JOURNAL OFVIROLOGY,
0022-538X/01/$04.00⫹0 DOI: 10.1128/JVI.75.21.10505–10510.2001Nov. 2001, p. 10505–10510 Vol. 75, No. 21 Copyright © 2001, American Society for Microbiology. All Rights Reserved.
Isomerization of a Uniquely Designed Amplicon during Herpes
Simplex Virus-Mediated Replication
HUA WANG,1XINPING FU,1ANDXIAOLIU ZHANG1,2*
Center for Cell and Gene Therapy1and Departments of Pediatrics and Molecular Virology & Microbiology,2
Baylor College of Medicine, Houston, Texas 77030 Received 18 April 2001/Accepted 22 July 2001
Herpes simplex virus (HSV) type 1 DNA isomerization was studied using a uniquely designed amplicon that mimics the viral genomic structure. The results revealed that amplicon concatemers frequently contain adjacent amplicon units with their segments in opposed orientations. These unusual concatemers were generated through homologous recombination, which does not require HSV DNA as the source of homology.
The herpes simplex virus type 1 (HSV-1) genome is a linear, double-stranded DNA molecule of 152 kb. It consists of two covalently linked segments designated long (L) and short (S). Each segment contains largely unique sequences (ULand US)
which are bracketed by inverted repeats. Previous studies have shown that HSV replicative intermediates are high-molecular-weight molecules in which the genomic termini are fused to-gether in a head-to-tail arrangement (2, 13, 14). These findings have led to a model in which the linear viral genome circular-izes immediately after infection and replicates unidirectionally by a rolling-circle mechanism. This mode of replication gener-ates a head-to-tail concatemer that is cleaved into unit-length genomes during packaging (10, 11, 19, 26). Consistent with this model, defective HSV-1 genomes are encapsidated as head-to-tail repeats (4, 28), structures that are compatible with a rolling-circle mechanism of DNA replication (21, 22).
A prominent feature of HSV DNA replication is the free inversion of the L and S segments relative to each other, generating four isomeric forms that occur naturally in equimo-lar proportions (5, 11). The molecuequimo-lar mechanism of segment inversion is poorly understood. The use of alternative cleavage sites during maturation and packaging of concatemeric inter-mediates can account for the generation of only two isomeric forms from a single monomeric template (25). The remaining isomers are thought to have been generated by homologous recombination, and the repeatedasequences appear to play an important role in this recombination-mediated segment inver-sion (3, 6, 7, 16–18, 24). However, recent studies have provided a different insight into the mechanism of HSV DNA isomer-ization. Analysis of replicative intermediates digested with re-striction enzymes that cleave once in the unique region of the virus genome has revealed that the concatemers very fre-quently contain adjacent genomic units with L segments in different orientations (called concatemers with internal seg-ment isomerization [concatemer-ISI]), from which all four pos-sible HSV isomers can be generated in an equal molar ratio through random cleavage and packaging (1, 15, 20, 27). This finding has suggested that HSV genome isomerization is
inti-mately linked to DNA replication, rather than to a late event associated with cleavage and packaging.
The mechanism for generating concatemer-ISI is not fully understood. Since conventional rolling-circle replication is un-able to generate such concatemers, they must arise by other, yet-unidentified mechanisms. Further elucidation of the mech-anism(s) requires a more detailed analysis of the viral replica-tive intermediates. However, the large size and complexity of the HSV genome make it time-consuming and technically dif-ficult to modify the viral genome in order to facilitate experi-mental designs. For the same reasons, it is also difficult to select unique restriction enzymes for Southern blot analysis. The HSV amplicon, which has a much smaller genome and contains only theciselements required for virus-mediated rep-lication and cleavage-packaging (i.e., the reprep-lication origin and the packaging signal) (8, 9), may be an alternative and simpli-fied tool for this purpose. Upon introduction into cells together with a helper virus, the HSV amplicon can be amplified, pre-sumably by a rolling-circle mechanism using the essential pro-teins provided by the helper virus. The replicated amplicon DNA can be subsequently packaged into viral particles, which are also provided by the helper virus, in a multiple-copy con-catemeric form.
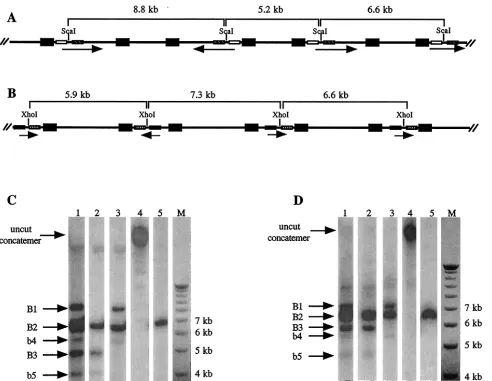
Earlier studies by us and others showed that neither isomer-ization nor concatemer-ISI could be detected on HSV ampli-cons harboring only a single set of viralciselements (18, 28). We therefore constructed a new version of the HSV amplicon that mimics the viral genomic structure; this amplicon has two sets of HSV repeat sequences, each composed of a replication origin (oriS) and ana sequence in different locations and in opposite orientations. This amplicon, called pSZ-dPac-EGFP, consequently has its own L and S segments bracketed by the HSV repeats (Fig. 1A), much like the native HSV genome. Purified pSZ-dPac-EGFP DNA was transfected into baby hamster kidney (BHK) cells, which were subsequently infected with a helper HSV (wild-type HSV strain 17). The amplicon was harvested and passaged twice before concatemers were extracted from packaged viral particles. DNA was digested with either ScaI, which has a single recognition site located close to one end of the L segment, orXhoI, which has a single recognition site located near one end of the S segment (Fig. 1A). If there is no internal segment isomerization and only head-to-tail concatemers are formed, then digestion with ei-* Corresponding author. Mailing address: Department of Pediatrics
and Center for Cell and Gene Therapy, Baylor College of Medicine, One Baylor Plaza, Houston, TX 77030. Phone: (713) 798-1256. Fax: (713) 798-1230. E-mail: [email protected].
10505
on November 9, 2019 by guest
http://jvi.asm.org/
ther of these enzymes will generate only the unit-length 6.6-kb fragment. Alternatively, if the orientation of the L segment is frequently opposed,ScaI digestion will generate two additional DNA fragments with sizes of 8.8 and 5.2 kb (Fig. 2A). Accord-ingly, if the orientation of the S segment is also frequently opposed, XhoI digestion will produce two additional DNA fragments with sizes of 7.3 and 5.9 kb (Fig. 2B).
Following restriction enzyme digestion, DNA fragments were separated by agarose gel electrophoresis, transferred to a nylon membrane, and hybridized with a probe (pC) which was made from the entire pSZ-dPac-EGFP sequence but lacked the oriS and a sequences (to limit cross-hybridization with DNA fragments from the helper virus genome).ScaI digestion of the packaged amplicon concatemer generated two strong bands of approximately 8.8 and 5.2 kb in size (designated B1 and B3, respectively) in addition to the 6.6-kb unit-length frag-ment (B2) (Fig. 2C, lane 1). A pair of additional bands 5.7 and 4.2 kb in size (designated b4 and b5, respectively) were also weakly visible; these bands represent two of the terminal frag-ments of the packaged amplicon concatemers.
To add further support to the segment isomerization pred-ication in Fig. 2A, we employed two additional probes, pS1 and pS2, that were made from different regions of the amplicon genome. Fitting the predicted isomerization pattern in Fig. 2A, the results in Fig. 2C (lanes 2 and 3) showed that pS1 hybrid-izes to the unit-length 6.6-kb fragment as well as the smaller 5.2-kb fragment, whereas pS2 hybridizes to the unit-length fragment and the larger 8.8-kb fragment. Digestion with the XhoI enzyme, which cuts within the S segment, also resulted in the predicted pattern (shown in Fig. 2B).XhoI digestion gen-erated two fragments (B1 and B3) in addition to the unit-length fragment (B2) when the digested concatemeric DNA was hybridized with probe pC (Fig. 2D, lane 1). Two weaker terminal fragments (b4 and b5, with sizes of 5.4 and 4.7 kb, respectively) were also visible. The disappearance of band B1
from the hybridization with the subgenomic probe pX1 (Fig. 2D, lane 2) and of band B3 from the hybridization with sub-genomic probe pX2 (Fig. 2D, lane 3) confirmed the specificity of each of the bands.
To estimate the percentage of concatemers containing L and S segments in opposing orientations, the intensity of each in-dividual band was quantified by phosphorimager analysis (Im-ageQuant). The results showed that the sum of the intensities of B1 and B3 was only slightly less than the intensity of B2 in bothScaI- andXhoI-digested DNA samples (data not shown, but see lanes 1 of Fig. 2C and D). Collectively, these results demonstrate that during HSV-mediated pSZ-dPac-EGFP rep-lication, a high percentage (30 to 50%) of concatemers contain L and S segments in opposite orientations in neighboring am-plicon genomes.
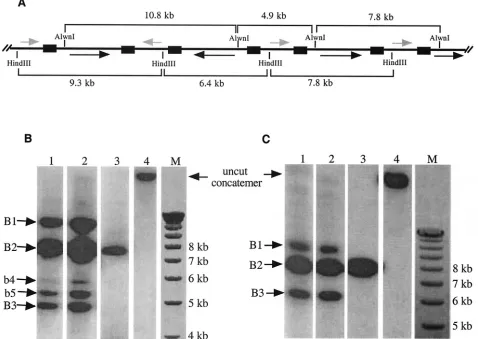
Next, we conducted experiments to determine if the gener-ation of concatemer-ISI is through homologous recombina-tion, as confirmation of this aspect would further support the hypothesis that there is a direct link between the production of concatemer-ISI and the generation of equimolar HSV iso-forms. To conduct this experiment, we constructed another amplicon plasmid, pW4.2 (Fig. 1B). In addition to a single copy of theoriSand theasequence of HSV-1, this amplicon con-tains two copies of the ampicillin gene (ampR) arranged in different loci of the plasmid and in opposite orientations. This arrangement creates the L and S segments of this plasmid, which are bracketed by the ampicillin genes rather than the repeated sequences of HSV. The same Southern blotting strat-egy described above was used to detect the generation of concatemer-ISI, with the exception that the amplicon concate-mers were digested with eitherAlwnI, which cuts the plasmid once at one end of the L segment, orHindIII, which cuts the plasmid once at one end of the S segment. The predicted isomerization patterns and the DNA fragments and their sizes followingAlwnI orHindIII digestion are shown in Fig. 3A. The FIG. 1. Structures of pSZ-dPac-EGFP and pW4.2. Short and long segments of the plasmids are labeled S and L, respectively. The orientations of the repeated regions of pSZ-dPac-EGFP includingoriSand theasequence (a-seq) are shown by arrowheads. The names and locations of the four subgenomic DNA fragments used for making probes are indicated. The orientations of the two ampicillin genes (AmpR) in pW4.2 are also labeled. The locations of the unique restriction enzymes used to determine the segment orientation in the concatemers are indicated by individual enzyme names.
on November 9, 2019 by guest
amplicon concatemers were extracted from stocks which had been passaged either once (Fig. 3B, lanes 1 and 4) or twice (lane 2).AlwnI digestion of the concatemeric DNA generated three major DNA bands (B1 to B3) of approximately 10.8, 7.8, and 4.9 kb, respectively (Fig. 3B, lanes 1 and 2). The weakly hybridizing terminal fragments (b4 and b5, of 5.3 and 5.5 kb, respectively) were also visible. HindIII digestion also gener-ated three strong bands with sizes of approximately 9.3, 7.8, and 6.4 kb (3C, lanes 1 and 2). Quantification of the bands representing concatemer-ISI (i.e., bands B1 and B3) by phos-phorimager analysis showed that the concatemer-ISI from pW4.2 occurred at a frequency similar to that from pSZ-dPac-EGFP (data not shown). The ratio remained almost un-changed during amplicon passages (compare lanes 1 and 2 in Fig. 3B and C). These results indicate that the ampicillin gene
sequence can fully replace the HSV repeated sequences to achieve high-frequency homologous recombination during HSV-1-mediated DNA replication, leading to the frequent generation of concatemer-ISI. These results therefore confirm that concatemer-ISI generation occurs through homologous recombination and that it can take place between non-HSV DNA elements.
One possible mechanism to generate concatemer-ISI during virus infection is through homologous recombination between two different isoforms of the viral genome. This would require the preexistence of at least two different isoforms in the input virus. Alternatively, if a single isoform of HSV was used, mul-tiple rounds of viral replication would be required to generate more than one HSV isoform. However, it is not technically possible to perform an experiment with a strict single-round FIG. 2. Southern blot analysis of pSZ-dPac-EGFP concatemers. (A and B) Schematic representations of pSZ-dPac-EGFP concatemers and possible orientations of the L (A) and S (B) segments; the orientation of each segment is indicated by an arrow. The repeated regions (oriSand theasequence) of each amplicon are represented by filled boxes. TheScaI andXhoI recognition sites are marked along the concatemers, and the sizes of the restriction fragments are noted. The locations of the subgenomic DNA fragments used for making probes are represented by open bars (pS1), hatched bars (pS2), crosshatched bars (pX1), and shaded bars (pX2). (C and D) Southern blot hybridization followingScaI (C) orXhoI (D) digestion. Lanes 1 to 3, digested concatemeric pSZ-dPac-EGFP DNA; lanes 4, undigested DNA; lanes 5, digested purified pSZ-dPac-EGFP plasmid DNA. The marker (lanes M) is the 1-kb ladder (Gibco-BRL). Each lane of the blot was hybridized as follows: in panels C and D, lanes 1, 4, and 5 were hybridized with probe pC; in panel C, lanes 2 and 3 were hybridized with pS1 and pS2, respectively; and in panel D, lanes 2 and 3 were hybridized with pX1 and pX2, respectively.
VOL. 75, 2001 NOTES 10507
on November 9, 2019 by guest
http://jvi.asm.org/
infection-replication because each plaque-purified virus would undergo many rounds of replication before a sufficiently large stock could be made for experimental characterization. In ad-dition, it is not possible to generate a virus stock containing only a single HSV isoform, since each new virus stock contains all four possible HSV isoforms in an equimolar ratio. Since the pSZ-dPac-EGFP amplicon generates a similar pattern of con-catemer-ISI, we queried whether a strict single-round DNA amplification of a single isoform could be performed with this amplicon. To test this, cells were transfected with pSZ-dPac-EGFP and were infected 16 h later with helper virus at 10 PFU per cell, a dose at which the majority of cells will be infected in the first round and will therefore prevent a subsequent second-round infection. We assumed that under these experimental conditions, the amplicon would undergo only a single round of amplification and packaging. Concatemeric DNA extracted from virions released from the cells was digested withScaI and hybridized with probe pC. In this single-step DNA replication setting, concatemer-ISI, which was represented by the appear-ance of B1 and B3 bands, was efficiently generated (Fig. 4, lane
1) and largely maintained during subsequent serial passages (Fig. 4, lanes 2, 3, and 4). Phosphorimager quantification showed that the ratio of the intensities of bands B1 and B3 to that of B2 in lane 1 of Fig. 4 was approximately 2:3 and that the ratio remained almost unchanged during serial passages of the stock (Fig. 4, lanes 2, 3, and 4). These results therefore suggest that generation of concatemer-ISI does not require the preex-istence of more than one isomer of pSZ-dPac-EGFP.
Our earlier studies showed that homologous recombination could occur between a wild-type HSV genome and a mutant HSV genome that has a single SpeI recognition site deleted (23). However, the relative amount of concatemers generated from recombination between mutant and wild-type HSV was far less than that of the concatemer-ISI generated from the wild-type virus genome alone. A possible explanation for this discrepancy could be the branched nature of HSV replicative intermediates and/or the relatively large sizes of the DNA molecules that were studied (180 to 260 kb), which may have limited efficient DNA transfer during Southern blotting pro-cedures. In order to measure the relative amounts of concate-FIG. 3. Generation of concatemer-ISI is through homologous recombination. (A) Schematic representation of a pW4.2 concatemer and possible orientations of the L (black arrows) and S segments (gray arrows) along the molecule. The repeated regions (ampicillin gene) of the construct are represented by filled boxes. TheAlwnI andHindIII sites are marked along the concatemer, and the sizes of the restriction fragments are noted. (B and C) Southern blot hybridization following digestion withAlwnI (B), which cuts once within the L segment of pW4.2, orHindIII (C), which cuts once within the S segment. Concatemeric DNA was extracted from amplicon stocks that had been passaged either once (lanes 1 and 4) or twice (lane 2). The DNA in lane 3 represents unit-length pW4.2 plasmid DNA. Hybridization was with a probe made from the entire pW4.2 genome but lacking theoriSandasequence. The marker (lanes M) is the 1-kb ladder.
on November 9, 2019 by guest
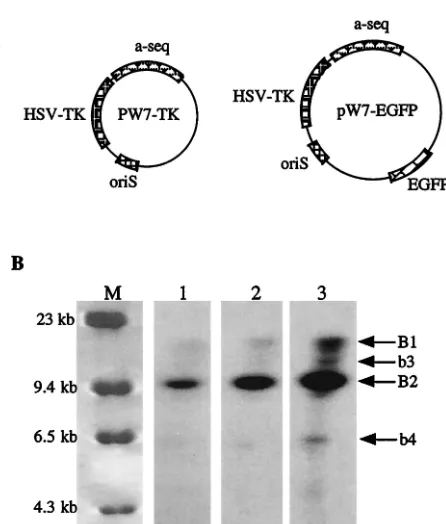
[image:4.587.54.532.74.413.2]mers more accurately, we carried out an experiment similar to the ones described above but employed two amplicons which had much smaller genomes than the virus. It has been reported that when two similar-sized plasmids carrying different marker genes were mixed at a 1:1 ratio for in vitro transfection, the majority of the transfected cells were found to express both marker genes (12). We therefore transfected BHK cells with a DNA mixture of two amplicons, pW7-TK and pW7-EGFP, at an equimolar ratio. Plasmid pW7-EGFP was constructed by inserting an enhanced green fluorescent protein (EGFP) gene cassette [containing the cytomegalovirus promoter and bovine growth hormone poly(A)] into the uniqueSapI site of pW7-TK, such that pW7-TK has 100% homology to pW7-EGFP outside the EGFP cassette (Fig. 5A). Sixteen hours after trans-fection with the DNA mixture, the cells were infected with the wild-type helper virus HSV strain 17. Virion DNA extracted from virus particles was digested withXhoI, which has a single recognition site located in the EGFP cassette region but does not cut within pW7-TK itself. The digested DNA was subjected to gel electrophoresis and Southern blot hybridization with a radioactive probe made from the EGFP gene (this probe will identify only pW7-EGFP but not pW7-TK unless it has recom-bined with pW7-EGFP). Frequent homologous recombination between these two amplicons during HSV-mediated DNA am-plification should generate a significant number of concatem-ers containing both amplicons intconcatem-erspconcatem-ersed along a single mol-ecule, which upon digestion withXhoI will produce a 19.5-kb DNA fragment in addition to the unit-length 11.7-kb pW7-EGFP fragment. Figure 5B shows that with a normal exposure time (lanes 1 and 2), only a single band (B2) representing the unit-length pW7-EGFP could be detected. However, after an extremely long exposure (lane 3), a band of approximately 19 kb was also visible, although the intensity of this band was only a small proportion of that of B2. The other two weakly hybrid-izing bands (b3 and b4, of approximately 6 and 14 kb, respec-tively) represent the terminal fragments of concatemers that contain the EGFP gene. These results are in agreement with
our previous observations (23) and indicate that random ho-mologous recombination between two different genomes does occur but at a frequency too low to contribute significantly to the observed large amount of concatemer-ISI during HSV-mediated DNA replication.
Based on these observations, we conclude that the genera-tion of concatemer-ISI is a generalized phenomenon that can occur in the absence of a complete HSV genome. Further-more, concatemer-ISI is the direct result of homologous re-combination, which does not specifically require that the ho-mologous sequence be from HSV DNA. The demonstration of a similar pattern of ISI during amplicon replication also indi-cates that uniquely designed amplicons such as pSZ-dPac-EGFP may be useful as a simplified model for further inves-tigation into HSV replication mechanisms.
We thank Malcolm K. Brenner for continuous support and C. Ma for technical assistance.
REFERENCES
1.Bataille, D., and A. Epstein.1994. Herpes simplex virus replicative concate-mers contain L components in inverted orientation. Virology203:384–388. 2.Ben Porat, T., A. S. Kaplan, B. Stehn, and A. S. Rubensten.1977.
Replica-tion of herpes DNA. II. SedimentaReplica-tion characteristics of newly synthesized DNA. Virology79:292–301.
3.Bruckner, R. C., R. E. Dutch, B. V. Zemelman, E. S. Mocarski, and I. R. Lehman.1992. Recombination in vitro between herpes simplex virus type 1 a sequences. Proc. Natl. Acad. Sci. USA89:10950–10954.
[image:5.587.312.535.64.326.2]4.Deiss, L. P., and N. Frenkel.1986. Herpes simplex virus amplicon: cleavage of concatemeric DNA is linked to packaging and involves amplification of FIG. 4. Generation of concatemer-ISI during a single-step
ampli-con replication. The ampliampli-con was hybridized with probe pC. Lane 1, transfection-infection; lane 2, passage 1; lane 3, passage 2; lane 4, passage 3; lane 5, unit-length pSZ-dPac-EGFP plasmid DNA; lane 6, undigested amplicon concatemer (from the passage 1 sample). The marker (lane M) is the 1-kb ladder.
FIG. 5. Homologous recombination between different amplicons is a rare event. (A) Schematic representations of TK and pW7-EGFP. (B) Southern blotting results. Following a mixed transfection of pW7-TK and pW7-EGFP and subsequent helper HSV infection, con-catemeric DNA was extracted, digested withXhoI, and hybridized with a probe made from the gene encoding EGFP. The DNA marker (lane M) is-HindIII (Gibco-BRL). Lanes 1 to 3 show different exposures of the same Southern blot: lane 1, short exposure; lane 2, medium expo-sure; lane 3, long exposure.
VOL. 75, 2001 NOTES 10509
on November 9, 2019 by guest
http://jvi.asm.org/
[image:5.587.45.284.75.235.2]restriction endonuclease cleavage sites. Proc. Natl. Acad. Sci. USA72:1768– 1772.
12.Horbinski, C., M. K. Stachowiak, D. Higgins, and S. G. Finnegan.2001. Polyethyleneimine-mediated transfection of cultured postmitotic neurons from rat sympathetic ganglia and adult human retina. BMC Neurosci.2:2. 13.Jacob, R. J., L. S. Morse, and B. Roizman.1979. Anatomy of herpes simplex
virus DNA. XIII. Accumulation of head to tail concatemers in nuclei of infected cells and their role in the generation of the four isomeric arrange-ments of viral DNA. J. Virol.29:448–457.
14.Jacob, R. J., and B. Roizman.1977. Anatomy of herpes simplex virus DNA. VIII. Properties of the replicating DNA. J. Virol.23:394–411.
15.McVoy, M. A., and S. P. Adler.1994. Human cytomegalovirus DNA repli-cates after early circularization by concatemer formation, and inversion occurs within the concatemer. J. Virol.68:1040–1051.
16.Mocarski, E. S., L. E. Post, and B. Roizman.1980. Molecular engineering of the herpes simplex virus genome: insertion of a second L-S junction into the genome causes additional genome inversions. Cell22:243–255.
17.Mocarski, E. S., and B. Roizman.1981. Site-specific inversion sequence of
viral DNA. J. Virol.73:810–813.
24.Smiley, J. R., B. S. Fong, and W. C. Leung.1981. Construction of a double-jointed herpes simplex viral DNA molecule: inverted repeats are required for segment inversion, and direct repeats promote deletions. Virology113:
345–362.
25.Varmuza, S. L., and J. R. Smiley.1985. Signals for site-specific cleavage of HSV DNA: maturation involves two separate cleavage events at sites distal to the recognition sequences. Cell41:793–802.
26.Vlazny, D., A. Kwong, and N. Frenkel.1982. Site-specific cleavage/packaging of herpes simplex virus DNA and the selective maturation of nucleocapsids containing full length viral DNA. Proc. Natl. Acad. Sci. USA79:1423–1427. 27.Zhang, X., S. Efstathiou, and A. Simmons.1994. Identification of novel herpes simplex virus replicative intermediates by field inversion gel electro-phoresis: implications for viral DNA amplification strategies. Virology202:
530–539.
28.Zhang, X., H. O’Shea, C. Entwisle, M. Boursnell, S. Efstathiou, and S. Inglis.1998. An efficient selection system for packaging herpes simplex virus amplicons. J. Gen Virol.79:125–131.