0022-538X/11/$12.00 doi:10.1128/JVI.01707-10
Copyright © 2011, American Society for Microbiology. All Rights Reserved.
Epitope Mapping of HIV-Specific CD8
⫹
T Cell Responses by Multiple
Immunological Readouts Reveals Distinct Specificities Defined by Function
䌤
Meika Richmond,
1Lyle R. McKinnon,
2,3Sandra A. Koesters Kiazyk,
4Charles Wachihi,
2Makobu Kimani,
2Joshua Kimani,
1,2Francis A. Plummer,
1,2,4and T. Blake Ball
1,2,4,5,6*
Department of Medical Microbiology, University of Manitoba, Winnipeg, Canada1; Department of Medical Microbiology,University of Nairobi, Nairobi, Kenya2; Department of Medicine, University of Toronto, Toronto, Canada3;
National Microbiology Laboratory, Public Heath Agency of Canada, Winnipeg, Canada4; Department of
Immunology, University of Manitoba, Winnipeg, Canada5; and National HIV and Retrovirology Laboratories,
Public Health Agency of Canada, Winnipeg, Canada6
Received 12 August 2010/Accepted 9 November 2010
The limited success of HIV vaccine candidates to date highlights our need to better characterize protective cell-mediated immunity (CMI). While HIV-specific CD8ⴙ T cell responses have been defined largely by measuring gamma interferon (IFN-␥), these responses are not always protective, and it is unclear whether the same epitopes would predominate if other functional parameters were examined. Here, we assessed the epitope specificity of HIV-specific CD8ⴙT cell responses by multiparametric flow cytometry, measuring five CD8ⴙT cell functions (IFN-␥, macrophage inflammatory protein 1[MIP-1], tumor necrosis factor alpha [TNF-␣], interleukin-2 [IL-2], and proliferative capacity) in 24 chronically HIV-infected individuals. Sixty-nine epitope-specific responses to 50 epitopes within p24 were measured. Surprisingly, most epitope-epitope-specific responses were IFN-␥negative (50/69 responses). Many responses had polyfunctional (33%) and proliferative (19%) compo-nents. An inverse association between IL-2 and proliferation responses was also observed, contrary to what was described previously. We confirm that long-term nonprogressors (LTNP) have more polyfunctional responses and also have higher-magnitude and broader p24-specific proliferation and higher levels of IL-2 and TNF-␣ production than do progressing controls. Together, these data suggest that the specificity of CD8ⴙ T cell responses differs depending on the immunological readout, with a 3.5-fold increase in breadth detected by including multiple parameters. Furthermore, the identification of epitopes that elicit polyfunctional responses reinforces the need for the comprehensive evaluation of HIV vaccine candidates, and these epitopes may represent novel targets for CMI-based vaccines.
As the human immunodeficiency virus (HIV)/AIDS epi-demic continues to grow, there is a desperate need for an effective vaccine. Most current HIV vaccine candidates aim to induce HIV-specific CD8⫹ T cell responses capable of con-taining viral replication and slowing disease progression. This vaccine concept is based on several lines of evidence suggesting that CD8⫹T cell responses can control viral replication (7, 25, 27, 40). However, HIV-specific CD8 responses are detected in nearly all HIV-positive (HIV⫹) subjects regardless of disease progression (8, 20, 21, 38). Several large studies have found no correlation between HIV-specific CD8⫹T cell gamma inter-feron (IFN-␥) secretion and viral load, and high-avidity re-sponses to autologous virus can be measured in subjects who are progressing to AIDS (16, 26). HIV-specific CD8⫹T cells are often exhausted or functionally inferior in chronic, pro-gressive HIV-1 infection, in some cases lacking perforin ex-pression, cytokine secretion, and proliferative capacity (30, 33). These data suggest that not all CD8⫹ T cell responses are effective, and responses that better correlate with protection need to be identified.
A subgroup of HIV-infected subjects, termed long-term
nonprogressors (LTNP), experience slower progression to AIDS and provide a valuable model for the study of cell-mediated immunity (CMI) responses that may be capable of controlling HIV. Previous work has demonstrated that these individuals maintain stronger HIV-specific CD8⫹T cell pro-liferation than do progressing controls (11, 30, 33). LTNP were also found to have more polyfunctional HIV-specific CD8⫹T cells, as defined by the concurrent expression of the cytokines IFN-␥, interleukin-2 (IL-2), and tumor necrosis factor alpha (TNF-␣); the chemokine macrophage inflammatory protein 1 (MIP-1); and the degranulation marker CD107a (1, 5). Poly-functional CD8⫹T cell responses have been measured in humans vaccinated with the highly efficacious smallpox virus vaccine (37), while polyfunctional CD4⫹T cells have been found to be protec-tive in settings where immunity is primarily cell mediated, for example, following tuberculosis vaccination and in murine models of Leishmania major (6, 10). The detection of polyfunctional CD8⫹T cells in HIV-1-exposed but -uninfected subjects poten-tially demonstrates that these polyfunctional responses may play a role in protection against HIV infection (17).
Most studies describing the epitope specificity of CD8⫹T cell responses in HIV infection have relied extensively on IFN-␥ enzyme-linked immunospot (ELISPOT) assays (44), which allow a rapid definition of positive responses. Recent studies have begun to call into question the reliability of ELISPOT assays for the assessment effective immune re-sponses (4, 45, 47). The use of a single readout may miss many * Corresponding author. Mailing address: Department of Medical
Microbiology, University of Manitoba, H3440-1015 Arlington St., Win-nipeg, MB R3E 3R2 Canada. Phone: (204) 784-8074. Fax: (204) 789-2018. E-mail: [email protected].
䌤Published ahead of print on 17 November 2010.
1275
on November 7, 2019 by guest
http://jvi.asm.org/
effective responses, particularly a readout that may not mea-sure responses capable of controlling HIV infection.
For example, little is known regarding the specificity of pro-liferative responses despite evidence that they are associated with the control of HIV infection (11, 24, 33). Data from our laboratory examining CD8⫹T cell responses to HIV Env using IFN-␥ELISPOT assays and 6-day carboxyfluorescein diacetate succinimidyl ester (CFSE) proliferation assays revealed sub-stantial differences in the epitopes recognized between assays (31). Here we extend these observations using an unbiased epitope-mapping approach to determine the specificity of poly-functional CD8⫹T cell responses. Although proliferation and polyfunctional responses have previously been associated with LTNP, those studies utilized peptide pools or predefined epitopes to measure responses. By epitope mapping of HIV-specific CD8⫹T cells with multiple parameters, we observed many readout-specific responses, including a response breadth of over 3.5 times that if IFN-␥was used alone.
MATERIALS AND METHODS
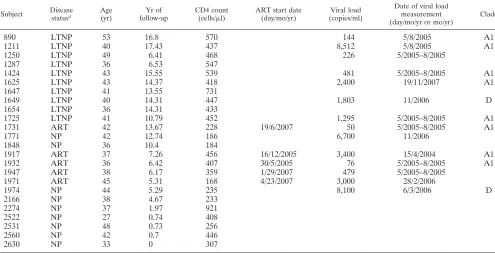
Subjects.Study participants (n⫽24) were all HIV infected and enrolled in a well-described longitudinal female sex worker cohort based in Nairobi, Kenya (18) (Table 1). Written informed consent was obtained from all study partici-pants, and ethics review boards from the University of Manitoba and the Ken-yatta National Hospital approved the study. Clinical and demographic data, including CD4 T cell counts, were collected from subjects biannually.
Antiret-roviral therapy (ART)-naïve subjects with CD4⫹counts above 400 cells/l for
over 6 years were classified as long-term nonprogressors (LTNP). LTNP were monitored for mean and median times of 11.98 and 14.31 years, respectively
(range, 6.41 to 17.43 years;n⫽10). Participants who did not meet the criteria for
LTNP were considered to be normal progressors (NP) (n⫽9); the time to
progression to AIDS has been demonstrated to be 3.5 years for this cohort (2). Additionally, five subjects on ART were included in the study.
Flow cytometry panels.HIV-specific responses were assessed by the simulta-neous measurement of four immunological parameters in a panel that included
CD3-AmCyan, CD8-allophycocyanin cynine 7 (APCCy7), IFN-␥–fluorescein
iso-thiocyanate (FITC), MIP-1–phycoerythrin (PE), IL-2–allophycocyanin (APC)
(BD Biosciences), and TNF-␣–Pacific Blue (eBiosciences). Proliferation was
assessed by using a flow cytometry panel that included carboxyfluorescein diac-etate succinimidyl ester (CFSE; Molecular Probes, Invitrogen), CD3-AmCyan, and CD8-APCCy7 (BD Biosciences).
Peptides.Peptides (9-mer peptides overlapping by 8 amino acids [aa]) derived from the HIV-1 p24 clade A1 ancestral sequence (Sigma-Genosys) were pooled in a matrix format using Deconvolute This!, version 1.0 (39) (courtesy of Mario Roederer, Vaccine Research Center, NIAID, NIH). Clade A1 is the predomi-nant circulating clade in this study population (12, 35, 36). Each peptide was represented twice, resulting in 16 pools with approximately 30 peptides per pool.
Peptide pools were used at 2g/ml/peptide, and stimulations were accompanied
by 2 positive controls, cytomegalovirus (CMV), Epstein-Barr virus, and influenza
virus (CEF) peptides (32 peptides/pool, 2g/ml/peptide; AnaSpec) and
Staph-ylococcus aureusenterotoxin B (SEB) (0.1g/ml; Sigma-Aldrich), and duplicate negative controls consisting of medium alone.
Intracellular cytokine staining.Fresh peripheral blood mononuclear cells (PBMC) were stimulated with peptide pools, CEF, SEB, or medium overnight for 14 h. The stimulation time of 14 h was chosen to optimize IL-2 expression.
PBMC were incubated at 1⫻106cells/ml in complete R-10 medium (RPMI 1640
[HyClone, Thermo Scientific] supplemented with 10% heat-inactivated fetal bovine serum [FBS] [Gibco, Invitrogen] and 1% Antimycotic Penstrep [Gibco, Invitrogen]). After 1 h of stimulation at 37°C, pretitrated amounts of monensin
(Golgistop, 1l/ml; BD Biosciences) and brefeldin A (Golgiplug, 1l/ml; BD
Biosciences) were added, followed by a further 13 h of incubation. Following incubation the cells were washed and stained according to the manufacturer’s protocol, using a Cytofix/Cytoperm kit (BD PharMingen). Stained cells were analyzed immediately.
Proliferation assays.PBMC were resuspended in phosphate-buffered saline (PBS) and loaded with CFSE according to the manufacturer’s protocols (Mo-lecular Probes). Cells were than stimulated with p24 peptide pools and incubated for 6 days at 37°C in R-10 medium. The same sets of negative and positive controls as those used for the overnight assays were used. On day 6, the cells were washed, stained using surface antibodies, fixed with 1% paraformaldehyde, and acquired immediately.
[image:2.585.43.538.82.335.2]Data acquisition and analysis.Cells were analyzed on an LSRII flow cytom-eter (BD Biosciences). Between 30,000 and 100,000 events were collected within the lymphocyte gate per sample. Data analyses were performed by using FlowJo software, version 8.7.3 (TreeStar). Boolean gates were applied to the 4 overnight
TABLE 1. Demographic data for study subjects
Subject Disease
statusa
Age (yr)
Yr of follow-up
CD4 count
(cells/l)
ART start date (day/mo/yr)
Viral load (copies/ml)
Date of viral load measurement (day/mo/yr or mo/yr)
Clade
890 LTNP 53 16.8 570 144 5/8/2005 A1
1211 LTNP 40 17.43 437 8,512 5/8/2005 A1
1250 LTNP 49 6.41 468 226 5/2005–8/2005
1287 LTNP 36 6.53 547
1424 LTNP 43 15.55 539 481 5/2005–8/2005 A1
1625 LTNP 43 14.37 418 2,400 19/11/2007 A1
1647 LTNP 41 13.55 731
1649 LTNP 40 14.31 447 1,803 11/2006 D
1654 LTNP 36 14.31 433
1725 LTNP 41 10.79 452 1,295 5/2005–8/2005 A1
1731 ART 42 13.67 228 19/6/2007 50 5/2005–8/2005 A1
1771 NP 42 12.74 186 6,700 11/2006
1848 NP 36 10.4 184
1917 ART 37 7.26 456 16/12/2005 3,400 15/4/2004 A1
1932 ART 36 6.42 407 30/5/2005 76 5/2005–8/2005 A1
1947 ART 38 6.17 359 1/29/2007 479 5/2005–8/2005
1971 ART 45 5.31 168 4/23/2007 3,000 28/2/2006
1974 NP 44 5.29 235 8,100 6/3/2006 D
2166 NP 38 4.67 233
2274 NP 37 1.97 921
2522 NP 27 0.74 408
2531 NP 48 0.73 256
2560 NP 42 0.7 446
2630 NP 33 0 307
aLTNP, long-term nonprogressor; NP, normal progressor; ART, subjects on antiretroviral therapy.
on November 7, 2019 by guest
http://jvi.asm.org/
functions, and the polyfunctionality of each response was assessed by using SPICE 5.1 (courtesy of Mario Roederer, Vaccine Research Center, NIAID, NIH).
Statistical analysis.HIV-specific CD8⫹T cell responses were considered
positive for a given parameter if they wereⱖ2-fold higher than the mean of their
respective negative controls. Data reported are values after background subtrac-tion. Polyfunctional responses were also background subtracted, and a lower threshold corresponding to the 90th percentile of distribution of negative values was built for each cytokine pattern, and values below this threshold were set to 0. For some participants limited cell numbers prevented the confirmation of all putative epitopes; in these cases the cutoff for a positive pool response was raised to 3-fold over background, and therefore, only the strongest peptides were confirmed. Statistical analyses were performed by using Graph Pad Prism 5.0 and SPICE 5.1. All correlations were determined by using Spearman’s rank correla-tion. Breadth and magnitude comparisons between subject groups were deter-mined by using Mann-Whitney tests.
RESULTS
Epitope mapping of p24 using parallel measurements of four CD8ⴙ T cell functions. HIV-specific CD8⫹ T cell re-sponses to p24 epitopes were mapped by simultaneously mea-suring IFN-␥, MIP-1, IL-2, and TNF-␣production following overnight stimulations with 16 peptide pools representing HIV-1 clade A1 p24. Representative intracellular and prolif-eration responses are shown in Fig. 1a. For this particular participant, CD8⫹T cell responses to a single peptide were detectable for all four effector functions, while proliferation
was detected for another peptide. The full complexity of the short-term response to all peptide pools was then examined by using Boolean gating, yielding 16 unique response combina-tions for the four individual readouts measured (Fig. 1b).
LTNP maintain a higher degree of HIV-specific CD8ⴙT cell functionality and stronger proliferation, IL-2, and TNF-␣ re-sponses. Previous work suggested that LTNP maintain a higher degree of polyfunctional HIV-specific CD8⫹T cell re-sponses than do NP (5, 48). However, this was examined mostly using predefined epitopes or peptide pools and not a comprehensive epitope-screening approach. After Boolean gating and stratification by disease status and ART use, we confirmed that a higher degree of CD8⫹T cell polyfunction-ality in LTNP was observed for our study (Fig. 1c). HIV-specific CD8⫹T cell responses from LTNP and ART subjects (Fig. 1b, blue and red bars, respectively) displayed a higher functional profile than did those from NP (P⬍0.0001) (Fig. 1b, green bars, and c). While responses that included all four functions were nearly absent from NP, they were observed at a low frequency in LTNP and at a relatively high frequency in subjects on ART, presumably due to reconstituted immune responses in the latter.
[image:3.585.47.542.70.251.2]Although their CD8⫹T cell responses were more polyfunc-tional, LTNP did not have a higher breadth of response than
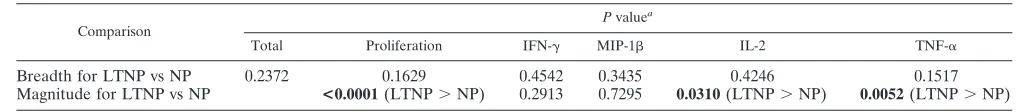
TABLE 2. Comparison of breadths and magnitudes of responses in LTNP and NP
Comparison
Pvaluea
Total Proliferation IFN-␥ MIP-1 IL-2 TNF-␣
Breadth for LTNP vs NP 0.2372 0.1629 0.4542 0.3435 0.4246 0.1517
Magnitude for LTNP vs NP <0.0001(LTNP⬎NP) 0.2913 0.7295 0.0310(LTNP⬎NP) 0.0052(LTNP⬎NP)
a
[image:3.585.35.543.660.716.2]Boldface type indicates a significant association.
FIG. 1. Representative polyfunctional intracellular cytokine staining and 6-day CFSE dilution. (A) Initial gating of forward-scatter area (FSC-A) versus forward-scatter height (FSC-H) used to eliminated doublets. The side-scatter area (SSC-A) versus the FSC-A was used to identify lymphocytes. After gating on CD3⫹CD8⫹cells, further gates were made for each of the respective functions (IFN-␥, MIP-1, TNF-␣, IL-2, and proliferation). Shown are representative data from subject ML2630, including overnight responses to the p24 peptide QGQMVHQSL (aa 7 to 14) and the day 6 response to the peptide SDIAGTTST (aa 102 to 110). (B) Boolean gates were applied to the 4 overnight functions. The 16 possible CD8⫹T cell response combinations are shown for LTNP, ART, and NP. IFN-␥, MIP-1, IL-2, and/or TNF-␣responses are shown as positive (⫹) and negative (⫺). (C) Comparison of the mean responses to p24 peptide pools between LTNP, ART, and NP. For simplicity, responses are grouped by number of functions, matched to the colored bars shown in B. LTNP and ART have a higher frequency of polyfunctional responses than do NP (P⬍0.001).
on November 7, 2019 by guest
http://jvi.asm.org/
did NP, regardless of the parameter taken into account (Table 2). Pool-specific responses in LTNP were typically of a higher magnitude than those observed for NP (proliferation, P ⬍ 0.0001; IL-2,P⫽0.0310; TNF-␣,P⫽0.0052 [by Mann-Whit-ney test]) (Table 2). The total breadth of responses, measured by the total number of responding pools regardless of param-eter or disease profile, was not associated with either viral load or CD4 count. However, CD4 counts were associated with the breadth of both pool-specific proliferation and TNF-␣ re-sponses (r⫽0.41 andP⫽0.0453, andr⫽0.41 andP⫽0.0461, respectively, by Spearman rank correlation) (Table 3). These associations suggest that polyfunctional p24 responses are an important correlate of LTNP in this cohort.
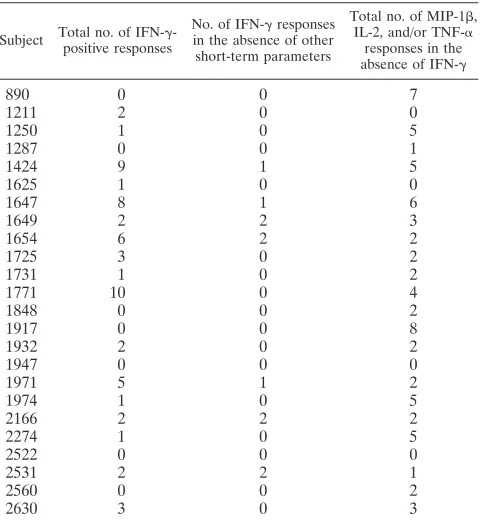
Different functional readouts define divergent specificities of CD8ⴙT cell responses.We hypothesized that regardless of the progression status, the specificity of responses to p24 pools would differ depending on the functional readout of the HIV-specific CD8⫹T cell response that was measured. In a propor-tion of participants (7/24 participants), pool-specific IFN-␥ re-sponses were observed in the absence of any other functional parameters; conversely, the majority of participants (20/24) displayed one or more of MIP-1, IL-2, and/or TNF-␣in the absence of IFN-␥(Table 4). These data suggest that many CD8
T cell responses are missed if IFN-␥was the only parameter measured.
For the majority of the subjects (67%; 16/24), at least one p24 pool was IFN-␥ positive (IFN-␥⫹) in the absence of a proliferative response. Similar data were obtained by examin-ing proliferation responses: 50% of subjects (12/24) had at least one proliferation-positive pool that was negative for IFN-␥. In total, the majority of IFN-␥responses did not have an accompanying proliferative response (91%; 54/59), and 25/30 (83%) of proliferation-positive pools lacked a corre-sponding IFN-␥response. Only 6% of responding pools were positive in both assays, all of which occurred in the LTNP, while 64% were IFN-␥⫹and proliferation negative, and 30% were IFN-␥negative (IFN-␥⫺) and proliferation positive (Fig. 2a). These data suggest that specific epitopes within these p24 pools are resulting in either an IFN-␥ or a proliferative re-sponse but rarely both in the same subject.
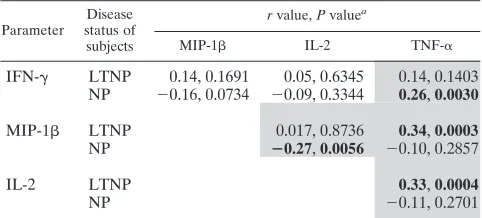
To better understand the relationship between specific ef-fectors and proliferative responses, we first determined the association between short-term pool-specific responses in LTNP and NP subjects. This approach allows the evaluation of both the concurrent expression of multiple cytokines, where one cell is responsible for the secretion of both effectors, and the coexpression of multiple cytokines, where the same sample is positive for two cytokines but is expressed by different cells. Interestingly, HIV-specific CD8⫹T cell responses from LTNP and NP differed in many associations, principally those involv-ing TNF-␣expression (Table 5). In general, LTNP exhibited more consistent associations between overnight parameters than did NP. This supports the observation that LTNP were more likely to have a concurrent expression of multiple param-eters and have a higher degree of polyfunctionality.
[image:4.585.43.543.81.136.2]We next examined the correlation between overnight and proliferation responses. Across the entire data set, the magni-tude of IFN-␥-positive responses correlated inversely with the magnitude of the proliferation response to the corresponding pool (r⫽ ⫺0.14 andP⫽0.0114 by Spearman’s rank correla-tion) (Fig. 2c). This is similar to what has previously been described, where substantial differences were observed in the epitope specificities between IFN-␥ELISPOT and 6-day pro-liferation assays (31). In the present study, propro-liferation re-sponses were inversely correlated with the magnitude of all overnight readouts, including MIP-1, IL-2, and TNF-␣ (all P⬍0.0001) (Fig. 2c and d). When stratified by disease profile, IFN-␥ and proliferation responses did not correlate in either LTNP or NP (Table 6). In addition, proliferation also correlated inversely with the all other effector functions in both groups (P⬍ 0.05). These data again demonstrate substantial differences in the antigen specificities of CD8⫹T cells measured by proliferative assays compared to any readout in overnight assays.
TABLE 3. Comparison of breadths of responses with CD4 counts and viral loads
Comparison
rvalue,Pvaluea
Total Proliferation IFN-␥ MIP-1 IL-2 TNF-␣
CD4 count 0.33, 0.1143 0.41,0.0453 ⫺0.12, 0.5781 ⫺0.39, 0.0596 ⫺0.002, 0.9934 0.41,0.0461 Viral load 0.23, 0.4354 0.43, 0.1439 0.15, 0.6329 0.24, 0.4332 0.39, 0.1836 ⫺0.18, 0.5577
a
Boldface type indicates a significant association.
TABLE 4. p24 pool responses showing disconnect between IFN-␥ and other short-term parameters
Subject Total no. of IFN-␥
-positive responses
No. of IFN-␥responses
in the absence of other short-term parameters
Total no. of MIP-1,
IL-2, and/or TNF-␣
responses in the
absence of IFN-␥
890 0 0 7
1211 2 0 0
1250 1 0 5
1287 0 0 1
1424 9 1 5
1625 1 0 0
1647 8 1 6
1649 2 2 3
1654 6 2 2
1725 3 0 2
1731 1 0 2
1771 10 0 4
1848 0 0 2
1917 0 0 8
1932 2 0 2
1947 0 0 0
1971 5 1 2
1974 1 0 5
2166 2 2 2
2274 1 0 5
2522 0 0 0
2531 2 2 1
2560 0 0 2
2630 3 0 3
on November 7, 2019 by guest
http://jvi.asm.org/
[image:4.585.44.284.462.724.2]FIG. 2. Disconnect between overnight and proliferation assays. (A) Percentage of CD8⫹T cells that were IFN-␥positive in the overnight assay and proliferation negative (Pro⫺) in the day 6 assay, those that were IFN-␥negative and proliferation positive, and those that were positive in both assays. (B) Percentage of CD8⫹that were IL-2 positive in the overnight assay and proliferation negative in the day 6 assay, those that were IL-2 negative proliferation positive, and those that were positive in both assays. (C) Inverse correlation between pool-specific proliferative responses and all four overnight readouts (Spearman’s rank correlation). Shown are percentages of day 6 and overnight responses for all subjects and all responses. (D) Background-adjusted CD8⫹T cell responses in subjects ML2531 (NP) (top) and ML1647 (LTNP) (bottom). In each of these instances, the only response to the respective pool was proliferation (pools 14 and 8, respectively).
on November 7, 2019 by guest
http://jvi.asm.org/
HIV-1-specific IL-2⫹ and proliferating CD8⫹ T cell re-sponses were previously observed to correlate strongly (48). Therefore, the inverse relationship between IL-2-secreting HIV-specific CD8⫹T cells and proliferation responses (r ⫽ ⫺0.49;P⬍0.0001) (Fig. 2c) observed in our study was unex-pected. In the present study, 70% (17/24) of subjects had at least one pool that was IL-2⫹but did not have an accompa-nying proliferative response. Similarly, 58% of subjects (14/24) had at least one proliferation-positive pool that was negative for IL-2. In total, only 5% (2/41) of IL-2 responses had an accompanying proliferative response, and only 7% (2/30) of the proliferation responses had a corresponding IL-2 re-sponse (Fig. 2b). While only 3% of pools were IL-2⫹ and proliferation positive, 56% were IL-2⫹ and proliferation negative, and 41% were IL-2⫺ and proliferation positive (Fig. 2b). The data suggest an even greater disparity be-tween proliferation and IL-2 than what was observed for IFN-␥and suggest that IL-2 might not always be a reliable predictor of proliferative capacity.
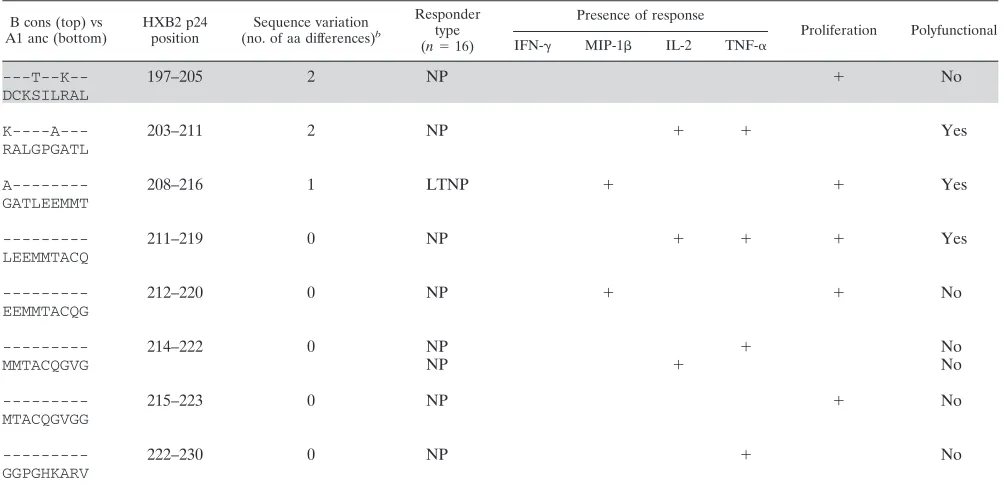
The epitope specificities of CD8ⴙ T cell responses differ based on functional readout. To further dissect differences between functional readouts at the epitope level, we confirmed the epitope specificities of pool responses. Putative responding peptides were identified by using the pooling matrix, and these were subsequently confirmed at later time points for 16/24 subjects (126 putative epitopes were tested with a range of 4 to 21 peptides tested/subject). Similar numbers of epitopes were tested between patient groups (data not shown). All 16 subjects responded to at least one of their putative epitopes, making a total of 69 responses to 50 epitopes (Table 7). There was no difference in the breadths of the response to the p24 epitopes between LTNP and NP (P⫽0.9423; mean numbers of peptides of 4.7 for LTNP and 4.5 for NP) (data not shown).
In agreement with the pool data, non-IFN-␥responses were observed for a high proportion (50/69; 73%) of peptide-specific responses. These data clearly show that a substantial portion of immune responses would remain undetected had we measured IFN-␥alone. Proliferation was a component of 13/69 (18%) responses, and only one response was proliferation positive and IFN-␥⫹. Consistent with pool-specific responses, the per-centages of epitope-specific IFN-␥-responding cells were in-versely correlated with the percentages of proliferating cells (r⫽ ⫺0.66; P⬍0.0001) (data not shown). Furthermore, the majority (9/13; 69%) of proliferative responses detected did
not have any corresponding responses measured in the over-night assay. Where a corresponding overover-night function was present, it was often IL-2 (3/4 [75%] [1 LTNP and 2 NP]). However, the majority (22/25) of the IL-2 responses did not have a corresponding proliferation response, and the percent-age of proliferating cells was inversely correlated with the magnitude of the IL-2 response (r⫽ ⫺0.72;P⬍0.0001) (data not shown).
To determine if the addition of extra parameters would lead to the definition of new epitopes, we compared epitopes iden-tified in our study with previously characterized epitopes. Of the 50 epitopes identified in this study, 12 (24%) are consid-ered best-defined epitopes (BDE) or can be found within a longer BDE (10 to 15 aa) (19), while the remaining epitopes have not been fully characterized (Table 7). To elucidate pos-sible sequence variations that may have led to the detection of novel epitopes, we compared the clade A1 ancestral sequence used in this study and the clade B consensus sequence used in previous epitope-mapping studies (Table 7) (19). Many of the previously uncharacterized epitopes (13/38; 33%) had ⱖ1 amino acid variation between the study sequence and the clade B consensus sequence, possibly accounting for their lack of recognition in previous studies. However, 100% of the 12 rec-ognized BDE also hadⱖ1 amino acid variation between the two sequences. Therefore, sequence differences are likely not the reason why these new epitopes have not been previously defined, and the addition of functional readouts could increase the number of HIV epitopes that are defined.
Interestingly, polyfunctionality (defined asⱖ2⫹responses) comprised 23/69 (33%) of the responses (Table 7). Both BDE and the newly described epitopes frequently stimulated poly-functional responses (5/17 and 18/52, respectively;P ⫽0.59) and responses in the absence of IFN-␥(11/17 and 39/52, re-spectively; P ⫽ 0.35). Interestingly, the uncharacterized epitopes were solely responsible for the proliferation responses that were observed (n⫽13). These data suggest not only that it is important to measure multiple functions but also that the measurement of responses at multiple time points is critical.
This polyfunctional epitope-mapping approach revealed a further disconnect between IFN-␥and proliferative responses. As one might predict from the pool data, individual epitopes often preferentially elicited IFN-␥and not proliferation, and visa versa. This is shown for subject ML1211 for pool 3 and pools 7 and 10, respectively (Fig. 3a and b) and for the decon-voluted peptides (Fig. 3b), with peptide 86 eliciting a polyfunc-tional 4⫹response, while two peptides from this subject (pep-tides 66 and 186) were positive for proliferation but none of the short-term parameters.
In addition, we observed numerous examples of individual epitopes selectively expressing one or more overnight param-TABLE 5. Correlations between short-term parameters stratified by
disease status
Parameter
Disease status of subjects
rvalue,Pvaluea
MIP-1 IL-2 TNF-␣
IFN-␥ LTNP 0.14, 0.1691 0.05, 0.6345 0.14, 0.1403 NP ⫺0.16, 0.0734 ⫺0.09, 0.3344 0.26,0.0030
MIP-1 LTNP 0.017, 0.8736 0.34,0.0003 NP ⴚ0.27,0.0056 ⫺0.10, 0.2857
IL-2 LTNP 0.33,0.0004
NP ⫺0.11, 0.2701
a
[image:6.585.300.542.89.143.2]Boldface type indicates a significant association. Shading indicates associa-tions that differ between LTNP and NP.
TABLE 6. Correlations between short-term parameters and proliferation stratified by disease status
Proliferation in subject
type
rvalue,Pvaluea
IFN-␥ MIP-1 IL-2 TNF-␣
LTNP ⫺0.10, 0.2828 ⴚ0.41,<0.0001 ⴚ0.51,<0.0001 ⴚ0.30,0.0010
NP ⫺0.07, 0.4207 ⴚ0.32,0.0007 ⴚ0.41,<0.0001 ⴚ0.32,0.0003
aBoldface type indicates a significant association.
on November 7, 2019 by guest
http://jvi.asm.org/
[image:6.585.43.284.90.199.2]TABLE 7. Fifty identified epitopes and their associated response profilesa
B cons (top) vs A1 anc (bottom)
HXB2 p24 position
Sequence variation
(no. of aa differences)b
Responder type
(n⫽16)
Presence of response
Proliferation Polyfunctional
IFN-␥ MIP-1 IL-2 TNF-␣
---A--- 3–11 1 LTNP ⫹ No
VQNLQGQMV
---AI 7–14 2 LTNP ⫹ ⫹ ⫹ ⫹ Yes
QGQMVHQSL NP ⫹ No
---AI---- 12–20 2 LTNP ⫹ No
HQSLSPRTL NP ⫹ No
LTNP ⫹ ⫹ ⫹ ⫹ Yes
AI--- 14–22 2 ART ⫹ No
SLSPRTLNA NP ⫹ ⫹ Yes
I--- 15–23 1 LTNP ⫹ ⫹ ⫹ Yes
LSPRTLNAW
--- 16–24 0 ART ⫹ No
SPRTLNAWV
--- 28–36 0 NP ⫹ ⫹ Yes
EEKAFSPEV LTNP ⫹ No
--- 30–38 0 ART ⫹ ⫹ ⫹ Yes
KAFSPEVIP
--- 32–40 0 LTNP ⫹ No
FSPEVIPMF LTNP ⫹ No
--- 36–44 0 LTNP ⫹ No
VIPMFSALS
--- 38–46 0 LTNP ⫹ No
PMFSALSEG NP ⫹ No
LTNP ⫹ No
--- 40–48 0 LTNP ⫹ No
FSALSEGAT
--- 43–51 0 NP ⫹ No
LSEGATPQD
---T 46–54 1 LTNP ⫹ No
GATPQDLNM LTNP ⫹ No
NP ⫹ ⫹ ⫹ Yes
---T-- 48–56 1 ART ⫹ No
TPQDLNMML
--T---T-- 52–60 2 ART ⫹ No
LNMMLNIVG NP ⫹ ⫹ ⫹ Yes
T---T---- LTNP ⫹ No
MMLNIVGGH NP ⫹ No
--T--- 56–64 2 LTNP ⫹ ⫹ Yes
LNIVGGHQA ART ⫹ No
--- 62–70 0 LTNP ⫹ No
HQAAMQMLK NP ⫹ No
--- 75–83 0 LTNP ⫹ No
EEAAEWDRL
--- 76–84 0 NP ⫹ ⫹ ⫹ Yes
EAAEWDRLH
Continued on following page
on November 7, 2019 by guest
http://jvi.asm.org/
TABLE 7—Continued
B cons (top) vs A1 anc (bottom)
HXB2 p24 position
Sequence variation
(no. of aa differences)b
Responder type
(n⫽16)
Presence of response
Proliferation Polyfunctional
IFN-␥ MIP-1 IL-2 TNF-␣
--- 77–85 0 ART ⫹ No
AAEWDRLHP
--- 82–90 0 LTNP ⫹ No
RLHPVHAGP
---A- 86–94 1 ART ⫹ No
VHAGPIPPG
--- 93–101 0 ART ⫹ No
PGQMREPRG
--- 96–104 0 LTNP ⫹ No
MREPRGSDI ART ⫹ No
--- 102–110 0 NP ⫹ No
SDIAGTTST
--- 104–112 0 ART ⫹ ⫹ ⫹ Yes
IAGTTSTLQ
--- 106–114 0 LTNP ⫹ ⫹ Yes
GTTSTLQEQ
--- 110–118 0 NP ⫹ ⫹ Yes
TLQEQIGWM
---N 112–120 1 LTNP ⫹ ⫹ Yes
QEQIGWMTG 119–127 1 ART ⫹ ⫹ Yes
--- 131–139 0 LTNP ⫹ No
KRWIILGLN
--- 133–141 0 LTNP ⫹ ⫹ ⫹ Yes
WIILGLNKI
--- 136–144 0 LTNP ⫹ No
LGLNKIVRM ART ⫹ No
---T--- 143–151 1 LTNP ⫹ No
RMYSPVIL
--- 150–158 0 LTNP ⫹ ⫹ Yes
ILDIRQGPK
--- 157–165 0 LTNP ⫹ No
PKEPFRDYV
---Y-- 163–171 1 NP ⫹ No
DYVDRFFKT
---Y--- 166–174 1 NP ⫹ ⫹ Yes
DRFFKTLRA
Y--- 169–177 1 LTNP ⫹ ⫹ ⫹ Yes
FKTLRAEQA
--- 187–195 0 NP ⫹ No
ETLLVQNAN
--- 190–198 0 NP ⫹ No
LVQNANPDC
---T--K 195–203 2 NP ⫹ ⫹ ⫹ Yes
NPDCKSILR
Continued on following page
on November 7, 2019 by guest
http://jvi.asm.org/
eters. Data for subject ML1932 (Fig. 3c and d) demonstrate the functional disparity of CD8 overnight responses at both the pool and epitope levels. This subject responded with mono-functional responses to 3/5 responsive pools (MIP-1 re-sponses to pool 14 and IL-2 rere-sponses to pools 5 and 11) but also had polyfunctional responses to pools 7 and 16, with the coexpression of IFN-␥and MIP-1. This participant showed similar selectivenesses in expression at the peptide level, with
[image:9.585.42.544.81.325.2]responses to peptides 202, 203, and 208 being monofunctional and independent from all other parameters (MIP-1⫹ and TNF-␣, respectively). Several polyfunctional responses were also seen: peptide 209 was IFN-␥⫹IL-2⫹, while peptide 210 was MIP-1⫹TNF-␣⫹. These data further show that the func-tional readout used to define CD8⫹T cell activity can play a major role in determining the epitope specificity of HIV-spe-cific CD8⫹T cells.
TABLE 7—Continued
B cons (top) vs A1 anc (bottom)
HXB2 p24 position
Sequence variation
(no. of aa differences)b
Responder type
(n⫽16)
Presence of response
Proliferation Polyfunctional
IFN-␥ MIP-1 IL-2 TNF-␣
---T--K-- 197–205 2 NP ⫹ No
DCKSILRAL
K----A--- 203–211 2 NP ⫹ ⫹ Yes
RALGPGATL
A--- 208–216 1 LTNP ⫹ ⫹ Yes
GATLEEMMT
--- 211–219 0 NP ⫹ ⫹ ⫹ Yes
LEEMMTACQ
--- 212–220 0 NP ⫹ ⫹ No
EEMMTACQG
--- 214–222 0 NP ⫹ No
MMTACQGVG NP ⫹ No
--- 215–223 0 NP ⫹ No
MTACQGVGG
--- 222–230 0 NP ⫹ No
GGPGHKARV
a
Shown is a comparison of B consensus (B cons) sequences with recognized peptides from the A1 uncestral (A1 anc) library used in the current study. Shading indicates epitopes that are considered best-defined epitopes (BDE) or can be found within a longer, 10-aa to 15-aa BDE (23).
b
Number of amino acid differences between the sequences noted.
FIG. 3. HIV-specific CD8⫹T cell specificity differs depending on the effector readout, as shown for a representative subject. (A) Pool-specific responses for ML1211. Proliferation responses were measured at day 6, and four different readouts were measured after overnight stimulation. This subject has divergent long-term and short-term responses, including proliferation only (pool 10) and 4⫹without proliferation (pool 3). Pool 15 is an example of a 4⫹polyfunctional response with accompanying proliferation. (B) Deconvoluted peptide responses for subject ML1211. Peptide 86 elicited a polyfunctional 4⫹response, while two peptides, peptides 66 and 186, preferentially elicited a proliferation-only response. (C) Pool-specific responses for subject ML1932. Many responses were 1⫹, with an MIP-1response to pool 14 and an IL-2 response to pools 5 and 11. Polyfunctional responses to pools 7 and 16 were seen with the concurrent expression of IFN-␥and MIP-1. (D) Deconvoluted peptide responses for subject ML1932. Responses to peptides 202, 203, and 208 were 1⫹, with an MIP-1response and a TNF-␣response, respectively. Peptides 209 and 210 elicited a 2⫹response, with an IFN-␥⫹IL-2⫹response and an MIP-1⫹TNF-␣⫹response, respectively.
on November 7, 2019 by guest
http://jvi.asm.org/
[image:9.585.47.542.489.642.2]DISCUSSION
Recent advances in immunology have allowed the opportu-nity to redefine the breadth and specificity of the large, com-plex antiviral CD8⫹ T cell responses, expanding our under-standing of protective immunity. For example, acute CD8⫹T cell responses to smallpox and yellow fever virus vaccinations, measured by cellular activation, intracellular cytokine staining, and tetramer frequencies, revealed that these responses are of a much higher magnitude than was previously appreciated (34). While assessments of HIV-specific responses have tradi-tionally relied on assays that measure IFN-␥ secretion, this cytokine does not always correlate with protective immunity, nor does it accurately describe the full breadth of HIV-specific CD8⫹T cell responses (15, 26, 45). Other studies have dem-onstrated that HIV⫹ LTNP maintain a more functional re-sponse than do those who progress to AIDS (1, 5), but whether these responses differ in specificity remains largely unexplored. Here we have mapped CD8⫹T cell epitopes more compre-hensively than has been previously described, using multiple overnight parameters and long-term proliferation assays. Crit-ically, over two-thirds of the CD8 responses detected in our study were detected in the absence of a parallel IFN-␥ re-sponse (50/69 rere-sponses). These data suggest that previous epitope-mapping studies may have missed out on important immune responses by focusing solely on IFN-␥. Our data fur-ther reveal that CD8⫹T cell responses are complex and that particular epitopes within a given individual can be associated with different functional profiles. Identifying and understand-ing epitopes that elicit polyfunctional and proliferative re-sponses will serve as a valuable resource for vaccine design and ensure that wider verities of protective responses are obtained. By epitope mapping using multiple parameters, we identi-fied 50 epitopes in HIV-1 p24, 38 of which have not yet been fully characterized according to the Los Alamos HIV Immu-nology Database. A number of factors could explain the iden-tification of these novel epitopes. For one, we used shorter peptides than most prior studies (9-mer peptides compared to 15-mer peptides), which have an increased sensitivity to detect responses (3, 13, 14, 41). Second, the database of optimal CD8⫹ T cell HIV epitopes may be biased toward clade B, while this epitope-mapping study was conducted in Kenya, where clade A1 predominates (12, 32, 35, 36). Although the use of the clade A1 ancestral sequence may be a contributing factor leading to the identification of novel epitopes, we also found that the 12 previously defined epitopes could also be recognized between clades, despite sequence differences. A third, and perhaps more important, factor is the inclusion of a greater number of functional parameters as readouts to define responses. The measurement of four functional parameters (IFN-␥, MIP-1, IL-2, and TNF-␣) plus proliferation ex-panded the breadths of the detected responses by over 3.5-fold. The majority of novel epitopes identified in this study did not include IFN-␥(39/52 responses), 13 of which elicited prolifer-ation responses. Interestingly, a number of these previously unidentified epitopes induced polyfunctional (ⱖ2⫹) responses (18/52) but not necessarily IFN-␥. One caveat of our study, however, was that the longer incubation time was not opti-mized to detect IL-2 and not IFN-␥. It is possible that we would have detected more epitope-specific responses
pro-ducing IFN-␥, had we used a standard 6-h assay. However, together, these data support the idea that the breadth of HIV-specific immunity may be even greater than previously appreciated.
There was a striking discordance between responses mea-sured in overnight assays and those meamea-sured in proliferation assays, with CD8⫹ T cells responding to epitopes by either secreting a cytokine or proliferating but rarely both. This sug-gests that a given epitope in a subject may preferentially induce either effector memory CD8⫹ T cell (TEM)-like (IFN-␥) or
central memory CD8⫹ T cell (TCM)-like (proliferative) re-sponses. This observation was not limited to IFN-␥but was apparent for all short-term readouts, as proliferation responses were inversely correlated with IFN-␥, MIP-1, and TNF-␣, all associated with TEM responses. This was even more nounced at the epitope level, where the vast majority of pro-liferation responses (9/13) were completely independent of overnight parameters. While an explanation for these data remains unclear, one hypothesis is that the cells that respond in an overnight assay are not equipped to survive for the length of a proliferation assay, which is more likely to identify cells that need time to become activatedin vitro. CD8⫹T cell survival may be an important precursor of proliferation, whereas the short-term expression of cytokines, particularly IL-2, may not require survival to the same extent. Some studies suggested that survival capabilities, measured by exhaustion markers, might be the key attribute of a TCMcell (9).
Contrary to what was observed in a previous study (48), our data consistently showed an inverse correlation between IL-2 and proliferation, which rarely corresponded. A possible ex-planation for these differences is that previous studies stimu-lated cells for 6 h to measure IL-2, whereas we measured this cytokine in 14-h assays. However, another important difference between our data and those of Zimmerli et al. (48) is that our correlations were based on an epitope-screening approach that used IL-2 and proliferation to define CD8⫹T cell specificity, compared to measuring IL-2⫹ CD8⫹ T cells that were also IFN-␥⫹. Since IL-2 and proliferation were both inversely cor-related with IFN-␥in our cohort, it is possible that by focusing solely on IFN-␥ responses, IL-2 and proliferation responses independent of IFN-␥may have been missed in previous stud-ies. A lack of an association between proliferation and IL-2 in the bulk of Gag-specific CD8⫹T cells was recently observed for a Chinese cohort, supporting our findings (29).
Virus-specific CD8⫹ T cells in chronic infections such as HIV experience substantial functional exhaustion. It has been suggested that this occurs in a hierarchical manner as the infection persists, where IL-2, cytolysis, and proliferation are the first to be lost, followed by the loss of TNF-␣and, finally, by the loss of IFN-␥and anergy/deletion (28, 42, 43, 46). Based on this model, the reliance on IFN-␥to screen immune re-sponses may overestimate the effectiveness of the immune response by measuring a significant proportion of CD8⫹ T cells that may be in the final stages of exhaustion. However, our data also challenge this model by finding many CD8⫹T cell responses that are independent of IFN-␥, including the display of IL-2 and proliferation in isolation from other func-tions. Many functional divisions in epitope specificity were seen among the overnight parameters. An examination of all responses clearly shows that epitopes can stimulate many
on November 7, 2019 by guest
http://jvi.asm.org/
binations of responses, often in the absence of IFN-␥. These data suggest that in a chronic viral infection, CD8⫹ T cell responses can be complex and heterogeneous.
In agreement with data from previous work, our study found that LTNP possess a more polyfunctional HIV-specific CD8⫹ T cell response than do NP (5). However, our polyfunctional profile is slightly different from those reported by previous studies, due to a longer incubation time, which likely underes-timates MIP-1responses that are optimally detected at 6 h. LTNP also responded with stronger p24-specific proliferation, IL-2, and TNF-␣responses than did NP. These data are con-sistent with data from other studies which found the level of proliferation to be significantly lower in progressive chronic infection than primary infection or LTNP (30) and that re-sponses to p24 may be an important target for the control of HIV replication (22). In addition, LTNP demonstrated more consistent associations between functional parameters than did progressors, including the rarely observed coexpression of pro-liferation and IFN-␥ responses. This may be important for HIV vaccines, based on a recent study with nonhuman pri-mates which showed that macaques vaccinated with a CMV vector expressing simian immunodeficiency virus (SIV) had robust TEMresponses, characterized by polyfunctionality, and were less likely to become infected following low-dose chal-lenge (23). However, the authors of that study stressed that the parallel induction of TCMresponses, similar to what occurs in
many LTNP, may provide a second line of defense in case infection becomes established.
In summary, we show that within a given HIV-infected sub-ject, it is possible to identify novel epitopes that induce qual-itatively distinct CD8⫹ T cell responses. Screening for re-sponses by the measurement of five CD8⫹T cell functions revealed a greater complexity of CD8⫹T cell immunology and described a greater breadth of response than would have been derived from the measurement of any one response on its own. We have shown that epitope specificity differs between short-and long-term assays as well as within the short-term assay. A multiparametric approach to defining epitope specificity is one that could be used in vaccine evaluations in order to deter-mine the true immunogenicity of a given vaccine candidate. Moreover, because polyfunctionality and proliferative re-sponses have been associated with an attenuated HIV-1 dis-ease course, these data and similarly comprehensive epitope-mapping studies could identify a number of additional epitopes that may serve as useful targets for successful HIV-1 vaccines that will presumably induce a wider range of epitope-specific responses.
ACKNOWLEDGMENTS
We thank the study participants and the staff at the Majengo clinic (1985 to present) for their dedication and perseverance. We also thank Mario Roederer at the Vaccine Research Center, NIAID, NIH, for providing Deconvolute This! and SPICE software. F.P. is a Tier I Canada Research Chair in susceptibility and resistance to infection.
M.R. is supported by the MHRC, CIHR, and CIHR IID and GH training program. This work was supported by Canadian Institute of Health Research grant HOP 86192. We declare that we have no conflicts of interest.
M.R., L.R.M., S.A.K.K., and T.B.B. conceived and designed the experiments. M.R. and L.R.M. performed the experiments. M.R. and L.R.M. analyzed the data. M.R., L.R.M., S.A.K.K., and T.B.B. wrote
the paper. J.K., C.W., M.K., F.A.P., and T.B.B. performed clinical work and cohort maintenance.
REFERENCES
1.Almeida, J. R., et al.2007. Superior control of HIV-1 replication by CD8⫹ T cells is reflected by their avidity, polyfunctionality, and clonal turnover. J.
Exp. Med.204:2473–2485.
2.Anzala, O. A., et al.1995. Rapid progression to disease in African sex workers with human immunodeficiency virus type 1 infection. J. Infect. Dis.
171:686–689.
3.Beattie, T., et al.2004. Screening for HIV-specific T-cell responses using
overlapping 15-mer peptide pools or optimized epitopes. AIDS18:1595–
1598.
4.Bennett, M. S., H. L. Ng, A. Ali, and O. O. Yang.2008. Cross-clade detection of HIV-1-specific cytotoxic T lymphocytes does not reflect cross-clade
anti-viral activity. J. Infect. Dis.197:390–397.
5.Betts, M. R., et al.2006. HIV nonprogressors preferentially maintain highly
functional HIV-specific CD8⫹T cells. Blood107:4781–4789.
6.Beveridge, N. E., et al.2007. Immunisation with BCG and recombinant MVA85A induces long-lasting, polyfunctional Mycobacterium
tuberculosis-specific CD4⫹memory T lymphocyte populations. Eur. J. Immunol.37:
3089–3100.
7.Borrow, P., H. Lewicki, B. H. Hahn, G. M. Shaw, and M. B. Oldstone.1994.
Virus-specific CD8⫹cytotoxic T-lymphocyte activity associated with control
of viremia in primary human immunodeficiency virus type 1 infection. J.
Vi-rol.68:6103–6110.
8.Borrow, P., et al.1997. Antiviral pressure exerted by HIV-1-specific cytotoxic T lymphocytes (CTLs) during primary infection demonstrated by rapid
se-lection of CTL escape virus. Nat. Med.3:205–211.
9.Chomont, N., et al.2009. HIV reservoir size and persistence are driven by T
cell survival and homeostatic proliferation. Nat. Med.15:893–900.
10.Darrah, P. A., et al.2007. Multifunctional TH1 cells define a correlate of
vaccine-mediated protection against Leishmania major. Nat. Med.13:843–
850.
11.Day, C. L., et al.2007. Proliferative capacity of epitope-specific CD8 T-cell responses is inversely related to viral load in chronic human
immunodefi-ciency virus type 1 infection. J. Virol.81:434–438.
12.Dowling, W. E., et al.2002. Forty-one near full-length HIV-1 sequences from Kenya reveal an epidemic of subtype A and A-containing recombinants.
AIDS16:1809–1820.
13.Draenert, R., et al.2003. Comparison of overlapping peptide sets for detec-tion of antiviral CD8 and CD4 T cell responses. J. Immunol. Methods
275:19–29.
14.Draenert, R., et al.2004. Impact of intrapeptide epitope location on CD8 T cell recognition: implications for design of overlapping peptide panels. AIDS
18:871–876.
15.Draenert, R., et al.2004. Immune selection for altered antigen processing leads to cytotoxic T lymphocyte escape in chronic HIV-1 infection. J. Exp.
Med.199:905–915.
16.Draenert, R., et al.2004. Persistent recognition of autologous virus by high-avidity CD8 T cells in chronic, progressive human immunodeficiency virus
type 1 infection. J. Virol.78:630–641.
17.Erickson, A. L., et al.2008. Potentially exposed but uninfected individuals produce cytotoxic and polyfunctional human immunodeficiency virus type
1-specific CD8(⫹) T-cell responses which can be defined to the epitope level.
Clin. Vaccine Immunol.15:1745–1748.
18.Fowke, K. R., et al.1996. Resistance to HIV-1 infection among persistently
seronegative prostitutes in Nairobi, Kenya. Lancet348:1347–1351.
19.Frahm, N., B. Baker, and C. Brander. 2008. Identification and optimal definition of HIV-derived cytotoxic T lymphocyte (CTL) epitopes for the
study of CTL escape, functional avidity and viral evolution, p. 3–24.In
B. T. M. Korber, et al. (ed.), HIV molecular immunology 2008. Los Alamos National Laboratory, Theoretical Biology and Biophysics, Los Alamos, NM. 20.Gea-Banacloche, J. C., et al.2000. Maintenance of large numbers of
virus-specific CD8⫹T cells in HIV-infected progressors and long-term
nonpro-gressors. J. Immunol.165:1082–1092.
21.Goulder, P. J., et al.1997. Late escape from an immunodominant cytotoxic T-lymphocyte response associated with progression to AIDS. Nat. Med.
3:212–217.
22.Goulder, P. J., and D. I. Watkins.2008. Impact of MHC class I diversity on immune control of immunodeficiency virus replication. Nat. Rev. Immunol.
8:619–630.
23.Hansen, S. G., et al.2009. Effector memory T cell responses are associated with protection of rhesus monkeys from mucosal simian immunodeficiency
virus challenge. Nat. Med.15:293–299.
24.Horton, H., et al.2006. Preservation of T cell proliferation restricted by protective HLA alleles is critical for immune control of HIV-1 infection.
J. Immunol.177:7406–7415.
25.Jin, X., et al.1999. Dramatic rise in plasma viremia after CD8(⫹) T cell depletion in simian immunodeficiency virus-infected macaques. J. Exp. Med.
189:991–998.
on November 7, 2019 by guest
http://jvi.asm.org/
26.Klenerman, P., Y. Wu, and R. Phillips. 2002. HIV: current opinion in
escapology. Curr. Opin. Microbiol.5:408–413.
27.Koup, R. A., et al.1994. Temporal association of cellular immune responses with the initial control of viremia in primary human immunodeficiency virus
type 1 syndrome. J. Virol.68:4650–4655.
28.Kristensen, N. N., J. P. Christensen, and A. R. Thomsen.2002. High
num-bers of IL-2-producing CD8⫹T cells during viral infection: correlation with
stable memory development. J. Gen. Virol.83:2123–2133.
29.Li, H., et al.2009. Proliferation, but not interleukin 2 production, of
Gag-specific CD8⫹T cells is associated with low HIV viremia and high CD4
counts in HIV-1-infected Chinese individuals. J. Acquir. Immune Defic.
Syndr.52:1–8.
30.Lichterfeld, M., et al.2004. Loss of HIV-1-specific CD8⫹T cell proliferation after acute HIV-1 infection and restoration by vaccine-induced
HIV-1-spe-cific CD4⫹T cells. J. Exp. Med.200:701–712.
31.McKinnon, L. R., et al.2008. Substantial intrapatient differences in the
breadth and specificity of HIV-specific CD8⫹T-cell interferon-gamma and
proliferation responses. J. Acquir. Immune Defic. Syndr.49:123–127.
32.McKinnon, L. R., et al.2009. Epitope mapping of HIV-specific CD8⫹T cells
in a cohort dominated by clade A1 infection. PLoS One4:e6965.
33.Migueles, S. A., et al. 2002. HIV-specific CD8⫹T cell proliferation is coupled to perforin expression and is maintained in nonprogressors. Nat.
Immunol.3:1061–1068.
34.Miller, J. D., et al.2008. Human effector and memory CD8⫹T cell
re-sponses to smallpox and yellow fever vaccines. Immunity28:710–722.
35.Neilson, J. R., et al.1999. Subtypes of human immunodeficiency virus type 1
and disease stage among women in Nairobi, Kenya. J. Virol.73:4393–4403.
36.Peters, H. O., et al.2008. An integrative bioinformatic approach for studying escape mutations in human immunodeficiency virus type 1 gag in the
Pum-wani sex worker cohort. J. Virol.82:1980–1992.
37.Precopio, M. L., et al.2007. Immunization with vaccinia virus induces
poly-functional and phenotypically distinctive CD8(⫹) T cell responses. J. Exp.
Med.204:1405–1416.
38.Price, D. A., et al.1997. Positive selection of HIV-1 cytotoxic T lymphocyte escape variants during primary infection. Proc. Natl. Acad. Sci. U. S. A.
94:1890–1895.
39.Roederer, M., and R. A. Koup.2003. Optimized determination of T cell
epitope responses. J. Immunol. Methods274:221–228.
40.Schmitz, J. E., et al.1999. Control of viremia in simian immunodeficiency
virus infection by CD8⫹lymphocytes. Science283:857–860.
41.Sekaly, R. P.2008. The failed HIV Merck vaccine study: a step back or a
launching point for future vaccine development? J. Exp. Med.205:7–12.
42.Shin, H., and E. J. Wherry.2007. CD8 T cell dysfunction during chronic viral
infection. Curr. Opin. Immunol.19:408–415.
43.Streeck, H., et al.2008. Antigen load and viral sequence diversification
determine the functional profile of HIV-1-specific CD8⫹T cells. PLoS Med.
5:e100.
44.Streeck, H., N. Frahm, and B. D. Walker.2009. The role of IFN-gamma
Elispot assay in HIV vaccine research. Nat. Protoc.4:461–469.
45.Valentine, L. E., et al.2008. Recognition of escape variants in ELISPOT
does not always predict CD8⫹T-cell recognition of simian
immunodefi-ciency virus-infected cells expressing the same variant sequences. J. Virol.
82:575–581.
46.Wherry, E. J., J. N. Blattman, K. Murali-Krishna, R. van der Most, and R. Ahmed.2003. Viral persistence alters CD8 T-cell immunodominance and tissue distribution and results in distinct stages of functional impairment.
J. Virol.77:4911–4927.
47.Yang, O. O.2003. Will we be able to ‘spot’ an effective HIV-1 vaccine?
Trends Immunol.24:67–72.
48.Zimmerli, S. C., et al.2005. HIV-1-specific IFN-gamma/IL-2-secreting CD8 T cells support CD4-independent proliferation of HIV-1-specific CD8 T
cells. Proc. Natl. Acad. Sci. U. S. A.102:7239–7244.