virionswerepelleted,and theviralproteinswere
immunoprecipitated bythepoolof S3antisera. Aproteinof 18 kDawas specificallydetected in EIAV virions (Fig. 4C,
lanec).The16-kDa band present(Fig. 4C,lanec)islikelyan
artifact,sinceitwasnotdetected whenvirionextractswere first immunoprecipitated and then analyzed by Western
immunoblottingwith the anti-S3 sera. Furthermore, afaint
16-kDabandcanbeseenin thecontrol lanesontheoriginal autoradiogram.
The results of the present study indicate that the three
speciesofcDNAs, p2/2, p176, andp20, expressed proteins
encodedbyS3. Theyallgaverisetoacommonproteinof16 to17kDa, and two,p2/2 andp176, expressedanadditional protein of 18 to 20 kDa. It is noteworthy that the Rev
transcriptof visnavirus alsoencodestwospeciesof Rev-like
proteins (29). On the basis ofsequenceanalysisof the three
transcriptsandtheirpredicted coding capacity,it ispossible
thatthe 18-kDa S3-encoded protein expressed by p2/2 and
p176 is a protein of 165 amino acids whose 29 N-terminal residues are derived from the surface glycoprotein (SU)
sequences(codons43to73)whicharefused in frameto S3.
The structure of this protein is analogous to those of the
predictedRevproteinsof otherungulate lentivirusessuchas visna virus (13, 29, 44), visna-like ovine virus SA-OMVV
(39), bovine immunodeficiency virus (12, 36), and caprine
arthritis encephalitisvirus(19, 43).
Alignmentofthepredicted18-kDaS3-encodedproteinof EIAV with the Rev proteins of other lentiviruses (47) re-vealed the presence of three conserved domains. The first domain contains a cluster of charged amino acids rich in
glutamylresidues(KEESKEEKRRNDWWKK, residues36 to 51 of the REV ORF). Although the N termini of the
primateRevproteinsarenotderived fromenv(6, 7, 16), they contain an analogous cluster ofcharged residues (RSGDS DEDLLKAVRLIK). Recent mutational analysis (17) sug-gestedthat this N-terminal stretchis involved in thebinding specificityofthe Revprotein for the Revresponseelement (RRE). Thesecond domain is also a highly charged region
composedofseveral basicamino acids in the secondcoding
A
on November 9, 2019 by guest
http://jvi.asm.org/
[image:4.612.88.548.71.228.2]A
B
a b c a b c d
0
C
0 30 90 150 210
TIME(min)
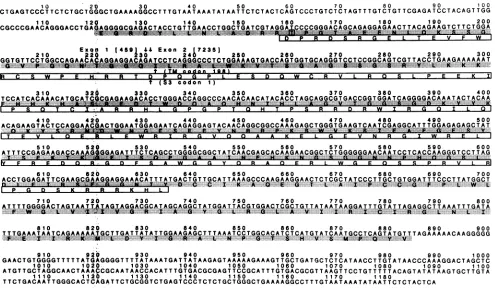
FIG. 5. Assay forRevactivity. The CAT reporterplasmid pCMV-CAT-RREwasconstructedby cloning the CAT coding region (the BglII-to-Bsp 1286 Ifragment whichspansnt-42to689, where+1referstotheAof the CAT initiationcodon) into theBamHI-HindIIIsites downstreamof theCMV promoter inpCEV21(36). Aportion of theenvregion of theEIAV provirus(theBglII-to-ApaI fragment [nt 4473 to7247]), which presumably contains theRRE but lacks thepresumedRevcodingsequence,wascloned into theBglII-ApaIsitesofpCEV21, downstream of the CATregion. Theinsulinpoly(A) signalis located downstream of the insertedenvregion.Asimilar construct,designated pCMV-CAT,whichlacksthe envregion, servedasacontrol.(A) Canine cellsweremock transfected(lane a)ortransfected with4 ,ugof DNA frompCMV-CAT-RRE(laneb)orpCMV-CAT(lanec). After48h, CAT levelswereassessedaspreviously described (45).Percentagesof
[14C]chloramphenicol
convertedto acetylatedderivativeswere asfollows: lane a,0; laneb, 23.0; and lane c, 98.1.(B) Caninecellswereco-transfectedwith 4 ,ugofDNAfrom pCMV-CAT-RREand 4 Lg of DNA from cDNA clone
p20
(lane a), p2/2 (lane b),orpl76
(lane c)orthe pCEV21vector (laned). After48h,CATlevelswereassessedand the percentages of[ C]chloramphenicol converted toacetylated derivativeswere asfollows:lane a,89.3; laneb, 98.4;lane c,99.2; and lane d, 35.9. (C) Duplicate samplesweretransfectedasdescribed for panelsAand B,andCATactivitywasdeterminedby themethod of Neumannetal.(32). Symbols: U,clonep176;El,clonep2/2;*,clone p20; 0,pCEV21.
exon
(RRDRWIRGQ, residues 97
to 105 of the REVORF),
reminiscent of the arginine-rich
motif
of the Revproteins
of primate lentiviruses. These residues arethought
to be in-volved in binding to the RRE(21,
25,27)
and are alsothought
to act asthenuclear localizationsignal
(4, 26, 38, 51,
53).
The third conserveddomain,
located at thecarboxy
terminus of the
putative
S3-encoded Revprotein,
spans residues 129 to 141(ELGEVNRGIWREL)
and may be analogous tothe humanimmunodeficiency
virus type1 Revsequence
(LPPLERLTLDCNED),
whichwassuggested
tofunctionas anactivation domain that bindstocellular factors involved in RNA transport
(24, 26, 37, 50).
The 16-kDa S3-encoded
protein,
which isexpressed
by
all threetranscripts, presumably
initiatesat anon-AUG codon locatedatthe 5' end of S3.Consequently,this
protein would lack thehighly charged
N-terminalglutamic
acid-rich do-main locatedatthe5' end of the 18-kDaprotein.
However, it wouldcontain the othertwoconservedregions
encodedby S3.To further substantiate thenotion that the three
species
of cDNAs representrevtranscripts,
weexamined whether theproteins
expressed by thesecDNAscouldovercomeintransthe effects of env sequences in
inhibiting expression
of achloramphenicol acetyltransferase
(CAT)-env
reporter con-struct(38).
Areporterplasmid
(pCMV-CAT-RRE)
in whichaportion ofenvlacking S3, but presumablycontaining the RRE, located downstream of the CAT coding sequence and under the control of thecytomegalovirus (CMV) promoter,
was constructed. The CAT activity of this construct was
23%
conversion (versus 97.2% for pCMV-CAT) (Fig.5A).
Significantly higher
CAT levels wereobserved in thepres-ence of each one of the rev cDNAs. Clone
p20,
which expresses a 16-kDa S3-encodedprotein,
enhanced CAT production by pCMV-CAT-RRE to a level of76.8%
conver-sion.
Moreover,
cDNA clonesp2/2
andp176,
which express18- and 16-kDa
protein
species, elevated CAT levels to 95.4 and97.9%, respectively.
Using
theFOLD programasimplemented
intheUniver-sity
of Wisconsin GeneticComputing
Center's sequenceanalysis
softwarepackage,
weperformed
exhaustiveanaly-ses of the EIAV env nucleotide sequence in attempts to
identify
apotential
RRE;however,
none was found.Thus,
the EIAVRRE
might
havea much more subtlesecondary
structure
than
thoseidentified in other lentiviruses or mayuse an
altogether
different sequence motif. In any case, it will be necessarytoidentify
the EIAV RREby recombinanttechniques and/or
direct RNAbinding experiments.
It will also be of interest to determine whether the EIAV Revprotein
canrecognizethe RREs of otherlentiviral
genomes. Our data indicate that the Revproteins
of EIAV areexpressed
viaalternatively spliced
mono-andpolycistronic
messages, asis thecasefor otherprimate
(2,
15, 41, 49,52)
andungulate
(10,
19,29, 36, 43)
lentiviruses. However, thepolycistronic
messagesofEIAV may expresstwospecies
of Revwith distinct N termini. In other systems, it has been shown that the use of alternativeinitiation
codons from within thesamemessageresults in differentialprotein
local-ization(1,
5,46).
It remains to bedetermined whether thetwo
putative
Revproteins
arelocalizeddifferently.
The Revprotein
of visna virus(19 kDa),
in contrast toprimate
lentiviral Rev
proteins,
accumulatedpreferentially
innuclei(48)
aswellasinthecytoplasmic
and membrane fractions of infected cells(28).
Moreover, the largestspecies
of the Rev protein of visna virus (19 kDa) was packaged into matureviral
particles,
as is shown here for EIAV. Thus, the Revprotein might
playarole in theassembly of viral particlesorin the
early
stepsofvirusinfection,
in additiontoregulating
the
cytoplasmic
expression ofunspliced
orsingly
spliced
viral
transcripts.
Nucleotide sequence accession numbers. The nucleotide
;VN-,El 2
?n ) # lp
.i-;%', r", E
u
4b 4'o
on November 9, 2019 by guest
http://jvi.asm.org/
[image:5.612.129.478.72.231.2]sequences
and deduced amino acid sequences of cDNAclones
p176
andp20
have beenlodged in the GenBank database under accession numbers X63059 and X63058,
respec-tively.
We thankS. A. Aaronson for continued support and
encourage-ment.
This studywas supported by a project grant from the United
States-Israel Binational ScienceFoundation. REFERENCES
1. Acland,P.,M.Dixon, G.Peters, and C. Dickson. 1990. Subcel-lular fate ofthe Int-2 oncoprotein is determined by choice of initiation codon.Nature (London) 343:662-665.
2. Arrigo,S.J.,S.Weitsman, J.A.Zack, and I. S. Y. Chen. 1990. Characterization and expression of novel singly spliced RNA species of human immunodeficiency virus type 1. J. Virol. 64:4585-4588.
3. Beisel, C.E., J.F.Edwards, L. L.Dunn, and N. R. Rice. 1993.
Analysis
ofmultiplemRNAs frompathogenic equineinfectious anemia virus(EIAV)inanacutely infected horsereveals a novelprotein,
Ttm, derived from thecarboxyterminus of the EIAV transmembraneprotein. J.Virol.67:832-842.4. Berger, S., C. Aepinus, M. Dobrovnik, B. Fleckenstein, J. Hauber, and E. Bohnlein. 1991. Mutational analysis of func-tional domains in the HIV-1 Rev trans-regulatory protein.
Virology
183:630-635.5. Bugler, B.,F.Amalric,and H. Prats. 1991.Alternativeinitiation oftranslation determinescytoplasmic ornuclear localization of basicfibroblastgrowth factor. Mol. Cell. Biol. 11:573-577. 6. Chakrabarti, L.,M. Guyader, M.Alizon, M. D. Daniel, R. C.
Desrosiers,P.Tiollais, and P.Sonigo. 1987. Sequence of simian immunodeficiencyvirus from macaque and its relationship to otherhuman and simian retroviruses. Nature(London) 328:543-547.
7. Colombini, S., S. K. Arya, M. S. Reitz, L. Jagodzinski, B. Beaver,and F.Wong-Staal. 1989. Structureofsimian
immuno-deficiency
virus regulatorygenes. Proc. Natl. Acad. Sci. USA 86:4813-4817.8. Cullen, B. R. 1992.Mechanism of action of regulatory proteins encodedby complexretroviruses. Microbiol. Rev. 56:375-394. 9. Curran, T.,M. B.Gordon,K. L.Rubino, and L. C. Sambucetti. 1987.Isolationandcharacterization ofthec-fos (rat) cDNA and
analysis
of post-translational modification in vitro. Oncogene 2:79-84.10.
Davis,
J. L., andJ. E. Clements. 1989. Characterization of a cDNA clone encoding the visna virus transactivating protein. Proc. Natl.Acad.Sci. USA86:414-418.11. Dorn, P.,L.DaSilva,L.Martarano, and D. Derse. 1990. Equine infectiousanemia virustat:insights intothe structure, function, and evolution of lentivirus trans-activator proteins. J. Virol. 64:1616-1624.
12. Garvey,K.J.,M.S.Oberste, J.E. Elser, M. J.Braun, and M. Gonda. 1990.Nucleotidesequenceand genome organization of
biologically
activeprovirusesof thebovine immunodeficiency-like virus.Virology 175:391-409.13. Gourdou, I.,V. Mazarin, G. Quert, N. Sauze, and R. Vigne. 1989. The open reading frame S ofvisna virus genome is a trans-activatinggene.Virology 171:170-178.
14. Greene,W.C. 1991. The molecularbiologyof human
immuno-deficiency
virus type 1infection.N.Engl.J. Med.324:308-317. 15. Guatelli, J. C., T. R. Gingeras, and D. D. Richman. 1990. Alternativespliceacceptorutilization duringhumanimmunode-ficiency
virus type 1 infection of cultured cells. J. Virol. 64:4093-4098.16. Guyader, M.,M.Emerman,P.Sonigo,F.Clavel, L.Montagnier, andM. Alizon. 1987. Genomeorganizationandtransactivation ofthe humanimmunodeficiencyvirus type 2. Nature (London) 326:662-669.
17. Hope,T.J.,D.McDonald,X.Huang, J. Low, and T. G. Parslow. 1990.Mutationalanalysisof the humanimmunodeficiencyvirus type 1 Rev transactivator: essential residues near the amino terminus. J. Virol. 64:5360-5366.
18. Horikami, S. M., and S. A. Moyer. 1991. Synthesis of leader RNA and editing of the P mRNA during transcription by
purified measlesvirus. J.Virol. 65:5342-5347.
19. Kalinski,H.,A.Yaniv,P.Mashiah,T.Miki, S. R.Tronick,and A. Gazit. 1991. Rev transcripts ofcaprine arthritis encephalitis virus. Virology 183:786-792.
20. Kawakami, T.,L.Sherman, J.E.Dahlberg,A.Gazit,A.Yaniv, S. R.Tronick, and S. A. Aaronson. 1987. Nucleotide sequence analysis of the integrated form of the genome ofequine infec-tiousanemiavirus. Virology 158:300-312.
21. Kjems, J., A. D. Frankel, and P. A. Sharp. 1991. Specific
regulation ofmRNAsplicingin vitroby apeptide from HIV-1 Rev. Cell 67:169-178.
22. Kozak, M. 1987. Effects of intercistronic lengthon theefficiency
of reinitiationby eucaryotic ribosomes. Mol. Cell. Biol. 7:3438-3445.
23. Kozak, M. 1989. Context effects and inefficient initiation at non-AUG codons in eucaryotic cell-free translation systems. Mol. Cell. Biol. 9:5073-5080.
24. Malim, M. H., S. Bohnlein, J. Hauber, and B.R. Cullen. 1989. Functional dissection of the HIV-1 Revtrans-activator deriva-tion ofatrans-dominant repressor of Rev function. Cell 58:205-214.
25. Malim, M. H., J. Hauber, S.-Y.Le, J. V. Maizel, and B. R. Cullen. 1989. The HIV-1 rev trans-activator acts through a structural target sequence to activate nuclear export of
un-spliced viralmRNA. Nature (London)338:254-257.
26. Malim, M. H., D. F. McCarn, L. S. Tiley, and B. R. Cullen. 1991. Mutational definition of the human immunodeficiency virus type 1 Rev activation domain. J. Virol. 65:4248-4254. 27. Malim, M. H., L. S. Tiley, D. F. McCarn, J. R. Rusche, J.
Hauber, and B. R. Cullen. 1990. HIV-1 structural gene expres-sion requires binding of the Rev trans-activator to its RNA target sequence. Cell 60:675-683.
28. Mazarin, V., I. Gourdou, G. Querat, N. Sauze, G.Audoly, C. Vitu, P. Russo, C. Rousselot, P. Filippi, and R. Vigne. 1990. Subcellular localization of rev-gene product in visna virus-infected cells. Virology 178:305-310.
29. Mazarin, V., I. Gourdou, G.
Querat,
N. Sauze, andR. Vigne. 1988. Genetic structure and function of an early transcript of visna virus. J. Virol. 62:4813-4818.30. Mild, T.,T.Matsui, M. A. Heidaran, and S. A. Aaronson. 1989. An efficient directional cloning system to construct cDNA libraries containing full-length inserts at high frequency. Gene 83:137-146.
31. Miller, M. A., R. F.
Garry,
J. M. Jaynes, and R. C. Montelaro. 1991. A structural correlation between lentivirus transmem-brane proteinsandnatural cytolytic peptides. AIDS Res.Hum. Retroviruses 7:511-519.32. Neumann, J. R., C. A. Morency, and K. 0. Russian. 1987. A novel rapid assay for chloramphenicol acetyltransferase gene expression. BioTechniques 5:444-447.
33. Noiman, S., A. Gazit, 0. Tori, L. Sherman, T. Miki, S. R. Tronick, and A.Yaniv. 1990. Identification of sequences encod-ing the equine infectious anemia virus tat gene. Virology 176: 280-288.
34. Noiman, S., A. Yaniv, L. Sherman, S. R. Tronick, and A. Gazit. 1990.Pattern of transcription of the genome of equine infectious anemia virus. J. Virol. 64:1839-1843.
35. Noiman, S., A. Yaniv, T. Tsach, T.Miki,S. R. Tronick, and A. Gazit. 1991. The Tat protein of equine infectious anemia virus is encoded by at least three types of transcripts. Virology 184:521-530.
36. Oberste, M. S., J. D. Greenwood, and M. A. Gonda. 1991. Analysis of the transcriptionpattern and mapping of the puta-tive rev and env splice junctions of bovine immunodeficiency-like virus. J. Virol. 65:3932-3937.
37. Olsen, H. S., P. Nelbock, A. W. Cochrane, and C. A. Rosen. 1990.Secondary structure is the major determinant for interac-tion of HIV rev protein with RNA. Science 247:845-848. 38. Perkins, A., A. W. Cochrane, S. M. Ruben, and C. A. Rosen.
1989. Structural and functional characterization of the human immunodeficiency virus Rev protein. J. Acquired Immune
on November 9, 2019 by guest
http://jvi.asm.org/
fic. Syndr.2:256-263.
39. Querat, G., G.Audoly,P. Sonigo, and R. Vigne. 1990. Nucle-otide sequence
analysis
of SA-OMVV, a visna-related ovine lentivirus: phylogenetic history of lentiviruses. Virology 175: 434 447.40. Rice, N. R., L. E. Henderson, R. C. Sowder, T. D. Copeland, S. Oroszlan, and J. F. Edwards.1990. Synthesis andprocessing of the transmembrane envelope protein of equineinfectious ane-mia virus.J.Virol. 64:3770-3778.
41. Robert-Guroff, M., M.Popovic,S. Gartner, P.Markham, R. C. Gallo,and M.S. Reitz.1990. Structureandexpression oftat-, rev-, andnef-specific transcripts of human immunodeficiency virustype1in infectedlymphocytes andmacrophages. J. Virol. 64:3391-3398.
42. Rushlow, K., K. Olsen, G. Stiegler, S. L. Oayne, R. C. Mon-telaro,andC. J. Issel. 1986. Lentivirusgenomic organization: the complete nucleotide sequence of the env gene region of equineinfectious anemiavirus. Virology 155:309-321. 43. Saltarelli, M., G. Querat, D. A. M. Konings, R. Vigne, and J. E.
Clements. 1990.Nucleotidesequenceandtranscriptional analy-sis of molecular clones of CAEVwhich generate infectious virus.Virology179:347-364.
44. Sargan,D.R., andI.D. Bennet.1989. Atranscriptionalmapof visnavirus: definition ofthesecond intronstructuresuggestsa
rev-likegeneproduct. J. Gen.Virol. 70:1995-2006.
45. Sherman, L., A.Gazit,A.Yaniv,T.Kawakami, J.E.Dahlberg, andS.R. Tronick. 1988. Localization ofsequencesresponsible
for trans-activation of theequine infectious anemia virus long
terminalrepeat.J.Virol. 62:120-126.
46. Spence, A. M.,P. C. Sheppard, J. R.Davie, Y. Matou, N. Nishi, W. L. McKeenan, J. G. Dodd, and R. J. Matusik. 1989. Regulation of a bifunctional mRNA results in synthesis of secreted and nuclear probasin. Proc. Natl. Acad. Sci. USA 86:7843-7847.
47. Stephens,R.M., D. Derse, and N. R. Rice. 1990. Cloning and characterization ofcDNAs encoding equineinfectiousanemia virus Tat and putative Revproteins. J. Virol.64:3716-3725. 48. Tiley, L.S., P. H. Brown,S.-Y. Le, J. V. Maizel, J. E. Clements,
and B. R. Cullen. 1990. Visna virus encodesa post-transcrip-tional regulator of viral structuralgene expression.Proc.Natl. Acad.Sci.USA87:7497-7501.
49. Unger, R. E., M. W. Stout, and P. A. Luciw. 1991. Simian immunodeficiency virus(SIVmac) exhibits complex splicing for tat, revandenvmRNA.Virology 182:177-185.
50. Venkatesh, L. K., and G. Chinnadura. 1990. Mutants in a conserved region near the carboxy-terminus of HIV-1 Rev identify functionally important residues and exhibitadominant negative phenotype. Virology 178:327-330.
51. Venkatesh, L. K., S. Mohammad, and G. Chinnadura. 1990. Functional domains oftheHIV-1 rev gene required for trans-regulation and subcellular localization. Virology 176:39-47. 52. Viglianti,G.A., P. L.Sharma,and J. I. Mullins.1990.Simian
immunodeficiency virus displays complex patterns of RNA splicing.J. Virol. 64:4207-4216.
53. Zapp, M. L., and M. R. Green.1989. SequencespecificRNA binding by the HIV-1 Revprotein. Nature(London) 342:714-716.

![FIG. 4.with32firstbypKS176reactionminthecells.poolofsupernatant(lanes[35S]cysteine-labelledandimmunoblotted EIAV-infected days Coding capacity of the Rev transcripts](https://thumb-us.123doks.com/thumbv2/123dok_us/1299207.83001/4.612.88.548.71.228/firstbypks-reactionminthecells-poolofsupernatant-cysteine-labelledandimmunoblotted-infected-capacity-transcripts.webp)