Copyright © 1999, American Society for Microbiology. All Rights Reserved.
Functional Interactions of the HHCC Domain of Moloney Murine
Leukemia Virus Integrase Revealed by Nonoverlapping
Complementation and Zinc-Dependent Dimerization
FAN YANG,1OSCAR LEON,2NORMA J. GREENFIELD,3ANDMONICA J. ROTH1*
Department of Biochemistry1and Department of Neuroscience and Cell Biology,3University of Medicine and Dentistry
of New Jersey-Robert Wood Johnson Medical School, Piscataway, New Jersey 08854, and Instituto de Bioquimica,
Facultad de Ciencias, Universidad Austral de Chile, Valdivia, Chile2
Received 6 August 1998/Accepted 9 December 1998
The retroviral integrase (IN) is required for the integration of viral DNA into the host genome. The N terminus of IN contains an HHCC zinc finger-like motif, which is conserved among all retroviruses. To study the function of the HHCC domain of Moloney murine leukemia virus IN, the first N-terminal 105 residues were expressed independently. This HHCC domain protein is found to complement a completely nonoverlapping construct lacking the HHCC domain for strand transfer, 3*processing and coordinated disintegration reac-tions, revealingtransinteractions among IN domains. The HHCC domain protein binds zinc at a 1:1 ratio and changes its conformation upon binding to zinc. The presence of zinc within the HHCC domain stimulates selective integration processes. Zinc promotes the dimerization of the HHCC domain and protects it from
N-ethylmaleimide modification. These studies dissect and define the requirement for the HHCC domain, the exact function of which remains unknown.
An essential step of the retroviral life cycle is the integration of the reverse-transcribed viral DNA into the host genome. This two-step process is carried out by the viral enzyme, inte-grase (IN), encoded by thepolgene (for a review, see reference 5). First, two deoxynucleotides are removed from both 39 ter-mini of the viral long terminal repeats (LTRs), exposing the conserved 59-CA-39at the recessed 39ends (39processing) (6, 29, 44, 54). The 39ends of the viral DNA are then joined to 59
staggered sites in the target DNA in a concerted transesterifi-cation reaction (strand transfer) (14, 27, 43). Both the 39 pro-cessing and strand transfer reactions are isoenergetic and oc-cur without ATP (27). Integration is completed by repair of the 59overhang of the viral LTR and the single-strand gaps flank-ing the integrated viral DNA, presumably by host enzymes. Repair of these single-strand gaps creates the hallmark dupli-cation of target DNA sequence flanking the retroviral integra-tion site. For Moloney murine leukemia virus (M-MuLV), the direct duplication is 4 bp long.
In vitro assays have been developed to study the mechanism of retroviral integration by using purified enzymes and short oligodeoxynucleotide duplexes mimicking the viral LTR ends (14, 44). Purified IN is also able to carry out disintegration reactions with either Y oligomer (13) or crossbone substrates (12, 15), which contain single and double LTRs, respectively. Sequence comparison and mutational analysis have identi-fied three functional domains in the IN protein (9, 24, 36, 40, 45, 46, 63, 67). The first two domains are highly conserved among all retroviruses and retrotransposons. The N-terminal region is characterized by an HHCC zinc finger motif (8, 9, 36, 68). Mutational studies of the conserved cysteine or histidine residues produced varied results regarding the importance of the HHCC domain (9, 10, 17, 26, 40, 42, 45, 47, 53, 55, 61–63).
The central region of IN is characterized by a D-D(35)-E motif. Mutation of the conserved aspartic or glutamic acid residues results in loss of all catalytic activities, indicating a role for these three residues in active site function (17, 26, 46, 47, 61, 62). The conserved residues in both the N terminus and the central core are also required for viral replication in vivo (25, 53, 56, 59, 65). The C-terminal region is less conserved and has been demonstrated to possess nonspecific DNA-binding activity (23, 45, 51, 52, 63, 66, 67).
There is strong evidence that the IN protein functions as a multimer (37). Within individual subdomains, homodimers have been identified. Dimers of the avian sarcoma virus and human immunodeficiency virus (HIV) core domains (7, 18), the HIV-1 C terminus (20, 50), and the HHCC region (68) have been identified. The association of these IN subdomains to form higher-ordered protein-DNA complexes is unknown. Complementation experiments employing different mutants of HIV-1 and M-MuLV IN support the multimerization of IN and provide an alternative approach in identifying protein-protein interactions (22, 24, 38, 60).
There is general agreement that the HHCC domain is es-sential for integration in vivo (25, 47, 53, 65); however, the function as assayed in vitro is less well defined. Although zinc finger domains frequently bind DNA, no evidence for DNA binding has been reported to date for the IN HHCC domain (45, 55); however, indirect effects have been noted (32, 53). Functions that might affect recognition (33, 45, 62) or posi-tioning (39, 61) of the viral LTRs have been observed to be associated with the HHCC domain. Formation of a stable IN-LTR complex requires the HHCC region (22, 64). MuLV LTR substrates which lack the 59 single-stranded (SS) tail require the presence of an HHCC domain for disintegration and coordinated disintegration reactions (15, 16). The N ter-minus of HIV-1 IN was found to cross-link to the target DNA (34, 35). The HHCC domain is also reported to be involved in protein-protein interactions and the process of tion (15, 22, 49, 68). Zinc is reported to promote multimeriza-* Corresponding author. Mailing address: Department of
Biochem-istry, University of Medicine and Dentistry of New Jersey-Robert Wood Johnson Medical School, 675 Hoes Ln., Piscataway, NJ 08854. Phone: (732) 235-5048. Fax: (732) 235-4783. E-mail: Roth@waksman .rutgers.edu.
1809
on November 9, 2019 by guest
http://jvi.asm.org/
tion of HIV-1 IN and to stimulate Mg21-dependent 39
pro-cessing and strand transfer activity (48, 49, 68).
Here, we report that the N terminus of M-MuLV IN con-taining the HHCC domain complements a completely non-overlapping domain of M-MuLV IN containing the core and C terminus. This isolated HHCC domain binds zinc with a 1:1 ratio. Zinc binding induces a conformational change of the HHCC domain and stimulates the catalytic activity of M-MuLV IN in complementation assays.
MATERIALS AND METHODS
Materials.Crude [g-32P]ATP (7,000 Ci/mmol) was purchased from ICN.
[a-32P]dATP and [a-32P]TTP were purchased from Amersham. T4
polynucle-otide kinase was obtained from New England Biolabs. Exonuclease-free Klenow fragment of DNA polymerase I was obtained from United States Biochemical. Ni21-nitrilotriacetic acid agarose was purchased from Qiagen. All restriction
enzymes were purchased from New England Biolabs. RecA protein was pur-chased from Promega. GlutathioneS-transferase–HMGI-C was kindly provided by Kiran Chada (69).
Oligonucleotides.DNA oligonucleotides were prepared by the University of Medicine and Dentistry of New Jersey Biochemistry Department Synthesis Fa-cility and purified by electrophoresis on 20% denaturing polyacrylamide gels. Oligonucleotides used in this study are referred to by their synthesis numbers and were labeled and prepared as described elsewhere (15, 16, 40). Oligonucleotides 2783 (59-GTCAGCGGGGGTCTTTCATT), 2784 (59-GTCAGCGGGGGTCTT TCA), and 2785 (59-AATGAAAGACCCCCGCTGAC) were used for 39 pro-cessing and strand transfer reactions. Oligonucleotides 4166 (59-AATGAAAG
TTCTTTCACGCTAGTCCTTGGAC), 4167 (59AATGAAAGTTCTTTCAAG
CGAGTCCTTGGAC), 5354 (59-TGAAAGTTCTTTCACGCTAGTCCTTGG
AC), and 5355 (59-TGAAAGTTCTTTCAAGCGAGTCCTTGGAC) were
substrates for the coordinated disintegration reactions. Oligonucleotide 7440
(59-ACCTGCGTAAGCAGGTAGACCGCAAGGTCTACTTTCGAATGCGA
AAGT) was used for the disintegration reaction.
Construction of M-MuLV HHCC mutant.Construction and expression of wild-type IN and mutants ND105 and CD232 were previously described (38, 40). M-MuLV HHCC mutant CD303 was constructed by PCR amplification with Vent DNA polymerase (New England Biolabs), with the plasmid CD232 as template and the T7 primer (59-TAATACGACTCACTATAGGG) (Promega) as upstream primer. The downstream primer, 6351 (59-CGGGATCCTAAGAC TTGCTGGCGTTGAC), encodes a stop codon and aBamHI site adjacent to sequence complementary to the IN mRNA coding region (indicated in boldface; the firstAis complementary to position 4925 in the MuLV RNA sequence [57]). The PCR product was digested withNdeI andBamHI and exchanged for the 1.2-kbNdeI andBamHI fragment from the construct CD232. The nucleotide sequence of the construct was verified by dideoxy sequencing with an AmpliCycle sequencing kit from Perkin-Elmer.
Purification of M-MuLV IN.Recombinant M-MuLV IN (WT, ND105, and CD232) containing a hexahistidine tag was expressed in Escherichia coli BL21(DE3) (Novagen) and purified by Ni21-nitriloacetate chromatography
(Qiagen) as previously described (38, 40). CD303 was expressed and purified similarly. CD303/zinc was renatured in the presence of 10mM ZnCl2; zinc was
omitted in the last step of renaturation. CD303 was further purified on a car-boxymethyl Sepharose column and eluted with a 0 to 1 M NaCl gradient in buffer containing 10 mM Tris-HCl (pH 7.5), 2 mM dithiothreitol (DTT), and 5% glycerol. The peak fractions were pooled and concentrated with a Centricon-10 concentrator (Amicon). The concentrated sample was then applied onto a Su-perose-12 fast protein liquid chromatography column, and fractions were col-lected by monitoring absorption at 280 nm. To remove the hexahistidine tag from CD303, the peak fractions from carboxymethyl Sepharose were dialyzed in 50 mM Tris-HCl (pH 7.5)–50 mM NaCl–2.5 mM CaCl2–2 mM DTT–0.1% Nonidet
P-40 (NP-40)–4% glycerol and then cleaved by thrombin (3 U/mg of IN) (Sigma) at room temperature for 1 h. The cleaved HHCC mutant was separated from the tagged protein by further purification on a P11 column. The cleaved protein was eluted from the P11 column with a 0 to 1 M NaCl gradient in buffer containing 10 mM Tris-HCl (pH 7.5), 2 mM DTT, and 5% glycerol.
In vitro assays.Strand transfer, 39processing, disintegration, and coordinated disintegration reactions were performed as previously described (15, 40). Typi-cally, reactions contained 1 pmol of labeled substrate and 7.5 to 10 pmol of IN protein. Complementation assays were performed by titrating the HHCC finger domain protein against a fixed level of ND105. The buffer for strand transfer and 39processing reactions contained 20 mM morpholineethanesulfonic acid (MES; pH 6.2), 10 mM DTT, 10 mM MnCl2, 10 mM KCl, and 10% glycerol. The buffer
for disintegration reaction had 20 mM piperazine-N,N9-bis(2-ethanesulfonic acid) (PIPES; pH 6.4), 5 mM CHAPS (3-[(3-cholamidopropyl)-dimethylammo-nio]-1-propanesulfonate), 10 mM DTT, 25 mM MnCl2, and 0.05% NP-40. The
buffer for coordinated disintegration reactions included 20 mM PIPES (pH 6.4), 10 mM CHAPS, 10 mM DTT, 10 mM MnCl2, 0.01% NP-40, 5% ethylene glycol,
and 10 mM NaCl. Ratios of HHCC domain to ND105 varied from 0.1:1 to 10:1 as indicated in each figure. For theN-ethylmaleimide (NEM) protection assay,
the proteins were treated with 10 mM NEM as described elsewhere (39). To label CD303 proteins with fluorescent maleimide, 2ml of 50 mMN(1-pyrene) maleimide in dimethyl sulfoxide was added to 10ml of CD303 protein sample at 40 pmol/ml and incubated on ice for 90 min. The reaction was stopped by adding 6ml of sodium dodecyl sulfate-polyacrylamide gel electrophoresis sample buffer, and the mixture was heated for 2 min at 100°C and subjected to electrophoresis on a 15% acrylamide gel. The fluorescent picture was taken with a Vilber Lourmat (France) transilluminator and a Canon A-1 camera with a yellow filter (Agfa G2). Protein concentration was measured by the method of Bradford (3) (Bio-Rad).
Circular dichroism (CD) spectroscopy.CD measurements were performed on an Aviv model 62D spectropolarimeter equipped with a thermally regulated cell holder. The CD far-UV spectra were analyzed at both 5 and 37°C from 250 to 200 nm, every 0.25 nm, with a 2-s collection time, in 1-mm rectangular quartz cuvettes. The content of secondary structures was calculated by using three different curve fitting programs: Lincomb (constrained least squares fit) (4), MLR (nonconstrained least squares fit) (4), and Selcon (41, 58) (for details of the programs, see the review in reference 31).
Zinc analysis by atomic absorption spectroscopy.Zinc concentration in the protein was measured on a Perkin-Elmer model 3100 atomic absorption spec-trometer, which was calibrated with a zinc standard curve made from a standard zinc solution (EM Science). The zinc concentration was measured by using the flame mode of the spectrometer. The protein concentrations of the samples were determined by a difference spectrum method (19, 28, 30), and the stoichiometry of zinc to HHCC domain was calculated.
RESULTS
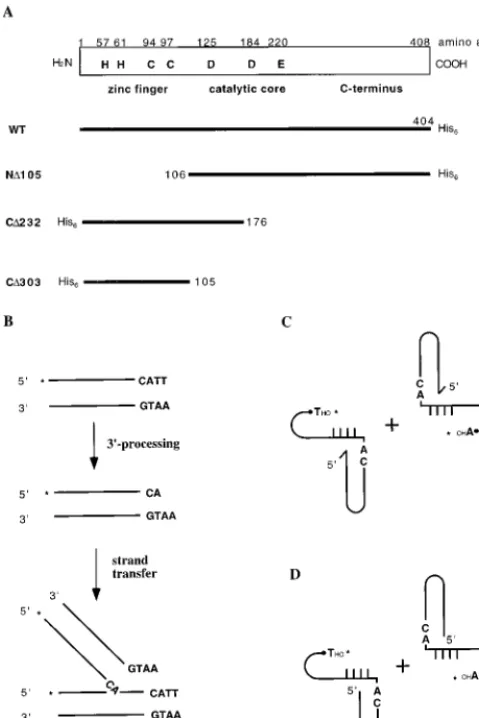
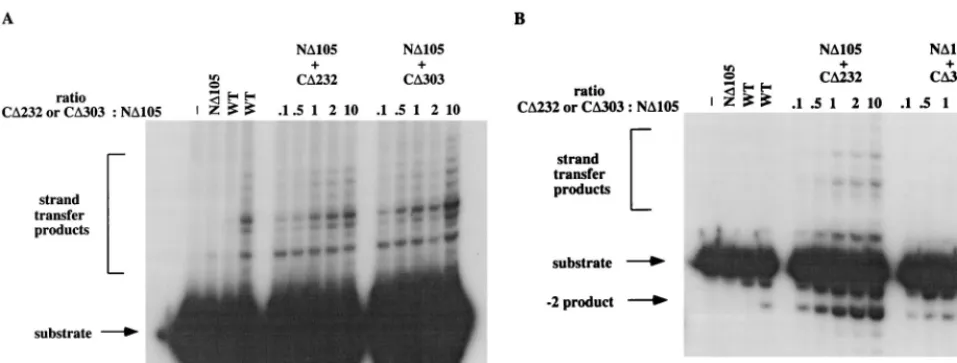
Complementation of ND105 by a nonoverlapping HHCC domain.The schematic representation of M-MuLV IN and IN mutants is shown in Fig. 1. Previous work in our laboratory has shown that the N-terminal deletion M-MuLV IN mutant ND105 (up to 40 pmol) produced a very low level of strand transfer at one preferential site, whereas no 39 processing could be detected (Fig. 2A, lane 2) (references 16 and 38 and data not shown). ND105 could be complemented by another mutant, CD232, which has a region of 71 amino acids overlap-ping with ND105 (38). To determine if this overlapping region is essential for CD232’s ability to rescue ND105 for strand transfer reactions, a smaller nonoverlapping HHCC construct containing amino acid residues 1 to 105 (CD303) was gener-ated. The ability of CD303 to complement ND105 for strand transfer and 39processing reactions was analyzed. For strand transfer reaction, a precleaved substrate, which lacks the two terminal bases lost during 39 processing, thus exposing the 59-CA-39end, was used (Fig. 1B). Addition of CD303 restores ND105’s ability to catalyze strand transfer reaction (Fig. 2A, lanes 10 to 14). The level of complementation achieved by CD303 is similar to that by CD232 (Fig. 2A, compare lanes 5 to 9 and lanes 10 to 14), indicating that the overlapping region from residues 106 to 176 is not required for the HHCC domain to complement ND105, at least for the strand transfer reaction. As a control, CD303 at 100 pmol had no strand transfer activity (data not shown). No complementing activity was detected when either RecA or glutathioneS-transferase–HMGI-C pro-tein was used to replace CD303 (data not shown).
In the 39processing reaction with a blunt-ended oligonucle-otide substrate (Fig. 1B), CD303 complements ND105 to pro-duce a “22” product. Interestingly, the efficiency of CD303 is less than that of CD232 (compare lanes 5 to 9 and lanes 10 to 14 in Fig. 2B), though the same fractions were used for strand transfer and 39processing reactions. This implies that the 71-amino-acid overlapping region is stabilizing the interaction between CD232 and ND105 for 39processing.
Zinc stimulates HHCC domain’s ability to complement ND105.Studies with HIV IN have shown that HIV IN binds zinc (8, 9, 32, 68), and zinc stimulates IN in strand transfer and 39processing reactions (48, 68). To see if zinc has any effect on M-MuLV IN, the HHCC domain protein CD303 was purified and refolded in the presence (CD303/zinc) or absence (CD303/ EDTA) of zinc (see Materials and Methods for details) and
on November 9, 2019 by guest
http://jvi.asm.org/
assayed for complementation of ND105 in strand transfer, 39
processing, and coordinated disintegration reactions.
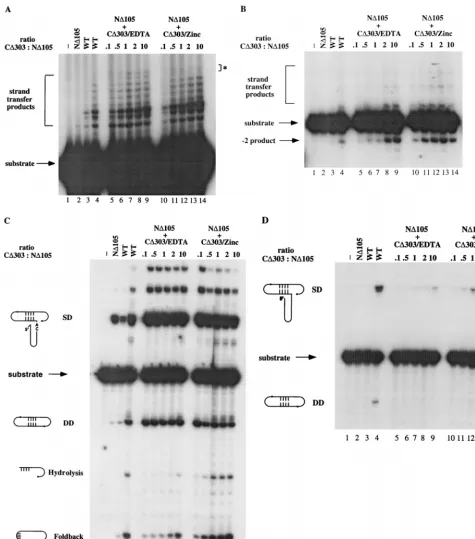
In the strand transfer reaction, both the CD303 proteins, with or without zinc, are capable of complementing ND105 (compare lanes 5 to 9 with lanes 10 to 14 in Fig. 3A). There is a subtle but distinct difference in the pattern of strand transfer products between CD303/zinc and CD303/EDTA. CD303/ EDTA complements ND105 to give rise to larger strand trans-fer products, which are barely detectable in the complementa-tion by CD303/zinc (see asterisk in Fig. 3A). The pattern for CD303/zinc (Fig. 3A, lanes 10 to 14) resembles more that of WT IN (Fig. 3A, lane 4), which was purified without zinc.
For 39processing and coordinated disintegration reactions, the effect of zinc within the HHCC domain is more obvious. For 39 processing, complementation of ND105 occurs with lower concentrations of CD303/zinc HHCC protein than of CD303/EDTA (compare lanes 5 to 9 with lanes 10 to 14 in Fig. 3B). CD303/zinc saturated ND105 at a ratio of 0.5 to 1 HHCC/ ND105 subunit, whereas CD303/EDTA required minimally two HHCC/ND105 subunits for maximal activity.
In coordinated disintegration reactions with two half-cross-bone substrates (Fig. 1C and D), four products have been previously identified, including single-end disintegration prod-uct (SD), a circular prodprod-uct resulting from disintegration of both LTR ends (DD), a small circular foldback product, and a hydrolysis product releasing the viral LTR (15). For coordi-nated disintegration reactions with the crossbone substrates containing 59-ss overhang, the presence of zinc stimulates the production of the foldback and hydrolysis products (Fig. 3C, compare lanes 5 to 9 and lanes 10 to 14). In the coordinated disintegration reaction with crossbone substrates lacking the 59-ss overhang, it was previously shown that the CD232 con-struct was capable of complementing ND105, yielding the dou-ble disintegration product (15). Interestingly, CD303/EDTA yielded little if any double disintegration product (lanes 5 to 9, Fig. 3D). However, CD303 renatured with zinc yielded in-creased levels of both single and double disintegration prod-ucts (lanes 10 to 14 in Fig. 3D). This reaction required excess CD303 to ND105, with maximal stimulation detected at a 10:1 ratio.
The histidine tag was removed from CD303/zinc and ana-lyzed for complementation with strand transfer, 39processing, and untailed coordinated disintegration reactions (data not shown). CD303/zinc without the tag maintained approximately 50 to 60% of its activity in all three assays compared to the mock-treated protein. Analysis of the protein by Coomassie blue staining indicated that more than 90% of the hexahisti-dine tag was removed by thrombin cleavage (data not shown), indicating that the hexahistidine tag is not contributing to the complementing activity.
[image:3.612.55.295.73.432.2]Zinc binding of the HHCC domain protects the HHCC domain from NEM alkylation.Previous work from our labo-ratory and others has shown that both M-MuLV IN (16, 40) and HIV IN (22) are sensitive to NEM modification. For M-MuLV IN, there are three cysteine residues in total: one in the core region (C209) and two in the HHCC region (C94 and C97). NEM modification of either ND105 or CD232 reduced the ability of the two IN mutants to complement each other (16). This was most clearly observed with a dumbbell single-end disintegration substrate lacking the 59-ss tail in the viral LTR (untailed disintegration). With this substrate, the reaction is dependent on both the catalytic core and the HHCC domain (16) (Fig. 4A, lanes 3 to 5). To determine if zinc has any effect on the NEM sensitivity of the HHCC domain, both CD303/zinc and CD303/EDTA were treated with NEM and then used to complement ND105 in the untailed disintegration reaction. As shown in Fig. 4A, ND105 is active at a very low level for the untailed disintegration reaction (lane 3). Addition of either CD303/zinc or CD303/EDTA restores ND105’s ability to cata-lyze the disintegration reaction (lanes 4 and 5). NEM treat-ment of CD303/EDTA abolishes its ability to complement ND105 (lane 9), while NEM-treated CD303/zinc is still active in complementing ND105 (lane 8). CD303/zinc with the hexahis-tidine tag removed remained resistant to NEM modification (data not shown). The accessibility of the cysteines in both CD303/zinc and CD303/EDTA was further probed withN (1-pyrene) maleimide, which allows direct visualization of the conjugation of the maleimide to the cysteine by fluorescence analysis. When equivalent amounts of the two proteins, as detected by Coomassie blue staining (Fig. 4B, lanes 3 and 4), were treated withN(1-pyrene) maleimide, only CD303/EDTA could be fluorescently labeled (Fig. 4B, lane 2). CD303/zinc was not modified (Fig. 4B, lane 1). These results indicate that coordination of zinc by the HHCC domain makes the two cysteines inaccessible to NEM, thus protecting the HHCC do-main from being alkylated.
FIG. 1. Illustration of WT M-MuLV IN, mutant INs, and assays used in this study. (A) Schematic representation of WT M-MuLV IN and IN mutants used in this study. The name of each mutant is indicated to the left of each protein. (B) DNA substrates and assays for 39processing and strand transfer reactions. (C) DNA substrates and assay for coordinated disintegration reactions with cross-bone substrates with 59-ss overhang in the LTR (tailed crossbone substrates). (D) DNA substrates and assay for coordinated disintegration reaction with substrates without 59-ss overhang in the LTR (untailed crossbone substrates). The asterisks indicate the ends labeled.
on November 9, 2019 by guest
http://jvi.asm.org/
Zinc promotes dimerization of the HHCC domain.To char-acterize the multimerization state of the HHCC domain in solution, CD303 was further purified after renaturation and applied to a Superose-12 sizing column (see Materials and Methods for details). CD303 refolded in the presence of zinc eluted as a single peak between 25- and 43-kDa protein mark-ers (Fig. 5). The calculated molecular mass from the standard curve derived from the protein standard markers was 32 kDa, corresponding to a dimer of CD303; the molecular mass of a CD303 monomer is 14.5 kDa. In the absence of zinc, CD303/ EDTA yielded no detectable dimers. Rather, these proteins chromatographed heterogeneously and eluted as an aggregate in the void volume and as monomers. In one preparation, a low level of hexamers was detected (data not shown). This is con-sistent with the solubility of the two proteins. CD303/zinc could be concentrated to greater than 20 mg/ml, whereas CD303/ EDTA precipitated from solution at concentrations greater than 2 mg/ml. The presence of histidine tag does not interfere with the multimerization of the HHCC domain since CD303/ zinc lacking the histidine tag was eluted as a dimer as well (data not shown). Neither manganese nor magnesium could replace zinc to promote the dimerization of the HHCC domain (data not shown).
Zinc binds to the HHCC domain at a 1:1 ratio and induces a conformational change of the HHCC domain.To determine the stoichiometry of zinc bound to the HHCC domain of M-MuLV IN and the effects of zinc on protein conformation, both zinc concentration and CD studies were performed on CD303, renatured with zinc or EDTA. For zinc content mea-surement, CD303/zinc and CD303/EDTA were analyzed by atomic absorption spectroscopy. For CD303/zinc, the calcu-lated average molar ratio of zinc to the HHCC domain is 1.1:1, and the ratio for CD303/EDTA is 0.1:1. The presence of his-tidine tag has no effect on the concentration of zinc, as CD303/ zinc without the His tag gave similar results.
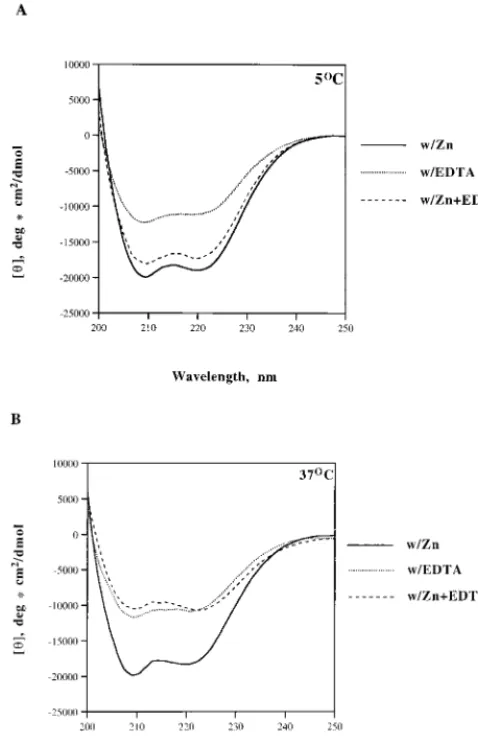
From the CD profile, both CD303 proteins (EDTA and zinc) are well structured overall, at either 5 or 37°C (Fig. 6). CD303/ EDTA contains more than 30% helical structure at both 5 and 37°C. This is consistent with results from in vitro activity assays, indicating that the HHCC protein renatured in EDTA main-tains the ability to complement the strand transfer reaction
(Fig. 3A). In the presence of zinc, the helical content of CD303 increases, with a corresponding decrease inbsheet, turn, and random-coil contents (Table 1). Addition of excessive EDTA to CD303/zinc at 37°C (Fig. 6B) would strip away zinc from the HHCC domain and result in a conformation similar to that of CD303/EDTA, which contains approximately 34% a-helix, 18%bsheet, 25% turn, and 24% random coil (Table 1). At 5°C, the CD303/zinc is generally stable despite the addition of excess EDTA by maintaining a helical content of 49% (Table 1).
DISCUSSION
Previous work in our laboratory has shown that M-MuLV IN mutant ND105 lacking the HHCC domain can be rescued by another mutant, CD232, lacking part of the core region and the C terminus (38). Our current study shows that the overlapping region between ND105 and CD232 was not required for the complementation. A new M-MuLV HHCC construct, CD303, which has only the complete HHCC domain and no common region with the ND105, could complement ND105 for strand transfer reaction. This nonoverlapping complementation indi-cates that (i) the N-terminal region of M-MuLV IN containing the HHCC domain is a structurally independent domain and (ii) this HHCC domain is able to function intransto comple-ment the core and C terminus. The N-terminal amino acids 1 to 105 behaved like a dimer, indicating that the HHCC domain interacts with itself as well as other parts of the IN (16). The ability of CD232 to assist the 39processing reaction at a lower protein level than CD303 implies that the region of overlap could provide an additional protein-protein interaction.
Zinc finger motifs have been found in many of the transcrip-tion factors and DNA binding proteins (1, 2). The spacing and positions of the histidine and cysteines within the highly con-served HHCC domain of retroviral IN are unique and can form a novel zinc finger-like motif. To date, the HHCC domain is directly related only to protein multimerization; however, there is no direct evidence that the HHCC domain of retroviral IN binds DNA (45, 55). Though HIV-1 IN has been shown to structurally bind zinc at the ratio of 1:1 through the HHCC domain (8, 9, 32), zinc was found recently to stimulate
mag-FIG. 2. Complementation of ND105 by nonoverlapping HHCC domain protein. (A) Complementation of ND105 by CD232/CD303 for strand transfer reaction. Lane 1, control buffer incubation; lane 2, 10 pmol of ND105; lanes 3 and 4, 10 and 40 pmol of WT IN, respectively; lanes 5 to 9, 10 pmol of ND105 plus CD232; the ratio of CD232 to ND105 is indicated at the top of the panel; lanes 10 to 14, 10 pmol of ND105 plus CD303; the ratio of CD303 to ND105 is indicated at the top of the panel. (B) Complementation of ND105 by CD232/CD303 for 39processing reaction. The lanes are the same as in panel A. Both CD232 and CD303 were renatured in the presence of EDTA; the reaction buffer contained 10 mM MnCl2.
on November 9, 2019 by guest
http://jvi.asm.org/
[image:4.612.64.543.71.252.2]nesium-dependent activities including strand transfer and 39
processing (48, 68). The results from the studies reported here demonstrate that incorporation of zinc into the HHCC domain of MuLV IN could stimulate the catalytic activities of
[image:5.612.62.537.70.609.2]M-MuLV IN, indicating that zinc binding is functionally impor-tant and biologically relevant. Zinc did not stimulate the over-all strand transfer activity of M-MuLV IN, as for HIV IN. Instead, the range of target sites selected was limited in the
FIG. 3. Zinc stimulates CD303’s ability to complement ND105. (A) Complementation of ND105 by CD303/EDTA or CD303/zinc for strand transfer reaction. Lane 1, control buffer incubation; lane 2, 10 pmol of ND105; lanes 3 and 4, 10 and 40 pmol of WT IN, respectively; lanes 5 to 9, 10 pmol of ND105 plus CD303/EDTA; lanes 10 to 14, 10 pmol of ND105 plus CD303/zinc. Ratios of CD303/zinc or CD303/EDTA to ND105 are indicated at the top of each panel. Lanes in panels B to D are the same as in panel A. p, difference in strand transfer products. (B) Complementation of ND105 by CD303/EDTA or CD303/zinc for 39processing reaction. (C) Complementation of ND105 by CD303/EDTA or CD303/zinc for coordinated disintegration reaction with tailed crossbone substrates. (D) Complementation of ND105 by CD303/EDTA or CD303/zinc for coordinated disintegration reaction with untailed crossbone substrate. CD303 was renatured in zinc or EDTA as indicated at the top of each panel; all reaction buffers contained MnCl2as indicated in Materials and Methods. SD, single disintegration product; DD, double disintegration product.
on November 9, 2019 by guest
http://jvi.asm.org/
presence of a zinc-coordinated HHCC region. This increased stringency may reflect the tightness of the assembled complex; zinc binding may stabilize the HHCC domain and the core-C terminus interactions (16).
NEM modification has been used to probe the role of the HHCC domain in in vitro catalytic activities (22, 39). For M-MuLV IN, both the N-terminal cysteines within the HHCC domain and one central cysteine were sensitive to NEM alky-lation. Alkylation of the HHCC domain protein CD232 abol-ished most of its ability to complement ND105 for strand trans-fer and coordinated disintegration reactions (16). The results here demonstrate that zinc binding by the HHCC domain protected it from NEM modification, indicating that zinc co-ordination by the His and Cys residues is stable and that the two cysteine residues are fully occupied by zinc. The results can explain why there was residual activity in complementation between ND105 and NEM-modified CD232 for strand transfer reactions (16), where a trace amount of zinc coordinated by a small amount of CD232 could be resistant to NEM. HHCC constructs renatured in the presence of EDTA maintained a 0.1:1 molar ratio of zinc to protein.
Various evidence has indicated that viral INs act as oli-gomers. Structural studies of the catalytic domains of HIV IN and avian sarcoma virus IN and of the C terminus of HIV IN showed that they exist as dimers in solution (7, 18, 20, 50). Though an earlier report on the HHCC domain of HIV IN did not find dimerization of the HHCC domain in the presence of zinc (8), recent structural studies of the HIV-1 IN (11) showed that the HIV-1 IN amino acids 1 to 55 formed a dimer in solution. In contrast, the nuclear magnetic resonance structure of HIV-2 IN amino acids 1 to 55 was not a dimer in solution (21). Although the individual domains have been identified as dimers, one key question which remains unanswered is how the components assemble into an active multimer. Complementa-tion studies of M-MuLV and HIV-1 IN also suggest that IN functions as a multimer (22, 24, 38, 60). Recent studies start to reveal the role of the HHCC domain in the multimerization process of IN. Our results here demonstrate that the HHCC domain of M-MuLV IN formed a dimer in solution and that zinc was required for the dimerization. These results are con-sistent with that of HIV-1 IN. However, since the HHCC domain of M-MuLV IN is much larger than that of HIV IN (105 versus 55 amino acids), we cannot exclude the possibility that the additional sequences in M-MuLV IN are involved in the dimerization of the HHCC domain of M-MuLV IN.
Stud-FIG. 4. Zinc binding protects the HHCC domain from NEM modification. (A) CD303/zinc remains active for complementation after NEM treatment. A disintegration assay with oligonucleotide 7440 as substrate was used to test the susceptibility of IN mutants to NEM modification. Lane 1, control buffer incu-bation; lane 2, WT IN; lane 3, ND105; lane 4, complementation of ND105 and CD303/zinc; lane 5, complementation of ND105 and CD303/EDTA; lane 6, NEM-modified WT IN; lane 7, NEM-modified ND105; lane 8, complementation of ND105 and NEM-modified CD303/zinc; lane 9, complementation of ND105 and NEM-modified CD303/EDTA. All NEM-treated proteins are underlined. (B) Only CD303/EDTA can be labeled by fluorescent maleimide. CD303/zinc and CD303/EDTA were modified withN(1-pyrene) maleimide as described in Ma-terials and Methods, and the protein bands were analyzed for fluorescence (lanes 1 and 2) and by Coomassie blue staining (lanes 3 to 5). Lanes 1 and 3, CD303/ zinc; lanes 2 and 4, CD303/EDTA; lane 5, protein molecular mass markers.
FIG. 5. Fast protein liquid chromatography elution profile of the HHCC domain protein CD303/zinc. CD303/zinc was chromatographed on a Superose-12 column. The protein standard markers are as follows: 1, bovine serum albumin, 67 kDa; 2, ovalbumin, 43 kDa; 3, chymotrypsinogen A, 25 kDa; 4, RNase A, 13.7 kDa. The elution time of the protein standards is indicated on the panel. OD 280, optical density at 280 nm; AU, absorbance units.
on November 9, 2019 by guest
http://jvi.asm.org/
ies are currently under way to determine regions within the N-terminal 105 amino acids of M-MuLV IN required for dimerization. For HIV IN, it has been shown that zinc can promote tetramerization of HIV IN, while the predominant form of IN is a dimer without zinc (49, 68). In light of the zinc-dependent dimerization of the HHCC domain, our
cur-rent working model is that the HHCC domains bridge two sets of existing dimers formed through the central core and the C terminus, resulting in formation of an IN tetramer. The mech-anism of how the dimerization of the HHCC domain is cou-pled to tetramerization and which oligomeric form is active remain to be determined.
Previous studies have shown that the HHCC domain of HIV-1 IN can bind zinc at a 1:1 molar ratio and changes its conformation upon zinc binding (8, 32, 68). The stoichiometry of zinc against the M-MuLV IN HHCC domain was 1:1, indi-cating that there is no zinc in the interface of the zinc finger dimer. By the inductively coupled plasma mass spectroscopy method, no other metal was found substituting for zinc at stoichiometric levels (unpublished results). The CD study of the M-MuLV IN amino acids 1 to 105 indicated that the addition of zinc induced conformational changes with in-creased alpha-helical content, as found for HIV IN (8, 68). This indicates that at least some portion of the HHCC domain has to adopt helix structure in order for the histidine and cysteine residues to coordinate the zinc. However, there are some differences between M-MuLV IN amino acids 1 to 105 (CD303) and HIV IN amino acids 1 to 55. From our CD study, the M-MuLV IN amino acids 1 to 105 were well structured even in the absence of zinc, as indicated by the helical content of CD303/EDTA. The HIV IN amino acids 1 to 55 were highly disordered without zinc (8, 68). This could be due to the fact that M-MuLV IN has a much larger N terminus than does HIV IN. M-MuLV IN has 50 amino acid residues N-terminal to the HHCC zinc finger motif. These extra 50 residues may help the HHCC motif fold and stabilize the domain, even in the ab-sence of zinc.
The finding that CD303/EDTA in the absence of stoichio-metric zinc could complement ND105 for both strand transfer and 39 processing reactions raises the possibility that the N terminus of M-MuLV IN could be further divided into two separate domains: the N-terminal domain consisting of the first 50 amino acids and the zinc finger domain containing the conserved HHCC residues. Preliminary evidence defining these separate functions is presented in the comparison of the unimolecular (dumbbell in Fig. 4) and biomolecular disinte-gration (coordinated disintedisinte-gration) (Fig. 3D) reactions with untailed substrates. In the absence of the 59-ss tail, both reac-tions required an HHCC N-terminal domain. For the unimo-lecular reaction, either the zinc- or EDTA-renatured protein sufficed, and yet treatment with NEM inactivated the EDTA-renatured sample. The bimolecular disintegration requires substrate assembly. In the absence of the 59-ss tail, the HHCC construct in the absence of zinc could only minimally catalyze a single-end disintegration. The presence of zinc may therefore assist in the dimerization of the protein scaffold, thereby asso-ciating with the DNA substrates. The reactions greatly stimu-lated by zinc, 39 processing, foldback, and hydrolysis, may re-quire a tightly assembled complex, inducing strain in the substrates and assisting the nucleophilic attack to release the LTR. Future studies are aimed at determining the function of the N-terminal 50 residues.
ACKNOWLEDGMENTS
We give special thanks to Tara Shukla for help with atomic absorp-tion spectroscopy analysis.
This work was supported by American Cancer Society grant RPG-95-056-03-VM, NIH grant CA76545, NSF-INT-9408501 (travel grant), and FONDECYT 1980982.
REFERENCES
[image:7.612.53.292.75.446.2]1.Berg, J. M., and Y. Shi.1996. The galvanization of biology: a growing appreciation for the roles of zinc. Science271:1081–1085.
FIG. 6. CD spectra of the HHCC domain proteins. CD spectra of CD303/ zinc, CD303/EDTA, and CD303/zinc plus 5 mM EDTA at 5°C (A) and 37°C (B) are shown.
TABLE 1. Calculated content of secondary structures of the HHCC domaina
Temp Value (%)
Mutant ahelix bsheet Turn Random
5°C CD303/zinc 53.2 7.8 19.6 18.8 CD303/EDTA 33.4 16.9 26.3 23.0 CD303/zinc1EDTA 49.3 11.4 27.1 15.3
37°C CD303/zinc 52.7 15.0 19.5 16.1 CD303/EDTA 37.5 18.7 26.3 22.2 CD303/zinc1EDTA 33.8 17.6 24.9 23.7
aThe value is derived from the CD spectra with three different curve fitting
programs (see Materials and Methods for details). Only the calculations from the Selcon program are shown here. The other two programs gave similar results.
on November 9, 2019 by guest
http://jvi.asm.org/
[image:7.612.55.293.602.704.2]2.Berg, J. M.1990. Zinc finger domains: hypotheses and current knowledge. Annu. Rev. Biophys. Biophys. Chem.19:405–421.
3.Bradford, M. M.1976. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye bind-ing. Anal. Biochem.72:248–254.
4.Brahms, S., and J. Brahms.1980. Determination of protein secondary struc-ture in solution by vacuum ultraviolet circular dichroism. J. Mol. Biol.138:
149–178.
5.Brown, P. O.1997. Integration, p. 161–203.InJ. M. Coffin, S. H. Hughes, and H. E. Varmus (ed.), Retroviruses. Cold Spring Harbor Laboratory Press, Plainview, N.Y.
6.Brown, P. O., B. Bowerman, H. E. Varmus, and J. M. Bishop.1989. Retro-viral integration: structure of the initial covalent product and its precursor, and a role for the viral IN protein. Proc. Natl. Acad. Sci USA86:2525–2529. 7.Bujacz, G., M. Jaskolski, J. Alexandratos, A. Wlodawer, G. Merkel, R. A. Katz, and A. M. Skalka.1995. High-resolution structure of the catalytic domain of avian sarcoma virus integrase. J. Mol. Biol.255:333–346. 8.Burke, C. J., G. Sanyal, M. W. Bruner, J. A. Ryan, R. L. LaFemina, H. L.
Robbins, A. S. Zeft, C. R. Middaugh, and M. G. Cordingly.1992. Structural implications of spectroscopic characterization of a putative zinc finger pep-tide from HIV-1 integrase. J. Biol. Chem.267:9639–9644.
9.Bushman, F. D., A. Engelman, I. Palmer, P. Wingfield, and R. Craigie.1993. Domains of the integrase protein of human immunodeficiency virus type 1 responsible for polynucleotidyl transfer and zinc binding. Proc. Natl. Acad. Sci. USA90:3428–3432.
10. Bushman, F. D., and B. Wang.1994. Rous sarcoma virus integrase protein: mapping functions for catalysis and substrate binding. J. Virol.68:2215–2223. 11. Cai, M., R. Zheng, M. Caffrey, R. Craigie, G. M. Clore, and A. M. Gronen-born.1997. Solution structure of the N-terminal zinc binding domain of HIV-1 integrase. Nat. Struct. Biol.4:567–577.
12. Chow, S. A., and P. O. Brown.1994. Juxtaposition of two viral DNA ends in a bimolecular disintegration reaction mediated by multimers of human im-munodeficiency virus type 1 or murine leukemia virus integrase. J. Virol.
68:7869–7878.
13. Chow, S. A., K. A. Vincent, V. Ellison, and P. O. Brown.1992. Reversal of integration and DNA splicing mediated by integrase of human immunode-ficiency virus. Science255:723–726.
14. Craigie, R., T. Fujiwara, and F. Bushman.1990. The IN protein of Moloney murine leukemia virus processes the viral DNA ends and accomplishes their integration in vitro. Cell62:829–837.
15. Donzella, G. A., C. B. Jonsson, and M. J. Roth.1996. Coordinated disinte-gration reactions mediated by Moloney murine leukemia virus integrase. J. Virol.70:3909–3921.
16. Donzella, G. A., O. Leon, and M. J. Roth.1998. Implication of a central cysteine residue and the HHCC domain of Moloney murine leukemia virus integrase protein in functional multimerization. J. Virol.72:1691–1698. 17. Drelich, M., R. Wilhelm, and J. Mous.1992. Identification of amino acid
residues critical for endonuclease and integration activities of HIV-1 IN protein in vitro. Virology188:459–468.
18. Dyda, F., A. B. Hickman, T. M. Jenkins, A. Engelman, R. Craigie, and D. R. Davies.1994. Crystal structure of the catalytic domain of HIV integrase. Science266:1981–1986.
19. Edelhoch, H.1967. Spectroscopic determination of tryptophan and tyrosines in proteins. Biochemistry6:1948–1954.
20. Eijkelenboom, A. P., R. A. Lutzke, R. Boelens, R. H. Plasterk, R. Kaptein, and K. Hard.1995. The DNA-binding domain of HIV-1 integrase has an SH3-like fold. Nat. Struct. Biol.2:807–810.
21. Eijkelenboom, A. P. A. M., F. M. I. van den Ent, A. Vos, J. F. Doreleijers, K. Hard, T. D. Tullius, R. H. A. Plasterk, R. Kaptein, and R. Boelens.1997. The solution structure of the amino-terminal HHCC domain of HIV-2 integrase: a three-helix bundle stabilized by zinc. Curr. Biol.7:739–746.
22. Ellison, V., J. Gerton, K. A. Vincent, and P. O. Brown.1995. An essential interaction between distinct domains of HIV-1 integrase mediates assembly of active multimer. J. Biol. Chem.270:3320–3326.
23. Engelman, A., A. B. Hickman, and R. Craigie.1994. The core and carboxyl-terminal domains of the integrase protein of human immunodeficiency virus type 1 each contribute to nonspecific DNA binding. J. Virol.68:5911–5917. 24. Engelman, A., F. D. Bushman, and R. Craigie.1993. Identification of dis-crete functional domains of HIV-1 integrase and their organization within an active multimeric complex. EMBO J.12:3269–3275.
25. Engelman, A., G. Englund, J. M. Orenstein, M. A. Martin, and R. Craigie.
1995. Multiple effects of mutations in human immunodeficiency virus type 1 integrase in viral replication. J. Virol.69:2729–2736.
26. Engelman, A., and R. Craigie.1992. Identification of conserved amino acid residues critical for human immunodeficiency virus type 1 integrase function in vitro. J. Virol.66:6361–6369.
27. Engelman, A., K. Mizuuchi, and R. Craigie.1991. HIV-1 DNA integration: mechanism of viral DNA cleavage and DNA strand transfer. Cell67:1211– 1221.
28. Fasman, G. D.1989. Practical handbook of biochemistry and molecular biology. CRC Press, Inc., Boca Raton, Fla.
29. Fujiwara, T., and K. Mizuuchi.1988. Retroviral DNA integration: structure
of an integration intermediate. Cell54:497–504.
30. Gill, S. C., and P. H. von Hipple.1989. Calculation of protein extinction coefficients from amino acid sequence data. Anal. Biochem.182:319–326. 31. Greenfield, N. J.1996. Methods to estimate the formation of proteins and
polypeptides from circular dichroism data. Anal. Biochem.235:1–10. 32. Haugan, I. R., B. M. Nilsen, S. Worland, L. Olsen, and D. E. Helland.1995.
Characterization of the DNA-binding activity of HIV-1 integrase using a filter binding assay. Biochem. Biophys. Res. Commun.217:802–810. 33. Hazuda, D. J., A. L. Wolfe, J. C. Hastings, H. L. Robbins, P. L. Graham,
R. L. LaFemina, and E. A. Emini.1994. Viral long terminal repeat substrate binding characteristics of the human immunodeficiency virus type 1 inte-grase. J. Biol. Chem.269:3999–4004.
34. Heuer, T. S., and P. O. Brown.1997. Mapping features of HIV-1 integrase near selected sites on viral and target DNA molecules in an active enzyme-DNA complex by photo-cross-linking. Biochemistry36:10655–10665. 35. Heuer, T. S., and P. O. Brown.1998. Photo-cross-linking studies suggest a
model for the architecture of an active human immunodeficiency virus type-1 integrase-DNA complex. Biochemistry37:6667–6678.
36. Johnson, M. S., M. A. McClure, D. F. Feng, J. Gray, and R. F. Doolittle.
1986. Computer analysis of retroviral pol genes: assignment of enzymatic functions to specific sequences and homologies with nonviral enzymes. Proc. Natl. Acad. Sci. USA83:7648–7652.
37. Jones, K. S., J. Coleman, G. W. Merkel, T. M. Laue, and A. M. Skalka.1992. Retroviral integrase functions as a multimer and can turn over catalytically. J. Biol. Chem.287:16037–16040.
38. Jonsson, C. B., G. A. Donzella, E. Gaucan, C. M. Smith, and M. Roth.1996. Functional domains of Moloney murine leukemia virus integrase defined by mutation and complementation analysis. J. Virol.70:4585–4597.
39. Jonsson, C. B., and M. Roth.1993. Role of the His-Cys finger of Moloney murine leukemia virus integrase protein in integration and disintegration. J. Virol.67:5562–5571.
40. Jonsson, C. B., G. A. Donzella, and M. J. Roth.1993. Characterization of the forward and reverse integration reactions of the Moloney murine leukemia virus integrase protein purified from Escherichia coli. J. Biol. Chem.268:
1462–1469.
41. Kabsch, W., and C. Sander.1983. Dictionary of protein secondary structure: pattern recognition of hydrogen-bonded and geometrical features. Biopoly-mers22:2577–2637.
42. Katz, R. A., G. Merkel, and A. M. Skalka.1996. Targeting of retroviral integrase by fusion to a heterologous DNA binding domain: in vitro activities and incorporation of a fusion protein in viral particles. Virology217:178–190. 43. Katz, R. A., G. Merkel, J. Kulkosky, J. Leis, and A. Skalka.1990. The avian retroviral IN protein is both necessary and sufficient for integrative recom-bination in vitro. Cell63:87–95.
44. Katzman, M., R. A. Katz, A. M. Skalka, and J. Leis.1989. The avian retroviral integration protein cleaves the terminal sequences of linear viral DNA at the in vivo sites of integration. J. Virol.63:5319–5327.
45. Khan, E., J. P. G. Mack, R. A. Katz, J. Kulkosky, and A. M. Skalka.1991. Retroviral integrase domains: DNA binding and the recognition of LTR sequences. Nucleic Acids Res.19:851–860.
46. Kulkosky, J., K. S. Jones, R. A. Katz, J. P. G. Mack, and A. M. Skalka.1992. Residues critical for retroviral integrative recombination in a region that is highly conserved among retroviral/retrotransposon integrases and bacterial insertion sequence transposases. Mol. Cell. Biol.12:2331–2338.
47. Leavitt, A. D., L. Shiue, and H. E. Varmus.1993. Site directed mutagenesis of HIV-1 integrase demonstrates differential effects on integrase functions in vitro. J. Biol. Chem.268:2113–2119.
48. Lee, S. P., and M. K. Han.1996. Zinc stimulates Mg21-dependent 39
-processing activity of human immunodeficiency virus type 1 integrase in vitro. Biochemistry35:3837–3844.
49. Lee, S. P., J. Xiao, J. R. Knutson, M. S. Lewis, and M. K. Han.1997. Zn21
promotes the self-association of human immunodefiency virus type-1 inte-grase in vitro. Biochemistry36:173–180.
50.Lodi, P. J., J. A. Ernst, J. Kuszewski, A. B. Hickman, A. Engelman, R. Craigie, G. M. Clore, and A. M. Gronenborn.1995. Solution structure of the DNA binding domain of HIV-1 integrase. Biochemistry34:9826–9833. 51. Lutzke, R. A., C. Vink, and R. H. Plasterk.1994. Characterization of the
minimal DNA-binding domain of HIV integrase protein. Nucleic Acids Res.
22:4125–4131.
52. Mumm, S. R., and D. P. Grandgenett.1991. Defining nucleic acid-binding properties of avian retrovirus integrase by deletion analysis. J. Virol.65:
1160–1167.
53. Roth, M. J., P. Schwartzberg, N. Tanese, and S. P. Goff.1990. Analysis of mutations in the integration function of Moloney murine leukemia virus: effects on DNA binding and cutting. J. Virol.64:4709–4717.
54. Roth, M. J., P. L. Schwartzberg, and S. P. Goff.1989. Structure of the termini of DNA intermediates in the integration of retroviral DNA: dependence of IN function and terminal DNA sequence. Cell58:47–54.
55. Schauer, M., and A. Billich.1992. The N-terminal region of HIV-1 integrase is required for integration activity, but not for DNA binding. Biochem. Biophys. Res. Commun.185:874–880.
56. Shin, C., B. Taddeo, W. A. Haseltine, and C. M. Farnet.1994. Genetic
on November 9, 2019 by guest
http://jvi.asm.org/
analysis of the human immunodeficiency virus type 1 integrase protein. J. Virol.68:1633–1642.
57. Shinnick, T. M., R. A. Lerner, and J. G. Sutcliffe.1981. Nucleotide sequence of Moloney murine leukaemia virus. Nature293:543–548.
58. Sreerama, N., and R. W. Woody.1994. Poly(pro)II helices in globular pro-teins: identification and circular dichroic analysis. Biochemistry33:10022– 10025.
59. Taddeo, B., W. A. Haseltine, and C. M. Farnet.1994. Integrase mutants of human immunodeficiency virus type 1 with a specific defect in integration. J. Virol.68:8401–8405.
60. van Gent, D. C., C. Vink, A. A. Groeneger, and R. H. Plasterk.1993. Comple-mentation between HIV integrase proteins mutated in different domains. EMBO J.12:3261–3267.
61. van Gent, D. C., A. A. M. O. Groeneger, and R. H. A. Plasterk.1992. Mutational analysis of the integrase protein of human immunodeficiency virus type 2. Proc. Natl. Acad. Sci. USA89:9598–9602.
62. Vincent, K. A., V. Ellison, S. A. Chow, and P. O. Brown.1993. Character-ization of human immunodeficiency virus type 1 integrase expressed in Esch-erichia coliand analysis of variants with amino-terminal mutations. J. Virol.
67:425–437.
63. Vink, C., A. M. Oude Groeneger, and R. H. Plasterk.1993. Identification of
the catalytic and DNA-binding region of the human immunodeficiency virus type 1 integrase protein. Nucleic Acids Res.21:1419–1425.
64. Vink, C., R. A. P. Lutzke, and R. H. A. Plasterk.1994. Formation of a stable complex between the human immunodeficiency virus integrase protein and viral DNA. Nucleic Acids Res.22:4103–4110.
65. Wiskerchen, M., and M. A. Muesing.1995. Human immunodeficiency virus type 1 integrase: effects of mutations on viral ability to integrate, direct viral gene expression from unintegrated viral DNA templates, and sustain viral propagation in primary cells. J. Virol.69:376–386.
66. Woerner, A. M., and C. J. Marcus-Sekura.1993. Characterization of a DNA binding domain in the C-terminus of HIV-1 integrase by deletion mutagen-esis. Nucleic Acids Res.21:3507–3511.
67. Woerner, A. M., M. Klutch, J. G. Levin, and C. J. Marcus-Sekura.1992. Localization of DNA binding activity of HIV-1 integrase to the C-terminal half of the protein. AIDS Res. Hum. Retroviruses8:297–304.
68. Zheng, R., T. M. Jenkins, and R. Craigie.1996. Zinc folds the N-terminal domain of HIV-1 integrase, promotes multimerization, and enhances cata-lytic activity. Proc. Natl. Acad. Sci. USA93:13659–13664.
69. Zhou, X., K. F. Benson, H. R. Ashar, and K. Chada.1995. Mutation respon-sible for the mouse pygmy phenotype in the developmentally regulated factor HMG1-C. Nature376:771–774.