0022-538X/95/$04.0010
Copyright 1995, American Society for Microbiology
Human Herpesvirus 6B Origin-Binding Protein: DNA-Binding
Domain and Consensus Binding Sequence
NAOKI INOUE1*
ANDPHILIP E. PELLETT2
National Institute of Health, Tokyo 162, Japan,1and Centers for Disease Control and Prevention, Atlanta, Georgia 303332
Received 9 December 1994/Accepted 21 April 1995
We previously demonstrated by a DNA-binding assay that the human herpesvirus 6B (HHV-6B) replication origin has a structure similar to those of alphaherpesviruses, although the HHV-6B and herpes simplex virus type 1 (HSV-1) origin-binding proteins (OBPs) and origins are not interchangeable. Here we describe addi-tional properties of the interaction between HHV-6B OBP and the HHV-6B origin. Competitive electrophoretic mobility shift assays (EMSAs) with DNA duplexes containing single-base alterations allowed deduction of a consensus DNA sequence for HHV-6B-specific OBP binding, YGWYCWCCY, where Y is T or C and W is T or A, while that for HSV-1-specific binding was reported to be YGYTCGCACT. By EMSA, the HHV-6B OBP DNA-binding domain was mapped to a segment containing amino acids 482 to 770. However, in Southwestern (protein-DNA) blotting, the region sufficient for the DNA binding encompassed only amino acids 657 to 770. Similarly, Southwestern blotting showed that amino acids 689 to 851 of HSV-1 OBP had HSV-1 origin-binding activity, although this region was insufficient for origin binding in the EMSA. Although the longer DNA-binding domains identified by EMSA have marginal overall homology among HHV-6B and alphaherpesvirus OBP homologs, the smaller regions sufficient for the binding observed by Southwestern blotting have signif-icant similarity. From these results, we propose a hypothesis that the DNA-binding domain of herpesvirus OBPs consists of two subdomains, one containing a conserved motif that contacts DNA directly, and another, less well conserved, that may modulate either the conformation or accessibility of the binding domain.
The genomes of human herpesvirus 6 variants A and B (HHV-6A and HHV-6B) consist of an approximately 140-kb unique region and a pair of 10- to 13-kb directly repeated terminal elements, DRL and DRR. HHV-6 nucleotide and encoded protein sequences indicate that these viruses are ge-netically collinear with human cytomegalovirus (HCMV) over much of their genomes, but also that there are significant differences with HCMV with the respect to genome size, G1C content, and the existence of unique genes. On the basis of this information, HHV-6 is classified in the betaherpesvirus sub-family (22, 23, 27, 28; reviewed in references 15 and 24).
An HHV-6B open reading frame (ORF), CH6R, encodes a homolog (OBPH6B) of the herpes simplex virus type 1 (HSV-1) origin-binding protein (OBPH1) encoded by the HSV-1 UL9 gene (5). We previously demonstrated that OBPH6Bbinds to two sites, OBP-1 and OBP-2, in the upstream region of the HHV-6B major DNA-binding protein homolog that is likely to serve as an origin of lytic replication (oriLyt), based on its genomic location and sequence properties (16). The region containing the OBPH6Bbinding sites has sequences and overall arrangements similar to those of the replication origins of several alphaherpesviruses (16). This region can support tran-sient plasmid replication in HHV-6-infected cells (8). Muta-tions in the OBP-1 sequence that abolish binding to OBPH6B do not support transient replication (9). Moreover, some lab-oratory-passaged virus stocks have undergone gene amplifica-tion in the oriLyt region (29), and a plasmid containing mul-tiple copies of the oriLyt region replicates with enhanced
efficiency in transient replication assays (9). These results sug-gest that OBPH6B functions in a manner analogous to the alphaherpesvirus OBPs and that initiation of HHV-6B DNA replication may resemble that of alphaherpesviruses.
OBPH1 has helicase, ATPase, and DNA-binding activities (2, 3, 10, 12). A series of conserved elements in the amino-terminal half of the protein are involved in the helicase and ATPase activities; the carboxyl-terminal region, encompassing amino acids 563 to 832, is responsible for HSV-1 origin-bind-ing activity (6, 21, 31). The product of varicella-zoster virus (VZV) gene 51, the VZV OBP homolog (OBPVZV), recog-nizes a binding site within the VZV origin of replication that is identical to 11 bp of the OBPH1-binding site box I sequence (30), and the OBPVZVorigin-binding domain is located in the carboxyl-terminal one-third of the protein (4, 30). In earlier work, we mapped the DNA-binding domain of OBPH6Bwithin its carboxyl 403 amino acids (16).
Because interaction between OBP and the replication origin is hypothesized to be required for initiation of viral DNA replication (9, 19, 30), characterization of both the cis- and trans-acting components is essential for understanding the mo-lecular aspects of viral replication. Although the OBPH6B -binding sites OBP-1 and OBP-2 each lie within 23-bp segments that contain sequences homologous to OBPH1-binding site box I, OBPH1and OBPH6Bdid not bind to the binding sites of their counterpart in electrophoretic mobility shift assays (EMSAs) (16). This result suggests that both the binding sequences and OBP-binding specificity are virus specific. In this study, we have more thoroughly studied the properties of the oriLyt-OBP interaction. We identified a consensus sequence for OBPH6B-binding and more precisely defined the DNA-binding domain of OBPH6B. Elements that appear to determine the DNA-binding specificity between HHV-6B and HSV-1 and * Corresponding author. Mailing address: Dept. Virology I,
Na-tional Institute of Health, 1-23-1 Toyama, Shinjuku-ku, Tokyo 162, Japan. Phone: 81-3-5285-1111. Fax: 81-3-5285-1150. Electronic mail address: [email protected].
4619
on November 9, 2019 by guest
http://jvi.asm.org/
glutathione-S-transferase (GST)-OBPH6Bfusion protein. The resulting plasmids were designated pH6Z-4061, -4071, -4063, -4064, -4068, -4069, and -4070, re-spectively. The EcoRI-NotI fragment of pH6Z-4009 was cloned between the
EcoRI and NotI sites of pGEX-5X-3 to construct pH6Z-4062. pH6Z-4062 was
digested with EcoT22I and NotI, treated with T4 DNA polymerase, and circu-larized to delete the region between the EcoT22I and NotI sites. The resulting plasmid was designated pH6Z-4066. The BamHI-XhoI fragment of pH6Z-4061 was cloned between the BamHI and XhoI sites of pGEX-5X-3 to create pH6Z-4065. The OBPH6Bregions in these plasmids are depicted in Fig. 4.
(iii) pBS-TATA.pBluescript II KS(1) (Stratagene, La Jolla, Calif.) was di-gested with SacI and BamHI and ligated with a linker containing an ATG sequence,
59-CTATAAATATGGG-39 39-TCGAGATATTTATACCCCTAG-59
The sequence between the SacI site and the ATG sequence of pBS-TATA is identical to the sequence between the SacI site and the first ATG sequence of the truncated OBPH6BORF in pH6Z-4009.
(iv) Plasmids for in vitro expression of truncated OBPH6B.pH6Z-4062, -4063, -4064, -4068, and -4071 DNAs were digested with NotI, treated with T4 DNA polymerase, and then digested with BamHI. The BamHI sites are located 9 to 17 bp upstream of the HHV-6B-derived regions in these plasmids. The BamHI-NotI fragments were cloned between the BamHI and EcoRV sites of pBS-TATA. The resulting plasmids were designated pH6Z-4012, -4013, -4014, -4018, and -4011. pH6Z-4009 was digested with XhoI and NotI, and a linker containing a stop codon,
59-TCGAGATAGCT-39 39-CTATCGACCGG-59
was inserted between the sites, resulting in pH6Z-4015.
pH6Z-4009 was digested with EcoT22I and NotI, and a linker containing a stop codon,
59-TAGCTAG-39 39-ACGTATCGATCCCGG-59
was cloned between the sites, resulting in pH6Z-4016.
(v) pH1F-4461 and pH1F-4462.pRB132 (a gift of B. Roizman, University of Chicago) contains the HSV-1 strain F BamHI C fragment (26), which contains a part of the UL9 ORF encoding the carboxyl domain of OBPH1. The BamHI-NotI fragment of pRB132 was subcloned between the BamHI and NotI sites of pBluescript II KS(1) to construct pH1F-4401. The EcoRI-NotI fragment of pH1F-4401 was cloned between the EcoRI and NotI sites of pGEX-5X-1 to construct pH1F-4461. pH1F-4401 was digested with BstBI, treated with T4 DNA polymerase, and then digested with NotI. The BstBI-NotI fragment was cloned between the SmaI and NotI sites of pGEX-5X-1, resulting in 4462. pH1F-4461 and -4462 encode GST fusions with OBPH1amino acids 537 to 851 and 689 to 851, respectively.
Expression of GST-OBPH6Band GST-OBPH1fusion proteins inEscherichia
coli.DH5acells containing plasmids for GST-OBP fusion proteins were cultured at 378C. At a cell density of 108
/ml, isopropyl-b-D-thiogalactopyranoside was added to the culture to a final concentration of 0.1 mM. The cells were cultured at 308C for 90 min and then harvested. Pelleted cells were suspended in 1/40 the original volume of buffer (20 mM Tris-HCl [pH 7.5], 150 mM NaCl, 1 mM EDTA) and sonicated to reduce viscosity. In the case of OBPH6B, urea was added to the lysate to a final concentration of 1.0 M. The lysate was fractionated by centrifugation (50,0003g, 30 min) into a soluble extract and an insoluble
fraction. The soluble extract was used for EMSA. The insoluble pellet was suspended with sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE) loading buffer and used for Southwestern (protein-DNA) blotting.
DNA-binding assays.Oligonucleotides were synthesized in DNA synthesizers (Oligo 1000, Beckman Instruments, Fullerton, Calif.; model 380, Applied
Bio-systems, Foster City, Calif.).32
P-labeled DNA duplex probes, 31-bp 1C contain-ing the HHV-6B OBP-1 sequence (Table 1) and 36-bp UL and 26-bp UL2 containing the HSV-1 ori box I sequence (17/18 and 7/8 in reference 11, respec-tively) were used in EMSA and Southwestern blotting assays. To demonstrate the specificity of DNA binding (Fig. 6), labeled single-stranded and double-stranded probes were prepared by 59-end labeling of 19-nucleotide (nt)-long oligonucleotides with T4 polynucleotide kinase and by annealing the labeled
3-17 *************** 15 10
3-15 ************* 13 10
WT ******************* 19 (100)
MXE AAG**************** 19 100
MYC *C***************** 19 100
MZA **A**************** 19 100
MZC **C**************** 19 100
M1G ***G*************** 19 65
M1A ***A*************** 19 55
M1T ***T*************** 19 95
M2A ****A************** 19 70
M2T ****T************** 19 5
M2C ****C************** 19 30
M3G *****G************* 19 25
M3A *****A************* 19 90
M3C *****C************* 19 25
M4G ******G************ 19 30
M4A ******A************ 19 40
M4C ******C************ 19 100
M5G *******G*********** 19 10
M5A *******A*********** 19 0
M5T *******T*********** 19 5
M6G ********G********** 19 10
M6A ********A********** 19 100
M6C ********C********** 19 20
M7G *********G********* 19 0
M7A *********A********* 19 20
M7T *********T********* 19 0
M8G **********G******** 19 0
M8A **********A******** 19 10
M8T **********T******** 19 30
M9G ***********G******* 19 45
M9A ***********A******* 19 5
M9T ***********T******* 19 100
M10T ************T****** 19 85
M10A ************A****** 19 75
M11G *************G***** 19 50
M11C *************C***** 19 80
M12T **************T**** 19 35
M12D **************AA*** 19 70
M13T ***************T*** 19 80
M14G ****************G** 19 70
M14D ****************GA* 19 10
a
The 13-bp core OBP-1 sequence is underlined.
on November 9, 2019 by guest
[image:2.612.315.553.83.642.2]strand with its unlabeled complement, respectively. EMSA and Southwestern blotting were done as described before (16) except that the following binding buffer was used in the blotting reactions: 20 mM Tris-HCl (pH 7.0), 1 mM EDTA, 50 mM NaCl, 0.01% Nonidet P-40, 1mg of poly(dI-dC)poly(dI-dC) per ml, 0.2mg of salmon testes DNA per ml, and 1 ng of32P-labeled DNA probe per ml.
Immunoblotting.Antiserum against the A9-3 oligopeptide was used as the anti-OBPH6Bantiserum in immunoblotting (16). Anti-GST antibody (Pharma-cia) was used to detect GST-OBPH6Band GST-OBPH1fusion proteins.
RESULTS
[image:3.612.151.465.70.287.2]Identification of a consensus core OBPH6B-binding se-quence.We previously defined the OBPH6B-binding sequences OBP-1 and OBP-2 within 23-bp segments in the oriLyt region. In addition, we demonstrated that neither OBPH6Bnor OBPH1 recognized the binding sequence of its counterpart (16), sug-gesting that they recognize different sequences. To determine a consensus core sequence for OBPH6B-specific binding, two approaches were taken: (i) identification of the core-binding region boundaries by competitive EMSA with native sequence DNA duplexes of various lengths and (ii) identification of sequence requirements in the core-binding region by satura-tion mutagenesis (Table 1). Representative results of EMSAs with mutated DNA duplexes are shown in Fig. 1. The effects of competitor DNA duplexes were determined by measuring ra-dioactivity in the shifted bands and expressed as the percentage of binding inhibition relative to the binding activity of the wild-type DNA duplex. Thus, 0% inhibition means that a DNA duplex used as a competitor had no DNA-binding activity, and 100% inhibition means that the competitor DNA had full activity. The percent inhibition by each competitor is listed in Table 1.
The 17-bp DNA duplex Y-15 completely inhibited binding, and DNA duplexes containing the central 13-bp sequence 59 -CGTTCTCCCGTCC-39had partial activity (35 to 70%). This indicated that the 13-bp sequence contains the core-binding sequence. The sequence requirements for OBPH6B-binding within this region are summarized in Fig. 2. The results
indi-cate that (i) mutations introduced outside of positions 1 to 9 have little effect on binding, indicating that the OBPH6B core-binding region lies within these 9 bp and that the additional flanking sequences are required to stabilize the binding, and (ii) the consensus sequence in the core region is YGWYC WCCY, where Y is T or C and W is T or A. Substitutions in the M14D DNA duplex abolished DNA-binding activity, although these substitutions are outside of the 13-bp core region. This exceptional result might be explained by the fact that these substitutions produce a 4-nt-long sequence, CGAA, that is complementary to the other end of the oligonucleotide, result-ing in a reduction of proper double-stranded DNA duplex formation.
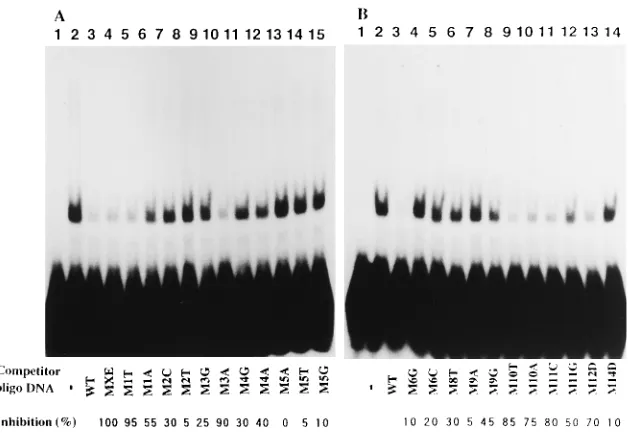
Strand-specific interactions identified by EMSA with het-eroduplex DNAs containing single mismatched base pairs.
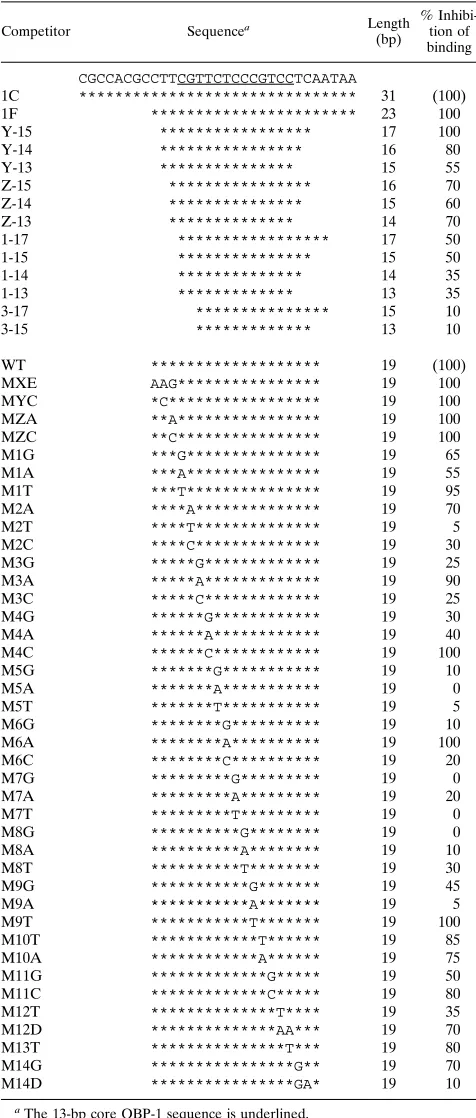
Substitutions of G with T at position 2 (M2T), C with A at position 5 (M5A), T with G at position 6 (M6G), C with G at FIG. 1. Competitive EMSA with mutated DNA duplexes.32
P-labeled DNA duplex 1C containing the OBP-1 sequence (Table 1) was reacted with purified GST-OBPH6Bfusion protein expressed from pH6Z-4061 (Fig. 4) (A, lanes 2 to 15; B, lanes 2 to 14) or without the protein (A and B, lanes 1), in the presence of 50-fold more unlabeled competitor DNA (Table 1) and then monitored for electrophoretic mobility shift. For quantitative analyses, radioactivity in the shifted bands was counted with a Fuji image analyzer BAS2000. The inhibition of binding of32P-1C DNA to OBPH6Bby each competitor DNA duplex is shown beneath the gel as the percent inhibition relative to the wild-type DNA duplex (WT). The other competitor DNA duplexes listed in Table 1 were analyzed similarly (data not shown).
FIG. 2. Effect of alterations in the 13-bp core OBP-1 sequence and the resulting consensus sequence of the OBPH6B-binding site. Open blocks indicate that the alteration did not affect the binding activity (.70% residual binding in Table 1). Shaded and solid blocks indicate that the alteration reduced the binding ability partially (20 to 70% residual binding) or severely (,20% residual binding), respectively. Base positions are numbered 59to 39relative to the 13-bp core sequence. The consensus OBPH1-binding sequence is shown beneath the consensus OBPH6B-binding sequence. OBP-2 indicates sequence differences that were not examined biochemically but are present in the wild-type OBP-2 se-quence.
on November 9, 2019 by guest
http://jvi.asm.org/
[image:3.612.319.550.557.645.2]position 7 (M7G), C with T at position 8 (M8T), and C with A at position 9 (M9A) reduced DNA binding to less than 30% of the wild-type sequence value (Fig. 1 and 2). Heteroduplex DNAs containing single mismatched base pairs at these
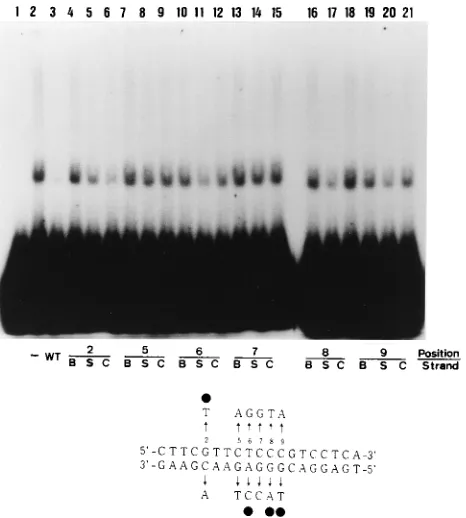
posi-GST fusions in E. coli. The plasmid construction procedures are described in Materials and Methods. Figure 4 depicts the domains of OBPH6B expressed in the fusion proteins. The proteins were separated into fractions soluble and insoluble in 1 M urea. Total protein and the insoluble fraction were sepa-rated by SDS-PAGE (Fig. 5B) and used in Southwestern blot-ting analyses (Fig. 5A). A32P-labeled DNA duplex containing the OBP-1 sequence was reacted with the proteins on the blots. All of the GST-OBPH6Bfusion proteins except those expressed from pH6Z-4070 and pH6Z-4065 bound to the OBP-1 se-quence. Therefore, the amino-terminal boundary of the DNA-binding domain is located between amino acids 657 and 701, and the carboxyl boundary lies between amino acids 744 and 770.
To examine the sequence specificity of OBP-1 DNA binding in Southwestern blotting, single-stranded and double-stranded DNAs containing wild-type OBP-1 sequence and those con-taining a C-to-A base alteration at position 5 (5A) in the sequence were used as probes (Fig. 6A to D). The intensities of the bands with single-stranded wild type, double-stranded 5A, and single-stranded 5A were 35, 20, and 2% of that with the double-stranded wild type probe, indicating that the South-western blotting detected both double-stranded DNA-specific FIG. 3. EMSA to measure binding activity of heteroduplexes.
[image:4.612.63.296.70.329.2]Nineteen-base-pair OBP-1 heteroduplex DNAs with a mutation on the 59-39sense (S) strand, those with a mutation on the 39-59complementary (C) strand, and DNA duplexes with a mutation on both (B) strands were used for competitive EMSA as described in the legend to Fig. 1. No protein was added in lane 1, and no competitor DNA (—) was added in lane 2. The sequence of the wild-type (WT) DNA duplex, base positions, and base alterations are described under the gel. Solid circles indicate that mutations on one strand of heteroduplexes abolished the DNA-binding activity more severely than did those on the other strand.
FIG. 4. Plasmids for expression of truncated OBPH6Bin in vitro translation and as GST-OBPH6Bfusion proteins, and summary of results from Fig. 5, 7, and 8. The procedures for construction of these plasmids are described in Materials and Methods. SacI (Sa), EcoRI (ER), BspHI (Bp), HincII (Hi), BsrBI (Br), BglII (Bg), PstI (Ps), SmaI (Sm), XhoI (Xh), EcoT22I (Ec), and NotI (No) sites are indicated. Domains A and B are explained in the text. Positive (1) and negative (2) reactions are indicated. NT, not tested; NA, no plasmid was available for the assay.
on November 9, 2019 by guest
and sequence-specific binding. However, the specificity in Southwestern blotting was weaker than that in the EMSA. To compare the optimal conditions in the Southwestern blotting assay with those in the EMSA, some reaction conditions were examined. Incubation at 48C but not at 378C decreased the binding (Fig. 6E and F). The optimal NaCl concentration for DNA binding in the Southwestern blotting assay was 50 mM, and the addition of MgCl2did not affect binding (Fig. 6G and H). These conditions are similar to the optima for EMSA (data not shown).
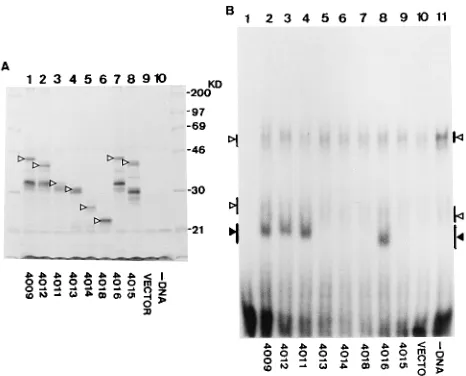
DNA-binding properties of GST-OBPH6B fusion proteins analyzed by EMSA.Soluble extracts of the GST-OBPH6B fu-sion proteins were reacted with32P-1C duplex DNA to exam-ine their DNA-binding activity by EMSA (Fig. 7B). GST-OBPH6B fusion proteins expressed from pH6Z-4061, -4062, and -4066 bound to the duplex, but those expressed from the other constructs had marginal binding activity. Therefore, the boundary of the amino terminus determined by EMSA is
lo-cated between amino acids 426 and 481, and the carboxyl boundary is located between amino acids 744 and 770. The amino-terminal boundary determined in this assay is different from that obtained by Southwestern blotting. The discrepancy cannot be explained by an absence of expressed protein in the soluble fraction used in EMSA, because (i) all GST-OBPH6B fusion proteins in the soluble fraction were detected by
[image:5.612.361.509.71.237.2]immu-FIG. 5. Southwestern blotting with GST-OBPH6Bfusion proteins. Total bac-terial lysate (lanes 1 to 3) and the fraction insoluble after extraction with 1 M urea (lanes 4 to 14) were separated in two SDS-PAGE gels. One gel (A) was used for Southwestern blotting as described previously (16). The other gel (B) was stained with Coomassie brilliant blue. The protein samples were obtained from bacteria containing the following plasmids: pH6Z-4061 (lane 4), -4062 (lanes 1 and 5), -4071 (lanes 2 and 6), -4063 (lane 7), -4064 (lane 8), -4068 (lane 9), -4069 (lane 10), -4070 (lane 11), -4066 (lane 12), and -4065 (lane 13) and pGEX-5X-1 (lanes 3 and 14). Open arrowheads indicate the GST-OBPH6Bfusion proteins and GST.
FIG. 6. Specificity of Southwestern blotting under various conditions. Insol-uble fractions prepared from lysates of bacteria containing the following plas-mids were analyzed by SDS-PAGE: lane 1, pH6Z-4062; lane 2, pH6Z-4071; lane 3, pH6Z-4069; lane 4, pGEX-5X-1. Binding buffer contained 50 mM NaCl (panels A to F), 200 mM NaCl (panel G), or 50 mM NaCl and 5 mM MgCl2 (panel H). Reaction temperatures were 258C (panels A to D and G), 378C (panel E), or 48C (panel F). Probes were wild-type (WT) double-stranded DNA (panels A, E, F, G, and H), WT single-stranded DNA (panel B), 5A double-stranded DNA (panel C), or 5A single-stranded DNA (panel D).
FIG. 7. EMSA with GST-OBPH6Bfusion proteins. (A) Detection of GST-OBPH6Bfusion proteins in soluble extracts from bacteria containing the indi-cated pH6Z plasmids (lanes 1 to 7) and pGEX-5X-1 (lane 8). GST-OBPH6B fusion proteins in the extracts were bound to glutathione-Sepharose 4B (Phar-macia), eluted with 1/40 volume of SDS-PAGE loading buffer, and then used for immunoblotting analyses with anti-GST antibody. The pH6Z-4070 and pGEX-5X-1 eluates were diluted 10- and 100-fold, respectively. Arrowheads indicate bands with the predicted molecular masses. (B) EMSAs with GST-OBPH6B fusion proteins were done as described in the legend to Fig. 1 without any added protein (lane 1) or with extracts from bacteria containing the indicated pH6Z plasmids (lanes 2 to 10) and pGEX-5X-1 (lane 11).
on November 9, 2019 by guest
http://jvi.asm.org/
[image:5.612.61.290.77.442.2] [image:5.612.319.549.439.634.2]noblotting with an GST antibody (Fig. 7A) and (ii) anti-OBPH6B antiserum that was obtained by immunizing rabbits with an oligopeptide deduced from the OBPH6Bsequence (16) also reacted with the GST-OBPH6Bproteins (data not shown). However, it is difficult to exclude the possibility that there is a difference in the quantity of available protein. In addition, there is a possibility that the 29-kDa GST domain of the fusion proteins inhibits the DNA binding of the shorter OBPH6B species by physical hindrance or by conformational alterations.
Determination of OBPH6BDNA-binding domain by EMSA with in vitro-translated OBPH6B. To eliminate the possible
artifacts caused by GST fusion, truncated OBPH6B proteins were expressed by in vitro transcription-translation coupled reactions. A series of plasmids for this purpose were con-structed as described in Materials and Methods. Figure 4 de-picts the truncated OBPH6Bregions. Only five to eight extra amino acid residues were attached to the truncated OBPH6B proteins at their amino-terminal end. Expression of the ex-pected proteins was confirmed by separation of35S-labeled in vitro reaction products by SDS-PAGE followed by fluorogra-phy (Fig. 8A). The DNA-binding activity of these truncated forms of OBPH6Bwas analyzed by EMSA (Fig. 8B). Although in vitro reaction lysate without added DNA contained proteins that gave mobility-shifted bands (open arrowheads), the OBPH6B-specific mobility shift (solid arrowhead) was observed with lysates programmed with pH6Z-4009, -4012, -4011, or -4016. In these experiments, the amino terminus of the DNA-binding domain of OBPH6Bmapped between amino acids 482 and 514, and the carboxyl terminus mapped between amino acids 744 and 770.
The experiments employing Southwestern blotting with GST fusion proteins, EMSA with GST fusion proteins, and EMSA
proteins.The conserved nature of domain B prompted exam-ination of the OBPH1 DNA-binding domain by EMSA and Southwestern blotting. Segments of OBPH1 containing resi-dues 537 to 851 and 689 to 851 were expressed as GST-OBPH1 fusion proteins from bacteria containing pH1F-4461 and pH1F-4462, respectively. Fusion protein expression was de-tected by staining the gel with Coomassie brilliant blue (data not shown) and by immunoblotting with an anti-GST antibody (Fig. 10A, lanes 4 and 5). The amount of GST-OBPH1fusion protein expressed from pH1F-4461 was less than that from pH1F-4462. Some smaller products from pH1F-4461 and pH1F-4462, which might be products of proteolysis, were present. As expected, the product from pH1F-4461 bound to the HSV-1 oriSbox I sequence in EMSA, but intact GST and the product from pH1F-4462 did not (Fig. 10D). However, the products from both pH1F-4461 and -4462 had DNA-binding activity in the Southwestern blotting assay (Fig. 10B). Signals obtained with the GST-OBPH1fusion protein from pH1F-4462 (lane 5) were significantly stronger than those obtained with GST-OBPH6Bfusion proteins (lanes 1 and 2) under conditions in which nearly equivalent amounts of the fusion proteins were applied to each lane. A probe containing the HHV-6 OBP-1 sequence did not bind to GST-OBPH1 fusion proteins (Fig. 10C). These results indicate that OBPH1does not bind to the HHV-6 OBP-1 sequence. The OBPH1region expressed from pH1F-4462 corresponds to domain B of OBPH6B(Fig. 9).
DISCUSSION
In this study, we characterized the OBPH6B binding se-quence and defined the DNA-binding domain of OBPH6B. In addition, we identified a conserved element in the DNA-bind-ing domains of OBP homologs and found that the OBPH1 conserved element retained HSV-1 origin-binding activity in Southwestern blotting.
Characterization of the binding sequence. Competitive EMSA with DNA duplexes showed that the 17-bp sequence of OBP-1, 59-TTCGTTCTCCCGTCCTC-39, was sufficient for full OBPH6B-binding activity. The central 13 bp of the 17-bp sequence had partial binding activity (30 to 60%), depend-ing on the length flankdepend-ing the sequence (Table 1). EMSA with mutated DNA duplexes identified a 9-bp segment internal to the 13-bp sequence within which sequence alterations re-duced binding activity. To obtain more direct evidence, we attempted exonuclease III footprinting with affinity-purified GST-OBPH6B protein. Although some experiments gave re-sults suggesting that the OBPH6Bbinding occurred within the 13 bp, it was difficult to obtain reproducibly sharp footprints (data not shown). This may have been due to the small amount of available protein, because OBPH6B expressed by recombi-FIG. 8. EMSA with in vitro lysates containing truncated OBPH6B. (A) In
vitro expression of truncated OBPH6B. Transcription-translation reaction mix-tures without added DNA (lane 10) or with plasmid DNA (lanes 1 to 9) were incubated in the presence of [35S]methionine (see Fig. 4 for the plasmid codes). Labeled protein was separated by SDS–12% PAGE and detected by fluorogra-phy. Arrowheads indicate bands corresponding to those predicted from the construction of plasmids. (B) EMSA without lysate (lane 1) or with the in vitro-translated lysate programmed with the following plasmids: pH6Z-4009 (lane 2), -4012 (lane 3), -4011 (lane 4), -4013 (lane 5), -4014 (lane 6), -4018 (lane 7), -4016 (lane 8), and -4015 (lane 9) and pBS-TATA (lane 10). Open arrow-heads indicate mobility shifts caused by proteins in the reticulocyte lysate, and solid arrowheads indicate OBPH6B-specific shifts. Because the electrophoresis was run longer to separate specific signals from nonspecific ones, unbound probe DNA nearly ran out from the bottom of the gel.
on November 9, 2019 by guest
[image:6.612.62.295.71.259.2]nant baculoviruses (16) and GST-, b-galactosidase-, and thi-oredoxin-OBPH6B fusion proteins expressed in E. coli are highly insoluble (data not shown). Although truncated OBPH6B proteins translated in vitro were soluble, the lysate containing the proteins did not provide conclusive footprints (data not shown).
Competitive EMSA with mutated DNA duplexes defined the consensus OBPH6B-binding site sequence as YGWYCWC CY (Table 1 and Fig. 1 and 2). Distinctive differences in the consensus HHV-6B and HSV-1 sequences occurred at position 6, W in place of G, and at position 8, C in place of A. These differences may determine the virus specificity of DNA repli-cation initiation.
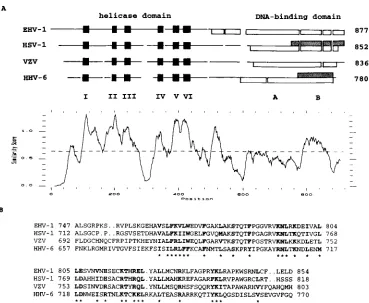
[image:7.612.122.491.68.371.2]The utility of DNA-binding assays with heteroduplex DNA was shown previously by Hazuda et al. (14). We applied the method to OBPH6B (Fig. 3). Although the experiment was done with a limited number of altered DNA duplexes, we observed that the heteroduplexes that contain single-base al-terations at position 2 on the complementary strand and at position 6, 8, or 9 on the sense strand competed with 32 P-labeled wild-type DNA duplex to bind OBPH6B, indicating that these single-base alterations were more tolerated than alter-ations either on the opposite strand or on both strands. This suggests that base-mediated interactions with OBPH6Bare lim-ited to one base of these base pairs. Although alteration of either base at positions 5 and 7 abolished the interaction, it is possible that the mismatched base pairs affect the DNA struc-ture. Substitutions to other bases at these positions will be FIG. 9. Sequence similarity in the OBP DNA-binding domain. (A) The amino acid sequences of the OBP homologs from EHV-1, HSV-1, VZV, and HHV-6B were aligned by the GCG PILEUP program (7), with a gap weight of 6 and a gap length weight of 0.3. The aligned sequences are depicted schematically. Helicase motifs I to VI (solid boxes) (21), DNA-binding domains determined by EMSA (open boxes) (4, 6, 18; this work), and DNA-binding domains determined by Southwestern blotting (shaded boxes, this work) are indicated. The similarity of the aligned sequences is plotted by the GCG PLOTSIMILARITY program (7), with a window size of 20. (B) Alignment of sequences corresponding to the DNA-binding domain of OBPH6Bsufficient for Southwestern blotting (domain B). Boldface letters and asterisks under the sequences indicate positions where residues in all four sequences have similarity scores higher than 0.5 in the modified Dayhoff table (13).
FIG. 10. DNA-binding domain of GST-OBPH1. GST-OBPH6Bfusion pro-teins expressed from pH6Z-4061 (lane 1), -4069 (lane 2), and -4070 (lane 3), GST-OBPH1fusion proteins expressed from pH1F-4461 (lane 4) and -4462 (lane 5), and GST (lane 6) were separated in three SDS–10% PAGE gels. One gel (A) was used for immunoblotting with an anti-GST antibody. The other gels were used for Southwestern blotting with the HSV-1-derived UL2 probe (B) and with the 1C probe (C). The conditions of the Southwestern blotting were the same as those for OBPH6B. Open and solid arrowheads indicate GST-OBPH6Band GST-OBPH1fusion proteins, respectively. (D) Soluble extracts from bacteria contain-ing pH1F-4461 (lane 2), pH1F-4462 (lane 3), and pGEX-5X-1 (lane 4) were used for EMSA with the UL probe. No protein was added in lane 1.
on November 9, 2019 by guest
http://jvi.asm.org/
[image:7.612.66.294.480.636.2]binding. The additional requirement for amino acids 426 to 481 in EMSA with GST-fusion proteins may be explained by an effect of the fused GST domain, for example, by physical hindrance caused by the GST portion. However, Southwestern blotting provided the surprising result that the region contain-ing amino acids 657 to 770 was sufficient for DNA bindcontain-ing (Fig. 5). The binding observed by Southwestern blotting was both double-stranded DNA specific and sequence specific, although less specific than that observed in EMSA; for example, a sin-gle-stranded oligonucleotide containing the wild-type binding sequence bound weakly (Fig. 6).
The DNA-binding domains of EHV-1, HSV-1, and VZV OBPs were defined within residues 500 to 887, 564 to 832, and 551 to 813, respectively, by EMSA (4, 6, 18). We compared the sequences of the DNA-binding domains and noticed that the homology of OBPH6Bwith the other three sequences is signif-icantly higher in domain B than elsewhere in the DNA-binding domain (Fig. 9A). Chen and Olivo (4) and Martin et al. (19) compared HSV-1 and VZV OBP sequences and noticed that the region corresponding to domain B is one of the five most conserved sequence blocks. Mutational analyses demonstrated the importance of the OBPH1region corresponding to domain B (1, 20). Because the DNA-binding sequences are very similar among HSV-1, VZV, EHV-1, and HHV-6B, this conserved domain of the OBPs may be the element directly responsible for DNA binding.
We propose the following hypothesis to explain our obser-vations. Domain B may directly contact DNA, and domain A may function to expose domain B to the outside of the mole-cule. Denaturation of the protein in the Southwestern blotting procedure may overcome the requirement for the exposure of domain B by domain A. The reduced specificity of domain B in the Southwestern blotting assay suggests that domain A is involved in increasing both the specificity and stability of the specific binding. This hypothesis is supported by the demon-stration that the OBPH1region corresponding to OBPH6B do-main B was sufficient for DNA binding in Southwestern blot-ting analysis (Fig. 10). An alternative, but less likely, possibility is that the 1 M urea-insoluble GST-OBP fusion proteins used for Southwestern blotting might differ from the soluble protein used for EMSA, for example, in phosphorylation. We were unable to test the materials in the complementary assays be-cause (i) the insoluble material used in Southwestern blotting is incompatible with EMSA, which is performed in solution, and (ii) the soluble material used in EMSA is difficult to produce in quantities sufficient for use in Southwestern blot-ting experiments.
To confirm our hypothesis, the following analyses will be useful. (i) Reactivities of monoclonal antibodies against do-main B with the series of truncated OBP proteins in solution
effect on HHV-6B DNA replication. In addition, the identifi-cation of domain B presents an inviting target for the devel-opment of molecules that interfere with the initiation of DNA replication.
ACKNOWLEDGMENTS
We thank Felicia Stamey, the CDC core facility, and Beckman Biolab (Tokyo) for synthesis of oligonucleotides. We also thank Ma-sato Tashiro and Steve Dewhurst for critical reading of the manuscript. We thank the Human Genome Center, Institute of Medical Science, The University of Tokyo for use of its UNIX computer system.
This work was supported in part by a grant from the Ministry of Education, Japan.
REFERENCES
1. Arbuckle, M. I., and N. D. Stow. 1993. A mutational analysis of the DNA-binding domain of the herpes simplex virus 1 UL9 protein. J. Gen. Virol.
74:1349–1355.
2. Boehmer, P. E., M. S. Dodson, and I. R. Lehman. 1993. The herpes simplex virus type-1 origin binding protein: DNA helicase activity. J. Biol. Chem.
268:1220–1225.
3. Bruckner, R. C., J. J. Crute, M. S. Dodson, and I. R. Lehman. 1991. The herpes simplex virus 1 origin binding protein: a DNA helicase. J. Biol. Chem.
267:2669–2674.
4. Chen, D., and P. D. Olivo. 1994. Expression of the varicella-zoster virus origin-binding protein and analysis of its site-specific DNA-binding proper-ties. J. Virol. 68:3841–3849.
5. Dambaugh, T. R., C. A. Greenamoyer, E. D. Anton, J. J. O’Brian, G.
Lindquester, and P. E. Pellett.Unpublished data (GenBank accession num-ber L14772).
6. Deb, S., and S. P. Deb. 1991. A 269-amino-acid segment with a pseudo-leucine zipper and a helix-turn-helix motif codes for the sequence-specific DNA-binding domain of herpes simplex virus type 1 origin-binding protein. J. Virol. 65:2829–2838.
7. Devereux, J., P. Haeberli, and O. Smithies. 1984. A comprehensive set of sequence analysis programs for the VAX. Nucleic Acids Res. 12:387–395. 8. Dewhurst, S., S. C. Dollard, P. E. Pellett, and T. R. Dambaugh. 1993.
Identification of a lytic-phase origin of DNA replication in human herpes-virus 6B strain Z29. J. Virol. 67:7680–7683.
9. Dewhurst, S., D. M. Krenitsky, and C. Dykes. 1994. Human herpesvirus 6B origin: sequence diversity, requirement for two binding sites for origin-binding protein, and enhanced replication from origin multimers. J. Virol.
68:6799–6803.
10. Dodson, M. S., and I. R. Lehman. 1993. The herpes simplex virus type 1 origin binding protein: DNA-dependent nucleoside triphosphatase activity. J. Biol. Chem. 268:1213–1219.
11. Elias, P., and I. R. Lehman. 1988. Interaction of origin binding protein with an origin of replication of herpes simplex virus 1. Proc. Natl. Acad. Sci. USA
85:2959–2963.
12. Fierer, D. S., and M. D. Challberg. 1992. Purification and characterization of UL9, the herpes simplex virus type 1 origin-binding protein. J. Virol. 66: 3986–3995.
13. Gribskov, M., and R. R. Burgess. 1986. Sigma factors from E. coli, B. subtilis, phage SP01, and phage T4 are homologous proteins. Nucleic Acids Res.
14:6745–6763.
14. Hazuda, D. J., H. C. Perry, A. M. Naylor, and W. L. McClements. 1991. Characterization of the herpes simplex virus origin binding protein interac-tion with oriS. J. Biol. Chem. 266:24621–24626.
15. Inoue, N., T. R. Dambaugh, and P. E. Pellett. 1994. Molecular biology of
on November 9, 2019 by guest
human herpesviruses 6A and 6B. Infect. Agent Dis. 2:343–360.
16. Inoue, N., T. R. Dambaugh, J. C. Rapp, and P. E. Pellett. 1994. Alphaher-pesvirus origin-binding protein homolog encoded by human herAlphaher-pesvirus 6B, a betaherpesvirus, binds to nucleotide sequences that are similar to ori regions of alphaherpesviruses. J. Virol. 68:4126–4136.
17. Inoue, N., S. Harada, T. Honma, T. Kitamura, K. Yanagi. 1991. The domain of Epstein-Barr virus nuclear antigen 1 essential for binding to oriP region has a sequence fitted for the hypothetical basic-helix-loop-helix structure. Virology 182:84–93.
18. Martin, D. W., and S. Deb. 1994. Cloning and expression of an equine herpesvirus 1 origin-binding protein. J. Virol. 68:3674–3681.
19. Martin, D. W., S. P. Deb, J. S. Klauer, and S. Deb. 1991. Analysis of the herpes simplex virus type 1 oriSsequence: mapping of functional domains. J. Virol. 65:4359–4369.
20. Martin, D. W., R. M. Mun˜oz, D. Oliver, M. A. Subler, and S. Deb.1994. Analysis of the DNA-binding domain of the HSV-1 origin-binding protein. Virology 198:71–80.
21. Martinez, R., L. Shao, and S. K. Weller. 1992. The conserved helicase motifs of the herpes simplex virus type 1 origin-binding protein UL9 are important for function. J. Virol. 66:6735–6746.
22. Nicholas, J. 1994. Nucleotide sequence analysis of a 21-kbp region of the genome of human herpesvirus-6 containing homologues of human cytomeg-alovirus major immediate-early and replication genes. Virology 204:738–750. 23. Nicholas, J., and M. E. D. Martin. 1994. Nucleotide sequence analysis of a 38.5-kilobase-pair region of the genome of human herpesvirus 6 encoding
human cytomegalovirus immediate-early gene homologs and transactivating functions. J. Virol. 68:597–610.
24. Pellett, P. E., and J. B. Black. Human herpesvirus 6. In B. N. Fields et al. (ed.), Virology, 3rd ed., in press. Raven Press, Ltd., New York.
25. Perry, H. C., D. J. Hazuda, and W. L. McClements. 1993. The DNA binding domain of herpes simplex virus type 1 origin binding protein is a transdomi-nant inhibitor of virus replication. Virology 193:73–79.
26. Post, L. E., A. J. Conley, E. S. Mocarski, and B. Roizman. 1980. Cloning of reiterated and nonreiterated herpes simplex virus 1 sequences as BamHI fragments. Proc. Natl. Acad. Sci. USA 77:4201–4205.
27. Sato, S., T. Yamamoto, Y. Isegawa, and K. Yamanishi. 1994. Identification of human herpesvirus 6 uracil-DNA glycosylase gene. J. Gen. Virol. 75:2349– 2354.
28. Schiewe, U., F. Neipel, D. Schreiner, and B. Fleckenstein. 1994. Structure and transcription of an immediate-early region in the human herpesvirus 6 genome. J. Virol. 68:2978–2985.
29. Stamey, F. R., G. Dominguez, J. B. Black, T. R. Dambaugh, and P. E. Pellett. 1995. Intragenomic linear amplification of human herpesvirus 6B oriLyt suggests acquisition of oriLyt by transposition. J. Virol. 69:589–596. 30. Stow, N. D., H. M. Weir, and E. C. Stow. 1990. Analysis of the binding sites
for the varicella-zoster virus gene 51 product within the viral origin of DNA replication. Virology 177:570–577.
31. Weir, H. M., J. M. Calder, and N. D. Stow. 1989. Binding of the herpes simplex virus type 1 UL9 gene product to an origin of viral DNA replication. Nucleic Acids Res. 17:1409–1425.