Distinct Mechanisms Regulate Exposure of Neutralizing Epitopes in
the V2 and V3 Loops of HIV-1 Envelope
Chitra Upadhyay,a,bLuzia M. Mayr,a,bJing Zhang,dRajnish Kumar,a,bMiroslaw K. Gorny,a,bArthur Nádas,cSusan Zolla-Pazner,a,b Catarina E. Hioea,b
Veterans Affairs New York Harbor Healthcare System, New York, New York, USAa
; Department of Pathologyb
and Institute of Environmental Medicine,c New York University School of Medicine, New York, New York, USA; Department of Microbiology, West China School of Preclinical and Forensic Medicine, Sichuan University, Chengdu, Sichuan, People’s Republic of Chinad
ABSTRACT
Broadly neutralizing antibodies targeting the HIV-1 envelope (Env) are key components for protection against HIV-1. However, many cross-reactive epitopes are often occluded. This study investigates the mechanisms contributing to the masking of V2i (variable loop V2 integrin) epitopes compared to the accessibility of V3 epitopes. V2i are conformation-dependent epitopes
en-compassing the integrin␣47-binding motif on the V1V2 loop of HIV-1 Env gp120. The V2i monoclonal antibodies (MAbs)
display extensive cross-reactivity with gp120 monomers from many subtypes but neutralize only few viruses, indicating V2i’s cryptic nature. First, we asked whether CD4-induced Env conformational changes affect V2i epitopes similarly to V3. CD4 treat-ment of BaL and JRFL pseudoviruses increased their neutralization sensitivity to V3 MAbs but not to the V2i MAbs. Second, the contribution of N-glycans in masking V2i versus V3 epitopes was evaluated by testing the neutralization of pseudoviruses pro-duced in the presence of a glycosidase inhibitor, kifunensine. Viruses grown in kifunensine were more sensitive to neutralization by V3 but not V2i MAbs. Finally, we evaluated the time-dependent dynamics of the V2i and V3 epitopes. Extending the time of virus-MAb interaction to 18 h before adding target cells increased virus neutralization by some V2i MAbs and all V3 MAbs tested. Consistent with this, V2i MAb binding to Env on the surface of transfected cells also increased in a time-dependent man-ner. Hence, V2i and V3 epitopes are highly dynamic, but distinct factors modulate the antibody accessibility of these epitopes. The study reveals the importance of the structural dynamics of V2i and V3 epitopes in determining HIV-1 neutralization by anti-bodies targeting these sites.
IMPORTANCE
Conserved neutralizing epitopes are present in the V1V2 and V3 regions of HIV-1 Env, but these epitopes are often occluded from Abs. This study reveals that distinct mechanisms contribute to the masking of V3 epitopes and V2i epitopes in the V1V2 domain. Importantly, V3 MAbs and some V2i MAbs display greater neutralization against relatively resistant HIV-1 isolates when the MAbs interact with the virus for a prolonged period of time. Given their highly immunogenic nature, V3 and V2i epitopes are valuable targets that would augment the efficacy of HIV vaccines.
T
he need for a safe and effective human immunodeficiencyvi-rus type 1 (HIV-1) vaccine is paramount, but despite intensive research, the scientific challenges facing its development remain formidable. The modest level of protection observed in the RV144 phase III vaccine trial indicated that protective immunity against
HIV-1 can be elicited through vaccination (1). High levels of
an-tibodies (Abs) specific for the variable loop 1 and 2 (V1V2) do-main of HIV Env gp120 in the vaccinees correlated with reduced
risk of HIV-1 acquisition (2–4). Follow-up sieve analyses of
break-through viruses showed increased vaccine efficacy against viruses with genetic signatures at two positions in V2, further supporting
the role of V2 in HIV-1 immunity (5). However, the exact
mech-anism(s) that contributed to a reduced risk of HIV-1 in RV144 vaccinees remains unknown, highlighting the importance of bet-ter understanding the antiviral functions of Abs against the V1V2 domain of HIV-1 gp120.
Monoclonal Abs (MAbs) targeting three different categories of epitopes in the V1V2 domain have been described. CH58 and CH59, isolated from an RV144 vaccinee, bind V2 peptides and monomeric gp120 proteins; their epitopes (designated V2p, for V2 peptide) are mapped to helical and loop structures modeled
from the C strand of V1V2 (6–8). MAbs, such as CAP256, CH01,
PG9, and PG16, recognize quaternary epitopes (V2q) in the V1V2 domain, which are preferentially expressed on the native trimeric
Env spike and comprised in part of N-glycans (9–11). Finally, a set
of seven MAbs used in our study recognize a third type of epitopes designated V2i (for V2 integrin). The V2i epitope depends on the proper conformation of the V1V2 domain and maps to the disor-dered V2 loop that connects the C and D strands, in particular, the
area overlapping with the integrin␣47-binding site, hence “V2i”
(6,12,13). The structure of this region is unknown, as it was not
resolved in the cryoelectron microscopy and crystal structures of
Env trimers and in the 1FD6-V1V2 scaffolded structures (14–16).
Notably, all seven V2i MAbs display extensive cross-reactivity in
Received18 July 2014 Accepted20 August 2014
Published ahead of print27 August 2014
Editor:F. Kirchhoff
Address correspondence to Catarina E. Hioe, [email protected].
C.U. and L.M.M. contributed equally to this work.
Copyright © 2014, American Society for Microbiology. All Rights Reserved.
doi:10.1128/JVI.02125-14
on November 7, 2019 by guest
http://jvi.asm.org/
binding monomeric gp120 proteins from tier 1, 2, and 3 viruses from different HIV-1 subtypes. However, V2i MAbs neutralize only a few tier 1 viruses and none of the tier 2 or tier 3 viruses in the
standard neutralization assay with TZM.bl target cells (17). These
data indicate that the V2i epitope is occluded from Ab recognition in resistant tier 2 and tier 3 viruses, reminiscent of cross-reactive neutralizing epitopes in the crown of variable loop 3 (V3). V3 epitopes are occluded in many viruses and pseudoviruses by the
V1V2 domain (18) and the N-glycans shrouding the HIV-1 Env
spike (19–21), and their exposure is augmented by CD4 binding to
Env (22,23) and when the virus is enriched for
high-mannose-type N-glycans (24,25). Nonetheless, factors regulating the
acces-sibility of V2i epitopes have not been described.
The present study was performed to elucidate whether similar mechanisms mask V2i and V3 epitopes. We evaluated the effects of CD4 binding, N-glycan composition, and kinetics of pseudovi-rus-MAb interaction on the neutralization of tier 1 and tier 2 viruses by V2i MAbs compared to V3 MAbs. The data demon-strate that distinct factors affect HIV-1 neutralization by V2i and V3 MAbs. Unlike V3 MAbs, neutralization by V2i MAbs was not affected by CD4 binding or Env enrichment with high-mannose glycans. However, like V3 MAb-mediated neutralization, neutral-ization by V2i MAbs was improved by increasing the virus-Ab preincubation time. Together with recent structural data showing the positions of V3 and V2i epitopes on the HIV-1 Env trimer, this study indicates that the exposure of V3 and V2i is regulated dif-ferently but both epitopes are subject to conformational masking, as they become more Ab accessible by prolonged Ab-virus inter-action. Although the conformational heterogeneity of V2 and V3 needs to be further investigated, these findings provide insight into the importance of the structural dynamics that govern the neutralizing activity of Abs against these sites and may be ex-ploited in designing vaccines against HIV.
MATERIALS AND METHODS
Cell lines, plasmids, and viruses.The TZM.bl cell line was obtained through the NIH AIDS Research and Reference Reagent Program (ARRRP), as contributed by J. Kappes and X. Wu (26). 293T/17 cells were obtained from the American Type Culture Collection (ATCC). Plasmids expressing Env of BaL.01 JRFL, REJO, and YU2 for generating pseudovi-ruses were obtained from the NIH ARRRP. Simian-human immunodefi-ciency viruses (SHIV) C.1157ipEL-p and HIV-1 92TH023 were also ob-tained from NIH ARRRP. CM244-OR Env was kindly provided by Sodsai Tovanabutra (MHRP and Henry M. Jackson Foundation for the Ad-vancement of Military Medicine). BaL.ec1 pseudovirus was kindly pro-vided by Victoria Polonis (Henry M. Jackson Foundation for the Ad-vancement of Military Medicine). BaL.ec1 was tested in CD4-dependent and time-dependent neutralization assays, whereas BaL.01 was used for kifunensine experiments. Plasmids pMX-gp160⌬cBaL-IRES-GFP and pMX-gp160⌬cJRFL-IRES-GFP (coexpressing gp160 with a deleted cyto-plasmic tail and green fluorescent protein [GFP] separated by an internal ribosome entry site [IRES]) (27,28) for cell binding assays were kindly provided by the Nussenzweig laboratory (The Rockefeller University).
Pseudoviruses were produced by cotransfecting 293T/17 cells with HIV-1rev- andenv-expressing plasmids and the pNL4 –3⌬env plasmid using jetPEI transfection reagent (Polyplus-transfect SA). Glycan-modi-fied pseudoviruses were generated as described above in the presence of 25
M kifunensine (Sigma). Supernatants were harvested after 48 h and clarified by centrifugation and 0.45-m filtration. Some virus stocks were concentrated with a 100-kDa Amicon filter (Millipore). Single-use ali-quots were stored at⫺80°C. Virus titration was performed on TZM.bl cells as described previously (29).
Human monoclonal antibodies.The V2i, V3, and control MAbs used in this study were produced in our laboratory as described previously (13,
17,30–34). MAbs b12 (provided by Dennis Burton and Carlos Barbas), 2G12 (provided by Hermann Katinger), and NIH45-46 (provided by Pamela Bjorkman) were obtained through the NIH ARRRP (35–37). V2q MAb PG9 was purchased from Polymun Scientific. V2p MAbs CH58 and CH59 were provided by Barton Haynes (Duke Human Vaccine Institute).
Neutralization assay. Virus neutralization was measured using a
-galactosidase-based assay (Promega) with TZM.bl target cells (38). Se-rial dilutions of MAbs were added to the virus in half-area 96-well plates (Costar) and incubated for designated time periods at 37°C. TZM.bl cells were then added along with DEAE-dextran (6.25g/ml; Sigma) and in-cubated for 48 h. Each condition was tested in duplicate or triplicate. Assay controls included replicate wells of TZM.bl cells alone (cell control) and TZM.bl cells with virus alone (virus control). Percent neutralization was determined on the basis of virus control under the specific assay condition. The virus input corresponded to titrated stocks yielding 150,000 to 200,000 relative luminescence units. For neutralization assays with soluble CD4 (sCD4), the virus was preincubated with recombinant human sCD4 (Progenics Pharmaceuticals) for 30 min at 37°C, before the addition of serially diluted MAbs. For temperature-dependent study, the virus-MAb mixture was incubated for 1 h at 37°C or 38.5°C prior to adding the TZM.bl cells. For experiments with different pHs, the pH of the medium was adjusted by the addition of HCl.
gp120 binding assay.The relative binding affinities of MAbs to gp120 from BaL and JRFL pseudoviruses was measured by a sandwich enzyme-linked immunosorbent assay (ELISA). The ELISA plates were coated with sheep anti-gp120 Ab (D7324, 2g/ml; Aalto BioReagents, Dublin, Ire-land), blocked with 2% bovine serum albumin (BSA) in phosphate-buff-ered saline (PBS), and incubated with 1% Triton X-100-treated pseudo-viruses. Serially diluted MAbs (0.01 to 10g/ml) were then added for 2 h, and the bound MAbs were detected with alkaline phosphatase-conjugated goat anti-human IgG Fc andp-nitrophenyl phosphate substrate.
Cell binding assay.As described previously (27), 293T/17 cells were grown to 70% confluence and transfected with pMX-gp160⌬cBaL -IRES-GFP or pMX-gp160⌬cJRFL-IRES-GFP (28), using FuGene HD (Roche). Two days later, the cells were washed with PBS, detached with trypsin-free cell-dissociation buffer (Invitrogen), and resuspended in PBS containing 2% fetal bovine serum and 2 mM EDTA. The transfected cells were stained on ice for the designated time periods with MAb (10g/ml), washed, and stained for 30 min with allophycocyanin (APC)-conjugated mouse anti-human IgG (BD Pharmingen). The cells were analyzed using a BD LSRFortessa.
Statistical analysis.Comparisons of virus neutralization and MAb binding were performed using S-Plus 6.1 (Insightful Corp.) or GraphPad Prism. Statistical analyses were performed on neutralization data that reachedⱖ50%.
RESULTS
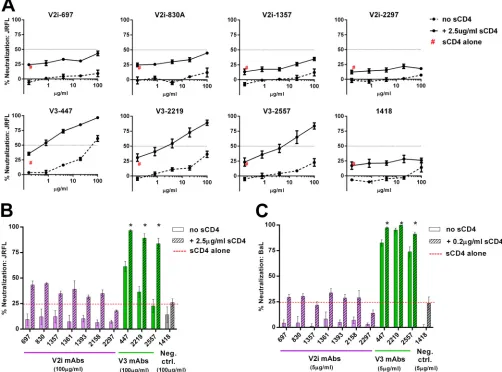
To investigate the mechanisms by which V2i and V3 epitopes are occluded from Ab recognition, we tested whether virus neutral-ization by V2i and V3 MAbs was influenced by the following three parameters: CD4 binding, N-glycan composition, and time. We compared the neutralization by seven V2i MAbs (697, 830A, 1357, 1361, 1393, 2158, and 2297) and three V3 MAbs (447, 2219, and 2557) against tier 1 and tier 2 viruses from different clades of HIV-1. MAb 1418, specific for human parvovirus B19, was in-cluded as a negative control. MAbs CH58 (V2p) CH59 (V2p), PG9 (V2q), 2G12, b12 (CD4-binding site [CD4bs]), and NIH45-46 (CD4bs) were also tested in some experiments for comparison.
CD4 engagement enhances HIV-1 neutralization by V3 but
not V2i MAbs.CD4 binding induces conformational changes in
HIV-1 Env that result in the exposure of otherwise inaccessible
regions, including cryptic Ab epitopes in the V3 loop (39–42).
on November 7, 2019 by guest
http://jvi.asm.org/
However, the influence of CD4 on V2i epitopes has not been in-vestigated. To address this gap in the field, we tested the capacity of V2i versus V3 MAbs to neutralize HIV-1 JRFL and BaL pseudovi-ruses that were pretreated with sCD4 or left untreated. No neu-tralization was detected with each of the seven V2i MAbs when
titrated against both untreated JRFL and untreated BaL (Fig. 1).
Upon pretreatment of these pseudoviruses with sCD4 at a fixed
concentration that caused⬃25% inhibition, neutralization by all
V2i MAbs remained below 50%, and no dose-dependent effect
was observed. Similar patterns were seen with JRFL and BaL (Fig.
1BandC). In contrast, sCD4 treatment significantly increased
JRFL neutralization by the V3 MAbs 447, 2219, and 2557 (P⬍
0.001). At the highest concentration tested (100g/ml), 84 to 97%
neutralization was reached by the V3 MAbs against JRFL in the presence of sCD4, while only 23 to 61% was achieved without
sCD4 (Fig. 1AandB). As expected, the increase in neutralization
against BaL, a tier 1 virus with relatively accessible V3 epitopes,
was less pronounced, albeit still significant (P⬍0.05) (Fig. 1C).
The control MAb 1418 did not show any neutralization in the presence or absence of sCD4 against either JRFL or BaL. These results indicate that, unlike that of V3, the accessibility of V2i epitopes is not modulated by CD4 interaction with HIV-1 Env.
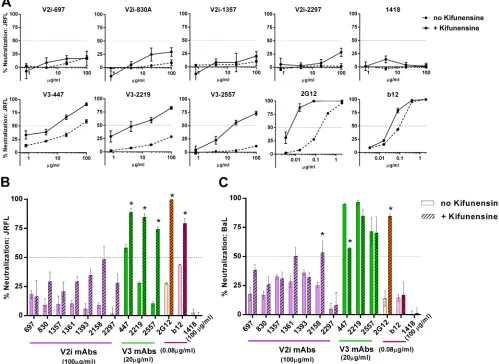
Changes in N-glycan sugar composition affect
neutraliza-tion by V3 MAbs but not V2i MAbs.The removal of N-glycans
from specific sites on HIV-1 Env enhances virus sensitivity to Abs
against V3 and other neutralizing epitopes on gp120 (23, 43).
Recent studies further demonstrate the influence of specific sugar
moieties of N-glycans on access to neutralizing V3 epitopes (24,
25). In particular, enrichment of high-mannose-type glycans
en-hanced virus neutralization by V3 Abs. To investigate whether V2i epitopes are similarly affected, we generated JRFL and BaL
pseu-doviruses in the presence of 25M kifunensine, a mannosidase
inhibitor that blocks the processing of complex-type N-glycans and enriches for the high-mannose type. The V2i MAbs displayed
FIG 1Influence of CD4 engagement on virus neutralization by V2i and V3 MAbs. (A) Neutralization of JRFL pseudovirus by V2i, V3, and control MAbs was tested following treatment with or without sCD4. Virus was pretreated with sCD4 at one designated concentration for 30 min at 37°C or left untreated and then incubated with serially diluted MAb for 1 h at 37°C. The mixture was added to TZM.bl target cells, and after 48 h, virus infection was measured based on
-galactosidase activity. (B, C) Comparison of neutralization by V2i, V3, and control MAbs in the presence or absence of sCD4 against JRFL and BaL.ec1. “sCD4 alone” denotes virus neutralization by sCD4 alone (no MAbs). Means and standard errors from two independent experiments (each in duplicate) are shown. Statistical analyses were performed on neutralization curves reachingⱖ50%. *,P⬍0.001 for JRFL (B) andP⬍0.05 for BaL (C), based on the two-way analysis of variance (ANOVA) test showing significant synergistic difference above the calculated sum of percent neutralization by titrated MAb plus sCD4.
Modulation of V2 versus V3 Epitopes
on November 7, 2019 by guest
http://jvi.asm.org/
[image:3.585.40.542.71.443.2]poor neutralizing activities against JRFL and BaL irrespective of
kifunensine treatment (Fig. 2AtoC). This contrasted with the
increased neutralization observed with all three V3 MAbs tested against JRFL produced in kifunensine-treated cells compared to
their neutralization of untreated JRFL (Fig. 2A andB). MAbs
2G12 and b12 were also tested in parallel as controls. Consistent
with previous reports (44), MAb 2G12, which recognizes a cluster
of N-glycans bearing the high-mannose type (37), showed greater
levels of neutralization against JRFL produced in the presence of kifunensine than against untreated JRFL. The irrelevant control MAb 1418 had no neutralizing activity against treated or un-treated viruses. In the case of BaL, kifunensine treatment rendered
the virus more resistant to neutralization by the V3 MAb 447 (Fig.
2C), whereas no pronounced effects were observed on
neutraliza-tion by 2219, 2557, and b12. However, kifunensine treatment
sig-nificantly increased BaL neutralization by the V2i MAb 2158 (P⬍
0.001), reaching up to 50% neutralization at the highest
concen-tration tested (100g/ml). As anticipated, BaL neutralization by
2G12 was also greatly enhanced. These results demonstrate that
V3 epitopes for MAbs 447, 2219, and 2557 are positively (in the case of JRFL) or negatively (in the case of BaL.01) modulated by the changes in N-glycan sugar composition, while V2i epitopes are not affected, except for 2158 against BaL. The results highlight the important role played by N-glycans in influencing V3 and V2i epitopes, beyond simple steric masking. Moreover, the differences observed between JRFL and BaL indicate that the impact of the N-glycan composition is dependent on the virus strain.
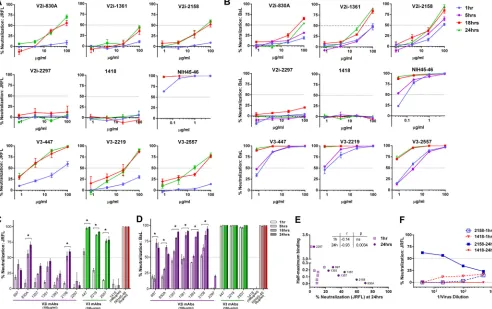
Prolonging the virus-MAb preincubation time increases
neutralization and binding by V2i and V3 MAbs.We next
inves-tigated the effects of the length of virus-MAb interaction before their exposure to TZM.bl target cells on the neutralizing activity of V2i versus V3 MAbs. Increasing the preincubation time signifi-cantly improved the neutralizing activity of several V2i MAbs
against both JRFL and BaL (Fig. 3). By prolonging the incubation
time from the standard 1 h to 18 h or 24 h,ⱖ50% neutralization
was reached with V2i MAbs 830A and 2158 against JRFL (Fig. 3A
andC). The V2i MAb 1361 also showed increased neutralization,
reaching nearly 50%. Neutralization by the other V2i MAbs also
FIG 2Changes in N-glycan sugar composition affect virus neutralization by V3 MAbs but not V2i MAbs. (A) V2i, V3, and control MAbs were titrated and tested for neutralizing activity against JRFL produced in the presence or absence of kifunensine. Virus and MAb were incubated for 1 h and then added to TZM.bl target cells. Virus infectivity was measured 48 h later based on-galactosidase activity. (B, C) Comparison of neutralization by V2i, V3, and control MAbs against JRFL or BaL.01 produced with or without kifunensine. Means and standard errors calculated from three different experiments (each in duplicate) are shown. *,P⬍
0.0001 for JRFL and BaL.01, based on the two-way ANOVA test comparing the neutralization curves of each MAb tested against virus produced with or without kifunensine.
on November 7, 2019 by guest
http://jvi.asm.org/
[image:4.585.40.539.72.436.2]increased but did not attain 50% (Fig. 3C). Only V2i MAb 2297 had no time-dependent neutralizing activity, similar to the con-trol MAb 1418; this V2i MAb had an exceptionally low binding
affinity for JRFL (Fig. 3E). Indeed, the levels of JRFL neutralization
achieved with the longer incubation time of 24 h but not the stan-dard 1-h incubation correlated with the relative affinities of the V2i MAbs as measured by half-maximal binding to JRFL gp120 (Fig. 3E). When tested against BaL, V2i MAbs displayed various
low levels of neutralization after 1 h of virus-MAb incubation (Fig.
3BandD). With prolonged incubation time, neutralization
in-creased significantly for all V2i MAbs except 2297. Interestingly, increased neutralization was observed after extending the
incuba-tion to 18 h but not to 5 h. With the incubaincuba-tion time ofⱖ18 h, V3
MAbs (447, 2219, and 2557) and the CD4bs MAb NIH45-46 also displayed greater neutralization against both JRFL and BaL, be-yond the levels observed with the standard 1-h incubation time.
To test whether differences in input virus infectivity account for the increased neutralization observed after 18 to 24 h versus 1 h of virus-MAb incubation, serially diluted JRFL was tested for
neutralization by a constant amount (100g/ml) of V2i MAb
2158 or control MAb 1418 following 1 h versus 24 h of virus-MAb incubation time. Untreated diluted virus was tested in parallel. No neutralization was observed with a 1-h incubation over the range
of virus input tested (Fig. 3F). In contrast, the 2158-mediated
neutralization observed after a 24-h incubation showed a recipro-cal relationship with the amount of virus input. These data indi-cate that the virus neutralization attained with a 24-h incubation time is dependent on both the MAb concentration and the virus dose, whereas the standard 1-h incubation time was not sufficient regardless of the virus dose and MAb concentration.
One likely reason for the need for longer virus-MAb preincu-bation to neutralize HIV-1 is that the V1V2 and V3 regions are highly dynamic on Env spikes, continually flickering from one conformation to another and rarely adopting the structures rec-ognized by the MAbs. To evaluate this idea, we asked whether the binding of V2i and V3 MAbs to Env spikes also occurred in a time-dependent manner. Therefore, 293T/17 cells were
trans-fected with plasmids expressing BaL or JRFL Env (27,28) for 48 h
and then incubated with V2i, V3, PG16, or control MAbs for var-ious periods of time. The binding of MAbs to the virus Env on the
FIG 3Time-dependent increase of virus neutralization by V2i and V3 MAbs. (A, B) Virus neutralization was measured after JRFL or BaL.ec1 was incubated with serially diluted MAbs for the designated period of time at 37°C prior to the addition of TZM-bl target cells. The CD4bs MAb NIH45-46 and the irrelevant control MAb 1418 were tested in parallel for comparison. (C, D) Comparison of neutralization by V2i, V3, CD4bs, and control MAbs against JRFL and BaL.ec1 after various times of virus-MAb preincubation. Means and standard errors from two to three experiments are shown. *,P⬍0.05 for V2i andP⬍0.001 for V3 MAbs, from the two-way ANOVA test comparing neutralization curves observed with virus-MAb preincubation of 1 h versus 18 h and 1 h versus 24 h. (E) JRFL neutralization achieved after a 24-h preincubation with V2i MAbs correlated with gp120 JRFL binding as measured by ELISA. Correlation was also observed with an 18-h incubation (P⫽0.033;r⫽ ⫺0.75) but not with a 1-h incubation (not shown). (F) JRFL neutralization after a 24-h incubation with V2i MAb displayed virus dose dependency, while no neutralization was achieved with a 1-h incubation regardless of the amount of input virus. Virus was titrated, treated with one fixed concentration of V2 MAb 2158 or control MAb 1418 (100g/ml) for 1 h or 24 h, and then added to the TZM.bl target cells. Virus infection was assessed 48 h later based on-galactosidase activity. At high virus dilutions yielding exceedingly low infectivity, some neutralization values reached below 0% and are denoted in the graph as 0%.
Modulation of V2 versus V3 Epitopes
on November 7, 2019 by guest
http://jvi.asm.org/
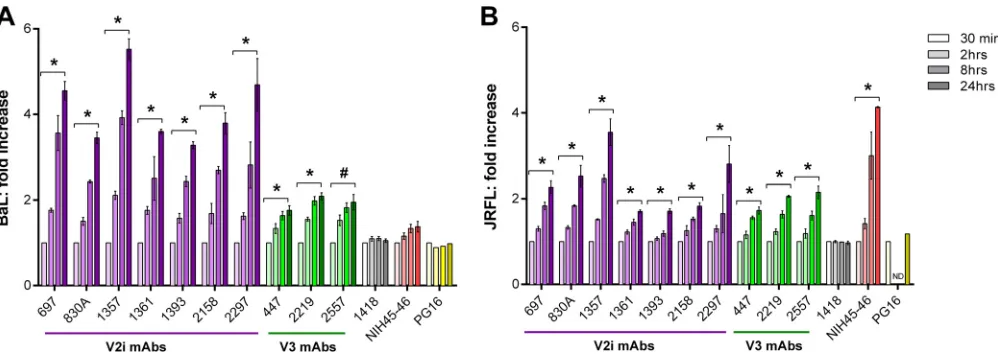
[image:5.585.46.538.70.379.2]cell surface was measured by flow cytometry (Fig. 4). Little bind-ing to BaL Env was observed for the V2i MAbs after 30 min, but a
significant linear increase was detected over time (Fig. 4A). After
24 h, the binding of all seven V2i MAbs increased 4-fold on aver-age when measured by mean fluorescence intensity (MFI) (aver-age MFI of 1,190 for the seven MAbs at 30 min versus MFI of 4,914 at 24 h). Higher levels of V3 than of V2i MAbs bound the BaL Env after only 30 min (average MFI, 6,369), and V3 MAb binding
increased ⬃1.9-fold after 24 h (Fig. 4A). The CD4bs MAb
NIH45-46 showed no time-dependent increase of binding to BaL Env, indicating that this highly potent neutralizing MAb attained maximal binding within 30 min (MFI of 1,512 at 30 min and MFI
of 1,894 at 24 h) (Fig. 4A). The increase in V2i MAb binding to
JRFL Env was less pronounced. Low levels of binding were ob-served after 30 min (mean MFI of 612), and after 24 h, a 2.25-fold
increase was detected (mean MFI of 1,467) (Fig. 4B). V3 MAbs
similarly showed an average 2-fold increase over 24 h. As with BaL-transfected cells, V3 MAbs bound JRFL-transfected cells at higher levels (mean MFI of 2,240 at 30 min and mean MFI of 4,428 at 24 h). The CD4bs MAb NIH45-46 displayed a time-dependent
4-fold increase in binding to JRFL Env (Fig. 4B). As a control to
verify the relative expression of trimeric Env spikes on the trans-fected cells over time, we also tested PG16 MAb, which recognizes a quaternary epitope on the Env trimer. The data show compara-ble binding of PG16 to BaL and JRFL after incubation for 30 min and up to 24 h, indicating that similar levels of properly folded trimeric spikes were expressed on the transfected cells throughout the assay period.
Two additional assay parameters were studied, pH and tem-perature. To study the effects of different pHs on virus neutraliza-tion, we first evaluated the infectivity of BaL upon incubation in medium of pH 4.2, 5.6, 6.6, or 7.4. The virus infectivity decreased
to⬍10% at pH 4.2 and 5.6 but remained unchanged at pH 6.6
compared to the infectivity at pH 7.4 (Fig. 5A), allowing us to
compare virus neutralization under the two latter conditions. BaL neutralization by V2i MAb 2158 and control MAb 1418 was as-sessed at pH 6.6 or pH 7.4 following 1 h or 5 h of virus-MAb
incubation. Lowering the pH to 6.6 had no significant effect on virus neutralization after 1 h or 5 h of incubation with the MAb (Fig. 5B). Similarly, when the temperature during virus-MAb in-cubation was raised from 37°C to 38.5°C, the neutralizing activi-ties of V2i or V3 MAbs against BaL and JRFL were comparable (Fig. 5C). Hence, lowering the pH to 6.6 and increasing the tem-perature to 38.5°C did not augment virus neutralization by V2i and V3 MAbs as was observed with prolonging the virus-MAb
incubation time toⱖ18 h.
Prolonging virus-MAb incubation enhances neutralization
of other HIV-1 isolates by V2i, V2p, V2q, and V3 MAbs.In
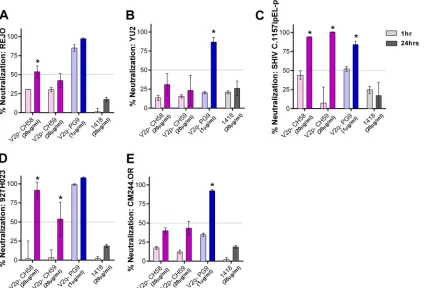
ad-dition to JRFL and BaL, we evaluated the neutralization of viruses from different clades, including REJO (clade B), YU2 (clade B), SHIV C.1157ipEL-p (clade C), 92TH023 (CRF_01AE), and CM244-OR (CRF_01AE). With the standard 1-h incubation, no neutralization was detected against REJO and YU2 by the V2i and
V3 MAbs tested (Fig. 6AandB). However, greater than 90%
neu-tralization was achieved when REJO was incubated for 24 h with
100g/ml of V2i MAbs 697, 1393, and 2158, while 79%
neutral-ization was reached with 1357 (Fig. 6A). Neutralization above
50% was also observed with these V2i MAbs at 20g/ml upon 24
h of virus-MAb incubation. Increased levels of neutralization after incubation for 24 h compared to the neutralization after 1 h were similarly obtained with V3 MAbs 2219 and 2557 but not with V3 MAb 447 and control MAb 1418. In the case of YU2,
neutraliza-tion reached 77% after 24 h of incubaneutraliza-tion with 100g/ml of V2i
MAb 697 but dropped to 41% with 20g/ml (Fig. 6B). The other
V2i MAbs also showed increased neutralization, but the levels
only reachedⱕ50%. Similarly, V3 MAbs 447 and 2219 but not
2557 exhibited greater neutralization, reaching 100% following 24 h of incubation. Increased neutralization was also observed with 24 h versus 1 h of incubation when V2i and V3 MAbs were tested
against SHIV C.1157ipEL-p (Fig. 6C) and HIV-1 92TH03 (Fig.
6D). In contrast, CM244.OR remained poorly neutralized (Fig.
6E), indicating that not all viruses become sensitive to
neutraliza-tion by V2i and V3 MAbs after the extended virus-MAb
incuba-FIG 4Time-dependent increase of Env spike binding by V2i and V3 MAbs. Relative levels of MAb binding to BaL (A) or JRFL (B) Env spikes on transfected 293T cells were measured by flow cytometry. Cells were transfected with gp160⌬cBaL-IRES-GFP for 48 h and treated with V2i, V3, or control MAbs at 4°C for various
times from 30 min to 24 h. MAb binding was detected with APC-conjugated anti-human IgG. Fold increase of MAb binding was calculated relative to mean fluorescence intensity measured at 30 min (normalized to 1). Means and standard errors from two to four independent experiments are shown. *,P⬍0.05 based on the one-tailttest comparing fold increase at 24 h versus 30 min; #,P⫽0.059; ND, 2 h and 8 h time points not tested for JRFL.
on November 7, 2019 by guest
http://jvi.asm.org/
[image:6.585.43.542.65.242.2]tion time. No neutralization was detected with the control MAb 1418 against any of the viruses tested.
To evaluate whether prolonging virus-MAb incubation had similar effects on MAbs targeting other epitopes in the V1V2 do-main, V2p and V2q, we tested MAbs CH58 (V2p), CH59 (V2p), and PG9 (V2q) against the same five viruses as were used in the
experiment whose results are shown inFig. 6. Increased
neutral-ization was seen with V2p MAbs, albeit only against SHIV
C.1157ipEL-p and HIV-1 92TH023 (Fig. 7); the two viruses are
known to be sensitive to CH58 and CH59 (7). In contrast, PG9
showed potent neutralizing activity against all viruses after 24 h of virus-MAb incubation, and the neutralization of YU2, SHIV C.1157ipEL-p, and CM244.OR was significantly enhanced with the longer incubation compared to the neutralization after the
standard 1 h (Fig. 7).
Taken together, these data demonstrate that, unlike the epitopes targeted by the highly potent MAbs, such as NIH45-46, neutralizing V2 and V3 epitopes on the Env of many HIV-1 iso-lates are not readily recognized by Abs at any one time but become accessible gradually over a period of time. Thus, a prolonged vi-rus-MAb incubation time is needed to detect neutralization by MAbs against these epitopes. This study also shows that different factors contribute to the occlusion of V2 and V3 epitopes from Ab recognition. Of the five parameters examined here, three influ-ence V3 epitopes, but only the time of MAb-virus preincubation modulates V2 epitopes.
DISCUSSION
The modest level of protection reported in the RV144 clinical trial has invigorated the HIV-1 research field and brought renewed attention to the V1V2 and V3 regions of HIV-1 Env gp120. How-ever, it remains unknown whether and how Abs target these loops to reduce infection by HIV. Although many MAbs against V1V2 and V3 cross-react with monomeric Env of HIV-1 isolates from different subtypes, they generally display weak neutralization
against tier 2 and tier 3 viruses (13,17,45,46), indicating poor
accessibility of the V1V2 and V3 epitopes on the Env spikes. The present study evaluates whether the same mechanisms underlie V2i and V3 epitope occlusion from and exposure to MAbs that target these epitopes. Interestingly, CD4-induced Env conforma-tional changes, which affect V3, had no effect on V2i epitopes. This finding may be informed by the V2i position revealed in the
recently published trimeric HIV-1 Env structures (14,15,47,48).
The V1V2 domain is localized at the apex of the trimer above and
distant from the CD4-binding site (14, 49, 50), and the V2i
epitopes in particular encompass the disordered, surface-exposed
region of V2 that contains the␣47-binding motif (6,12). The V3
loop, on the other hand, is beneath V1V2, with its epitope-bearing crown pointing toward the inner trimer axis, away from the trimer
surface (15), albeit this is only a single structure of an envelope
from a single strain, constrained by crystal packing and bound Ab. V1V2 deletion and CD4 treatment augment V3 epitope exposure,
FIG 5Effect of pH and temperature on virus neutralization by V2i and V3 MAbs. (A) To measure the effect of pH on virus infectivity, BaL.ec1 pseudovirus was incubated for 1 h in medium adjusted to pH 4.2, 5.6, 6.6, or 7.4 prior to the addition of TZM.bl target cells. Virus infectivity was measured 48 h later based on-galactosidase activity. (B) To compare BaL.ec1 neutralization by V2i MAb 2158 and control MAb 1418 at pH 6.6 versus pH 7.4, virus-MAb mixtures were incubated at the designated pH for 1 h or 5 h before adding the target cells. Means and standard errors from representative experiments (each in duplicate) are shown. (C) To evaluate the effect of higher temperature on virus neutralization, 5-fold serial dilutions of V2i and V3 MAbs were incubated with JRFL and BaL.ec1 at 37°C versus 38.5°C for 1 h before the addition of TZM-bl target cells. The irrelevant control MAb 1418 was used as a negative control. The cells were then cultured at 37°C for 48 h and infection measured based on-galactosidase activity. Means and standard errors of two experiments are shown.
Modulation of V2 versus V3 Epitopes
on November 7, 2019 by guest
http://jvi.asm.org/
[image:7.585.41.538.70.342.2]thereby promoting virus capture and neutralization by V3 MAbs
(22,51,52). Indeed, CD4 binding induces the crown of V3 to flip
up or protrude from the surface of the Env trimer, as seen in the
monomeric gp120 core (53), but is not likely to modify the
acces-sibility of V2i epitopes as seen in this study.
We also investigated the contribution of N-glycans in masking the V2i versus V3 epitopes. N-glycans play an important role in viral escape by shielding the key vulnerable epitopes on the Env
spike (23), including V3 epitopes. For example, the removal of
N-glycans at positions 187 and 197 in the V2 loop and at position 301 on the V3 base improved the antigenicity and immunogenic-ity of V3, CD4bs, and membrane proximal external region
(MPER) epitopes (19,54–56). Enriching the N-glycans on HIV
Env with the high-mannose type by growing viruses in
kifu-nensine or in a cell line lackingN-acetylglucosamine transferase I
also increased virus sensitivity to V3 MAbs (24,25). Conversely,
for the relatively sensitive tier 1 virus BaL, kifunensine reduced neutralization by V3 MAb 447, suggesting the potential role of glycans in governing V3 exposure on the more open Env trimers of the tier 1 viruses. In contrast, kifunensine treatment had no positive or negative effects on neutralization with the seven V2i MAbs tested, indicating that the N-glycan composition regulating V3 epitopes is not a factor underpinning poor Ab recognition of V2i epitopes. All N-glycans observed in the crystal structures of
scaffolded V1V2 point away from the V2i (6,16,57). Hence,
al-though N-glycans influence the Ab recognition of V2i epitopes
(13), they function mainly to induce and maintain proper folding
of the V1V2 domain. N-glycans are not part of the V2i epitope, in contrast to the V2q epitopes recognized by PG9 and PG16, and do not participate in shielding V2i epitopes from neutralizing Abs.
Finally, we evaluated the time-dependent dynamics of the V2i and V3 epitopes to determine whether these epitopes continu-ously sample different conformations, only rarely adopting those recognizable by the Abs at any one time. If this idea is correct, the epitope occupancy by the Abs would increase over time, leading to greater virus binding and neutralization with prolonged virus-Ab incubation time. The inability to resolve the V2 loop structure where the major element of V2i epitopes is located points to the highly flexible and dynamic nature of this region on the HIV Env trimer. In the case of V3 epitopes, multiple conformations have been observed in the crystal structures of V3 peptides bound by
each of the various V3 MAbs studied (58,59), which yet differ
from those seen on monomeric gp120 and trimeric gp140 pro-teins. The data from the present study agree with this idea, since virus neutralization by most V3 and V2i MAbs increased
signifi-cantly withⱖ18 h of virus-MAb preincubation. A previous study
by Binley et al. (60) also showed that prolonging the incubation
time from 1 h to 18 h improved virus neutralization by MAbs against V3, MPER, and the CD4-binding site. Here, we report for the first time time-dependent neutralization by MAbs targeting different epitopes in the V1V2 domain, including V2i, V2p, and V2q. Notably, the enhanced neutralization attained following the prolonged incubation time with these V2 MAbs was observed against specific virus isolates. For example, V2p MAbs which are not broadly reactive were effective only against tier 1 viruses SHIV C.1157ipEL-p and HIV-1 92TH023, while V2i MAbs were more cross-reactive and displayed potent neutralization against tier 1 viruses BaL, SHIV C.1157ipEL-p, and 92TH023 and tier 2 viruses JRFL and REJO, but only two V2i MAbs neutralized YU2 (tier 2) and none neutralized CM244.OR (tier 2). The specificity of virus
FIG 6Time-dependent increase of neutralization against different viruses by V2i and V3 MAbs. Virus neutralization was measured after REJO (A), YU2 (B), SHIV C.1157ipEL-p (C), HIV-1 92TH023 (D), and CM244.OR (E) vi-ruses were incubated with serially diluted MAbs for 1 h versus 24 h at 37°C prior to the addition of TZM-bl target cells. Means and standard errors from two to three experiments are shown. Statistical analyses were done on neutral-ization data reachingⱖ50%. *,P⬍0.001 for REJO, YU2, and SHIV andP⬍
0.05 for CM244.OR and HIV-1 92TH023.
on November 7, 2019 by guest
http://jvi.asm.org/
[image:8.585.39.278.64.602.2]neutralization detected under the prolonged virus-MAb incuba-tion was also evident from the lack of virus neutralizaincuba-tion by V2i
MAbs that do not recognize the virus envelope (17; also data not
shown) and by the irrelevant control MAb 1418.
Among the different parameters evaluated in this study, only virus-MAb incubation time was found to influence the neutraliz-ing activities of V2i MAbs. The neutralization levels achieved by the different V2i MAbs following 24 h of incubation correlated with the MAb binding affinities, while the neutralization levels with the standard 1-h incubation did not. In contrast, increasing the temperature during virus-MAb incubation from 37°C to 38.5°C had no effect on the neutralizing activity of V2i or V3 MAbs. A previous study has shown, however, that higher temper-atures increase the fluidity of membranes and facilitate HIV-1 entry via the fusion pores, thereby promoting HIV-1 infectivity
(61). We also studied the effect of lower pHs on virus infectivity
and neutralization by V2i MAbs. Virus infectivity decreased to
⬍10% at pH 4.2 and 5.6 but remained unchanged at pH 6.6
com-pared to the infectivity at pH 7.4. In agreement with these data,
past studies demonstrate that the acidic pH of⬍5.0 in the vaginal
mucosa is detrimental to the virus but that semen neutralizes the
environment, promoting virus transmission (62–64). At pH 6.6
and pH 7.4, the levels of virus neutralization by V2i MAb 2158 were comparable whether the virus was incubated with the MAb for 1 h or 5 h. Hence, unlike incubation time, pH and temperature in the limited ranges tested have minimal effects on virus neutral-ization by V2i MAbs.
The virus neutralization observed after 18 h or 24 h of incuba-tion cannot be explained simply by lower virus input due to virus decay over time. With 1 h of incubation, the V2i MAb did not mediate any neutralization even when smaller amounts of input virus were used. Indeed, no neutralization was observed over the entire range of virus dilutions examined. On the other hand, with a 24-h incubation, the neutralization levels correlated inversely with the virus input. The longer time needed for V2i MAbs to neutralize HIV-1 suggests their relatively low occupancy rate, or on rate, for binding to the Env spike. This is supported by our data showing that the levels of V2i MAb binding to Env on the cell surface after the initial 30 min were low relative to those seen with potently neutralizing MAbs, such as NIH45-46. However, V2i MAb binding continued to increase over time, and as more epitopes are occupied, the virus neutralization threshold is reached after 18 h. Based on Haim’s model of virus- and
Ab-specific factors determining HIV-1 neutralization (65), the V2i
MAbs fall into the Ab class with a high perturbation factor, which requires high degrees of conformational changes in the Env or epitope region for Ab recognition. This is consistent with the
flex-ible nature of the V2i region (14–16,49) and with the idea that the
V2i conformation recognized by the Ab is displayed only for a brief period of time and by a fraction of the Env. Altogether, these data support the notion that V2i exposure is not influenced by N-glycans or CD4 binding; rather, the V2i epitopes situated on the surface-accessible region of the trimeric Env spike are subjected to conformational masking. Similar to V2i, the structural dynamics
FIG 7Virus neutralization by V2p and V2q MAbs after prolonged virus-MAb incubation. Virus neutralization by V2p (CH58 and CH59) and V2q (PG9) MAbs was measured after 1 h versus 24 h of virus-MAb preincubation. REJO (A), YU2 (B), SHIV C.1157ipEL-p (C), 92TH023 (D), and CM244.OR (E) were incubated with MAbs for the designated period of time at 37°C prior to the addition of TZM-bl target cells. Means and standard errors from two to three experiments are shown. Statistical analyses were done as described in the legend toFig. 6. *,P⬍0.001 for REJO, YU2, SHIV (exceptP⬍0.05 for PG9), HIV-1 92TH023, and CM244.OR.
Modulation of V2 versus V3 Epitopes
on November 7, 2019 by guest
http://jvi.asm.org/
[image:9.585.83.507.68.356.2]of V2p and V2q epitopes also influence the neutralization activity of MAbs targeting these epitopes, and the effects vary depending on the virus strains.
The significance of neutralization mediated by V2i Abs follow-ing an 18-h or longer incubation with the virus is yet unknown.
Moreover, relatively high concentrations (20 to 100g/ml) are
needed to achieveⱖ50% neutralization. However, high levels of
Abs to the variable loops are attainable in chronically
HIV-in-fected patients (66), although it remains unknown whether these
levels can be reached by immunization, particularly locally at the site of virus entry. Perhaps a cocktail of MAbs or polyclonal Abs consisting of V2i Abs and/or other specificities may reach suffi-ciently high concentrations to mediate neutralization and act in a
synergistic or additive manner (67–69). Since virus neutralization
after prolonged incubation with V2i MAbs is inversely correlated with the amount of virus input, these Abs are likely to have an impact on the low-dose virus transmission that occurs via sexual transmission and not the high-dose infection among intravenous drug users. Indeed, HIV-1-specific antibodies with modest virus-neutralizing activity have shown protection against SHIV
trans-mission in low-dose challenge studies (70). The half-life of plasma
virions was reported to be between 6 h and 1.2 days (71,72). With
18 h required for V2i Abs to exert significant binding and neutral-ization, these Abs could reduce the virus infectivity and the size of
the infectious inoculumin vivo. Nonetheless, virus entry to target
cells is much faster than 18 h, as the entire sequence of CD4 bind-ing, chemokine receptor engagement, and virus fusion occurs in
⬃60 min (73). Cell-to-cell transmission of HIV can be completed
even more rapidly, within 30 min or less, in the tight cell-cell
junction microenvironment of the virological synapse (74–76).
Whether V2i Abs requiring the prolonged virus-Ab interaction would be able to exert any inhibitory effects against virus acquisi-tionin vivorequires further investigation.
The steps in the virus life cycle that can be affected by V2i Abs also need to be better understood. In this study, the TZM.bl assay was used to measure virus neutralization. The target cells used in
this standard assay express CD4 and CCR5 but not␣47, which
has been implicated in facilitating virus attachment to
gut-muco-sal CD4 target cells, in part by interacting with V2 (77). If V2i Abs
are present and interact with transmitted virus at the mucosal port of virus entry, they may reduce infection of CD4 T cells by
block-ing virus attachment via␣47. Nonetheless, a more recent study
demonstrates that␣47 binding may not be a general property of
the Envs of most HIV-1 variants (78).
Notably, this study provides novel evidence that neutralization of relatively resistant HIV-1 isolates, such as JRFL, REJO, and YU2, may be mediated by V3 and V2i MAbs, which previously were deemed to be nonneutralizing against these and other tier 2
viruses (17). Considering that induction of V3 and V2i Abs by
immunization is achievable with vaccine immunogens already tested in human clinical trials and that reduced risk of infection
is correlated with antibodies of these specificities (2–4,79), the
data presented here help us better understand the potential mechanisms by which V3 and V2i Abs may decrease HIV-1 acquisition.
ACKNOWLEDGMENTS
This study was supported in part by NIH grants R01 AI102740 (C.E.H.) and HIVRAD P01 AI100151 (S.Z.-P.) and by research funds from the
Henry M. Jackson Foundation and the Department of Veterans Affairs, Veterans Health Administration, Office of Research and Development.
We thank Constance Williams and Vincenza Itri for providing MAbs, Florian Klein for pMX-gp160⌬c-IRES-GFP plasmids, Victoria Polonis, Sebastian Mark Molnar, and Morgan Watts for the BaL.ec1 pseudovirus, Sodsai Tovanabutra for the CM244.OR plasmid, Michael Tuen for gen-eral laboratory support, and Susanne Tranguch for reviewing the manu-script.
REFERENCES
1.Rerks-Ngarm S, Pitisuttithum P, Nitayaphan S, Kaewkungwal J, Chiu J, Paris R, Premsri N, Namwat C, de Souza M, Adams E, Benenson M, Gurunathan S, Tartaglia J, McNeil JG, Francis DP, Stablein D, Birx DL, Chunsuttiwat S, Khamboonruang C, Thongcharoen P, Robb ML, Mi-chael NL, Kunasol P, Kim JH. 2009. Vaccination with ALVAC and AIDSVAX to prevent HIV-1 infection in Thailand. N. Engl. J. Med.361: 2209 –2220.http://dx.doi.org/10.1056/NEJMoa0908492.
2.Haynes BF, Gilbert PB, McElrath MJ, Zolla-Pazner S, Tomaras GD, Alam SM, Evans DT, Montefiori DC, Karnasuta C, Sutthent R, Liao HX, DeVico AL, Lewis GK, Williams C, Pinter A, Fong Y, Janes H, DeCamp A, Huang Y, Rao M, Billings E, Karasavvas N, Robb ML, Ngauy V, de Souza MS, Paris R, Ferrari G, Bailer RT, Soderberg KA, Andrews C, Berman PW, Frahm N, De Rosa SC, Alpert MD, Yates NL, Shen X, Koup RA, Pitisuttithum P, Kaewkungwal J, Nitayaphan S, Rerks-Ngarm S, Michael NL, Kim JH.2012. Immune-correlates analysis of an HIV-1 vaccine efficacy trial. N. Engl. J. Med.366:1275–1286.http: //dx.doi.org/10.1056/NEJMoa1113425.
3.Zolla-Pazner S, Decamp A, Gilbert PB, Williams C, Yates NL, Williams WT, Howington R, Fong Y, Morris DE, Soderberg KA, Irene C, Reich-man C, Pinter A, Parks R, Pitisuttithum P, Kaewkungwal J, Rerks-Ngarm S, Nitayaphan S, Andrews C, O’Connell RJ, Yang ZY, Nabel GJ, Kim JH, Michael NL, Montefiori DC, Liao HX, Haynes BF, Tomaras GD.2014. Vaccine-induced IgG antibodies to V1V2 regions of multiple HIV-1 subtypes correlate with decreased risk of HIV-1 infection. PLoS One9:e87572.http://dx.doi.org/10.1371/journal.pone.0087572. 4.Zolla-Pazner S, deCamp AC, Cardozo T, Karasavvas N, Gottardo R,
Williams C, Morris DE, Tomaras G, Rao M, Billings E, Berman P, Shen X, Andrews C, O’Connell RJ, Ngauy V, Nitayaphan S, de Souza M, Korber B, Koup R, Bailer RT, Mascola JR, Pinter A, Montefiori D, Haynes BF, Robb ML, Rerks-Ngarm S, Michael NL, Gilbert PB, Kim JH.2013. Analysis of V2 antibody responses induced in vaccinees in the ALVAC/AIDSVAX HIV-1 vaccine efficacy trial. PLoS One8:e53629.http: //dx.doi.org/10.1371/journal.pone.0053629.
5.Rolland M, Edlefsen PT, Larsen BB, Tovanabutra S, Sanders-Buell E, Hertz T, deCamp AC, Carrico C, Menis S, Magaret CA, Ahmed H, Juraska M, Chen L, Konopa P, Nariya S, Stoddard JN, Wong K, Zhao H, Deng W, Maust BS, Bose M, Howell S, Bates A, Lazzaro M, O’Sullivan A, Lei E, Bradfield A, Ibitamuno G, Assawadarachai V, O’Connell RJ, deSouza MS, Nitayaphan S, Rerks-Ngarm S, Robb ML, McLellan JS, Georgiev I, Kwong PD, Carlson JM, Michael NL, Schief WR, Gilbert PB, Mullins JI, Kim JH.2012. Increased HIV-1 vaccine efficacy against viruses with genetic signatures in Env V2. Nature490:417– 420.http://dx.doi.org/10.1038/nature11519.
6.Spurrier B, Sampson J, Gorny MK, Zolla-Pazner S, Kong XP. 2014. Functional implications of the binding mode of a human conformation-dependent V2 monoclonal antibody against HIV. J. Virol.88:4100 – 4112.
http://dx.doi.org/10.1128/JVI.03153-13.
7.Liao HX, Bonsignori M, Alam SM, McLellan JS, Tomaras GD, Moody MA, Kozink DM, Hwang KK, Chen X, Tsao CY, Liu P, Lu X, Parks RJ, Montefiori DC, Ferrari G, Pollara J, Rao M, Peachman KK, Santra S, Letvin NL, Karasavvas N, Yang ZY, Dai K, Pancera M, Gorman J, Wiehe K, Nicely NI, Rerks-Ngarm S, Nitayaphan S, Kaewkungwal J, Pitisuttithum P, Tartaglia J, Sinangil F, Kim JH, Michael NL, Kepler TB, Kwong PD, Mascola JR, Nabel GJ, Pinter A, Zolla-Pazner S, Haynes BF.2013. Vaccine induction of antibodies against a structurally heteroge-neous site of immune pressure within HIV-1 envelope protein variable regions 1 and 2. Immunity 38:176 –186. http://dx.doi.org/10.1016/j .immuni.2012.11.011.
8.Bonsignori M, Pollara J, Moody MA, Alpert MD, Chen X, Hwang KK, Gilbert PB, Huang Y, Gurley TC, Kozink DM, Marshall DJ, Whitesides JF, Tsao CY, Kaewkungwal J, Nitayaphan S, Pitisuttithum P, Rerks-Ngarm S, Kim JH, Michael NL, Tomaras GD, Montefiori DC, Lewis
on November 7, 2019 by guest
http://jvi.asm.org/
GK, DeVico A, Evans DT, Ferrari G, Liao HX, Haynes BF. 2012. Antibody-dependent cellular cytotoxicity-mediating antibodies from an HIV-1 vaccine efficacy trial target multiple epitopes and preferentially use the VH1 gene family. J. Virol.86:11521–11532.http://dx.doi.org/10.1128 /JVI.01023-12.
9.Walker LM, Phogat SK, Chan-Hui PY, Wagner D, Phung P, Goss JL, Wrin T, Simek MD, Fling S, Mitcham JL, Lehrman JK, Priddy FH, Olsen OA, Frey SM, Hammond PW, Kaminsky S, Zamb T, Moyle M, Koff WC, Poignard P, Burton DR.2009. Broad and potent neutralizing antibodies from an African donor reveal a new HIV-1 vaccine target. Sci-ence326:285–289.http://dx.doi.org/10.1126/science.1178746.
10. Bonsignori M, Hwang KK, Chen X, Tsao CY, Morris L, Gray E, Marshall DJ, Crump JA, Kapiga SH, Sam NE, Sinangil F, Pancera M, Yongping Y, Zhang B, Zhu J, Kwong PD, O’Dell S, Mascola JR, Wu L, Nabel GJ, Phogat S, Seaman MS, Whitesides JF, Moody MA, Kelsoe G, Yang X, Sodroski J, Shaw GM, Montefiori DC, Kepler TB, Tomaras GD, Alam SM, Liao HX, Haynes BF.2011. Analysis of a clonal lineage of HIV-1 envelope V2/V3 conformational epitope-specific broadly neutral-izing antibodies and their inferred unmutated common ancestors. J. Virol. 85:9998 –10009.http://dx.doi.org/10.1128/JVI.05045-11.
11. Moore PL, Gray ES, Sheward D, Madiga M, Ranchobe N, Lai Z, Honnen WJ, Nonyane M, Tumba N, Hermanus T, Sibeko S, Mlisana K, Abdool Karim SS, Williamson C, Pinter A, Morris L.2011. Potent and broad neutralization of HIV-1 subtype C by plasma antibodies targeting a quaternary epitope including residues in the V2 loop. J. Virol.85:3128 – 3141.http://dx.doi.org/10.1128/JVI.02658-10.
12. Mayr LM, Cohen S, Spurrier B, Kong XP, Zolla-Pazner S.2013. Epitope mapping of conformational V2-specific anti-HIV human monoclonal an-tibodies reveals an immunodominant site in V2. PLoS One8:e70859.http: //dx.doi.org/10.1371/journal.pone.0070859.
13. Gorny MK, Moore JP, Conley AJ, Karwowska S, Sodroski J, Williams C, Burda S, Boots LJ, Zolla-Pazner S.1994. Human anti-V2 monoclonal antibody that neutralizes primary but not laboratory isolates of human immunodeficiency virus type 1. J. Virol.68:8312– 8320.
14. Lyumkis D, Julien JP, de Val N, Cupo A, Potter CS, Klasse PJ, Burton DR, Sanders RW, Moore JP, Carragher B, Wilson IA, Ward AB.2013. Cryo-EM structure of a fully glycosylated soluble cleaved HIV-1 envelope trimer. Science342:1484 –1490.http://dx.doi.org/10 .1126/science.1245627.
15. Julien JP, Cupo A, Sok D, Stanfield RL, Lyumkis D, Deller MC, Klasse PJ, Burton DR, Sanders RW, Moore JP, Ward AB, Wilson IA.2013. Crystal structure of a soluble cleaved HIV-1 envelope trimer. Science342: 1477–1483.http://dx.doi.org/10.1126/science.1245625.
16. McLellan JS, Pancera M, Carrico C, Gorman J, Julien JP, Khayat R, Louder R, Pejchal R, Sastry M, Dai K, O’Dell S, Patel N, Shahzad-ul-Hussan S, Yang Y, Zhang B, Zhou T, Zhu J, Boyington JC, Chuang GY, Diwanji D, Georgiev I, Kwon YD, Lee D, Louder MK, Moquin S, Schmidt SD, Yang ZY, Bonsignori M, Crump JA, Kapiga SH, Sam NE, Haynes BF, Burton DR, Koff WC, Walker LM, Phogat S, Wyatt R, Orwenyo J, Wang LX, Arthos J, Bewley CA, Mascola JR, Nabel GJ, Schief WR, Ward AB, Wilson IA, Kwong PD.2011. Structure of HIV-1 gp120 V1/V2 domain with broadly neutralizing antibody PG9. Nature 480:336 –343.http://dx.doi.org/10.1038/nature10696.
17. Gorny MK, Pan R, Williams C, Wang XH, Volsky B, O’Neal T, Spurrier B, Sampson JM, Li L, Seaman MS, Kong XP, Zolla-Pazner S.2012. Functional and immunochemical cross-reactivity of V2-specific mono-clonal antibodies from HIV-1-infected individuals. Virology427:198 – 207.http://dx.doi.org/10.1016/j.virol.2012.02.003.
18. Krachmarov CP, Honnen WJ, Kayman SC, Gorny MK, Zolla-Pazner S, Pinter A.2006. Factors determining the breadth and potency of neutral-ization by V3-specific human monoclonal antibodies derived from sub-jects infected with clade A or clade B strains of human immunodeficiency virus type 1. J. Virol.80:7127–7135.http://dx.doi.org/10.1128/JVI.02619 -05.
19. Li Y, Cleveland B, Klots I, Travis B, Richardson BA, Anderson D, Montefiori D, Polacino P, Hu SL.2008. Removal of a single N-linked glycan in human immunodeficiency virus type 1 gp120 results in an en-hanced ability to induce neutralizing antibody responses. J. Virol.82:638 – 651.http://dx.doi.org/10.1128/JVI.01691-07.
20. Guttman M, Kahn M, Garcia NK, Hu SL, Lee KK. 2012. Solution structure, conformational dynamics, and CD4-induced activation in full-length, glycosylated, monomeric HIV gp120. J. Virol.86:8750 – 8764.http: //dx.doi.org/10.1128/JVI.07224-11.
21. Lusso P, Earl PL, Sironi F, Santoro F, Ripamonti C, Scarlatti G, Longhi R, Berger EA, Burastero SE.2005. Cryptic nature of a conserved, CD4-inducible V3 loop neutralization epitope in the native envelope glycopro-tein oligomer of CCR5-restricted, but not CXCR4-using, primary human immunodeficiency virus type 1 strains. J. Virol.79:6957– 6968.http://dx .doi.org/10.1128/JVI.79.11.6957-6968.2005.
22. Mbah HA, Burda S, Gorny MK, Williams C, Revesz K, Zolla-Pazner S, Nyambi PN.2001. Effect of soluble CD4 on exposure of epitopes on primary, intact, native human immunodeficiency virus type 1 virions of different genetic clades. J. Virol.75:7785–7788.http://dx.doi.org/10.1128 /JVI.75.16.7785-7788.2001.
23. McCaffrey RA, Saunders C, Hensel M, Stamatatos L.2004. N-linked glycosylation of the V3 loop and the immunologically silent face of gp120 protects human immunodeficiency virus type 1 SF162 from neutraliza-tion by anti-gp120 and anti-gp41 antibodies. J. Virol.78:3279 –3295.http: //dx.doi.org/10.1128/JVI.78.7.3279-3295.2004.
24. Kumar R, Tuen M, Liu J, Nadas A, Pan R, Kong X, Hioe CE.2013. Elicitation of broadly reactive antibodies against glycan-modulated neu-tralizing V3 epitopes of HIV-1 by immune complex vaccines. Vaccine 31:5413–5421.http://dx.doi.org/10.1016/j.vaccine.2013.09.010. 25. Binley JM, Ban YE, Crooks ET, Eggink D, Osawa K, Schief WR, Sanders
RW.2010. Role of complex carbohydrates in human immunodeficiency virus type 1 infection and resistance to antibody neutralization. J. Virol. 84:5637–5655.http://dx.doi.org/10.1128/JVI.00105-10.
26. Platt EJ, Wehrly K, Kuhmann SE, Chesebro B, Kabat D.1998. Effects of CCR5 and CD4 cell surface concentrations on infections by macrophag-etropic isolates of human immunodeficiency virus type 1. J. Virol.72: 2855–2864.
27. Klein F, Gaebler C, Mouquet H, Sather DN, Lehmann C, Scheid JF, Kraft Z, Liu Y, Pietzsch J, Hurley A, Poignard P, Feizi T, Morris L, Walker BD, Fatkenheuer G, Seaman MS, Stamatatos L, Nussenzweig MC.2012. Broad neutralization by a combination of antibodies recogniz-ing the CD4 bindrecogniz-ing site and a new conformational epitope on the HIV-1 envelope protein. J. Exp. Med.209:1469 –1479.http://dx.doi.org/10.1084 /jem.20120423.
28. Pietzsch J, Scheid JF, Mouquet H, Klein F, Seaman MS, Jankovic M, Corti D, Lanzavecchia A, Nussenzweig MC.2010. Human anti-HIV-neutralizing antibodies frequently target a conserved epitope essential for viral fitness. J. Exp. Med.207:1995–2002.http://dx.doi.org/10.1084/jem .20101176.
29. Li M, Gao F, Mascola JR, Stamatatos L, Polonis VR, Koutsoukos M, Voss G, Goepfert P, Gilbert P, Greene KM, Bilska M, Kothe DL, Salazar-Gonzalez JF, Wei X, Decker JM, Hahn BH, Montefiori DC. 2005. Human immunodeficiency virus type 1 env clones from acute and early subtype B infections for standardized assessments of vaccine-elicited neutralizing antibodies. J. Virol. 79:10108 –10125.http://dx.doi.org/10 .1128/JVI.79.16.10108-10125.2005.
30. Nyambi PN, Mbah HA, Burda S, Williams C, Gorny MK, Nadas A, Zolla-Pazner S.2000. Conserved and exposed epitopes on intact, na-tive, primary human immunodeficiency virus type 1 virions of group M. J. Virol.74:7096 –7107.http://dx.doi.org/10.1128/JVI.74.15.7096 -7107.2000.
31. Pinter A, Honnen WJ, He Y, Gorny MK, Zolla-Pazner S, Kayman SC. 2004. The V1/V2 domain of gp120 is a global regulator of the sensitivity of primary human immunodeficiency virus type 1 isolates to neutralization by antibodies commonly induced upon infection. J. Virol.78:5205–5215.
http://dx.doi.org/10.1128/JVI.78.10.5205-5215.2004.
32. Gorny MK, Conley AJ, Karwowska S, Buchbinder A, Xu JY, Emini EA, Koenig S, Zolla-Pazner S.1992. Neutralization of diverse human immu-nodeficiency virus type 1 variants by an anti-V3 human monoclonal an-tibody. J. Virol.66:7538 –7542.
33. Gorny MK, Williams C, Volsky B, Revesz K, Wang XH, Burda S, Kimura T, Konings FA, Nadas A, Anyangwe CA, Nyambi P, Krachma-rov C, Pinter A, Zolla-Pazner S.2006. Cross-clade neutralizing activity of human anti-V3 monoclonal antibodies derived from the cells of individ-uals infected with non-B clades of human immunodeficiency virus type 1. J. Virol.80:6865– 6872.http://dx.doi.org/10.1128/JVI.02202-05. 34. Hioe CE, Wrin T, Seaman MS, Yu X, Wood B, Self S, Williams C,
Gorny MK, Zolla-Pazner S.2010. Anti-V3 monoclonal antibodies dis-play broad neutralizing activities against multiple HIV-1 subtypes. PLoS One5:e10254.http://dx.doi.org/10.1371/journal.pone.0010254. 35. Diskin R, Scheid JF, Marcovecchio PM, West AP, Jr, Klein F, Gao H,
Gnanapragasam PN, Abadir A, Seaman MS, Nussenzweig MC,
Bjork-Modulation of V2 versus V3 Epitopes
on November 7, 2019 by guest
http://jvi.asm.org/
man PJ.2011. Increasing the potency and breadth of an HIV antibody by using structure-based rational design. Science334:1289 –1293.http://dx .doi.org/10.1126/science.1213782.
36. Burton DR, Barbas CF, III, Persson MA, Koenig S, Chanock RM, Lerner RA.1991. A large array of human monoclonal antibodies to type 1 human immunodeficiency virus from combinatorial libraries of asymp-tomatic seropositive individuals. Proc. Natl. Acad. Sci. U. S. A.88:10134 – 10137.http://dx.doi.org/10.1073/pnas.88.22.10134.
37. Trkola A, Purtscher M, Muster T, Ballaun C, Buchacher A, Sullivan N, Srinivasan K, Sodroski J, Moore JP, Katinger H.1996. Human mono-clonal antibody 2G12 defines a distinctive neutralization epitope on the gp120 glycoprotein of human immunodeficiency virus type 1. J. Virol. 70:1100 –1108.
38. Montefiori DC.2005. Evaluating neutralizing antibodies against HIV, SIV, and SHIV in luciferase reporter gene assays. Curr. Protoc. Immu-nol. Chapter 12:Unit 12.11. http://dx.doi.org/10.1002/0471142735 .im1211s64.
39. Decker JM, Bibollet-Ruche F, Wei X, Wang S, Levy DN, Wang W, Delaporte E, Peeters M, Derdeyn CA, Allen S, Hunter E, Saag MS, Hoxie JA, Hahn BH, Kwong PD, Robinson JE, Shaw GM. 2005. Antigenic conservation and immunogenicity of the HIV coreceptor binding site. J. Exp. Med.201:1407–1419.http://dx.doi.org/10.1084 /jem.20042510.
40. Hoffman TL, LaBranche CC, Zhang W, Canziani G, Robinson J, Chai-ken I, Hoxie JA, Doms RW.1999. Stable exposure of the coreceptor-binding site in a CD4-independent HIV-1 envelope protein. Proc. Natl. Acad. Sci. U. S. A.96:6359 – 6364.http://dx.doi.org/10.1073/pnas.96.11 .6359.
41. Salzwedel K, Smith ED, Dey B, Berger EA. 2000. Sequential CD4-coreceptor interactions in human immunodeficiency virus type 1 Env function: soluble CD4 activates Env for coreceptor-dependent fusion and reveals blocking activities of antibodies against cryptic conserved epitopes on gp120. J. Virol.74:326 –333.http://dx.doi.org/10.1128/JVI.74.1.326 -333.2000.
42. Xiang SH, Doka N, Choudhary RK, Sodroski J, Robinson JE.2002. Characterization of CD4-induced epitopes on the HIV type 1 gp120 en-velope glycoprotein recognized by neutralizing human monoclonal anti-bodies. AIDS Res. Hum. Retroviruses18:1207–1217.http://dx.doi.org/10 .1089/08892220260387959.
43. Li Y, Rey-Cuille MA, Hu SL.2001. N-linked glycosylation in the V3 region of HIV type 1 surface antigen modulates coreceptor usage in viral infection. AIDS Res. Hum. Retroviruses17:1473–1479.http://dx.doi.org /10.1089/08892220152644179.
44. Scanlan CN, Ritchie GE, Baruah K, Crispin M, Harvey DJ, Singer BB, Lucka L, Wormald MR, Wentworth P, Jr, Zitzmann N, Rudd PM, Burton DR, Dwek RA.2007. Inhibition of mammalian glycan biosynthe-sis produces non-self antigens for a broadly neutralising, HIV-1 specific antibody. J. Mol. Biol.372:16 –22.http://dx.doi.org/10.1016/j.jmb.2007 .06.027.
45. Israel ZR, Gorny MK, Palmer C, McKeating JA, Zolla-Pazner S.1997. Prevalence of a V2 epitope in clade B primary isolates and its recognition by sera from HIV-1-infected individuals. AIDS11:128 –130.
46. Shotton C, Arnold C, Sattentau Q, Sodroski J, McKeating JA.1995. Identification and characterization of monoclonal antibodies specific for polymorphic antigenic determinants within the V2 region of the human immunodeficiency virus type 1 envelope glycoprotein. J. Virol.69:222– 230.
47. Mao Y, Wang L, Gu C, Herschhorn A, Desormeaux A, Finzi A, Xiang SH, Sodroski JG.2013. Molecular architecture of the uncleaved HIV-1 envelope glycoprotein trimer. Proc. Natl. Acad. Sci. U. S. A.110:12438 – 12443.http://dx.doi.org/10.1073/pnas.1307382110.
48. Bartesaghi A, Merk A, Borgnia MJ, Milne JL, Subramaniam S.2013. Prefusion structure of trimeric HIV-1 envelope glycoprotein determined by cryo-electron microscopy. Nat. Struct. Mol. Biol.20:1352–1357.http: //dx.doi.org/10.1038/nsmb.2711.
49. Mao Y, Wang L, Gu C, Herschhorn A, Xiang SH, Haim H, Yang X, Sodroski J.2012. Subunit organization of the membrane-bound HIV-1 envelope glycoprotein trimer. Nat. Struct. Mol. Biol.19:893– 899.http: //dx.doi.org/10.1038/nsmb.2351.
50. Khayat R, Lee JH, Julien JP, Cupo A, Klasse PJ, Sanders RW, Moore JP, Wilson IA, Ward AB.2013. Structural characterization of cleaved, solu-ble HIV-1 envelope glycoprotein trimers. J. Virol.87:9865–9872.http://dx .doi.org/10.1128/JVI.01222-13.
51. Wyatt R, Moore J, Accola M, Desjardin E, Robinson J, Sodroski J.1995. Involvement of the V1/V2 variable loop structure in the exposure of hu-man immunodeficiency virus type 1 gp120 epitopes induced by receptor binding. J. Virol.69:5723–5733.
52. Gzyl J, Bolesta E, Wierzbicki A, Kmieciak D, Naito T, Honda M, Komuro K, Kaneko Y, Kozbor D.2004. Effect of partial and complete variable loop deletions of the human immunodeficiency virus type 1 en-velope glycoprotein on the breadth of gp160-specific immune responses. Virology318:493–506.http://dx.doi.org/10.1016/j.virol.2003.10.009. 53. Huang CC, Tang M, Zhang MY, Majeed S, Montabana E, Stanfield RL,
Dimitrov DS, Korber B, Sodroski J, Wilson IA, Wyatt R, Kwong PD. 2005. Structure of a V3-containing HIV-1 gp120 core. Science310:1025– 1028.http://dx.doi.org/10.1126/science.1118398.
54. Pantophlet R, Wilson IA, Burton DR.2004. Improved design of an antigen with enhanced specificity for the broadly HIV-neutralizing anti-body b12. Protein Eng. Des. Sel.17:749 –758.http://dx.doi.org/10.1093 /protein/gzh085.
55. Ma BJ, Alam SM, Go EP, Lu X, Desaire H, Tomaras GD, Bowman C, Sutherland LL, Scearce RM, Santra S, Letvin NL, Kepler TB, Liao HX, Haynes BF.2011. Envelope deglycosylation enhances antigenicity of HIV-1 gp41 epitopes for both broad neutralizing antibodies and their unmutated ancestor antibodies. PLoS Pathog.7:e1002200.http://dx.doi .org/10.1371/journal.ppat.1002200.
56. Kolchinsky P, Kiprilov E, Bartley P, Rubinstein R, Sodroski J.2001. Loss of a single N-linked glycan allows CD4-independent human immu-nodeficiency virus type 1 infection by altering the position of the gp120 V1/V2 variable loops. J. Virol.75:3435–3443.http://dx.doi.org/10.1128 /JVI.75.7.3435-3443.2001.
57. Pancera M, Shahzad-Ul-Hussan S, Doria-Rose NA, McLellan JS, Bailer RT, Dai K, Loesgen S, Louder MK, Staupe RP, Yang Y, Zhang B, Parks R, Eudailey J, Lloyd KE, Blinn J, Alam SM, Haynes BF, Amin MN, Wang LX, Burton DR, Koff WC, Nabel GJ, Mascola JR, Bewley CA, Kwong PD.2013. Structural basis for diverse N-glycan recognition by HIV-1-neutralizing V1-V2-directed antibody PG16. Nat. Struct. Mol. Biol.20:804 – 813.http://dx.doi.org/10.1038/nsmb.2600.
58. Stanfield R, Cabezas E, Satterthwait A, Stura E, Profy A, Wilson I.1999. Dual conformations for the HIV-1 gp120 V3 loop in complexes with dif-ferent neutralizing fabs. Structure7:131–142.http://dx.doi.org/10.1016 /S0969-2126(99)80020-3.
59. Jiang X, Burke V, Totrov M, Williams C, Cardozo T, Gorny MK, Zolla-Pazner S, Kong XP.2010. Conserved structural elements in the V3 crown of HIV-1 gp120. Nat. Struct. Mol. Biol.17:955–961.http://dx.doi .org/10.1038/nsmb.1861.
60. Binley JM, Wrin T, Korber B, Zwick MB, Wang M, Chappey C, Stiegler G, Kunert R, Zolla-Pazner S, Katinger H, Petropoulos CJ, Burton DR. 2004. Comprehensive cross-clade neutralization analysis of a panel of an-ti-human immunodeficiency virus type 1 monoclonal antibodies. J. Virol. 78:13232–13252.http://dx.doi.org/10.1128/JVI.78.23.13232-13252.2004. 61. Harada S, Yusa K, Monde K, Akaike T, Maeda Y.2005. Influence of membrane fluidity on human immunodeficiency virus type 1 entry. Biochem. Biophys. Res. Commun. 329:480 – 486. http://dx.doi.org/10 .1016/j.bbrc.2005.02.007.
62. Aldunate M, Tyssen D, Johnson A, Zakir T, Sonza S, Moench T, Cone R, Tachedjian G.2013. Vaginal concentrations of lactic acid potently inactivate HIV. J. Antimicrob. Chemother.68:2015–2025.http://dx.doi .org/10.1093/jac/dkt156.
63. Klebanoff SJ, Coombs RW. 1991. Viricidal effect of Lactobacillus acidophilus on human immunodeficiency virus type 1: possible role in heterosexual transmission. J. Exp. Med.174:289 –292.http://dx.doi.org /10.1084/jem.174.1.289.
64. Klebanoff SJ, Kazazi F.1995. Inactivation of human immunodeficiency virus type 1 by the amine oxidase-peroxidase system. J. Clin. Microbiol. 33:2054 –2057.
65. Haim H, Salas I, McGee K, Eichelberger N, Winter E, Pacheco B, Sodroski J.2013. Modeling virus- and antibody-specific factors to predict human immunodeficiency virus neutralization efficiency. Cell Host Mi-crobe14:547–558.http://dx.doi.org/10.1016/j.chom.2013.10.006. 66. Zolla-Pazner S.2005. Improving on nature: focusing the immune
re-sponse on the V3 loop. Hum. Antibodies14:69 –72.
67. Barouch DH, Whitney JB, Moldt B, Klein F, Oliveira TY, Liu J, Stephenson KE, Chang HW, Shekhar K, Gupta S, Nkolola JP, Seaman MS, Smith KM, Borducchi EN, Cabral C, Smith JY, Blackmore S, Sanisetty S, Perry JR, Beck M, Lewis MG, Rinaldi W, Chakraborty AK,