0022-538X/96/$04.0010
Copyrightq1996, American Society for Microbiology
Mutations of Basic Amino Acids of NCp7 of Human
Immunodeficiency Virus Type 1 Affect
RNA Binding In Vitro
EDITH SCHMALZBAUER,† BETTINA STRACK, JENS DANNULL,‡
STEPHAN GUEHMANN,†ANDKARIN MOELLING*
Institut fu¨r Medizinische Virologie, Universita¨t Zu¨rich,
CH-8028 Zu¨rich, Switzerland
Received 22 May 1995/Accepted 18 October 1995
The nucleocapsid (NC) protein of human immunodeficiency virus type 1 is required for packaging of viral RNA and for virion assembly. It contains two clusters of basic amino acids, consisting of five and four amino acid residues, flanking the first of its two zinc fingers. These amino acid residues have been mutagenized to neutral ones individually, as well as in various combinations, by site-directed mutagenesis. Wild-type NCp7 and the mutant proteins were expressed as recombinant proteins inEscherichia coli, with six histidines as tags at their amino termini in order to allow efficient purification. The purified proteins were analyzed for RNA binding in vitro with human immunodeficiency virus type 1 5* leader RNA transcribed in vitro. Assays comprised Northwestern blots at various salt concentrations and filter binding tests which allowed determi-nation of the dissociation constants of the various mutants. The results indicated that mutations of the amino acid R-7 and of R-32 and K-33 were more critical for RNA binding than other mutations. Mutation of the other amino acid residues reduced the binding affinity in proportion to the number of mutations. Mutation of seven of the nine basic amino acid residues reduced the binding of RNA by 50- to 90-fold.
Retroviruses code for nucleocapsid (NC) proteins, which are essential for various steps during retroviral replication. The NC proteins of retroviruses select the specific viral RNA from the pool of cellular RNAs and facilitate dimerization and pack-aging of two RNA genomes into the maturing virion. Further-more, NC proteins enhance binding of the tRNA primer to the primer binding site of the viral RNA (2, 28). NC proteins also stimulate the reverse transcriptase (41), improve the efficiency of the integrase (19), and protect the RNA against nucleases (18). The avian myeloblastosis virus NC protein has been shown to melt RNA of the same virus, leading to hyperchro-micity (39). The NC protein of human immunodeficiency virus type 1 (HIV-1) is, like that of avian and leukemia retroviruses, a structural component of the viral core and is essential for viral replication (1, 21, 27). Recently, the NC protein of HIV-1 has been shown to enhance the catalytic rate of a hammerhead ribozyme in vitro (24, 40). The NCp7 protein of HIV-1 also binds to single-stranded DNA in vitro (18, 38).
The NCp7 protein of HIV-1 is a polypeptide of 55 amino acid residues which arises by proteolytic processing from the polyprotein Gag precursor, which gives rise to a p15 protein from which p7, p1, and p6 are produced by further processing (13, 14). The NCp7 protein contains two zinc finger structures; the first one is more highly conserved than the second one. The first zinc finger is flanked by two basic domains containing five and four basic amino acid residues. The effect of zinc on the conformation has recently been demonstrated by nuclear mag-netic resonance (23, 34, 35, 37).
Most functions of the NC protein involve nonspecific RNA interaction and do not require the presence of zinc fingers (9, 28). However, selection of virus-specific RNA containing the packaging site PSI from the pool of cellular RNAs has been shown to require an intact first zinc finger in vivo (10, 12, 21) and in vitro (8). The HIV Gag or NC proteins have been shown in gel shift studies to bind to PSI-containing RNA indepen-dently of the presence of the zinc fingers (3). The NC protein exhibits two clusters of basic amino acid residues which are characteristic for several RNA-binding proteins, such as Tat or Rev (20). In the case of NC proteins, this cluster is bipartite, flanking the first zinc finger. Deletions of short sequences con-taining these basic residues lead to a complete loss of nucleic acid binding activity in vitro (9). Furthermore, spumaviruses, retrotransposons, and copia-like retroelements contain basic amino acids but no zinc finger, supporting the importance of these sequences for RNA binding (6). This study addresses questions concerning the function of the individual basic amino acid residues in RNA binding in vitro.
MATERIALS AND METHODS
Cloning, expression, and purification of recombinant NCp7 proteins.The gene coding for NCp7 was amplified by standard PCR techniques from the pBH10 template (30). NCp7 was cloned via a PCR primer introduced by NcoI and BamHI restriction sites into a modified pET3c vector (32), which allows expression of recombinant histidine fusion proteins carrying a Met-(His)6-Ala
tag amino terminally. The PCR-derived sequences were confirmed by dide-oxynucleotide sequencing (33) with the Sequenase version 2.0 kit from U.S. Biochemical Corp., Cleveland, Ohio. The pET3c6HisNCp7 recombinant plasmid served as a template for PCR mutagenesis, resulting in the formation of mutants
mt1 through mt8 and mt15, containing mutations amino terminal to the first zinc
finger, or mutants mt11 and mt12, carrying mutations carboxy terminal to the first zinc finger. Mutants mt11 and mt12 were used as templates in a second round of PCR mutagenesis to produce mutants mt9, mt10, mt13, mt14, and mt16, which exhibit mutations in both basic regions flanking the first zinc finger of the NCp7 protein. PCR was performed in a BIOMED thermocycler with Taq polymerase (Promega, Madison, Wis.) according to the instructions of the manufacturer. For site-directed mutagenesis, the following oligonucleotides were synthesized in an Applied Biosystems DNA synthesizer (underlining indicates the mutations):
* Corresponding author. Mailing address: Institut fu¨r Medizinische Virologie, Universita¨t Zu¨rich, Gloriastrasse 30, CH-8028 Zu¨rich, Swit-zerland. Phone: 41-1-257-2652. Fax: 41-1-257-6967.
† Present address: Max-Planck-Institut fu¨r Molekulare Genetik, D-14195 Berlin, Federal Republic of Germany.
‡ Present address: Robert Koch-Institut, D-13353 Berlin, Federal Republic of Germany.
771
on November 9, 2019 by guest
http://jvi.asm.org/
Arg-33Asn, 59
-GGAGCCATGGCGATGCAGAATGGCAATTTTAGGAAC-39; Arg-73Asn, 59-GGAGCCATGGCGATGCAGAGAGGCAATTTTAATA
AC-39; Arg-3–Arg-7–Arg-103Asn-Asn-Asn, 59GGAGCCATGGCGATGCAG AATGGCAATTTTAATAACCAAAATAAGATGGTT-39; Arg-103Asn, 59-G GAGCCATGGCGATGCAGAGAGGCAATTTTAGGAACCAAAATAA GATGGTT-39; Lys-113Gln, 59-GGAGCCATGCGCATGCAGAGAGGCAA TTTTAGGAACCAAAGACAGATGGTT-39; deletion of Arg-3 to Arg-10, 59 -GGAGCCATGGCGATGCAGAAGATGGTTAAGTGT-39; Lys-143Gln, 59 -G -G A -G C C A T -G -G C -G A T -G C A -G A -G A -G -G C A A T T T T A -G -G A A C C A A A GACAGATGGTTCAGTGTTTC-39; Arg-29–Arg-32–Lys-33–Lys-343 Ser-Ser-Asn-Asn, 59-AATTGCAGTGCCCCTAGTAACAACGGCTGT-39; Arg-32– Lys-333Ser-Asn, 59-AATTGCAGGGCCCCTAGTAACAAGGGCTGT-39; Arg-3–Arg-7–Arg-10–Lys-11–Lys-143Asn-Asn-Asn-Gln-Gln, 59-GGAGCCAT GGCGATGCAGAATGGCAATTTTAATAACCAAAATCAGATGGTT CAGTGTTTC-39; Lys-11–Lys-143Gln-Gln, 59-GGAGCCATGGCGATGCA
GAGAGGCAATTTTAGGAACCAAAGACAGATGGTTCAGTGTTTC-39.
The resulting cDNA clones were sequenced across the PCR fragments. Ex-pression of recombinant proteins following transformation of Escherichia coli BL21(DE3)pLysS was induced as described previously (36). Purification was performed by nickel (Ni)-chelating column chromatography (16); proteins were stored in 8 M urea–10 mMb-mercaptoethanol, pH 4.5, and prior to use they were dialyzed against a solution containing 50 mM Tris-HCl (pH 8.0), 50 mM NaCl, and 10mM ZnCl2.
In vitro synthesis and labelling of HIV-1 RNA.For in vitro transcription of the HIV-1 59leader RNA (wild-type [wt] RNA containing the packaging signal PSI, orCRNA), the pGEM4Z plasmid (Promega) containing a 431-bp HindIII-AccI fragment of the HIV-1 clone D10.3 was used as the template (8). Radioactively labelled RNA was transcribed in vitro with 50 U of T7 RNA polymerase (Strat-agene) in 25-ml volumes for 60 min at 378C. Reaction mixtures contained 2mg of linear plasmid DNA; 40 mM Tris-HCl, pH 8.0; 8 mM MgCl2; 2 mM spermidine;
500mM (each) rATP, rGTP, and rCTP; 100 mM rUTP; 50mCi (corresponding to 125 pmol) of [a-32P]UTP; 15 mM dithiothreitol; and 40 U of the human
placental RNase inhibitor RNasin (Promega). The reaction products were treated for 30 min at 378C with 1 U of DNase I to remove the template, extracted twice with a mixture of phenol and chloroform and once with chloroform, and precipitated twice in ethanol. As measured by scintillation counting, the specific activity of the in vitro-transcribed RNA was 63105cpm/mg. RNA with theC
deletion (positions 299 to 307) was obtained by recombinant PCR (8). Northwestern blot.To prepare bacterial extracts, pelleted cells from a stan-dard induction were heated to 1008C in sodium dodecyl sulfate (SDS) loading buffer; the lysates or the purified proteins were separated on SDS–15% poly-acrylamide gels and transferred by electroblotting to a nitrocellulose membrane. Simultaneously an identical polyacrylamide gel was stained with Coomassie bril-liant blue to verify the equality of protein amounts. The nitrocellulose mem-branes were incubated for at least 12 h at 48C in standard Northwestern buffer (30 mM HEPES [N-2-hydroxyethylpiperazine-N9-2-ethanesulfonic acid]-KOH [pH 7.5], 400 mM KCl, 10mM ZnCl2, 2 mM dithiothreitol, 0.5 mg of heparin per
ml) or in Northwestern buffer with modified salt and heparin concentrations as indicated. RNasin was added to inhibit degradation of RNA by RNase. The nitrocellulose membrane was incubated at 228C for 1 h in standard Northwestern buffer containing 33106cpm of radioactively labelled RNA with a specific
activity of 63105
cpm/mg. The membrane was washed twice at room temper-ature in Northwestern buffer containing heparin and twice in the buffer lacking heparin. The membrane was dried and subjected to autoradiography. Binding of the radioactive RNA probe was determined by phosphorimaging.
Filter binding assay.The nitrocellulose filter binding assays were derived from the methods of Riggs and Bourgeois (31) and Daly et al. (7). Prior to use, proteins were dialyzed against a solution containing 50 mM Tris-HCl (pH 8.0), 50 mM NaCl, and 10mM ZnCl2. RNasin (10 U) was added to all samples to
inhibit degradation of RNA by RNase. Proteins and RNA (23104
cpm, with a specific activity of 63105
cpm/mg) were incubated for 15 min at room temper-ature in a solution containing 50 mM Tris-HCl (pH 8.0), 50 mM NaCl, and 10
mM ZnCl2in a final volume of 120ml. After filtration through 25-mm-diameter,
0.45-mm-pore-size nitrocellulose disks (Millipore), the filters were washed twice with 1 ml of reaction buffer and dried. The radioactivity retained on the filters was determined by scintillation counting.
RESULTS
Cloning, expression, and purification of NCp7 protein.The NCp7 gene of HIV-1 was cloned into the pET3c vector and expressed as a recombinant protein with a fusion of six
histi-dines [(His)6] at the amino terminus. In order to analyze the
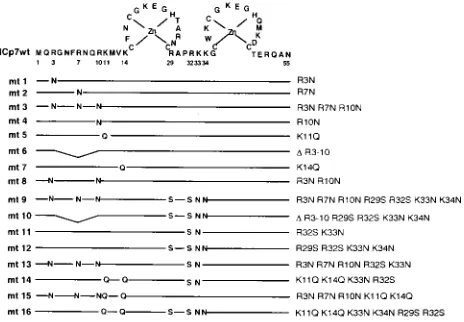
RNA binding properties of NCp7 and to evaluate the role of basic amino acids in binding, site-directed mutagenesis of in-dividual basic amino acid residues or various combinations thereof was performed, resulting in a total of 16 mutants. The mutations were conceived to evaluate the contributions of each of the two basic regions to RNA binding in vitro. Figure 1
shows the amino acid sequence of NCp7, consisting of 55 amino acids, with the two zinc fingers indicated schematically. Basic amino acid residues were mutated to neutral ones as indicated; the neutral amino acids were selected in such a way
that thea-helical angle of the NCp7 structure was not affected
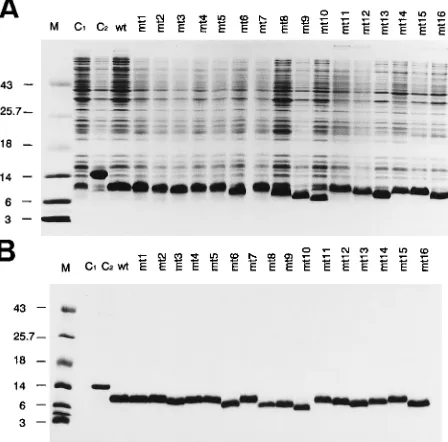
(38). The resulting mutants are numbered mt1 through mt16. Transformation of the wt and mutant (mt) clones into E. coli led to overexpression of the NC proteins. The bacterial ex-tracts were separated by SDS-polyacrylamide gel electrophore-sis, and the proteins were stained with Coomassie brilliant blue
(Fig. 2A). The (His)6tag allows purification of the (His)6-NC
fusion proteins by Ni-chelating column chromatography. Fig-ure 2B shows the purified and dialyzed proteins after SDS-polyacrylamide gel electrophoresis and staining with Coom-assie brilliant blue. The electrophoretic mobilities of some of the mutants were increased (e.g., mt6 and mt16).
RNA-NC protein interaction tested by Northwestern blot analysis. A subset of mutants were analyzed for their RNA binding abilities with bacterial lysates containing overex-pressed wt and mt NC protein (as shown in Fig. 2A) in a Northwestern blot employing in vitro-transcribed RNA. The
RNA corresponds to the 59end of the HIV-1 RNA
(nucleoti-des 76 to 508) of the HIV-1 clone D10.3. The RNA was
uniformly labelled by incorporation of [a-32P]UTP. In order to
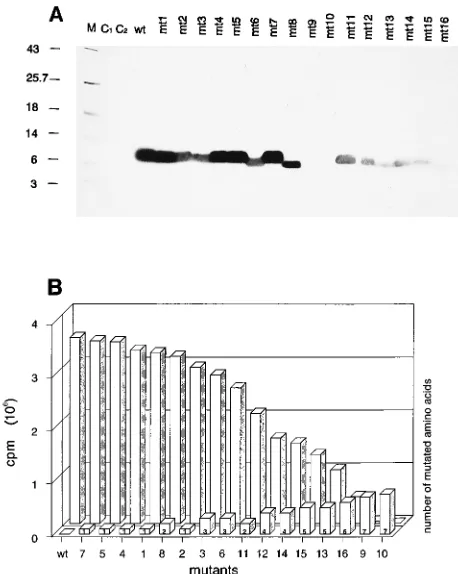
allow discrimination between high- and low-affinity RNA bind-ing of the NC mt proteins, the Northwestern blots were per-formed at various stringencies, with 100, 200, 400, and 600 mM KCl in the buffer during incubation with the RNA (Fig. 3A, B, C, and D, respectively). The results of the Northwestern blot analysis were quantitated by phosphorimaging and are shown as a block diagram in Fig. 3E.
Under the most stringent reaction conditions (600 mM KCl), the binding of mt2, mt3, and mt6 to RNA was substan-tially reduced compared with the binding of the wt protein. Whereas in the case of mt3 and mt6, three of four basic amino
acids 59 to the first zinc finger (R-3, R-7, and R-10) were
affected by the mutation, only R-7 was exchanged in the case of mt2. Since RNA binding of these mutants was reduced to a similar extent, the arginine at position 7 turned out to be of particular importance. Single mutations in the basic amino acids R-3, R-10, K-11, and K-14 (resulting in mutants mt1, mt4, mt5, and mt7, respectively) did not lead to significant
differ-FIG. 1. Amino acid sequence of the NCp7 protein of HIV-1 in single-letter code. Note the two zinc fingers, indicated schematically, wherein C, C, H, and C are coordinated by Zn. The nomenclature of the mutations comprises either the original amino acid residue, its position, and the substituting residue or a dele-tion (D).
on November 9, 2019 by guest
http://jvi.asm.org/
[image:2.612.320.552.73.234.2]ences in RNA binding compared with the wt protein under the conditions used.
In order to analyze the contribution of specific recognition
of the HIV-1 59 leader RNA by the NCp7 wt protein, we
performed Northwestern blot analysis with wt RNA containing
the packaging signal PSI (CRNA) and with a deletion mutant
(CRNAD299–307) as the control (Fig. 4). Deletion of
nucle-otides 299 to 307 resulted in a loss of specific binding of RNA by the nucleocapsid protein, as we demonstrated recently with competitive filter binding assays (8). Under standard assay conditions for Northwestern blot analysis (see Materials and
Methods), binding of theCRNAD299–307 deletion mutant to
the NCp7 wt protein was reduced to about 40% of theCRNA
binding. On the other hand, no differences in binding of NCp7 to RNA containing or lacking the packaging signal could be detected with filter binding assays unless a nonspecific
com-petitor such as tRNA orCRNA antisense was added (data not
shown). These results indicate that specific RNA binding con-tributes differently to the binding efficiencies measured by Northwestern blot analysis and filter binding assays.
Expression of the mutants as (His)6-tagged fusion proteins
allowed their purification to homogeneity by Ni-chelating col-umn chromatography, as shown in Fig. 2B. The purified pro-teins were analyzed for RNA binding by Northwestern blot analysis at 400 mM salt because 600 mM KCl abolished RNA binding of mt10 through mt16. The results of RNA binding with matched amounts of the purified proteins (shown in Fig. 2B) are presented in Fig. 5A. Quantitation of RNA binding activity is shown in Fig. 5B. mt2, mt3, and mt6 show reduced RNA binding, again providing evidence of the importance of R-7. mt5, and mt7 bind with efficiencies similar to that of the wt, indicating that single mutations of K-11 and K-14 are of less
importance than a mutation of R-7. Even the presence of two mutations, as in mt8 (R-3 and R-10), has less of an effect on RNA binding affinity than a single mutation affecting R-7 (mt2). Furthermore, mt11 demonstrates the relevance of R-32 or K-33, since mutation of both amino acid residues reduces RNA binding more than the simultaneous mutation of three basic amino acid residues amino terminal to the first zinc finger (mt3). Additional mutations of R-29 and K-34 (mt12) did not cause a further reduction of RNA binding. R-32 has been shown to be of importance for specific binding of the NC protein to the packaging site PSI (8). When significant num-bers of mutations took place in both domains simultaneously, the RNA binding ability was completely lost (mt9 and mt10).
[image:3.612.63.287.74.295.2]Filter binding assays.In order to quantitate the interaction of the wt and mt NC proteins with RNA, filter binding assays were performed with the purified proteins. Proteins were in-cubated with radioactively labelled RNA in solution and sub-sequently filtered through nitrocellulose filters. The RNA bound by the NC proteins was retained on the filter, and the radioactivity was determined by liquid scintillation counting.
FIG. 2. Expression and purification of recombinant wt and mt NCp7 pro-teins. (A) E. coli bacteria transformed with the pET3c6HisNCp7 plasmid were induced with isopropyl-b-D-thiogalactopyranoside for protein expression.
Pro-teins of total bacterial lysates were separated by SDS gel electrophoresis on 15% polyacrylamide gels and stained with Coomassie brilliant blue. The mutant num-ber is on top of each lane (for nomenclature, see Fig. 1). C1represents a control
lysate of bacteria transfected with the parental plasmid (pET8c); C2represents
a control lysate containing a deletion mutant of c-Myb. M, molecular weight marker. (B) Same as panel A except that the wt and mt NC proteins were purified by Ni-chelating column chromatography and directly applied to the gel.
FIG. 3. Optimization of assay conditions for Northwestern blot analysis in-vestigating NC protein-RNA interaction. Proteins of bacterial lysates separated by SDS-polyacrylamide gel electrophoresis, shown in Fig. 2A, were blotted onto nitrocellulose membranes. The membranes were probed with radioactively la-belled HIV-1 59 leader RNA (5mg), which was transcribed in vitro from a linearized recombinant pGEM4Z plasmid (63105
cpm/mg of RNA) (for details see Materials and Methods). RNA binding was performed under standard assay conditions, except that the concentrations of salt (KCl) and heparin were mod-ified as follows. (A) 100 mM KCl, 0.5 mg of heparin per ml; (B) 200 mM KCl, 0.5 mg of heparin per ml; (C) 400 mM KCl, 1 mg of heparin per ml; (D) 600 mM KCl, 1 mg of heparin per ml. mt1 through mt7 were analyzed; see the legend to Fig. 1 for nomenclature. For definitions of M, C1, and C2, see the legend to Fig.
2. (E) The amount of radioactively labelled RNA bound by the NC proteins was determined by phosphorimaging, and the mean values calculated from different measurements were plotted as column diagrams.
on November 9, 2019 by guest
http://jvi.asm.org/
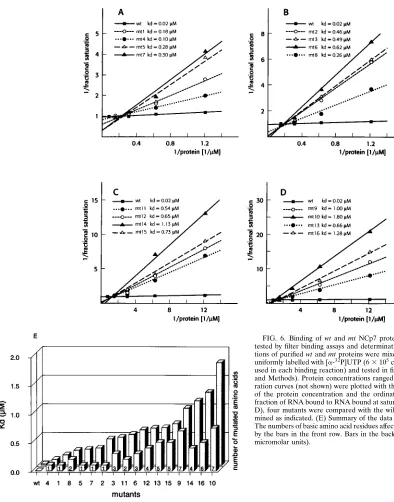
[image:3.612.318.550.300.603.2]The filter binding assays were performed with increasing NC protein concentrations, which resulted in saturation curves. The data were plotted as double-reciprocal plots, with the abscissa showing the reciprocal of the protein concentration and the ordinate indicating the reciprocal of the ratio of RNA bound to RNA bound at saturation (38) (Fig. 6A to D), which
allows determination of the dissociation constants (Kds) from
the slopes. The data in Fig. 6E show a plot of increasing Kds
which are correlated with the number of basic amino acids affected by the mutation. wt NCp7 protein bound to RNA with
a dissociation constant of 0.02mM. mt2, which contains only a
single mutation, exhibited a particularly high Kd(0.46mM) in
comparison with other mutants containing single amino acid mutations, again supporting the importance of R-7 for RNA
binding. mt3, containing three mutations, exhibited a Kd(0.49
mM) similar to that of mt2, which again demonstrated that R-3
and R-10 are less important for RNA binding. mt11, with two mutations (R-32 and K-33), also binds RNA with a significantly
reduced affinity (Kd, 0.54mM), underlining the importance of
these two amino acid residues. Two additional mutations of the adjacent amino acids, R-29 and K-34 (mt12), only slightly re-duced the binding. In all other cases, the number of mutations
directly correlated with the increase in Kd (e.g., mt9, mt10,
mt14, and mt16) (Fig. 6C and D).
Under the conditions used for the filter binding experiments, mutation of the lysine residues, especially K-11 and K-14 (mt5 and mt7, respectively), seems to have a greater influence than mutation of the arginine residue R-3 or R-10 (mt1 and mt4, respectively) in the amino-terminal region of the first zinc finger.
DISCUSSION
We have analyzed the RNA binding properties of recombi-nant wt and mt NCp7 proteins using Northwestern blots and filter binding assays. The results indicate that the importance of individual basic amino acid residues for binding of the NC protein to RNA is not uniform. Mutations of the basic amino acid residues in the amino-terminal region of the first zinc finger are not of equal importance. One mutation, a change of R-7 to N (mt2), reduces the binding affinity more than any other single mutation tested (mt1, mt4, mt5, or mt7). Mutation
of two basic amino acids on either side of R-7, R-3, and R-10 (mt3) or deletion of all amino acid residues from R-3 to R-10
(mt6) does not lead to a significantly increased Kdin
compar-ison with that of mt2. mt8, which contains an intact R-7 but mutated R-3 and R-10 residues, binds to RNA with higher affinity than mt2. These results support the importance of R-7 for RNA binding. Previously, mutagenesis of two lysine resi-dues located carboxy terminal to the first zinc finger of Rous sarcoma virus NC protein led to the loss of viral RNA pack-aging (11). Interaction of NC and PSI-containing RNA resem-bles the specific binding of Tat to TAR, which is also mediated by a single R residue within a stretch of basic amino acids consisting of eight lysines and only one arginine, at the fourth position (29).
[image:4.612.318.547.347.634.2]Apart from protecting RNA against nucleases (18), the nu-cleocapsid protein of HIV-1 was shown in vitro to stimulate different steps in the viral life cycle, such as annealing of tRNA to viral RNA (2, 28) and strand transfer (42). Nonspecific RNA binding is a prerequisite for these activities. tRNA-RNA annealing and strand transfer are steps of the replication cycle, and a defect in nonspecific RNA binding of nucleocapsid pro-tein should therefore result in a reduction of infectivity of the virus. This phenotype was described in vivo for strains with mutations in the basic amino acid residues on both sides of the
FIG. 4. Binding of NCp7 wt protein to the HIV-1 59leader RNA and to a deletion mutant, as tested by Northwestern blot analysis. Purified recombinant NCp7 wt protein (0.5 mg [lane 1] and 1 mg [lane 2]) was probed withCRNA and
CRNAD299–307. RNA binding with 33106cpm (63105cpm/mg of RNA) of
radioactively labelled transcript was performed under standard assay conditions (see Materials and Methods). Binding of radioactively labelled RNA to immo-bilized NCp7 wt protein was visualized by autoradiography.
FIG. 5. (A) Northwestern blot analysis of NC protein-RNA interaction using purified wt and mt NCp7 proteins. For nomenclature see the legend to Fig. 1. Matched amounts of purified recombinant wt and mt NCp7 proteins isolated by Ni-chelating column chromatography, shown in Fig. 2B, were probed with ra-dioactively labelled HIV-1 59leader RNA (63105
cpm/mg of RNA; 33106
cpm was used in each assay) in Northwestern blots with 400 mM KCl and 0.5 mg of heparin per ml. (B) Quantification by phosphorimaging of radioactive RNA bound to various mutants. The numbers of basic amino acid residues affected by the mutations are illustrated by the bars in the front row. Bars in the back row reflect the mean values calculated from different measurements of the radioac-tivity retained on the membrane.
on November 9, 2019 by guest
http://jvi.asm.org/
zinc finger of the Moloney murine leukemia virus NCp10 pro-tein (17).
Mutation of two of the four basic amino acid residues in the carboxy-terminal basic region, specifically, R-32 and K-33 in mt11, leads to a significant reduction in RNA binding. Addi-tional mutation of the flanking basic amino acid residues R-29 and K-34 (mt12) results in a slight further reduction of binding activity. Thus, it can be concluded that the amino acid residues R-7, R-32, and K-33 are the most relevant for the interaction of NCp7 with viral RNA.
It is interesting that the RNA-binding proteins are charac-terized by a highly conserved consensus sequence, RXXRR or RXXRK, where X represents any amino acid residue (20). Such an RNA binding motif is present in both basic domains
flanking the first zinc finger of HIV-1. R-7 corresponds to the first arginine of the RXXRK motif amino terminal to the zinc finger, and R-32–K-33 corresponds to the latter basic amino acids of the motif located carboxy terminal to the zinc finger. This motif is also found in other retroviruses such as simian immunodeficiency virus, HIV-2, Rous sarcoma virus, and Moloney murine leukemia virus (unpublished observation). Interestingly, many nonretroviral RNA-binding proteins, such as ribosomal proteins and the 70-kDa U1 small nuclear ribo-nucleoprotein, contain this motif. Since we found that a num-ber of distinct amino acid residues within this motif are re-sponsible for nonspecific interaction with RNA, it is not surprising that nonspecific nucleic acid-binding proteins such as protamines and histones also show similarities to this
do-FIG. 6. Binding of wt and mt NCp7 proteins to HIV-1 59leader RNA as tested by filter binding assays and determination of Kds. Increasing
concentra-tions of purified wt and mt proteins were mixed with in vitro-transcribed RNA uniformly labelled with [a-32P]UTP (63105cpm/mg of RNA; 23104cpm was
used in each binding reaction) and tested in filter binding assays (see Materials and Methods). Protein concentrations ranged from 0.0625 to 1mM. The satu-ration curves (not shown) were plotted with the abscissa showing the reciprocal of the protein concentration and the ordinate showing the reciprocal of the fraction of RNA bound to RNA bound at saturation. For each panel (A through D), four mutants were compared with the wild type and their Kds were
deter-mined as indicated. (E) Summary of the data obtained by filter binding assays. The numbers of basic amino acid residues affected by the mutation are illustrated by the bars in the front row. Bars in the back row represent the Kdvalues (in
micromolar units).
on November 9, 2019 by guest
http://jvi.asm.org/
[image:5.612.61.455.66.568.2]main (25). The RNA binding motif RXXRR differs from the
LAST motif [(K/R)3(S/T)2], which is typical for single-stranded
DNA-binding proteins such as the Gene32 protein of phage T4 (5). Such proteins are helix-destabilizing proteins with melting but not hybridization ability. Thus, the two repeats of the RNA binding motif may be responsible for the unique hybridization properties of NC proteins, which enhance RNA dimerization or binding of tRNA to the RNA primer binding site (2, 27) as well as enhancing the turnover rates of the interaction of a ribozyme with its RNA substrate (24, 40).
We demonstrated previously (8) that in particular, the amino acid residues R-3, R-7, and R-10, located amino termi-nal to the first zinc finger and R-29 and R-32, located carboxy terminal to that zinc finger, are essential for specific binding to the packaging site PSI of HIV-1 RNA. These findings are supported by the data presented here. By employing stringent conditions such as high salt concentrations and addition of heparin for Northwestern blot analysis, nonspecific ionic inter-actions between amino acids and RNA are reduced. There-fore, this assay allows the analysis of more specific types of interactions with higher sensitivity. The distinct reduction in RNA binding obtained for mutations affecting R-7 and those affecting R-32 and K-33 implicates the participation of these residues in the recognition of RNA. Northwestern blot and filter binding assays show slightly different results with regard to the influence of specific mutations on RNA binding. This may be due to a marked influence of ionic interaction on RNA binding. The lysine residues K-11 and K-14, which were found to be unimportant for RNA binding by Northwestern blot analysis, were subsequently analyzed in filter binding assays
and found to have Kdvalues very similar to that of mt8, which
has mutations affecting two basic amino acids, R-3 and R-10. These results might reflect the finding that mutation of the lysine residues within the basic amino acid motifs on either side of the zinc finger leads to a more pronounced reduction in binding to an RNA molecule that does not contain the HIV-1 PSI site (reference 24 and data not shown). The helical wheel projection of the first 15 amino acid residues (38) placed K-11 and K-14 in the immediate vicinity of R-7.
Apart from the exceptions mentioned above, the reduction of RNA binding activity adheres to the following rule: the fewer the basic amino acid residues, the lower the RNA bind-ing activity.
Recently a synthetic analog with a modification of the con-figuration of the proline residue at position 31 (P-31) located in the second basic region was studied by nuclear magnetic resonance and fluorescence techniques (22). P-31 induces a kink in the linker between the two zinc fingers, which causes the second basic region to be fully exposed to the solvent and to easily react with RNA. It was found that the change in conformation reduces the ability to promote HIV-1 RNA dimerization in vitro. A substitution of L for P-31 resulted in a drastic modification of virion formation and infectivity.
Our results clearly demonstrate that two distinct basic amino acid domains are involved in nonspecific NC protein-RNA interaction. Some of these residues were previously shown to be important for the specific interaction between the packaging signal of the HIV-1 RNA and the NC protein (8). NC protein was recently shown to enhance the activity of a hammerhead ribozyme (24, 40). This activity is shared by the heterogeneous nuclear ribonucleoprotein A1 (4) as well as a peptide polymer containing six to eight repeats of the carboxy-terminal RGG box motifs of the A1 protein (15). Even though nonspecific RNA binding is a prerequisite for facilitation of hammerhead ribozyme catalysis, it is not sufficient for stimulation of multiple turnover reactions (15). These findings indicate that repetition
of an RNA binding motif, as seen for the NC protein and the carboxy-terminal domain of the heterogeneous nuclear ribo-nucleoprotein A1, may be a common feature of proteins in-volved in processes that require association as well as dissoci-ation of RNA molecules. Our data show that three distinct amino acid residues, R-7, R-32, and K-33, flanking the first zinc finger of HIV-1 NCp7 are the most essential ones for RNA binding in vitro. Thus, in planning the construction of possible antiviral therapeutics involving NCp7-RNA binding, the unique role of these three particular amino acids should be considered.
ACKNOWLEDGMENTS
This work was supported by the Deutsche Forschungsgemeinschaft (SFB344), the Dr. Mildred Scheel Stiftung (F77), the Vaillant Stiftung, and the Swiss National AIDS Research Program.
REFERENCES
1. Aldovini, A., and R. A. Young. 1990. Mutations of RNA and protein se-quences involved in human immunodeficiency virus type 1 packaging result in production of noninfectious virus. J. Virol. 64:1920–1926.
2. Barat, C., V. Lullien, O. Schatz, G. Keith, M. T. Nugeyre, F. Gru ¨ninger-Leitch, F. Barre´-Sinnoussi, S. F. J. LeGrice, and J. L. Darlix.1989. HIV-1 reverse transcriptase specifically interacts with the anticodon domain of its cognate primer tRNA. EMBO J. 8:3279–3285.
3. Berkowitz, R. D., J. Luban, and S. P. Goff. 1993. Specific binding of human immunodeficiency virus type 1 gag polyprotein and nucleocapsid protein to viral RNAs detected by RNA mobility shift assays. J. Virol. 67:7190–7200. 4. Bertrand, E. L., and J. J. Rossi. 1994. Facilitation of hammerhead ribozyme catalysis by the nucleocapsid protein of HIV-1 and the heterogeneous nu-clear ribonucleoprotein A1. EMBO J. 13:2904–2912.
5. Casas-Finet, J. R., K. R. Fisher, and R. L. Karpel. 1992. Structural basis for the nucleic acid binding cooperativity of bacteriophage T4 gene 32 protein: the (Lys/Arg)3(Ser/Thr)2 (LAST) motif. Proc. Natl. Acad. Sci. USA 89: 1050–1054.
6. Covey, S. N. 1986. Amino acid sequence homology in the gag region of reverse transcribing elements and the coat protein gene of cauliflower mo-saic virus. Nucleic Acids Res. 14:623–633.
7. Daly, T. J., K. S. Cook, T. E. Maione, and J. R. Rusche. 1989. Specific binding of HIV-1 recombinant Rev protein to the Rev-responsive element in vitro. Nature (London) 342:816–819.
8. Dannull, J., A. Surovoy, G. Jung, and K. Moelling. 1994. Specific binding of HIV-1 nucleocapsid protein to PSI RNA in vitro requires N-terminal zinc finger and flanking basic amino acid residues. EMBO J. 13:1525–1533. 9. DeRocquigny, H., C. Gabus, A. Vincent, M. C. Fournie-Zaluski, B. Rocques,
and J. L. Darlix.1992. Viral RNA annealing activities of human immuno-deficiency virus type 1 nucleocapsid protein require only peptide domains outside the zinc fingers. Proc. Natl. Acad. Sci. USA 89:6472–6476. 10. Dorfman, T., J. Luban, S. P. Goff, W. A. Haseltine, and H. G. Go¨ttlinger.
1993. Mapping of functionally important residues of a cysteine-histidine box in the human immunodeficiency virus type 1 nucleocapsid protein. J. Virol. 67:6159–6169.
11. Fu, X., R. A. Kats, A. M. Skalka, and J. Leis. 1988. Site-directed mutagenesis of the avian nucleocapsid protein, pp12. J. Biol. Chem. 263:2140–2145. 12. Gorelick, R. J., D. J. Chabot, A. Rein, L. E. Henderson, and L. O. Arthur.
1993. The two zinc fingers in the human immunodeficiency virus type 1 nucleocapsid protein are not functionally equivalent. J. Virol. 67:4027–4036. 13. Henderson, L. E., M. A. Bowers, R. C. Sowder II, S. A. Serabyn, D. G. Johnson, J. W. Bess, Jr., L. O. Arthur, D. K. Bryant, and C. Fenselau.1992. Gag proteins of the highly replicative MN strain of human immunodeficiency virus type 1: posttranslational modifications, proteolytic processings, and complete amino acid sequences. J. Virol. 66:1856–1865.
14. Henderson, L. E., R. C. Sowder, T. D. Copeland, S. Oroszlan, and R. E. Benveniste.1990. Gag precursors of HIV and SIV are cleaved into six proteins found in mature virions. J. Med. Primatol. 19:411–419.
15. Herschlag, D., M. Khosla, Z. Tsuchihashi, and R. L. Karpel. 1994. An RNA chaperone activity of non-specific RNA binding proteins in hammerhead ribozyme catalysis. EMBO J. 13:2913–2924.
16. Hochuli, E., W. Bannwarth, H. Do¨berli, R. Gentz, and D. Stu¨ber.1988. Genetic approach to facilitate purification of recombinant proteins with a novel metal chelate adsorbant. Bio/Technology 6:1321–1325.
17. Housset, V., H. De Rocquigny, B. P. Roques, and J.-L. Darlix. 1993. Basic amino acids flanking the zinc finger of Moloney murine leukemia virus nucleocapsid protein NCp10 are critical for virus infectivity. J. Virol. 67: 2537–2545.
18. Karpel, R. L., L. E. Henderson, and S. Oroszlan. 1987. Interactions of retroviral structural proteins with single-stranded nucleic acids. J. Biol. Chem. 262:4961–4967.
on November 9, 2019 by guest
http://jvi.asm.org/
19. Lapadat-Tapolski, M., H. DeRocquigny, D. van Gent, B. Roques, R. Plas-terk, and J. L. Darlix.1993. Interactions between HIV-1 nucleocapsid pro-tein and viral DNA may have important functions in the viral life cycle. Nucleic Acids Res. 21:831–839.
20. Lazinski, D., E. Grzadzilska, and A. Das. 1989. Sequence specific recognition of RNA hairpins by bacteriophage antiterminators requires a conserved arginine-rich motif. Cell 59:207–218.
21. Me´ric, C., and S. P. Goff. 1989. Characterization of Moloney murine leuke-mia virus mutants with single-amino-acid substitutions and the Cys-His box of the nucleocapsid protein. J. Virol. 63:1558–1568.
22. Morellet, N., H. DeRocquigny, Y. Me´ly, N. Jullian, H. De´me´ne´, M. Ottmann, D. Ge´rard, J. L. Darlix, M. C. Fournie-Zaluski, and B. P. Roques.1994. Conformational behavior of the active and inactive forms of the nucleocap-sid NCp7 of HIV-1 studied by NMR. J. Mol. Biol. 235:287–301. 23. Morellet, N., N. Jullian, H. DeRocquigny, B. Maigret, J. L. Darlix, and B. P.
Rocques.1992. Determination of the structure of the nucleocapsid protein NCp7 from the human immunodeficiency virus type 1 by1
H NMR. EMBO J. 11:3059–3065.
24. Mueller, G., B. Strack, J. Dannull, B. S. Sproat, A. Surovoy, G. Jung, and K. Moelling.1994. Amino acid requirements of the nucleocapsid of HIV-1 for increasing activity of a Ki-ras ribozyme in vitro. J. Mol. Biol. 242:422–429. 25. Nagai, K., C. Oubridge, T. H. Jessen, J. Li, and P. R. Evans. 1990. Crystal
structure of the RNA-binding domain of the U1 small nuclear ribonucleo-protein A. Nature (London) 348:515–520.
26. Portman, D. S., and G. Dreyfuss. 1994. RNA annealing activity in Hela nuclei. EMBO J. 13:213–221.
27. Prats, A. C., V. Housset, G. de Billy, F. Cornille, H. Prats, B. Roques, and J. L. Darlix.1991. Viral RNA annealing activities of the nucleocapsid pro-tein of Moloney murine leukemia virus are zinc dependent. Nucleic Acids Res. 19:3541–3553.
28. Prats, A. C., L. Sarih, C. Gabus, S. Litvak, G. Keith, and J. L. Darlix. 1988. Small finger protein of avian and murine retroviruses has nucleic acid an-nealing activity and positions the replication primer tRNA onto genomic RNA. EMBO J. 7:1777–1783.
29. Puglisi, J. D., T. Ruoying, B. J. Calnan, A. D. Frankel, and J. R. Williamson. 1992. Conformation of the TAR RNA-arginine complex by NMR spectros-copy. Science 257:76–79.
30. Ratner, L., W. Haseltine, R. Patarca, K. J. Livak, B. Starcich, S. F. Josephs,
E. R. Doran, J. A. Rafalski, E. A. Whitehorn, K. Baumeister, L. Ivanoff, S. R. Petteway, Jr., M. L. Pearson, J. A. Lautenberger, T. S. Papas, J. Grayeb, N. T. Chang, R. C. Gallo, and F. Wong-Staal.1985. Complete nucleotide sequence of the AIDS virus, HTLV-III. Nature (London) 313:277–284. 31. Riggs, A. D., and S. Bourgeois. 1968. On the assay, isolation and
character-ization of the lac repressor. J. Mol. Biol. 34:361–364.
32. Rosenberg, A. H., B. N. Lade, D. S. Chui, S. W. Lin, J. J. Dunn, and F. W. Studier.1987. Vectors for selective expression of cloned DNAs by T7 RNA polymerase. Gene 56:135–145.
33. Sanger, F., S. Nicklen, and A. R. Coulson. 1977. DNA sequencing with chain-terminating inhibitors. Proc. Natl. Acad. Sci. USA 74:5463–5467. 34. South, T. L., P. R. Blake, R. C. Sowder, L. O. Arthur, L. E. Henderson, and
M. F. Summers.1990. The nucleocapsid protein isolated from HIV-1 par-ticles binds zinc and forms retroviral-type zinc fingers. Biochemistry 29: 7786–7789.
35. South, T. L., and M. F. Summers. 1993. Zinc- and sequence-dependent binding to nucleic acids by the N-terminal zinc finger of the HIV-1 nucleo-capsid protein: NMR structure of the complex with the Psi-site analog, dACGCC. Protein Sci. 2:3–19.
36. Studier, F. W., and A. Moffat. 1986. Use of bacteriophage T7 RNA poly-merase to direct selective high-level expression of cloned genes. J. Mol. Biol. 189:113–130.
37. Summers, M. F., T. L. South, B. Kim, and D. R. Hare. 1990. High-resolution structure of an HIV zinc fingerlike domain via a new NMR-based distance geometry approach. Biochemistry 29:329–340.
38. Surovoy, A., J. Dannull, K. Moelling, and G. Jung. 1993. Conformational and nucleic acid binding studies on the synthetic nucleocapsid protein of HIV-1. J. Mol. Biol. 229:94–104.
39. Sykora, K. W., and K. Moelling. 1981. Properties of the avian viral protein p12. J. Gen. Virol. 55:379–391.
40. Tsuchihashi, Z., M. Khosla, and D. Herschlag. 1993. Protein enhancement of hammerhead ribozyme catalysis. Science 262:99–102.
41. Volkmann, S., J. Dannull, and K. Moelling. 1993. The polypurin tract, PPT, of HIV as a target for antisense and triple-helix-forming oligonucleotides. Biochimie (Paris) 75:71–78.
42. You, J. C., and C. S. McHenry. 1994. Human immunodeficiency virus nu-cleocapsid protein accelerates strand transfer of the terminally redundant sequences involved in reverse transcription. J. Biol. Chem. 269:31491–31495.