0022-538X/05/$08.00⫹0 doi:10.1128/JVI.79.14.9046–9053.2005
Copyright © 2005, American Society for Microbiology. All Rights Reserved.
Structural and Functional Analysis of the
cis
-Acting Elements
Required for Plus-Strand RNA Synthesis of
Bamboo Mosaic Virus
Jen-Wen Lin,† Hsiao-Ning Chiu,† I-Hsuan Chen, Tzu-Chi Chen, Yau-Heiu Hsu,
and Ching-Hsiu Tsai*
Graduate Institute of Biotechnology, National Chung Hsing University, Taichung 402, Taiwan
Received 6 December 2004/Accepted 13 March 2005
Bamboo mosaic virus(BaMV) has a single-stranded positive-sense RNA genome. The secondary structure of
the 3ⴕ-terminal sequence of the minus-strand RNA has been predicted by MFOLD and confirmed by enzymatic structural probing to consist of a large, stable stem-loop and a small, unstable stem-loop. To identify the promoter for plus-strand RNA synthesis in this region, transcripts of 39, 77, and 173 nucleotides (Ba-39, Ba-77, and Ba-173, respectively) derived from the 3ⴕterminus of the minus-strand RNA were examined by an in vitro RNA-dependent RNA polymerase assay for the ability to direct RNA synthesis. Ba-77 and Ba-39 appeared to direct the RNA synthesis efficiently, while Ba-173 failed. Ba-77/⌬5, with a deletion of the 3ⴕ-terminal UUUUC sequence in Ba-77, directed the RNA synthesis only to 7% that of Ba-77. However, Ba-77/⌬16 and Ba-77/⌬31, with longer deletions but preserving the terminal UUUUC sequence of Ba-77, restored the template activity to about 60% that of the wild type. Moreover, mutations that changed the sequence in the stem of the large stem-loop interfered with the efficiency of RNA synthesis and RNA accumulation in vivo. The mutant with an internal deletion in the region between the terminal UUUUC sequence and the large stem-loop reduced the viral RNA accumulation in protoplasts, but mutants with insertions did not. Taken together, these results suggest that threecis-acting elements in the 3ⴕend of the minus-strand RNA, namely, the terminal UUUUC sequence, the sequence in the large stem-loop, and the distance between these two regions, are involved in modulating the efficiency of BaMV plus-strand viral RNA synthesis.
There are at least two major steps involved in the replication of positive-sense viral RNA: the first is the synthesis of the complementary strand of genomic RNA, which is followed by the generation of progeny RNAs using the newly synthesized minus-strand RNA templates (37). For these reactions to pro-ceed faithfully, the 3⬘ ends of both plus- and minus-strand RNAs must contain important elements, the sequences and/or structures needed to direct the replicase to initiate the RNA synthesis (7, 18). The 3⬘-terminal secondary structures of
Al-falfa mosaic virusRNAs (12, 13, 33),Turnip crinkle virusRNAs
(39), andTomato bush stunt virus defective interfering RNA (8) and the 3⬘ tRNA-like structures (TLS) of Brome mosaic
virus(BMV),Tobacco mosaic virus, andTurnip yellow mosaic
virus(TYMV) RNAs have been identified to be important for
minus-strand RNA synthesis (5, 6, 17, 26, 36).
Bamboo mosaic virus(BaMV) is a single-stranded
positive-sense RNA virus. Its 6,366-nucleotide (nt) genome [excluding the poly(A) tail] contains a 5⬘ m7GpppG structure and a 3⬘ poly(A) tail (24). Five open reading frames (ORF) encoding polypeptides of 155, 28, 13, 6, and 25 kDa could be identified in the genome. Besides the genomic RNA which encodes the 155-kDa protein comprising a capping enzyme domain (21, 22), a 5⬘ triphosphatase and helicase-like domain (9, 11, 16), and an RNA-dependent RNA polymerase (RdRp) activity
do-main (1, 19, 20), two major 3⬘-coterminal subgenomic RNAs of 2.0 kb and 1.0 kb in length encode the proteins for virus movement and encapsidation, respectively (23). ORF 2 to 4 are overlapping and are known as the “triple gene block” required for viral movement, as demonstrated withWhite
clo-ver mosaic virus(2). ORF 5 encodes the 25-kDa coat protein.
The tertiary structure of the 3⬘untranslated region (UTR) of BaMV has been identified to comprise a cloverleaf-like struc-ture, with a large stem-loop with a bulge and an internal loop, and a pseudoknot domain (3, 41). These structures were dem-onstrated to be important for minus-strand RNA synthesis in vitro by use of a partially purified replicase from infected
Nicotiana benthamiana(4). The interactions of the 3⬘UTR of
BaMV RNA with theEscherichia coli-expressed viral RdRp core domain have been demonstrated by electrophoretic mo-bility shift assay and footprinting analysis (15). Mutants with substitutions, deletions, and insertions introduced into stem-loop D failed to replicate efficiently in vivo, indicating that maintaining the integrity of the stem-loop structure is impor-tant for BaMV RNA replication (3). An active replicase com-plex or recombinant RdRp is a useful tool for identifying the RNA sequences and/or secondary structures required for viral RNA synthesis in vitro. A specific region in the 3⬘terminus of
Hepatitis C virus RNA has been shown to interact
preferen-tially with recombinant RdRp (25, 32). The replicase complex isolated from infected plants, containing the virus-encoded RdRp and host factors, has been used to identify the sequences required for efficient transcription in vitro for BMV (31),
Cu-cumber mosaic virus (CMV) (10), Turnip crinkle virus (38),
Tobacco mosaic virus(26), TYMV (5, 36),Potato virus X(29),
* Corresponding author. Mailing address: Graduate Institute of Bio-technology, National Chung Hsing University, Taichung 402, Taiwan. Phone: (886) 4-22840451. Fax: (886) 4-22860260. E-mail: chtsai1@dragon .nchu.edu.tw.
† Both authors contributed equally to this work.
9046
on November 8, 2019 by guest
http://jvi.asm.org/
Cucumber necrosis virus(27, 28), and BaMV (4). Here we have used the enzymatic-structure probing technique to identify the 3⬘-terminal structure of the BaMV minus-strand RNA. The biological function of this structure was examined by an in vitro RdRp activity assay and an in vivo protoplast inoculation anal-ysis.
MATERIALS AND METHODS
Mutant construction and RNA preparation.Short fragments containing a T7 promoter and sequences derived from the full-length infectious cDNA clone pBaMV40A were generated by PCR amplification for minus-strand RNA syn-thesis. Minus-strand RNA transcripts with 3⬘cotermini (39, 77, and Ba-173) of three different sizes (39, 77, and 173 nt, respectively), numbered from the first nucleotide of the 3⬘end of the minus-strand RNA (Fig. 1), were synthesized from the PCR-amplified DNA fragments with the upstream primer BaMV 5⬘(⫹) (5⬘GAAAACCACTCCAAACGAAA3⬘) and the downstream primers BaMV/ T7(⫺)39 (5⬘TAATACGACTCACTATAGGGTCTTTTCTTTTCTTTTCTTTT CG3⬘, T7 promoter is underlined), BaMV/T7(⫺)77 (5⬘TAATACGACTCACT ATAGGGCGATTGTAG3⬘), and BaMV/T7(⫺)173 (5⬘TAATACGACTCACT ATAGGGAGTGCGCT TCCTCTT3⬘), respectively. The other three deletion mutants of the minus-strand RNA transcripts with 5⬘ cotermini (Ba-77/⌬5, Ba-77/⌬16, and Ba-77/⌬31) were synthesized by the same method with primers BaMV/T7(⫺)77 and BaMV5⬘(⫹5) (5⬘CCACTCCAAACGAAAAGA3⬘), BaMV5⬘
(⫹16) (5⬘GAAAAGAAAAGAAAAGAAAAGACGAA3⬘), and BaMV5⬘(⫹31) (5⬘GAAAAGACGAAGAAAACAAA3⬘), respectively. To generate mutants of Ba-77 with abolished lower stems (LM mutants), abolished upper stems (UM mutants), compensatory restored lower stems (LR mutants), and compensatory restored upper stems (UR mutants), four sets of primer pairs were used: BaMV 5⬘(⫹)51mer (5⬘GAAAACCACTCCAAACGAAAAGAAAAGAAAAGAAAA GACGAAGAAAACAAA3⬘) and BaMV T7(⫺)72 M (5⬘TAATACGACTCAC TATAGGTGCTTTGTAGGGGACAATTGGCAGTTTGTTTTCT3⬘) for the LM mutant; BaMV 5⬘(⫹) 41R (5⬘GAAAACCACTCCAAACGAAAAGAAAA GAAAAGAAAAGCGCTAGAAAACAAA3⬘) and BaMV T7(⫺)72 M for the LR mutant; BaMV 5⬘(⫹)51mer and BaMV T7(⫺)60 M (5⬘TAATACGACTCA CTATAGGGCGATTGTAGGGGTGTATTGGCAGTTTGTTTTCTTCGT3⬘) for the UM mutant; and BaMV 5⬘(⫹)49R (5⬘GAAAACCACTCCAAACGAA AAGAAAAGAAAAGAAAAGACGAAGAAATGTAACTGCCAA3⬘) and BaMV T7(⫺)60R (5⬘TAATACGACTCACTATAGGGCGATTGTAGGGGT GTATTGGCAGTTACAT3⬘) for the UR mutant. (The bold letters correspond to the mutated positions.) The PCR products were cloned into SmaI-digested pUC119 vectors, and the clones were confirmed by sequencing. These four clones were subsequently used as templates for PCR with primers BaMV 5⬘(⫹) and T7 (5⬘TAATACGACTCACTATAG3⬘) to generate LM, LR, UR, and UM
mutant DNA fragments and for in vitro transcription. An in vitro transcription reaction was carried out at 37°C for 2 h with a 50-l reaction mixture containing 5l T7 RNA polymerase (20 U/l), 80 mM HEPES-KOH (pH 7.5), 12 mM MgCl2, 2 mM spermidine, 10 mM dithiothreitol, 3 mM nucleoside triphosphate,
and 5g DNA template. RNA transcripts were eluted from denaturing (8 M urea) polyacrylamide gels by soaking them in elution buffer (0.5 M ammonium acetate, 10 mM magnesium acetate, 1 mM EDTA, and 0.1% sodium dodecyl sulfate) with shaking overnight at room temperature and precipitated with eth-anol.
End labeling of the RNAs.In order to label the 5⬘ends of the Ba-39 and Ba-77 RNAs, 1.5g of gel-purified transcripts was dephosphorylated with 1.5 U of shrimp alkaline phosphatase at 37°C for 1 h, followed by phenol-chloroform extraction and ethanol precipitation prior to kinase treatment (35). A kinase treatment was performed with a total volume of 10l containing 3l (10Ci/l) [␥-32P]ATP and 5 U of T4 polynucleotide kinase at 37°C for 30 min, followed by
heat inactivation at 65°C for 10 min. Labeled transcripts were separated in a 10% sequencing gel and eluted from the gel by soaking them in elution buffer and shaking them overnight at room temperature.
Structure prediction and mapping of the 3ⴕend of BaMV minus-strand RNA.
The structure of the 3⬘terminus of the BaMV minus-strand RNA was predicted by the MFOLD program (43). The cleavage sites of the structural probe were determined by electrophoretic separation of RNA fragments on denaturing (7 M urea) polyacrylamide gels. The full-length labeled RNAs denatured in boiling water for 2 min and partially digested with alkaline buffer (55.5 mM Na2CO3[pH
9.0], 1.1 mM EDTA) were used as ladder markers. To prepare the sequencing ladders, the labeled RNAs were denatured in boiling water for 90 s following cleavage with RNase T1(10 to 15 U) and for 30 s following cleavage with RNase
A (3 ng) at 55°C and on ice for 10 min. The structural probing was done by digestion of the 5⬘-end-labeled transcripts (50,000 to 70,000 cpm) with RNases A, T1, T2, and V1at 20°C in 60l of digestion buffer (30 mM Tris [pH 7.5], 3 mM
EDTA, 200 mM NaCl, 100 mM LiCl) (40); 10 mM MgCl2was supplemented for
RNase V1digestion.
RdRp activity assay.The detergent-solubilized RdRp preparation used for the exogenous template activity assay was treated with 20g/ml micrococcal nucle-ase (U.S. Biochemicals) in the presence of 2 mM CaCl2at 30°C for 30 min; the
reaction was stopped by adding 16 mM EGTA (4). Subsequently, 20l of RdRp was included in the 50-l reaction mixture containing 10 mM dithiothreitol; 3 mM MgCl2; 2 mM each of ATP, CTP, and GTP; 2M UTP; 66 nM [␣
-32
P]UTP (3,000 Ci/mmol); 300g bentonite; and 5 pmol of RNA templates (4). The reaction mixtures were incubated at 30°C for 1 h, and the reactions were stopped by phenol-chloroform extraction and ethanol precipitation.
In vitro translation.In vitro translation was carried out with a 25-l reaction mixture containing 3g of transcript with a half volume of nuclease-treated rabbit reticulocyte lysate (RRL; Promega) supplemented with all of the amino acids except methionine. Proteins were labeled by the incorporation of 5Ci of
L-[35
S]methionine (1,000 Ci/mmol; Amersham) into the reaction mixture. Incu-bations were performed at 30°C for 1 h and terminated by the addition of the Laemmli sample buffer. Translation products were resolved by 12% sodium dodecyl sulfate-polyacrylamide gel electrophoresis. Gels were fixed, dried, and analyzed by phosphorimager (Fujifilm BAS 1500).
Protoplast inoculation.Protoplasts (4⫻105) prepared fromN. benthamiana
leaves were inoculated with 5g of the transcripts and incubated for 48 h at 25°C under constant light as described previously (41). For Northern blot analysis, total RNAs were extracted from the protoplasts, glyoxalated, electrophoresed through a 1% agarose gel, and transferred to a membrane (Zeta-Probe; Bio-Rad) as described previously (41). The hybridization probes were 0.6- and 0.7-kb
32P-labeled RNA transcripts derived from HindIII-linearized pBaMV-O/SB2.6
(14) and BglI-linearized pBaMV-O (41), complementary to the 3⬘ends of the plus and minus strands of BaMV RNA, respectively. The banding signals were scanned and quantified with a phosphorimager (Fujifilm BAS 1500).
RESULTS
[image:2.585.46.282.73.238.2]Computer prediction of the 3ⴕ-terminal structure of BaMV minus-strand RNA. The 3⬘-terminal structure of the minus strand of the BaMV RNA was predicted by the MFOLD program (43) to consist of two simple stem-loops separated by 8 nucleotides (nt 28 to 35; nucleotides are numbered from the 3⬘end) (Fig. 1). The small stem-loop is positioned at nt 6 to 27 with a U-rich loop. The large stem-loop positioned at nt 36 to 77 comprises a 4-nucleotide bulge (nt 68 to 71), an internal
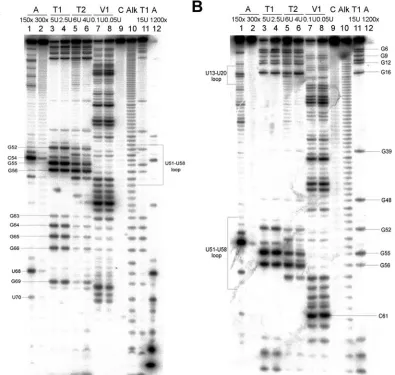
FIG. 1. Secondary structure of the 77 3⬘-terminal nucleotides of the BaMV minus-strand RNA, as predicted by the MFOLD program (43), and summary of the results of enzymatic structural probing. Nucleo-tides are numbered from the 3⬘-end cytosine. The positions of cleav-ages by single- or double-strand RNA-specific enzymes are indicated by symbols explained in the key in the figure.
VOL. 79, 2005 PROMOTER FOR BaMV PLUS-STRAND RNA SYNTHESIS 9047
on November 8, 2019 by guest
http://jvi.asm.org/
loop (nt 44 and 45 on the 3⬘side and nt 64 and 65 on the 5⬘ side), and an 8-nucleotide loop. The sequences GGGCGA77-72 base paired with UCGUCU41-36and AG67-66base paired with CU44-43were together designated the lower stem, and the se-quence GACAA63-59base paired with sequence UUGUU50-44 was designated the upper stem.
Structural probing of the 3ⴕ end of BaMV minus-strand RNA. Enzymatic structural probing of the 5⬘-end-labeled Ba-77 transcript showed that nucleotides U70, U68, U57, C54, U18, C17, and U8, which were predicted to reside in a single-stranded region, were sensitive to RNase A (Fig. 2). The nu-cleotides corresponding to the UUUUC repeats (nt 35 to 23) were shown to yield minor cleavage products with RNase A (Fig. 2). Prominent products were derived at nucleotides G69, G65, G64, G56, G55, G52, and G16 with RNase T1cleavage, implying that they are positioned in a single-strand region. Nucleotides G66, G63, G12, G11, G9, G7, and G6, located at the predicted junction regions of the stem-loop and internal or apical loops, were also sensitive to RNase T1, possibly due to
[image:3.585.100.495.70.445.2]breathing at the junction (Fig. 1 and 2), whereas G48 and G39, located in the stem, were resistant to RNase T1. Cleavage patterns derived from RNases T2and V1, specific for single-and double-strsingle-anded regions in RNA, respectively, did not overlap each other (Fig. 2). Nucleotides in the double-strand regions of the large stem-loop were cleaved efficiently with RNase V1and resistant to RNase T2. Nucleotides U51 to U58 in the apical loop were cleaved by RNase T2but not by RNase V1(Fig. 2). Nucleotides in the bulge (U68 and G69) and the internal loop (G63 to G66) of the large stem-loop were sensi-tive to RNase T2and resistant to RNase V1(Fig. 2A). Taken together, the structural probing results correlate well with the computer-predicted stem-loops in solution. The cleavages of the internal loop in the large stem-loop by RNase V1at U45 to U44 and by RNase T1at G65 to G64 implied that this internal loop could likely dynamically switch to two GU pairs (Fig. 2). Since the stem of the small stem-loop comprises the UUUUC repeats, it could potentially shift to base pair with a different unit of the repeat. Therefore, the cleavage pattern of the small
FIG. 2. Enzymatic probing of 5⬘-end-labeled Ba-77 RNA. (A) The RNAs were treated with RNase A (lanes A), RNase T1(lanes T1), RNase T2(lanes T2), or RNase V1(lanes V1). The concentration of the enzyme used in each reaction is indicated above each lane. Lanes C, untreated RNA sample; lanes Alk, partial digestion with alkaline buffer; lanes T1 (15 U) and lanes A (1200⫻), RNA sequencing reactions created markers for identifying the cleavage sites. The cleavage products were resolved on a 10% sequencing gel. (B) Electrophoresis of the same enzymatic cleavage products as shown in panel A on a 10% sequencing gel with a longer running time to resolve cleavage products further downstream.
on November 8, 2019 by guest
http://jvi.asm.org/
stem-loop appeared to be dynamically sensitive to both single-and double-strsingle-anded RNA-specific RNases (Fig. 2). There-fore, we specify these base pairs with broken lines (Fig. 1) to indicate that this region could be unstructured or has poten-tially unstable base pairing.
Ba-77 contains the promoter sequence for BaMV plus-strand RNA synthesis.To localize the promoter sequence for BaMV plus-strand RNA synthesis, the replicase complex iso-lated from infectedN. benthamianaplants (4) was used for the in vitro RNA synthesis assay. Three 3⬘-coterminal minus-strand transcripts of 41, 77, and 173 nt in length, designated Ba-39 (the terminal 39 nt plus two nonviral Gs at the 5⬘end), Ba-77, and Ba-173, respectively, were used as the exogenous templates for the in vitro RNA synthesis. The radioactive RNA products were resolved on an 8% polyacrylamide gel after the reaction and scanned with a phosphorimager (Fig. 3A). De-rived from densitometry and normalized with the number of uridines incorporated, the levels of products synthesized from Ba-39, Ba-173, and the 3⬘TLS of CMV RNA (4) were 72, 13, and 2% that of Ba-77, respectively (Fig. 3 and Table 1). These
results indicated that Ba-77, comprising both stem-loops, could serve as an efficient template for plus-strand RNA synthesis. Ba-39, comprising only the small stem-loop, yields two-thirds the activity of Ba-77, suggesting that the terminal 39 nt consists of most of the elements needed to promote BaMV plus-strand RNA synthesis in vitro. The large stem-loop could possibly enhance the template activity. However, Ba-173 failed to direct the plus-strand RNA synthesis efficiently, likely due to the sequence between nt 77 and 173 that folds back and either blocks the terminal sequence or abolishes the structure that initiates the plus-strand RNA synthesis.
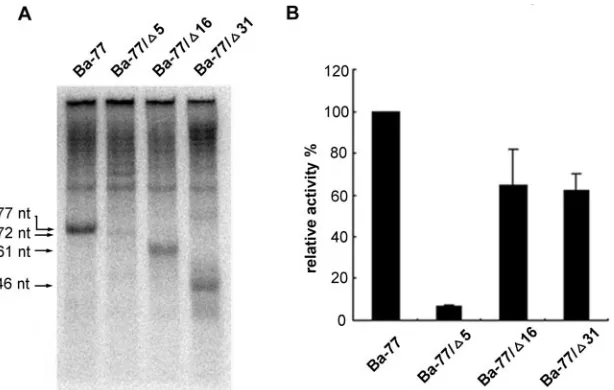
The 3ⴕ-end sequence is important for BaMV plus-strand RNA synthesis.Ba-77, containing both stem-loop sequences, is likely to be recognized by the BaMV replicase to initiate the RNA synthesis. To confirm the importance of the terminal UUUUC sequence for efficient BaMV RNA synthesis, the mutant Ba-77/⌬5, with a 5-nt deletion at the 3⬘ terminus of RNA, was constructed. The template activity of Ba-77/⌬5 was only 7% that of Ba-77, indicating that the terminal repeating sequence should play an important role in initiating the plus-strand RNA synthesis (Fig. 4). Interestingly, there are four consecutive internal UUUUC repeats from nt 36 to 17 in Ba-77 (Fig. 1). Therefore, mutants Ba-77/⌬16 and Ba-77/⌬31, with 16- and 31-nt deletions at the 3⬘terminus of the RNA, respectively, were constructed to investigate whether these re-peating units are involved in the initiation of plus-strand RNA synthesis in vitro. The template activities of Ba-77/⌬16 and Ba-77/⌬31 were 64 and 62% that of Ba-77, respectively (Fig. 4 and Table 1). These results indicated that, despite the loss of the small stem-loop sequence, the retention of the repeating sequence that mimics the structure of Ba-77 at the 3⬘end could direct BaMV RNA synthesis in vitro. Although Ba-77/⌬5 con-tains most of the template sequence, it failed to synthesize RNA efficiently without the repeating unit at the 3⬘end. Here we conclude that the repeating unit UUUUC at the 3⬘ end
[image:4.585.43.283.596.724.2]FIG. 3. Exogenous RNA template activities of BaMV RdRp. (A) Autoradiogram of the template activities, determined by using 2 pmol of each RNA transcript derived from the 3⬘ends of BaMV minus-strand RNA and CMV TLS, as indicated above each lane. The micrococcal-nuclease-treated BaMV RdRp was used. The products of the RdRp reactions are indicated by arrows. (B) Quantification of the relative RdRp template activities of the RNAs derived from the 3⬘ends of BaMV minus-strand RNA and CMV TLS. Note that the template activity of Ba-77 was designated 100%. All data represented are the averages from at least three independent experiments.
TABLE 1. Relative activities of templates used in RdRp assay
Template No. of uridines Normalized activity (%) (mean⫾SD)
Ba-173 27 13⫾8
Ba-77 7 100
Ba-39 1 72⫾13
CMV TLS 61 2⫾1
Ba-77/⌬5 7 7⫾1
Ba-77/⌬16 6 64⫾18
Ba-77/⌬31 6 62⫾8
Ba-77/UM 7 56⫾6
Ba-77/UR 9 55⫾5
Ba-77/LM 7 18⫾5
Ba-77/LR 7 21⫾1
VOL. 79, 2005 PROMOTER FOR BaMV PLUS-STRAND RNA SYNTHESIS 9049
on November 8, 2019 by guest
http://jvi.asm.org/
could serve as the initiation site of plus-strand RNA synthesis in vitro.
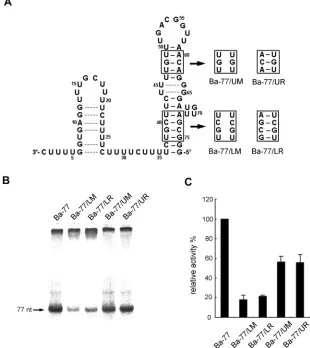
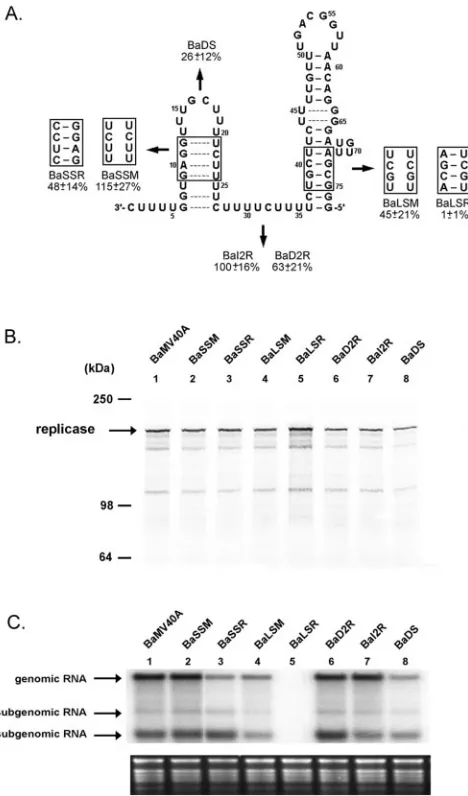
The sequence in the lower stem of the large stem-loop could modulate the template activity.The results derived from the template activities of Ba-77/⌬31 and Ba-39 indicated that the large stem-loop is critical in modulating BaMV plus-strand RNA synthesis. In order to examine whether the large stem-loop structure contributes to the promoter activity for plus-strand RNA synthesis, mutations were introduced to abolish and restore the stem structures. These mutants were desig-nated Ba-77/LM (lower stem abolished), Ba-77/LR (lower stem restored with compensatory sequence), Ba-77/UM (up-per stem abolished), and Ba-77/UR (up(up-per stem restored with compensatory sequence) (Fig. 5A). Five picomoles (about 130 ng) of each template, including Ba-77 and its derivatives, was used for the in vitro RdRp assay; results showed that the template activities of Ba-77/LM and Ba-77/LR reached about 20% that of Ba-77 (Fig. 5 and Table 1). The template activities of Ba-77/UM and Ba-77/UR achieved about 56% that of Ba-77 (Fig. 5 and Table 1). These results suggest that the sequence of the lower stem might be involved in efficient BaMV RNA synthesis. However, the structure and sequence of the upper stem appeared to be less critical than those of the lower stem. The length and the sequence of the 3ⴕend of BaMV minus-strand RNA are critical in viral RNA accumulation in vivo.To study the effects of these mutations in vivo, mutations were introduced into the full-length infectious cDNA clone, pBaMV40A. To investigate whether these mutations in the 5⬘ UTR are involved in translation, transcripts of BaMV40A and its derivatives were translated in vitro using RRL. The BaMV RdRp products of all mutants were detected at significant levels (Fig. 6B), suggesting that the mutants’ failure to accu-mulate the viral RNA products in protoplasts such as BaLSR (with the lower stem restored) and BaDS (with the small stem-loop deleted) was not due to the effects on the translation process that occurred when the mutations were introduced into the 5⬘UTR of BaMV RNA.
BaSSM, with the mutation in the stem of the small stem-loop that abolished the base pairing predicted by MFOLD, accumulated genomic RNA to a level similar to that of the wild type in protoplasts. However, BaSSR, the mutant with strengthened stem stability, produced a level of RNA only 48% that of the wild type (Fig. 6). These results suggest that no specific secondary structure is required to be involved in viral RNA accumulation in protoplasts in this region. In a test to determine whether the number of UUUUC repeats has any effect on the viral RNA accumulation, BaI2R (with two extra UUUUC repeats inserted) showed no variation in RNA accu-mulation from that of the wild type, whereas BaD2R (with two UUUUC repeats deleted) showed a reduction of RNA accu-mulation to 63% that of the wild type (Fig. 6). With BaDS, which has the entire small stem-loop deleted, the RNA accu-mulation dropped to 26% that of the wild type (Fig. 6). These results suggest that the distance between the 3⬘ end and the large stem-loop is critical to efficient viral RNA accumulation in vivo. Overall, the shortening of this region by removing the small stem-loop or two repeats (10 nt) or by strengthening the stability of the stem (an alternative way to reduce its length) reduced the levels of viral RNA accumulation in protoplasts.
BaLSM, with a disruption of the lower stem of the large stem-loop, showed an RNA accumulation that was 45% of that of the wild type. Restoration of the base pairing with a different sequence in BaLSR led to a viral RNA accumulation of 1% that of the wild type (Fig. 6). Together, these results and the data derived from the RdRp template activity assay suggest that the sequence and structure of the lower stem of the major stem-loop might be important for plus-strand RNA synthesis.
DISCUSSION
[image:5.585.141.448.68.263.2]Without the introduction of any nonviral nucleotides, the 77 3⬘-terminal nucleotides of the BaMV minus-strand RNA have been shown to be the minimal sequence needed to form two stem-loops in the T7 in vitro transcription system (24) and to constitute the most efficient RNA template in the in vitro
FIG. 4. Role of the 3⬘-end repeats in plus-strand initiation in vitro. (A) Autoradiogram of the template activities, determined by using the micrococcal-nuclease-treated BaMV RdRp. The templates, Ba-77 and its derivatives, are indicated above each lane. The products of the RdRp reactions are indicated by arrows. (B) Quantification of the relative template activities of Ba-77 and its derivatives from the in vitro RdRp assay. Note that the template activity of Ba-77 was designated 100%. All data represented are the averages from at least three independent experiments.
on November 8, 2019 by guest
http://jvi.asm.org/
BaMV RdRp assay. The enzymatic structural probing data supported the existence of the predicted structure, especially the large stem-loop in Ba-77 (Fig. 1), and the stem of the small stem-loop in Ba-77 and Ba-39 (data not shown) was not as stable as the stem in the large loop. For the small stem-loop, the ambiguity of the structural probing results in stem formation may be due to the UUUUC repeats located between nt 17 and 36 (Fig. 1), leading to alternate pairing and possibly a base-to-base stacking structure. Moreover, the free energy of the small stem is only⫺2.3 kcal/mol, as predicted by MFOLD calculation, indicating that the stem is not stable.
The transcript of Ba-39 containing the terminal UUUUC sequence was shown to be an efficient template for BaMV plus-strand RNA synthesis in vitro (Fig. 3). Mutants with a deletion of 16 or 31 nt from the 3⬘ end but retaining the internal UUUUC repeating unit as the terminal UUUUC se-quence could partially preserve the template activity for plus-strand RNA synthesis. However, the Ba-77/⌬5 mutant with the terminal UUUUC deletion failed to be an efficient template for plus-strand RNA synthesis in vitro. Taken together, these
results show that the UUUUC repeating unit at the 3⬘end is important for plus-strand RNA synthesis. The terminal UUUUC sequence could play a role similar to that of the CCA triplet found in TYMV, BMV, andCitrus tristeza virus(
Clos-teroviridae) RNA that is critical for minus-strand RNA
synthe-sis (34).
[image:6.585.135.445.74.422.2]The large stem-loop in the 3⬘-terminal region appears not to be essential for, but to enhance the efficiency of, BaMV RNA synthesis. However, mutations that change the sequence of the lower stem significantly reduced RNA synthesis in vitro and viral RNA accumulation in vivo, suggesting that the sequence of the stem is involved in regulating BaMV plus-strand RNA synthesis. In other words, the promoter for BaMV plus-strand RNA synthesis includes the large stem-loop. The results de-rived from an in vitro UV cross-linking experiment showed that the helicase-like domain of the BaMV replicase specifi-cally interacted with RNA from this region (unpublished data). Further experiments are required to characterize the interac-tions between the RdRp and the promoter for BaMV plus-strand RNA synthesis.
FIG. 5. Effects of mutations in the stem of the large stem-loop of Ba-77, determined by an in vitro RdRp assay. (A) Schematic representation of Ba-77; the sequences of the stem shown in boxes were replaced with the sequences indicated in boxes to the right of the arrows. The assigned names of the mutants are noted under the mutated sequences. (B) Autoradiogram of the template activities of Ba-77 and its derivatives, as indicated above each lane, determined by using the micrococcal-nuclease-treated BaMV RdRp. The products of the RdRp reactions are indicated by arrows. (C) Quantification of the relative RdRp template activities of Ba-77 and its derivatives. Note that the template activity of Ba-77 was designated 100%. All data represented are the averages from at least three independent experiments.
VOL. 79, 2005 PROMOTER FOR BaMV PLUS-STRAND RNA SYNTHESIS 9051
on November 8, 2019 by guest
http://jvi.asm.org/
Besides the terminal UUUUC and the large stem-loop, the distance between these two regions could also play a role in regulating the efficiency of BaMV plus-strand RNA synthesis. Mutants with shortened distances between these two regions, Ba-77/⌬16 and Ba-77/⌬31 in vitro and BaDS, BaD2R, and
BaSSR (with the stronger stem formation) in vivo, showed reduced levels of plus-strand RNA synthesis efficiency (Fig. 4) and of viral RNA accumulation (Fig. 6), likely because the interactions between the replicase and the promoter sequence require a particular amount of space to fulfill the specificity in initiating the viral plus-strand RNA synthesis. In combination with all of the different template activities, the replicase com-plex (the RdRp preparation) could have at least two different binding domains. One is the binding domain in the catalytic core that requires the template containing the UUUUC ter-minal sequence, and the other is the specificity domain that recognizes the large stem-loop. Ba-39 is a short template with a UUUUC terminal sequence that could fit into the catalytic core directly and initiate the RNA synthesis. However, an efficient template for a large molecule like Ba-77 must fit both domains. Therefore, Ba-77, which contains the UUUUC ter-minal sequence and the large stem-loop, does have the full activity. It is possible that Ba-77/LM and Ba-77/LR failed to fit into the specificity domain of the replicase complex, since they showed template activities lower than that of Ba-77. Therefore, shortening the distance between these two RNA moieties will interfere with the efficiency of RNA synthesis. However, in-creasing the distance or altering the sequence in the promoter may result in a tolerable or more flexible RNA structure to bind the replicase, bringing about less of an effect on RNA synthesis.
If the 77 3⬘-terminal nucleotides of the BaMV minus-strand RNA alone could form a promoter for efficient plus-strand RNA synthesis, this sequence would be free from the double-stranded replicative intermediate in vivo. A similar model has been described previously, indicating that release of the 3⬘ terminus of minus-strand RNA from its double-strand form (through its helicase function) is an important step in initiating BMV plus-strand RNA synthesis (30). Therefore, we hypoth-esize that the helicase may trigger the unwinding process of the double-stranded RNA and subsequently discharge the 77 nt of the 3⬘end of the minus-strand RNA from the double-stranded replicative intermediate and interact with the large stem-loop. The RdRp domain of the BaMV replicase should recognize the terminal UUUUC and initiate plus-strand RNA synthesis. Overall, these results suggested that three cis-acting ele-ments in the 3⬘ end of BaMV minus-strand RNA, i.e., the terminal UUUUC, the sequence of the large stem-loop, and the distance between these two regions, are important to ini-tiate efficient BaMV viral RNA synthesis. Although the sequences of the 5⬘UTR of BaMV RNA are not highly con-served among different strains (42), the 3⬘termini of minus-strand RNA all preserve these three cis-acting elements, as predicted by phylogenetic comparison.
ACKNOWLEDGMENTS
We thank Pei-Yu Lee of the Institute of Medical Biotechnology, Chungtai Institute of Health Sciences and Technology, for editorial help.
This work was supported by grants from National Science Council Projects NSC 91-2311-B-005-023 and 91-2311-B-005-036.
REFERENCES
1.Argos, P.1988. A sequence motif in many polymerases. Nucleic Acids Res.
16:9909–9916.
[image:7.585.47.281.69.467.2]2.Beck, D. L., P. J. Guilford, D. M. Voot, M. T. Andersen, and R. L. S. Forster. FIG. 6. Effects of mutations in the 5⬘UTR of BaMV RNA,
deter-mined by in vitro translation and in vivo protoplast inoculation assays. (A) Schematic illustration of the 3⬘end (77 nt) of BaMV minus-strand RNA and its derivatives. The names of the mutants and their RNA accumulation levels (mean⫾standard deviation) relative to that of the wild type (100%) are indicated. The data presented are the averages from at least five independent protoplast inoculation assays, and at least three Northern blots were done for each inoculation. (B) In vitro translation assay in RRL with in vitro-generated transcripts of pBaMV40A and its derivatives. Molecular size markers are indicated on the left. The arrow indicates the position of the replicase. (C) Northern blot analysis of total RNAs prepared from the inoculated
N. benthamianaprotoplasts at 48 h postinfection. RNAs were probed
with a32P-labeled RNA transcript complementary to 0.6 kb of the 3⬘ end of BaMV genomic RNA; rRNAs below the blot are the loading controls. The positions of the genomic RNA (6.4 kb) and the two subgenomic RNAs (2.0 and 1.0 kb) are indicated on the left. The levels of genomic RNA from individual inoculated samples are compared to that of BaMV40A harvested at 48 h postinfection.
on November 8, 2019 by guest
http://jvi.asm.org/
1991. Triple gene block proteins of white clover mosaic potexvirus are re-quired for transport. Virology183:695–702.
3.Cheng, C.-P., and C.-H. Tsai.1999. Structural and functional analysis of the 3⬘untranslated region of bamboo mosaic potexvirus genomic RNA. J. Mol. Biol.288:555–565.
4.Cheng, J.-H., M.-P. Ding, Y.-H. Hsu, and C.-H. Tsai. 2001. The partial purified RNA-dependent RNA polymerases from bamboo mosaic potexvirus and potato virus X infected plants containing the template-dependent activ-ities. Virus Res.80:41–52.
5.Deiman, B. A. L. M., R. M. Kortlever, and C. W. A. Pleij.1997. The role of the pseudoknot at the 3⬘end of turnip yellow mosaic virus RNA in minus-strand synthesis by the viral RNA-dependent RNA polymerase. J. Virol.
71:5990–5996.
6.Deiman, B. A. L. M., A. K. Koenen, P. W. G. Verlaan, and C. W. A. Pleij.
1998. Minimal template requirements for initiation of minus-strand synthesis in vitro by the RNA-dependent RNA polymerase of turnip yellow mosaic virus. J. Virol.72:3965–3972.
7.Dreher, T. W.1999. Functions of the 3⬘-untranslated regions of positive strand RNA viral genomes. Annu. Rev. Phytopathol.37:151–174. 8.Fabian, M. R., H. Na, D. Ray, and K. A. White.2003. 3⬘-terminal RNA
secondary structures are important for accumulation of tomato bushy stunt virus DI RNAs. Virology313:567–580.
9.Gorbalenya, A. E., and E. V. Koonin.1989. Viral proteins containing the purine NTP-binding sequence pattern. Nucleic Acids Res.17:8413–8440. 10.Hayes, R. J., and K. W. Buck.1990. Complete replication of a eukaryotic
virus RNA in vitro by a purified RNA-dependent RNA polymerase. Cell
63:363–368.
11.Hodgman, T. C.1988. A new superfamily of replicative proteins. Nature
333:22–23.
12.Houser-Scott, F., M. L. Baer, K. F. Liem, Jr., J.-M. Cai, and L. Gehrke.1994. Nucleotide sequence and structural determinants of specific binding of coat protein or coat protein peptides to the 3⬘ untranslated region of alfalfa mosaic virus RNA 4. J. Virol.68:2194–2205.
13.Houser-Scott, F., P. Ansel-McKinney, J.-M. Cai, and L. Gehrke.1997. In vitro genetic selection analysis of alfalfa mosaic virus coat protein binding to 3⬘-terminal AUGC repeats in the viral RNAs. J. Virol.71:2310–2319. 14.Huang, C.-Y., and C.-H. Tsai.1998. Evolution of bamboo mosaic virus in a
nonsystemic host results in mutations in the helicase-like domain that cause reduced RNA accumulation. Virus Res.58:127–136.
15.Huang, C.-Y., Y.-L. Huang, M. Meng, Y.-H. Hsu, and C.-H. Tsai.2001. Sequences at the 3⬘untranslated region of the bamboo mosaic potexvirus RNA interact with the viral RNA-dependent RNA polymerase. J. Virol.
75:2818–2824.
16.Huang, Y.-L., Y.-T. Han, Y.-T. Chang, Y.-H. Hsu, and M. Meng.2004. Critical residues for GTP methylation and formation of the covalent m7GMP-enzyme intermediate in the capping enzyme domain ofBamboo
mosaic virus. J. Virol.78:1271–1280.
17.Kao, C. C., and J.-H. Sun.1996. Initiation of minus-strand RNA synthesis by the brome mosaic virus RNA-dependent RNA polymerase: use of oligori-bonucleotide primers. J. Virol.70:6826–6830.
18.Kao, C. C., P. Singh, and D. J. Eckert.2001. De nova initiation of viral RNA-dependent RNA synthesis. Virology287:251–260.
19.Koonin, E. V.1991. The phylogeny of RNA-dependent RNA polymerases of plus-strand RNA viruses. J. Gen. Virol.72:2197–2206.
20.Li, Y.-I., Y.-M. Cheng, Y.-L. Huang, C.-H. Tsai, Y.-H. Hsu, and M. Meng.
1998. Identification and characterization of theEscherichia coli-expressed RNA-dependent RNA polymerase of bamboo mosaic virus. J. Virol.72:
10093–10099.
21.Li, Y.-I, Y.-J. Chen, Y.-H. Hsu, and M. Meng.2001. Characterization of the AdoMet-dependent guanylyltransferase activity that is associated with the N terminus of bamboo mosaic virus replicase. J. Virol.75:782–788. 22.Li, Y.-I., T.-W. Shih, Y.-H. Hsu, Y.-T. Han, Y.-L. Huang, and M. Meng.2001.
The helicase-like domain of plant potexvirus replicase participates in forma-tion of RNA 5⬘cap structure by exhibiting RNA 5⬘-triphosphatase activity. J. Virol.75:12114–12120.
23.Lin, N.-S., B.-Y. Lin, N.-W. Lo, C.-C. Hu, T.-Y. Chow, and Y.-H. Hsu.1994. Nucleotide sequence of the genomic RNA of bamboo mosaic potexvirus. J. Gen. Virol.75:2513–2518.
24.Lin, N.-S., F.-Z. Lin, T.-Y. Huang, and Y.-H. Hsu.1992. Genome properties of bamboo mosaic virus. Phytopathology82:731–734.
25.Oh, J.-W., T. Ito, and M. M. C. Lai.1999. A recombinant hepatitis C virus RNA-dependent RNA polymerase capable of copying the full-length viral RNA. J. Virol.73:7694–7702.
26.Osman, T. A. M., and K. W. Buck.1996. Complete replication in vitro of tobacco mosaic virus RNA by a template-dependent, membrane-bound RNA polymerase. J. Virol.70:6227–6234.
27.Panavas, T., J. Pogany, and P. D. Nagy.2002. Analysis of minimal promoter sequences for plus-stranded synthesis by the cucumber necrosis virus RNA-dependent RNA polymerase. Virology296:263–274.
28.Panavas, T., and P. D. Nagy.2003. The RNA replication enhancer element of tombusviruses contains two interchangeable hairpins that are functional during plus-strand synthesis. J. Virol.77:258–269.
29.Plante, C. A., K. H. Kim, N. Pillai-Nair, T. A. Osman, K. W. Buck, and C. L. Hemenway. 2000. Soluble, template-dependent extracts from Nicotiana benthamianaplants infected with potato virus X transcribe both plus- and minus-strand RNA templates. Virology275:444–451.
30.Pogue, G. P., and T. C. Hall.1992. The requirement for a 5⬘stem-loop structure in brome mosaic virus replication supports a new model for viral positive-strand RNA initiation. J. Virol.66:674–684.
31.Quadt, R., and E. M. J. Jaspars.1990. Purification and characterization of brome mosaic virus RNA-dependent RNA polymerase. Virology178:189– 194.
32.Reigadas, S., M. Ventura, L. Sarih-Cottin, M. Castroviejo, S. Litvak, and T. Astier-Gin.2001. HCV RNA-dependent RNA polymerase replicates in vitro the 3⬘terminal region of the minus-strand viral RNA more efficiently than the 3⬘terminal region of the plus RNA. Eur. J. Biochem.268:5857–5867. 33.Reusken, C. B. E. M., and J. F. Bol.1996. Structural elements of the
3⬘-terminal coat protein binding site in alfalfa mosaic virus RNAs. Nucleic Acids Res.24:2660–2665.
34.Satyanarayana, T., S. Gowda, M. A. Ayllon, M. R. Albiach-Marti, and W. O. Dawson.2002. Mutational analysis of the replication signals in the 3⬘ -non-translated region of citrus tristeza virus. Virology300:140–152.
35.Silberklang, M., A. M. Gillum, and U. L. RajBhandary.1977. The use of nuclease P1in sequence analysis of end group labeled RNA. Nucleic Acids
Res.4:4091–4108.
36.Singh, R. N., and T. W. Dreher.1997. Turnip yellow mosaic virus RNA-dependent RNA polymerase initiation of minus strand synthesis in vitro. Virology233:430–439.
37.Sivakumaran, K., and C. C. Kao.1999. Initiation of genomic plus-strand RNA synthesis from DNA and RNA templates by a viral RNA-dependent RNA polymerase. J. Virol.73:6415–6423.
38.Song, C., and A. E. Simon.1994. RNA-dependent RNA polymerase from plants infected with turnip crinkle virus can transcribe (⫹)- and (⫺)-strands of virus-associated RNAs. Proc. Natl. Acad. Sci. USA91:8792–8796. 39.Song, C., and A. E. Simon.1995. Requirement of a 3⬘-terminal stem-loop in
in vitro transcription by an RNA-dependent RNA polymerase. J. Mol. Biol.
245:6–14.
40.Tsai, C.-H., and T. W. Dreher.1992. Second-site suppressor mutations assist in studying the function of the 3⬘noncoding region of turnip yellow mosaic virus RNA. J. Virol.66:5190–5199.
41.Tsai, C.-H., C.-P. Cheng, C.-W. Peng, B.-Y. Lin, N.-S. Lin, and Y.-H. Hsu.
1999. Sufficient length of the poly(A) tail for the formation of a potential pseudoknot is required for efficient replication of bamboo mosaic potexvirus RNA. J. Virol.73:2703–2709.
42.Yang, C. C., J. S. Liu, C. P. Lin, and N.-S. Lin.1997. Nucleotide sequence and phylogenetic analysis of a bamboo mosaic potexvirus isolated from common bamboo (Bambusa vulgarisMcClure). Bot. Bull. Acad. Sin.38:77– 84.
43.Zuker, M., D. H. Mathews, and D. H. Turner.1999. Algorithms and ther-modynamics for RNA secondary structure prediction: a practical guide, p. 11–43.InJ. Barciszewski and B. F. C. Clark (ed.), RNA biochemistry and biotechnology. Proceedings of the NATO Advanced Research Workshop, Poznan, Poland, 10 to 17 October 1998. Kluwer Academic Publishers, Dor-drecht, The Netherlands.
VOL. 79, 2005 PROMOTER FOR BaMV PLUS-STRAND RNA SYNTHESIS 9053