Inhibition
of
Herpes Simplex
Virus
Type
2
Replication by
Thymidine
GARY H. COHEN, MINDIE N. FACTOR, AND MANUEL PONCE DELEON
DepartmentofMicrobiology, School of DentalMedicine, Universitvof Pennsylvaniaand Center for Oral HealthResearch, Philadelphia, Pennsylvania 19174
Received for publication 13March 1974
Replication of herpes si7aplex virus type2 (HSV-2) wasimpeded inKB cells
which were blocked in their capacity to synthesize DNA by 2mMthymidine
(TdR). The degree of inhibition was dependentupon the concentration of TdR.
In marked contrast, HSV-1 is able to replicate under these conditions. The
failure of HSV-2 to replicate is probably due to the inhibition ofviral DNA
synthesis; therewas a marked reduction in therateof DNAsynthesis aswellas
the total amount of HSV-2 DNA made in the presence of 2 mM TdR. We
postulated that the effect of TdR on viral replication occurs at the level of
ribonucleotide reductase in a manner similar to KBcells. However, unlike KB
cells,an alteredribonucleotide reductaseactivity, highly resistanttothymidine
triphosphateinhibition, wasfound in extracts ofHSV-2-infected KB cells. This
activitywaspresentinHSV-2-infectedcellsincubated in thepresence orabsence
ofTdR.Ribonucleotide reductase activity inextractsofHSV-1-infected KBcells
showed asimilarresistanceto thymidine triphosphate inhibition. These results
suggest that the effect ofTdRon HSV-2 replication occurs at a stage of DNA
synthesis other than reduction of cytidine nucleotides to deoxycytidine
nucleo-tides.
We reported recently that herpes simplex virus type 1 (HSV-1) replication is unimpeded in KBcells incubated in the presence of 2 mM
thymidine (TdR)
(3). This concentration of TdRblocks host cell DNAsynthesis
and subse-quent cell division (1, 2, 3, 15, 17). These observationssuggested
that at least one aspect of DNAreplication
utilizedby
HSV differssignificantly
from that of the host cell. This difference wasexplained by
thefinding
of a"new" viral-induced ribonucleotide reductase
enzyme activity in the cell after infection and
by
the observation that thisenzymewas mark-edly resistant to inhibitionby
thymidine
tri-phosphate (TTP),anegativeeffector ofribonu-cleotide reductase (1, 3, 9, 15). The "new"
enzymerelievedthe TdR blocktoDNA
synthe-sisby insuringa
pool
ofdeoxycytidylate
precur-sors forreplication of virus.The oral form of HSV has been shown to
differ
significantly
from thegenital
form(HSV-2)in anumberof
biological, biochemical,
and
biophysical
characteristics(12). Therefore,
we
investigated
thecapacity
ofHSV-2 torepli-cateinKBcellsinthepresence ofexcessTdR.
Wepresent evidence heretoshowthatHSV-2
is unable to replicate in the presence of2 mM TdR. The failureto
replicate
isprobably
dueto20
theinhibition ofviralDNAsynthesis. However, aribonucleotidereductase with properties
simi-lartotheenzymefound in HSV-1-infected cells
was also present in HSV-2-infected cells. This
suggeststhattheTdR block occurs at a stage of
DNA replication other than reduction of cyti-dine nucleotidesto deoxycytidine nucleotides.
MATERIALS ANDMETHODS
Cell cultures. Conditions for growth and mainte-nance of KB cells andbaby hamster kidney (BHK)
cells have beenpreviouslydescribed (3, 4).
Virus strains and titrations. Three strains of
HSV-1, HF, KOS, and CL, and three strains of
HSV-2, Savage(SAV) HSV (381) andVF, wereused. TheKOS, HSV (381), andVF strainswerekindly pro-vided by L. H. Moss III, Ohio State College of Medicine.StrainsVFand CLwererecentisolatesand were passed threetimes inBHKmonolayers priorto
use. Preparationofhigh-titeredvirusstockaswellas
assay by plaque titrations were
performed
as de-scribed previously (3).Cellsynchronization. KB cellswere
synchronized
by the double TdR block method(3, 4).Briefly,
KB cellsweretreated with2mM TdRfor 18to20h.The cells were washedfree ofTdRby centrifugation andresuspended in fresh warm medium to reverse the
block, and then allowedtogrowforanadditional9to 12h. Atthistime, 2mM TdRwasaddedforanother
on November 10, 2019 by guest
http://jvi.asm.org/
INHIBITION OF HSV-2 REPLICATION
period of 18to20h.Infectionswerestarted 1 hpriorto reversal of thesecond-TdRblock.
DNA synthesis in synchronized cell cultures. Pulse-labeling experimentswerecarriedoutas
previ-ously described (3, 4) employing 0.2 ACi of
['H]hypoxanthine per ml (specific activity 5 to 10
Ci/mmol, NewEngland Nuclear Corp.).Methods for the separation of DNA and RNA as well as the
determination of radioactivity were previously de-scribed(3,4).Radioactive-labeledDNAwasobtained as previously described except that ['H ]hypoxan-thinewas employedasthe isotope (3, 4).Separation
of viral DNA fromcellular DNA was carriedout by CsCl density gradient centrifugation (3, 4).
Protein and RNA synthesis in synchronized cell cultures. Infectedoruninfected KB cells(generally2
x 10'cells/ml)wereincubated at the times indicated with0.2ACiof [3H]L-aminoacid mixtureperml(New
England Nuclear Corp.)tomeasureproteinsynthesis
or [3H]hypoxanthine (0.2 MCi/ml) to estimate RNA synthesis. Incorporation of label was stopped by
pouring the cellsontoicedphosphatebufferedsaline (PBS). The cellswerecollectedby centrifugation and
resuspended in 5 ml ofcold 5% trichloroacetic acid. Theresulting precipitatewaswashed three times with trichloroacetic acid and dissolved in 0.1 N NaOH. ThesampleswereaddedtoAquasoland the radioac-tivitywasmeasured.
Preparation ofextracts forenzyme assay. The
preparation ofcell-freeextracts has been previously reported (3).Theextractswerestoredat -100 C ina
Union Carbide liquid nitrogen refrigerator. Under thesestorageconditionsribonucleotide reductase
ac-tivity remains stableforseveralyears.
Ribonucleotide reductase assay. The conversion
ofcytidine diphosphate (CDP) to deoxycytidine di-phosphate (dCDP)wasmeasured byamodificationof
the method describedby Cohen (3) toinclude
proce-dures in the assay described by Setlow (14). The
standard reaction mixture (0.25 ml) contained: 10 mMTris, pH7.8, 2.5mMATP,1.4mMdithiothreitol (DTT); 2.8 mM MgCl,; 0.02 mM CDP, 1.5
ACi
of ['H]CDP (specific activity 13 Ci/mmol; Schwartz-Mann) and enzymeextract. Thesamples wereincu-bated at 30C for 15 min, and the reaction was
stopped by addition of 0.25 ml of 2 N HCl. The precipitated material wasseparated from the
super-natantfluid bycentrifugation. Thesupernatantfluid
was heatedat 100C for 15 min, then evaporatedto dryness in a desiccator containing solid KOH and
P20,. Sampleswereredissolvedin 50glitersofwater and a 25-gliter portionwas analyzed bypaper
chro-matography (3). The rate of formation ofdCMP in cell extracts prepared from uninfected KB cells and KB cells infected with HSV-1 or HSV-2 was linear
for at least 25 min and was proportional to extract addedinthe rangeof 0.2 to0.8 mgofprotein. Each
assay wasrun induplicate and theresults presented
are the average ofduplicate assays whichvaried by
10%. One unit ofribonucleotide reductase activity is definedasthatamountofenzymewhich catalyzes
for-mation of dCMP at a rate of1 nmol per mg per h.
Protein concentration wasdetermined by the method
of Lowry et al. (8).
RESULTS
Replication of infectiousvirus in the pres-ence and abspres-ence of2 mMthymidine.Table 1 shows a typical experiment in which the num-ber of PFUformed by three strains ofHSV-1 in theTdR-blocked KB cellswas
nearly
ashighas the number formed in the control cells (no TdR). In contrast, TdR hadasignificant
inhibi-tory affect on replication of the three genital strains. Thetiters were200- to400-fold lower in KB cells containing TdR than in controls(no
TdR) and were similar to
background
levels found in 3 h-infected cultures. Furthermore, this decrease in titer in the presence of TdRwas not due to release ofHSV-2 into the supema-tant fluid. The total amount of extracellular virus was at least onelog
lower than that of cell-associated virus. These results indicate thatHSV-2
replicationinsynchronized
KBcellswas inhibitedby
2mMTdR. In thefollowing
experi-ments, one strain of HSV-2
(Savage)
and one strain ofHSV-1
(HF)
wasemployed.
Dose response relationship of thymidine concentration to virus synthesis. Figure 1 shows the dose response
relationship
between the amount ofHSV-2synthesized by
KB cells and the concentration of TdR present in the medium. In the range 2iM
to 2mM,
every 10-fold increase in TdR concentrationproduced
approximately
a onelog
decreaseinvirus titer.Complete
inhibitionoccurredat 2mMTdRand
no inhibition was observed at 2
jiM.
These results indicate thatHSV-2
replication
was inhibitedat aconcentration ofTdR which also inhibited cellular DNA synthesis and division in uninfected KB cells (3, 4) and that thisTABLE 1. Effect of excess thymidine on the replication of HSV types 1 and 2
PFUa HSV virus strain employed No Thymidine
thymidine (20h) (20h)
HSV-1
HF... 2.8x
107
1.6x107KOS ... 1.6x 108 6.0x 107 CL (wild) ... 6.8 x 107 2.0 x 107 HSV-2
SAV . ... 2.6 x 108 1.0 x 104° HSV (381) ... 3.6 x 107 1.0 x 105° VF (wild) ... 4.0 x 10, 1.8 x 104°
aEach value represents the average of duplicate
samples. See legend in Fig. 1 for procedure.
°Values similar to background of PFU found at 3 h postinfection.
21
VOL. 14,1974
on November 10, 2019 by guest
http://jvi.asm.org/
[image:2.499.251.447.464.627.2]106
PFU
SAMPLE
0
0
0
0~~~~~~~/
~
I}
110 2XIO3 2XIO-2
2XIO-l
2THYMIDINE CONCENTRATION (mM)
FIG. 1. Dose response relationship of TdR concen-trations to virus synthesis. Cultures were svnchro-nized by the double TdR block method. The cells werewashed to remove TdR and infected at an input multiplicitv of 10PFUIcellin thepresence ofvary'ing
concentrationsof TdR as indicated in the figure. The viruswasallowedtoabsorb for1h. Unabsorbed virus waseliminated by three washes with minimal essen-tial medium containing the corresponding TdR con-centration. Twenty-four hours later the cells were harvested and washed with PBS, and cell associated virus wasdetermined. Symbols: 0, PFU per sample with varying concentrations of TdR; 0, background virus in KB cells containing TdR harvested at 3 h
postinfection.
inhibition of new virus synthesis was dose dependent.
Rate of DNA
synthesis
in HSV-2-infected and uninfected KB cellsin thepresenceand absence ofexcessTdR.Synchronized
KBcellswere inf'ected with HSV-2 in the presence and absence of 2 mM TdR. The rate of DNA synthesis was measured by pulse
labeling
cell samples for 1 h with[3H]hypoxanthine.
Figure 2A shows therateof DNA
synthesis
inuninfected KB cells. When KB cells were
washed free of'TdR, a wave ofDNA
synthesis
occurred. Therate ofDNA
synthesis
increased rapidly for3 to 4 h.then decreasedby
8h. The cellnumberremainedconstant for 8to10h postreversal and then doubled. A second round of
DNAsynthesis
(corresponding
to the second Sphase) commencedat12 to 14h.When the cells
wereincubatedinthe presence of
TdR,
therateofhostcell DNAsynthesisremainedconstantat
less than
8%
ofthe reversed culture and therewas noincrease in cell number. Theresults are
essentially the same as previously reported (3).
The pattern of DNA synthesis in
synchro-nized cultures infected with HSV-2 was
mark-edlydifferent (Fig. 2B). In the absence of TdR,
the rateofDNA synthesis reached a
maximnum
approximately 5 h after infection and was never
greater than 30% of that found for uninfected
KB cells. DNA synthesis in the infected cells
thendeclined slowly for at least 16 h.
In contrast, the rate of DNA synthesis in TdR-blocked cells increased only slightly by 3 h after inf'ection and subsequently returned to a
lowlevel. These results show that DNA
synthe-sis in TdR-blocked infected cells occurs at a
muchreduced rate and suggests that the
inhibi-14 r A
12
10 6
5 -14
lo 4
x
UJ3 z
-2 z x 0 I = 4
3n
2
o
-B
A
/ \HSV-2 no TdR
6~~~~~
-/& oe.-HSV-2+ TdR
A A
O 2 4 6 8 10 12 14 16 18
HOURS POST REVERSAL
FIG. 2. Rate of DNA synthesis in synchronized culturesofuninfectedKBandHSV-2-infectedcells in the presence and absence of TdR. The rates were determinedby pulse labeling5.0-ml cellsamples (2x
105 cells/ml) with [3H]hypoxanthine for 1 h, as
indicated in MaterialsandMethods. Symbols: panel
A, 0,uninfected cells; 0, uninfectedin thepresence of2mM;panelB,A,infected cells, andA,infectedin thepresence of2mM TdRcells wereinfected at an input multiplicity of20PFU/cell.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:3.499.62.247.67.300.2] [image:3.499.267.449.238.569.2]INHIBITION OF HSV-2 REPLICATION
tion of viral replication may be due to an
inhibition of viral DNA synthesis.
Before conclusions could be drawn concerning
HSV-2 productionand the level of DNA
synthe-sis, itwasnecessarytoshow thatthe radioactive
DNA made after infection was viral and to
determine how much viral DNA was made in
thepresence of TdR.
CsCl density gradient analysis of DNA
synthesizedinHSV-2-infected KB cells in the
presence or absence ofexcess TdR.
Synchro-nized KB cellsinfected with HSV-2 were
incu-bated in the presence or absence of TdR.
Tritium-labeledhypoxanthinewasaddedtothe
cells 1 h afterreversal and 9 h laterthey were
harvested. Theextracted DNAwasanalyzed by
buoyant density ultracentrifugation in CsCl.
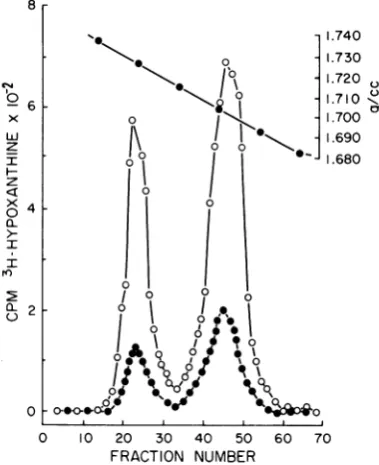
The radioactive profiles obtained are shown in
Fig. 3. Two bands of radioactivityweredetected
in infected cells incubated in the presence or
absence ofTdR. One band (1.700 g/cm3)
corre-sponds in buoyant densityto KB DNA (3) and
thesecond (1.725g/cm3) toHSV DNA (3). The
amount ofviral DNA synthesized in the
pres-8
6
x
w
z
I
z
x 4
0~
I I
10
D-2
u
1.740 1.730
.720 , 1.710 -" 1.700 O
1.690
1.680
0 10 20 30 40 50 60 70
FRACTION NUMBER
FIG. 3. CsCl density gradient analysis of DNA synthesizedinKB cellsinthepresenceorabsenceof
excess TdR. In each case. [3H]hypoxanthine was
added at a time corresponding to I h after TdR reversal. The cells werethen incubated foran
addi-tional 9h. Symbols: 0,DNA (2.8 gg) isolated from
HSV-2-infectedKBcells whichwereincubatedinthe
absenceofexcess TdR;*,DNA (5.8 Mg)isolated from
HSV-2-infected KB cellsand incubated in the pres-enceofexcess TdR.
enceof
TdR
wasmarkedly reduced.
From thesestudies
and othersweestimate thatonly
10% of the HSV-2 DNA found in the controls was synthesized in the presence of TdR. These results differfromthosepreviously
reported
for HSV-1 (3). In that case excess TdR had little effect on the rate or amount of viral DNA synthesis.Viral inhibition
of
KB cell RNAand
pro-tein synthesis. The observation thatnewviral DNA was made inHSV-2-infected
cells in thepresence ofTdRsuggests thatsome viral
func-tions were
expressed.
Supportive
evidence was obtainedby
examining anothercharacteristic ofHSV
infection.We asked whether HSV-2 infection of
TdR-blocked
KB cells resulted in the inhibition of RNA and proteinsynthesis. Cellular RNA and proteinsynthesis are notmarkedly
inhibited in mammalian cells by excess TdR (2, 16, 17). When cellsareinfected withHSV,
inhibition of host macromolecular synthesis isdependent
upon production ofviral-induced protein(s) (6, 13). Therefore, inhibition of host RNA and proteinsynthesis would constitute indirect evi-dence that viral functions are
expressed.
Table 2 shows the effect of2 mM TdR on RNA and protein synthesis in uninfected and infected KB cells. When the cells were incu-batedinthe presenceof
TdR,
neither RNAnor protein synthesis weremarkedly
inhibited. Incontrast, both RNA and protein synthesis in
HSV-2-infected KB cells were markedly inhib-ited, and the extent of this inhibition was enhanced
by
the presence of TdR.Ribonucleotide
reductaseactivity in HSV-infected cells grown in the presence and absenceof
excess TdR.HSV-1
replicates
in TdR-blocked KB cells (3). This characteristic has been ascribedtothe productionof a "new" or altered ribonucleotide reductase activity which is not subjectto allosteric regulation by TTP (3). DNA replication by this virus is not regulatedinthesame manner as host cell DNATABLE 2. Effect of TdRonuninfected and infected KB cell RNA andprotein synthesis
KBcell+TodrR %Inhibition
KB cell
Tdor
-___ ____RNAa Proteina
Uninfected 0 0
Uninfected + 5 12
Infected _ 57 68
Infected + 93 88
a
[3H
lhypoxanthine
or3H-labeled
aminoacids was addedat atimecorrespondingto1hafterTdR rever-sal. The cellswerethen incubatedforanadditional9 hinthe presenceorabsenceofTdR.23
VOL.14,1974
on November 10, 2019 by guest
http://jvi.asm.org/
[image:4.499.46.236.339.572.2] [image:4.499.251.444.549.626.2]COHEN, FACTOR, ANDPONCEDELEON
synthesis. Thus it was possible that the failure ofHSV-2 toreplicate would be reflected by the absence of a "new" ribonucleotide reductase activity similar to that found with HSV-1. Studies were undertaken to compare the ribo-nucleotide reductase activity found in unin-fected cells and in cells infected with HSV-1 and HSV-2.
Ribonucleotide reductaseactivity in synchro-nized KB cells is periodic, being highest in
extracts prepared at orjust prior to reversalof
the TdR block (3). Enzyme activity in the washedcellsremains at a maximum for 6 to 8 h post reversal, then decays rapidly during the
courseofthemitotic cycle (3). Incontrast,if the
TdR block is not reversed, the activity of the
enzyme remains essentiallyunchanged.
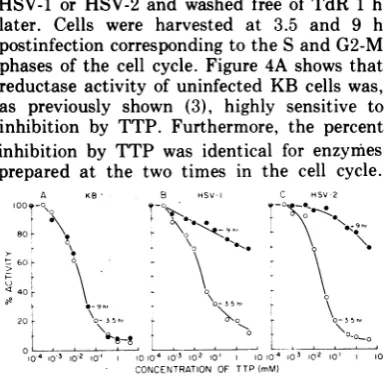
Synchronized KB cells were infected with HSV-1 or HSV-2 and washed free of TdR 1 h later. Cells were harvested at 3.5 and 9 h postinfection corresponding to the S and G2-M phases of the cell cycle. Figure 4A shows that reductase activity of uninfected KB cells was, as previously shown (3), highly sensitive to inhibition by TTP. Furthermore, the percent inhibition by TTP was identical for enzymes prepared at the two times in the cell cycle.
A KS B HSV-I C HSV-2
80 5X
<40.r E o TT o
ac y 9in,ea- of u e n i
20 Aur
~~~~L
of5 cs snh 35the V10-410 3 10-2 10-1 1010-4 10-3 12 tol 1010-4 103 IU2 1olCON\CENTRATION\OF TTP(mM)
FIG. 4. Effect ofTTPonribonucleotide reductase
activity in extracts of uninfected and infected KB
cells. A cultureofKB cellswass,vnchrovnizedbythe double TdR blockmethod. One hourpriortoreversal,
onethirdofthe culturewasinfectedwithHSV-1,one third with HSV-2, and the other third was
mock-infected. Cellswere infectedat aninputmultiplicity
of25PFUIcell, for both viruses.After 1hof incuba-tion the cultures were washed by centrifugation,
re-suspended in fresh medium, and allowed to grow.
Samplesweretakenat3.5and9hafterTdR reversal. Theactivitiesofeachpreparationweredetermined in the presence ofvarying concentrations ofTTP. The standard assay conditionswere used as described in Materials andMethods, except that TTPwasadded
to samples as indicated in the figure. One hundred
percent activity was determined by performing the assay in theabsence ofTTP. Symbols: 0, 3.5h;*, 9 h. PanelA, ribonucleotide reductaseactivity in ex-tracts prepared from uninfected cells (3.5 h = 1.94 U,9h = 1.84U); panel B, enzymeactivityinextracts
prepared fromHSV-1-infected cells (3.5 h = 1.42 U,
9h = 0.76 U);panel C, enzymeactivity inextracts
prepared fromHSV-2-infectedcells(3.5h = 1.52U,9
h = 0.91 U).
Figure 4B and C show that enzyme activity in
extracts prepared at 3.5 h postinfection either
with HSV-1 or HSV-2 had the samesensitivity
to TTP as did the enzyme prepared from KB
cells. However, by 9 h postinfection (Fig. 4B and C) ribonucleotide reductaseactivity in both HSV-1 and HSV-2 infected cell extracts was markedly resistanttoinhibition.More than 70%
ofthe maximum activity was present even in
the presence of 4.0 mM TTP. These findings indicate that a "new" reductaseactivity is also present in extracts obtained from the HSV-2-infected cells. Moreover, the extent of TTP inhibition was the same for both viral-induced enzymes.
Even though HSV-2 induceda"new" ribonu-cleotide reductase, itwas possible that the virus
was unable to induce the enzyme in the
pres-enceofTdR. Therefore, KB cells were infected
with HSV-2 in the presence of TdR. We ob-served that the presence of TdR during infec-tionhadnoeffect onthe specific activity of the
enzyme and no effect on the sensitivity ofthe
enzymetoTTP.Moreover, the observation that dialysis had no effect on thereductase activity
suggests that the insensitivity of the isolated
HSV-2 enzyme to exogenous TTP was not due to an alteration in nucleotide pool size. No synergistic effect on ribonucleotide reductase
activity wasdetectablewheninfected and
unin-fected extracts were mixed. The activity of a mixture of equal amounts of the two extracts wasequivalenttothesum of the activity of each
extract measured separately. From these
stud-ies, we conclude that the failure of HSV-2 to replicate in the presenceofexcess TdR cannot be explained by the inability of this virus to inducea "new" TTP-insensitive ribonucleotide reductaseactivity.
DISCUSSION
The present
study
compared
the pattern ofreplication of HSV-1 and HSV-2 in KB cells growing in the presence or absence of excess
TdR. Ourfindingssuggestthat TdRhas
differ-ent effects on the
replication
of HSV-1 andHSV-2. This observation adds an additional
biochemical
criterion whichdistinguishes
HSV-1 from HSV-2
(12).
Evidence has beenpresented
that this difference is not due to afailure of HSV-2 to infect TdR-blocked KB cells. This evidence includes detection of new
viral DNA albeit in low
quantity,
inhibition of hostcell RNA andprotein
synthesis,
andinduc-tion ofan altered ribonucleotide reductase
ac-tivity. Taken
together,
these resultssuggest
that the failureof HSV-2 to
replicate
inTdR-blocked KB cells occurred at the level ofviral DNA
replication
or later. Evidence that the24 J. VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:5.499.62.255.260.448.2]INHIBITION OF HSV-2REPLICATION
failure of HSV-2 to replicate in TdR-blocked
cells is at the level of DNA replication was
provided by the finding that both the rate of
DNA synthesis as well as the total amount of
viral DNAmadeweredrastically reducedinthe
presence ofexcessTdR.
A problem raised by this study concerns the
mechanism by which TdR inhibits HSV-2
repli-cation. It would be reasonable thatTdRaffects
viral replication at the level ofribonucleotide
reductase as it does in the host cell (1, 2, 15).
However, this explanation is not borne out by
ourresults. Unlike the uninfected hostcell, and
similarlytoHSV-1-infected cells,a"new"
ribo-nucleotide reductase activityhigflyresistantto
TTP inhibition is induced in HSV-2-infected
cells regardless of the presence or absence of
TdR.
Assuming that the ribonucleotide reductase is
functional in HSV-2-infected cells in the
pres-enceofTdR, what function(s) is inhibitedsoas
toprevent viralreplication?
TheTdR block may occurata stageof DNA
replication other than reduction of cytidine
nucleotide todeoxycytidine nucleotide. Recent
evidenceindicates that theproperties ofsomeof
theviral-inducedenzymesfound inHSV-1- and
HSV-2-infected cell extracts differina number
ofcharacteristics including inhibition by T1P.
Cooper (5) showed that TTP (0.1 mM)
inhib-ited the phosphorylation of deoxycytidine by
85% in extracts ofHSV-2-infected cells.
How-ever, TTP concentrations of 0.2 mM did not
significantly inhibit deoxycytidine
phosphoryl-ation in extracts of uninfected or
HSV-1-infected cells. Ogino etal. (10) showeda
differ-enceintheeffect of TdRnucleotideson
HSV-1-and HSV-2-induced TdR kinase. HSV-2 TdR
kinasewasmuchmoresensitivetoinhibition by
TdR nucleotides thanHSV-1 kinase.
Another possibility is that replication of
HSV-2 may be dependent upon somephase of
the host cell cycle. The replication of some
viruses including equine abortion virus (7) and
SV40 (11) is dependent upon some function(s)
(reviewed in ref. 7 and 11) present only during
the early S phase of the cell cycle. However,
HSV-1replication isindependent of the KB cell
cycle (3).
Experiments are now in progress to
investi-gate the mode of action of TdR on HSV-2
replication.
ACKNOWLEDGMENTS
Thisinvestigationwassupported by PublicHealthService grant DE-02623 from the National Institute of Dental Re-search.G.H.Cohenwassupported byaPublic Health Service
Research Career Development award A1-23801 from the National Institute of Allergy and Infectious Diseases.
WethankRoselyn J. Eisenberg, Lewis Pizer, and Weslev Wilcox for their generous help in the preparation of'this manuscript. Weacknowledge the excellent technical
assist-anceofMadeline Cohen.
LITERATURE CITED
1. Bjursell,G., and P. Reichard.1973.Effectsofthvmidine ondeoxyribonucleosidetriphosphate poolsand deoxy-ribonucleic acid svnthesis in Chinese hamster ovary cells. J. Biol. Chem.248:3904-3909.
2. Cleaver, J. E. 1967. Thymidine metabolism and cell kinetics. In A. Neuberger and E. L. Tatum (ed.). Frontiersofbiology. vol.6.North-HollandPublishing Co. Amsterdam.
3. Cohen, G. H.1972. Ribonucleotidereductaseactivityof synchronized KB cells infected with herpes simplex virus.J. Virol. 9:408-418.
4. Cohen, G.H.,R. K.Vaughan,and W. C.Lawrence.1971. Deoxyribonucleicacidsynthesisinsynchronized mam-malian KB cellsinfected withherpessimplexvirus.J.
Virol.7:783-791.
5. Cooper, G. M. 1973. Phosphorvlation of 5-bromodeox-ycytidine incells infected withherpes simplex virus.
Proc.Nat.Acad. Sci. U.S.A. 70:3788-3792.
6. Kaplan,A.S. 1973.Abriefreviewofthe biochemistry of herpes virus host-cell interaction. Cancer Res. 33:1393-1398.
7. Lawrence, W. C. 1971. Evidence for a relationship
be-tweenequine abortion(herpes)virusdeoxyribonucleic acid synthesis and the Sphaseofthe KB cellmitotic
cycle. J. Virol.7:736-748.
8. Lowry, 0. H., N. J. Rosebrough, A. L. Farr, and R. J. Randall.1951. Protein measurement with the Folin phenol reagent. J. Biol. Chem. 193:265-275. 9. Morris, N.R., and G. A.Fischer.1963.Studies
concern-ing the inhibition of cellular reproduction by deox-yribonucleosides. I. Inhibition ofthe synthesis of de-oxycytidine bya phosphorylatedderivativeof thymi-dine. Biochim.Biophys. Acta68:84-92.
10. Ogino, T., R.Shiman, and F.Rapp. 1973. Deoxythyini-dine kinase from rabbit kidney cells inf'ected with herpes simplex virus types 1 and 2. Intervirology 1:80-95.
11. Pages, J., S. Manteuil, D. Stehelin. M. Fiszman. M. Mark, and M. Girard. 1973. Relationship between replicationofsimianvirus 40DNAand specificevents
ofthe hostcellcycle.J. Virol. 12:99-107.
12. Rapp, F., and R. Duff. 1972.In vitro celltransformation by herpesviruses. Fed. Proc.31:1660-1668.
13. Roizman, B. 1971. Biochemical featuresofherpes
virus-infected cells. In W. Nakahara, K. Nishioka. T. Hi-rayama and Y. Ito (ed.), Recent advances in human
tumor virology and immunology. University Park Press, Baltimore.
14. Setlow, P. 1973. Deoxyribonucleic acid synthesis and deoxynucleotide metabolism during bacterial spore germination. J. Bacteriol. 114:1099-1107.
15. Skoog, K. L., B. A. Nordenskjold, and K. G. Bjursell.
1973. Deoxyribonucleoside-triphosphate pools and
DNAsynthesisinsynchronizedhamstercells.Eur.J.
Biochem.33:428-432.
16. Studzinski, G. P., andW. C. Lambert. 1969.Thymidine
as asynchronizing agent. I.Nucleicacid and protein
formation insynchronized HeLa cultures treated with
excessthymidine.J.Cell.Phvsiol. 73:109-118.
17. Xeros, N. 1962.Deoxyriboside control and
svnchroniza-tion of mitosis. Nature(London) 194:682-683.
25
VOL. 14,1974