JOURNALOFVIROLOGY, Nov. 1971, p. 716-721 Vol. 8,No. 5 Copyright©1971 AmericanSociety for Microbiology PrintedinU.S.A.
Quantitative
Binding of
125I-Concanavalin
A to
Normal and
Transformed Cells
DONNA J. ARNDT-JOVIN' AND PAUL BERG
Department of Biochemistry,Stanford University School of Medicine, Stanford, California 94305
Received for publication 2 August 1971
We have measured the quantitative binding of the radioactively labeled ag-glutinin1251-concanavalinA to normal mammalian cells and simian virus 40- and polyoma virus-transformed cells from tissue culture. Parallel measurements ofthe amountof 125I-concanavalin Anecessary to causeagglutinationof thecellsin sus-pensionwerecarriedout. Thetransformedand nontransformedcellsused for these
experiments showlarge differences in their ability to be agglutinated by
12qI-con-canavalin A. However, these cell lines have the same number ofspecific binding sites and similar affinities for the agglutininwhether transformed, trypsinized, or nontransformed. We conclude that the differential capacityofconcanavalin A to agglutinate transformed cells relativetonormalcells does not result fromdifferences in the number ofbinding sites betweenthetwo types of cells.
A number of proteins, termed
phytohemag-glutinins becausetheyagglutinateredbloodcells, have beenisolatedfromplants (2).Low concen-trations of several of these proteins agglutinate
cells transformed chemically or by
deoxyribo-nucleic acid (DNA) tumor viruses, whereas ag-glutination of nontransformed cellsrequires rela-tively high concentrations of the proteins (1, 4, 10,20). Theconcentration ofagglutinin required
to agglutinate cells is
inversely
related to thesaturationdensities reached in culture (18).
The phytohemagglutinins precipitate different
macromolecular carbohydrates andprobably in-teractwiththecellsurfacedat "exposed"
carbo-hydrate moieties; as
expected,
the agglutininactivity can be inhibited or even reversed by carbohydrate haptens specific for each phyto-hemagglutinin (4, 10). Nontransformed cells be-come agglutinable by the activity ofproteases; this observation suggested that nontransformed cellscontain"cryptic"
binding
sites for thephyto-hemagglutinins and that these sitescanbeexposed byproteasesor byarearrangementin the archi-tecture of the cell surface
by
transformation (3,11).
This hypothesis was based upon quantitative differences in the agglutination of different cell lines (3) and from areport that the amount of 63Ni-labeled concanavalin A (con A) bound to
IFellow of the Jane Coffin ChildsMemorialFund forMedical Research. Present address: Abteilung fur Molekulare Biologie, Max-Planck-Institut fUrBiophysikalischeChemie,D-3400 Goet-tingen, West Germany.
transformed cells was considerably greater than that bound to nontransformed cells (11). How-ever, the quantitative relationship between the amount of phytohemagglutin from red kidney beans neededto cause cellagglutinationand the amount ofthe agglutinin bound tothe cell sur-face has been obscure (20). Moreover, in the study with the 63Ni-labeledconA (11), the radio-active marker was not covalently linked to the protein and thus it is difficultto be certainthat thedeterminations of 88Ni bindingtothe cells ac-tually measured complex formation betweenthe con A and the cell surface; the fact that the amount of 65Ni binding was inhibited less than
50%
by
an excess ofthehapten
isparticularly
unsettling.
Wefelt that moredirect measurements of ag-glutinin bindingtocells and the correlation of that
bindingtothe phenomenon ofagglutinationwere needed to substantiate the "cryptic-site" hy-pothesis(11). SinceconA canbeeasily purifiedto physical homogeneity and many ofits physical and carbohydrate bindingproperties arealready
known (12, 14, 15, 19, 21), it was selected for iodination with 125I (17) to produce a stable covalent linkage between the labeled group and the agglutinin. Although the amount of 1251-labeled con A needed to agglutinate nontrans-formed cellswas muchgreater than that needed for nontransformedcells,wecouldnotdetectany significant difference in the binding of labeled proteintothetwotypesofcells.
716
on November 11, 2019 by guest
http://jvi.asm.org/
MATERIALS AND METHODS Tissue culture. Mouse fibroblast cell lines 3T3M-T20 (clonedfrom aline fromR. Dulbecco) and SV-3T3-26-d2 (alsofrom R.Dulbecco)andbaby hamster kidney cell lines BHK-21 (original clone from labora-toryof M. Stoker) and MT-1 (apolyoma-transformed clonederivedfrom thisline) were cultured in plastic petri dishes (Nunc, 100 by 20 mm, orFalcon, 35by 10 mm) in Dulbecco's modified Eagle's medium (GrandIslandBiologicalCo.)supplementedwith10% calfserum and 25,g ofchlortetracycloneperml.
Con A.Con A was extracted fromjack beanmeal (Sigma ChemicalCo.) andcrystallized bythe proce-dureof Sumner andHowell (21). Somepreparations were treatedwith 1Macetic acidfor 20 min and dia-lyzed to render the protein water-soluble and free from any carbohydrates (14). Allpreparations were adsorbedto aSephadexG150columnandelutedwith buffers atlowpH asdescribed by Olsonand Liener (14).
1251-labeled Con A. Specific iodination oftyrosine residues with lactoperoxidase (the kind gift of M. Morrison)wascarriedout asdescribedbyPhillipsand Morrison (17) with modifications ofconcentrations asfollows.The con A at a tetramerconcentrationof 10-4 Mto 1.6 X 10-4M wasdissolved in0.1 Msodium phosphatebuffer (pH7.4) containing Na125I at4 X 10-4to4 X 10- M(specific activity,200 to 500IuCi/
,umole).The reactionwasinitiatedbytheaddition of lactoperoxidase to aconcentration of 7.7 X 10-7 M and hydrogenperoxideat 8 X 10-5 to 1.6 X 10-4M. The incorporation of radioactivity was followed by precipitation of the protein with 5% trichloroacetic acidforcounting.
1251 counting. A Nuclear-Chicago model 1085 gamma counter was used for determining the 1251 radioactivity.
Agglutination assays.Cellagglutinationwasassayed byamodification of theproceduredescribedby Burger and Noonan (5). Subconfluent cells were removed from theplateswith 4 X 1o-4M ethylenediaminetetra-acetic acid (EDTA) in buffered saline at 37 C and washed oncewith buffered salinecontainingMg2+and Ca2+beforebeingwashed and suspendedin buffered salinelacking Mg2+and Ca2+. A100-jslitersolution of 2 X 105 to 106 cells/ml was mixed with an equal sample ofvarious concentrations of agglutinin dis-solvedinbuffered salinein the wells ofaporcelainspot plate.Theplatewaskeptingentlemotionat22C,and samplesof 20to50,literswereremovedat5,10,and and 30 min and observed in thewellsofplastic spot plates (Linbro Chemicals FB48) with an inverted microscope.
Bindingassays. Method 1was asfollows. Subcon-fluentcellswereremoved fromplateswith4 X 14M
EDTA in buffered saline lacking Mg2+ and Ca2+, washedoncewith buffered salinecontainingMg2+and Ca2+ and then washed and suspended in buffered salinewithoutMg2+and Ca2+at adensityof 5 X 105 to 106cells/ml. Different concentrationsof125I-labeled con Awithorwithout 0.3 M a-methyl-D-glucopyrano-side,ahaptenforconA,wereadded,and the cellswere
incubatedwith slowagitationfor 15 minat0C.After
centrifugation at 700 X g for 1min,the cell pellet was washed twotimes with coldbufferedsalineand then solubilizedwith 0.5 N NaOH; a sample was counted directlyinagamma counterand assayed for total pro-tein by the method ofLowry et al. (13). W
Method 2 was as follows. The cells remained at-tached to the culture plates. Subconfluent cells on 35-mmplastic petri plates were washed two times with cold buffered saline lacking Mg2+ andCa2+and then wereincubated with 0.3 ml of a solution containing different concentrations of'25I-con A with and without hapten. After 15minat 0C, the liquid was aspirated and theplates were washed twice with cold buffered saline.After the cells were solubilized in 0.5 N NaOH, they were analyzed as mentioned above. Control plates incubatedwithoutagglutininweretrypsinized, and the cellswerecountedin a hemocytometer.
RESULTS
Radioactivelabelingofcon A.Afterreactionof conAwith1251 in thepresence oflactoperoxidase and peroxide, the proteinwas dialyzed and then adsorbedto aSephadex G150 column. Figure 1 shows the elution profile of the radioactivity. Acrylamide gels of the material specifically bound toandeluted fromSephadexgaveonlyonepeak in the normalposition for purified conA. From the specific activity of the iodine, it was
deter-mined that one residue of tyrosine per 27,000 molecular weighthadbeeniodinated. This molec-ularweight isassumed to be theintactmonomer (22); however, atthepH of thebinding studies,
ml
FIG. 1. Dextranadsorptionandelutionof5 mg of 125I-con A (acetic acid-treated). The proteini was
adsorbedtoacolumn(I by27cm) ofG-150Sephadex
in 0.01 m tris(hydroxymethyl)aminomethane (Tris)-hydrochloride buffer (pH 7.2) containing 103 M
CaCl2and10-3 M MgCl2. Theeffluent wasmonitored for 125I radioactivity in a gamma counter. Adsorbed materialwaseluted with 0.02 Mglycine-HCI (pH 2.0) and dropped into tubes containing 0.075 M Tris-hydrochloride (pH9.0) to bring the solution to
neu-trality.
on November 11, 2019 by guest
http://jvi.asm.org/
[image:2.485.244.434.399.543.2]ARNDT-JOVIN AND BERG
the con A probably exists as a tetramer (15, 21, 22).
The'251-conAretained itshaptenpropertiesas demonstrated by its ability to be adsorbed onto Sephadex and eluted by either 0.1 M glucose or low pH and by the fact that the bindingtocells can be competed for or reversed by a-methyl-D-glucopyranoside butnotby N-acetylglucosamine. Competition experiments were done by binding '251-con A to cells in the presence of cold, un-modified con A. Allof the '25I-con A was com-peted by unmodifiedconA, and thecompetition experiments gave the same binding constants for 1251-con A asunmodified conA.
Agglutination of transformed and nontrans-formed cells. Table 1 givesthe levels of'251-con A and unlabeled conArequired to obtain aggluti-nationofmousefibroblasts [3T3 M and SV-3T3 (simian virus40-transformed) or 3T6 (non-viral-transformed)] and baby hamster kidney cells [BHK-21 (nontransformed) or MA-8 (abortively transformed) and MT-1 (polyoma virus-trans-formed)] and the levels required after mild trypsinization.
TABLE 1. Levels of '251-coni A or tlnlabeled conl A
requiredforagglutinationi of mouisefibroblasts andbaby hamsterkidney cells
Cellline
Concnof
'251-conA
orunlabeled
conA (,ug/ml) for half-maxi-mal
aggluti-nation
Mouse Normal
3T3M...
3T3M after0.01%trypsin for
10min...
Transformed
SV3T3...
SV3T3 after 0.01% trypsin for
10 min...
3T6...
3T6 after0.01% trypsin for
10 min...
Hamster Normal
BHK-21...
BHK-21 after0.01% trypsin for
10min...
MA-8...
MA-8 after0.01% trypsin for
10min... Polyomavirus-transformed
MT-1...
MT-1 after0.01% trypsinfor
10min...
70-120
7 7-15
7 7-10
7-10
25-30 7
100-120
10 7 7-10
16
12
cpm xio-3 8
4
16
12
cpm t10-3 8
4 A
0 40 80, 2,~ 120. 160 200
ggIml"''I-con A
B
//
II
0 40 80 120 160 200
[image:3.485.255.444.65.367.2]ggl1m251-conA
FIG. 2. Binidinigof"5I-coniA to cells in
solutiont
by method 1. (A) 3T3M cells (105) at 0 C; (B) SV3T3 cells (105) at 0 C. Symbols: X, untreated cellsplus 'I5I-con A, counts perminuiite bouind; Oi, cells treated with 0.01%'o trypsillfor 10millplus 125I-conA, counts perminute bounid; A, cellsplus 0.3 -4f a-methylgluco-pyraniosideplus '251-con A, colltits perminiute
bounlid.
The transformedand nontransformed cells used for these experiments exhibited significant dif-ferencesintheir ability to be agglutinated by con A. Thesamedifferenceswereobserved for wheat germ agglutinin activity with these cells (unpub-lisheddata), andtheyconfirmthedifferencesseen inagglutinabilityby others (1, 3, 4, 10, 20). How-ever, these cell lines have the same number of con A binding sites available whether trans-formed, trypsinized, or nontransformed as mea-suredbythespecificbindingof1251-labeledconA, calculatedonthebasis ofequal concentrations of cellprotein, orhave a slightlygreater number of binding sites for nontransformed cells compared totransformed or trypsinized cells calculated on the basis ofequalcellnumbers.
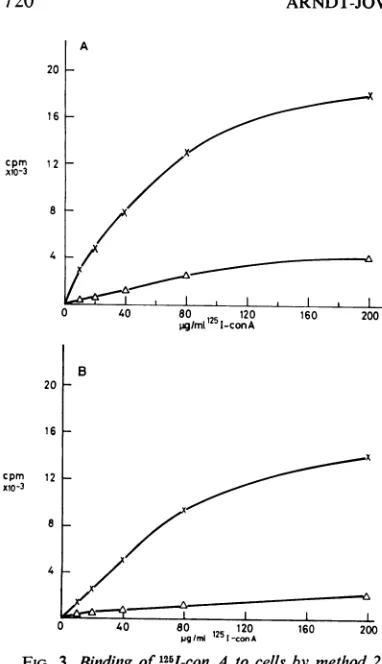
Binding of 1251-concanavalin A to transformed and nontransformed cells. Figure 2 shows typical
bindingcurvesfor1251-conA to 3T3and SV-3T3 cells in solution. The half-maximal binding of
718 J. VIROL.
on November 11, 2019 by guest
http://jvi.asm.org/
[image:3.485.50.245.337.646.2]TABLE 2. Half-maximal binding ofi5I-con A to 3T3 versus SV-3T3 anidBHK versuspolyoma-BHK cells
Concnof 125I-con A Cell line Cell number Method ofbindinebnig (ug/ml)for
half-maximalbinding
Mouse Normal
3T3M...X...SX 105 1 22 40-50
3T3M... 5 X1011 0 60-80
3T3M... 105 1 0 60-&0
3T3M, 0.01%trypsin for 10 min 105 1 0 60
3T3M... 3 X 105 2 0 75
3T3M... 105 2 0 50-60
Simian virus 40-transformed
SV3T3.... 5 X 105 1 22 40
SV3T3... 5X105 1 0 60-80
SV3T3... ... 105 1 0 60-80
SV3T3, 0.01% trypsin for 10 min.... 10, 1 0 60-80
SV3T3... 3X105 2 0 60-80
SV3T3 ..105 2 0 50
Hamster Normal
BHK,MA-8... 2 X 105 1 22 30
BHK,MA-8 ... ... 105 1 0 35-40
Polyomavirus-transformed
Polyoma-BHK,MT-1.... 2 X 105 1 22 40
Polyoma-BHK, MT-1...1 105 1 0 35-40
125I-con A (at different cell concentrations, at temperaturesof22or0C, withor without mild trypsindigestion)to3T3versusSV-3T3and BHK
versuspolyoma-BHKcells isgiveninTable2. Since proteolytic digestion ofcells resulted in
greater agglutinability, it was conceivable that
small amounts of proteolytic enzyme released fromthecellswhentheywereremovedfromthe plates couldcause erroneousinterpretation of the
specific binding results. To overcome this prob-lem, the binding assay was done with cells still
attached totheplatesaswellaswiththecellsin solution.Typical bindingcurvesfor 251I-conAto
3T3M and SV-3T3 cells at 105 cells per 35-mm
plate are given in Fig. 3 and reveal the same
numberofsitespercell for normalortransformed cells, 10' to3 x 101. Thehalfmaximalbinding
of125I-conA tothesecelllinesand toBHK
nor-mal and polyoma-transformed cell lines is
con-stantfor 3 X 104to105cellsper35-mmplate and
occursat a conA concentration of -70
Ag/ml;
the binding is inhibited by 0.03 M
a-methyl-D-glucopyranoside and is completely competed for by unmodifiedconA.Nodifferenceintheamount
ofbinding to normal and transformed cells was
observed for con A labeled with
'4C-iodoaceta-mide.
DISCUSSION
Ourdata indicate that there isanequal number
ofconAbinding sitesonthe cell surfaceof
trans-formed andnontransformed cells.Therefore, the differential capacity of con A to agglutinate transformed cells relativetonontransformed cells doesnotresult fromdifferences in the number of binding sites between thetwo typesofcells.
If there isindeed no difference in the number ofconA binding sites or indeed of other agglu-tinin-binding sites (20) between transformed and nontransformed cells, then how can one explain the differential agglutinability of the cell lines? Three models can be generated to explain the phenomenon: (i) steric interference, (ii) allosteric conversion, and (iii) ionic interaction. Steric inter-ference wouldrequire that theagglutinin-binding
site be buriedonthe surface ofthe normal, non-transformed cell, such that the second hapten binding site of the agglutinin is unavailable for bindingto asiteonanadjacent cell. Conversely, the transformed cell binding site would be more external, allowing the second hapten binding of theagglutininsitetoattachtoanother cell.
Allosteric conversion would require that the binding ofagglutinintothetransformedcellcause a rearrangement in the structure of the surface, such that the lectin is ableto interact with
adja-centcells,orthatthebinding ofagglutininto the nontransformedcellcause arearrangementinthe
cellsurface such that the second hapten siteonthe agglutinin cannot participate in second binding interactions with anothercell, orboth.
on November 11, 2019 by guest
http://jvi.asm.org/
ARNDT-JOVIN AND BERG
2C
16
cpm 12 x10-3
8
4
A
I
/
0 40 80 120
g/ml"'1-conA
20F
16
12
4
0
B
grees oftrypsinization causethe cells to become monodisperse.
Although we do not know enough about the x actual differences in cell surfaces between trans-formed and normal cells to distinguish clearly a mechanism foragglutination,wehave shown that
transformation does not create or uncover new binding sites for the agglutinin con A, nor has such a difference been observed for the
phyto-hemagglutininfrom redkidney beans;perhapsno difference exists for any ofthe agglutinins. The models which can explain suchobservations re-quire more sophisticated understanding of the 1: 2{i0 interaction of cell surfaces and their structural
makeup.
Similar results for thespecific binding of lectins totransformed and normal cells havebeenfound concurrentlyinother laboratories (6,16).
0
40 80 120
.igtmi 1251-conA
FIG. 3. Binding of'251-conA tocellsi (A) 3T3Mcells (101) at 0C; (B) SV31
at 0C. Symbols: X, cellsplus '25I-con,
minute bound; A, cells plus 0.03 M
az-pyranoside plus '25I-con A, counts per
The argument of ionic interactic that theability of cells to growon t
cellsortobe contact-inhibitedbythe adjacentcells andtheabilityofcellst
oneanother inthepresenceofcopoly
specific proteins may both depend (
figuration and localized charge whici
on adjacent cells. It is possible that membrane potential common to cel ionically masked on the surface of
formedcell. This modelrequires noc
number ofbinding sites foragglutinil formedand normalcellsbutsimplya(
thecharge repulsionofthecells.The
cellsurfacesby proteolyticenzymesd( the amount ofspecific binding of cc
described above but could drastical ionicmakeupofthe cells. Indeed, so] trypsinization alone can cause cells
sticky and adhere to one another i
presence of external proteins, whereE
ACKNOWLEDGMENTS
This investigation was supported by Public Health Service grantGM 13235 fromtheNationalInstitute of General Medical Sciencesand byAmerican Cancer SocietygrantVC-23A. It was alsoaided bya grantfrom theJane CoffinMemorial Fund for Medical Research.
LITERATURE CITED
1. Aub, J. C., B. H. Sanford, and M. N. Cote. 1965. Studieson
reactivity of tumor and normal cells to a wheat germ
agglutinin. Proc. Nat. Acad. Sci. U.S.A. 54:396-399. i 2. Boyd, W. C. 1963. The lectins: their present status. Vox.
160 200 Sang.8:1-32.
3.Burger, M. M. 1969. A difference in the architecture of the bymethod 2. surface membrane of normal andvirally transformed cells.
F3cells (105) Proc. Nat. Acad. Sci. U.S.A 62:994-1001.
4, counts per 4. Burger, M. M., and A. R. Goldberg. 1967. Identification of a
-methylgluco- tumor-specific determinant on neoplastic cell surfaces.
iinute bound. Proc. Nat. Acad. Sci. U.S.A. 57:359-366.
5. Burger, M. M., and K. D. Noonan. 1970. Restoration of normalgrowth by covering of agglutinin sites ontumor
cell surface. Nature (London) 228:512-515. an suggests 6. Cline, M. J., and D. C.Livingston. 1971. Binding of
3H-;op of other concanavalin Aby normal and transformedcells. Nature
presenceof (London) 232:155-156.
toadhere to 7. Duksin, D., E. Katchalski, and L. Sachs. 1970. Specific
aggregationofSV40-transformedcells byornithine,leucine
rmers (7)or copolymers. Proc. Nat. Acad. Sci. U.S.A. 67:185-192. on the con- 8. Forrester,J.A.,E. J. Ambrose, and I. Macpherson. 1962.
cells"see" Electrophoretic investigations of a clone of hamster fibro-the strong blasts and polyoma-transformed cells from the same
e s ong population. Nature (London) 196:1068-1070.
Us (8, 9) is 9. Hause,L.L., R. A. Pattillo, A.Sances, Jr., and R. F.
Mat-r the trans- tingly.1970.Cellsurfacecoatingsand membranepotentials difference in of malignant and nonmalignant cells. Science 169:601-603.
O. Inbar, M., andL.Sachs. 1969.Interactionof the
carbohydrate-ns ontrans- binding protein concanavalin A withnormal and
trans-differenceim formedcells. Proc.Nat. Acad.Sci.U.S.A. 63:1418-1425.
digestion of 11. Inbar,M.,andL.Sachs. 1969. Structural difference in sites oesnot alter on the surface membrane of normal and transformedcells.
Nature (London) 223:710-712.
on A as we 12. Kalb, A.J., andA. Levitzki. 1968. Metal-bindingsites of
ly alter the concanavalin A and their roleinthebinding of
a-methyl-melevels of D-glucopyranoside. Biochem. J. 109:669-672.
to become 13. Lowry, 0. H., N. J. Rosebrough, A. L. Farr, and R. J. without the Randall. 1951. Proteinmeasurement withthe Folinphenol
without the reagent. J.Biol.Chem. 193:265-275.
as other de- 14. Olson, M.0. J.,andI. E.Liener. 1967. Some physical and
720 J. VIROL.
cpm xlo-3
I
8
I on November 11, 2019 by guest
http://jvi.asm.org/
[image:5.485.53.244.51.383.2]chemical properties of concanavalin A,the phytohemagglu-tinin ofthejack bean. Biochemistry6:105-111. 15. Olson, M.0.J.,andI.E.Liener.1967. The associationand
dissociation of concanavalin A, the phytohemagglutinin of thejack bean. Biochemistry6:3801-3808.
16. Ozanne,B., and J.Sambrook.1971. Binding ofradioactively labelled concanavalin A and wheat germ agglutinin to
normalandvirus-transformed cells. Nature (London) 232: 156-160.
17. Phillips,D.R.,and M. Morrison. 1970.The arrangement of proteinsin the human erythrocytemembrane. Biochem. Biophys. Res. Commun. 40:284-289.
18. Pollack, R. E., and M. M. Burger. 1969. Surface-specific characteristics ofa contact-inhibited cell line containing
theSV40 viralgenome.Proc. Nat. Acad. Sci. U.S.A. 62: 1074-1076.
19. So, L. L., andI. J.Goldstein. 1968. Protein carbohydrate
interaction. XX. On the number of combining sites on
concanavalin A thephytohemagglutinin of the jack bean. Biochim. Biophys.Acta 165:398-404.
20. Steck,T. L.,and D. F. H. Wallach. 1965.Thebinding of kidney-beanphytohemagglutinin by Ehrlich ascites
carci-noma. Biochim. Biophys. Acta 97:510-522.
21.Sumner,J.B.,and S. F.Howell. 1936. Theidentification of thehemagglutininofthejack bean withconcanavalin A. J. Bacteriol. 32:227-237.
22.Wang,J.L.,B. A.Cunningham,and D. M.Edelman. 1971. Unusualfragments in the subunitstructureof concanavalin A.Proc.Nat. Acad.Sci.U.S.A.68:1130-1134.
on November 11, 2019 by guest
http://jvi.asm.org/