Transcription and Replication of Vesicular
Stomatitis Virus:
Effects of
Temperature-Sensitive
Mutations in
Complementation Group IV
ARLETTE COMBARD, CLAIRE MARTINET, CHRISTIANE PRINTZ ANE, ANNICK FRIEDMAN, AND PIERRE PRINTZ
Institut deMicrobiologie, Universite Paris-Sud, Centre d'Orsay, 91405 Orsay,France Received forpublication3 December 1973
Temperature-sensitive (ts) mutants ofvesicular stomatitisvirus belongingto
the RNA-complementationgroupIVwereinvestigatedunder various conditions tostudy both their RNA and protein syntheses. In infectedcells maintained at
39.2 C, viral RNA species were recovered only in the 13 to 15S region ofthe gradient in an amount depending on the ts mutant used. In the presence of
cycloheximide at 39.2C, the primary transcription wasdeficient, especially for
28SmRNA production. When mutant-infected cells wereshiftedto
nonpermis-sivetemperature, ashutofiof28S mRNAsynthesis occurredas ageneral feature.
On the contrary under thiscondition, thetwo mutants chosen, ts IV100 andts
IV111, behaved very differently in their 13 to 15S and 38S RNA production.
However, treatment with cycloheximide at the time of the transfer to 39.2 C resulted inasimilarrecoveryof 13to15SRNA in bothmutants,whereas the 28S remained very depressed. The viral proteins synthesized by cells infected with
thesametwomutantsalsoshowedadistinctpattern, especially regarding the N protein;acorrelation between 38S RNA andprotein N syntheseswastentatively
drawn. The whole set of data suggested that the lesion in group IV mutants
concerned aviralstructural proteinrequiredfortheprocessof in vivo transcrip-tion and which probably intervened inthereplication mechanism.
Progress hasrecently been madeinthestudy
ofprocessesinvolvedinthe productionof vesic-ular stomatitis virus (VSV), making use of mutants isolated invarious laboratories (9, 11,
30). Temperature-sensitive (ts) mutants,
una-bleto synthesize sizableamountsofviralRNA
at high temperature (RNA-), have been
ac-tively investigated as being the most useful
toolstodelineate the mechanisms ofboth viral
transcription and replication. New facts
emerged from our previous studies (16, 20) of
twomutants, tsIandtsIV100, representative of two RNA- complementation groups, I and IV
(F. Lafay, thesis, University of Paris, France, 1972) respectively. Firstly, transcription and
replication appear to be tightly linked, and we
suggest that a common subunit intervenes in vivo in transcription and replication. In addi-tion,results obtained withts IV100prompteda new hypothesis, in which the enzyme which functions in vitro as a transcriptase (2) would require another virus-coded polypeptide to
pro-duce faithful transcription in vivo. In this
paper, evidence obtained by studying various
group IV mutants and which further supports
thehypothesis ispresented.
MATERIALS AND METHODS
Cells. In all RNA analyses,HeLaconfluent
mono-layers were used under conditions already described (20). Forsomeproteinstudies,chickenembryo fibro-blasts (CEF)wereculturedasreported(9).
Virus.Temperature-sensitive(ts)mutantsofVSV (Indiana strain), herein investigated, belong exclu-sively to complementation group IV. Virus stocks containing only standard B particles wereobtained andwerechecked for low percentage of revertants,no
residual growth at 39.2C, and by complementation
test as described (20). The wild-type virus grew at 39.2C, aswellas at30C.
Chemicals. Actinomycin D, a generous gift from
MerckSharp andDohme, French branch,wasusedat a final concentration of 10 gg/ml. Cycloheximide, dilutedin medium to 100
ltg/ml,
andsodiumdodecyl sulfate (SDS) were purchased from Serva, Heidel-berg, Germany. RNase T,wasobtained from Sankyo (Tokyo, Japan) and pancreatic RNase A was obtainedfrom Nutritional Biochemicals Corp. (Cleveland, Ohio). All labeled compounds were from C.E.A.
(Saclay, France): [9Hluridine(20Ci/mmol), [3H
]leu-922
on November 10, 2019 by guest
http://jvi.asm.org/
VSV COMPLEMENTATION GROUP IV TS MUTANTS
cine (33Ci/mmol), [3H]tyrosine (49 Ci/mmol), and a mixture of [14C]amino acids (100 mCi/mmol).
Analysis of viral RNA. HeLa cells were infected (-10 PFU/cell, unless otherwise specified) and treated with actinomycin D as previously described (20). When cycloheximide was used, addition of radioactive nucleoside occurred 15 min later. After labelingw4h [3H luridine(20
gCi/ml
unless otherwise specified in figure legends), cells were harvested at the endofthe indicated incubation periods. From cyto-plasmic extracts in reticulocyte standard buffer, made1%with respect toSDS, solubilized RNAs were analyzed by centrifugation in the Spinco SW 27.1 rotor in 15 to 30%sucrose-SDS gradients. All methods and buffer compositions were previously described (20). After centrifugation (21,000 rpm at 17 C for 15 h), the gradients were fractionated and monitored as previously described (20).Annealing conditions. Annealing of RNA species was achieved as follows: portions (1,500 3H-counts/ min) were mixed with virion RNA (4 to 10
Ag/ml)
extracted according to the method of Montagnier (17). Duplicate samples were made toafinal volume of 400 gliters and a final concentration of 2 x standard saline citrate (0.15 M NaCl, 0.015 M sodium citrate). They were boiled for 3 min, hybrid-ized 1 h at 70 C, and then divided into two equal portions. One of them was digested for 30min at 37 C by pancreatic RNase A (5Ag/ml) andT, (30 units/ml). All samples were then acid-precipitated inthe presence of 0.08M sodium pyrophosphate and 100,g
ofyeast carrier RNA per ml. Materials trapped on membrane filters, type HA, were assayedfor radioac-tivity as usual. Controls simultaneously conducted lacked added virion RNA.
Labeling and analysis of viral proteins. CEFor
HeLa cells were first exposed for 90min at 37 C to
actinomycinD (1 ug/ml) and, afterviral adsorption (40 min at room temperature), were covered with diluted 1:10 (vol/vol) Eagle minimal essential
me-dium(Eurobio) containing 1%calfserum.Amixture of [3HIleucine and [3H]tyrosine was added at inter-vals after infection for 60-min pulses (20 MCi/ml).
Cellularextracts werepreparedbydisrupting the cells with aDounce homogenizer in awaysimilartothat used for RNA extraction(20). Proteinswereextracted
from cell fractions in precisely the same way as
previously described (21). After adding one-tenth volume of glacial acetic acid, the suspensions were
made0.5 M withurea and 1%with SDS, incubated for1hat37C,andfinally dialyzedatroom
tempera-turefor 16 h againstphosphate buffer (pH 7.2, 0.01
M) containing 0.1% SDS, 0.5 M urea, and 0.1% 2-mercaptoethanol. Viral proteins were analyzed by
polyacrylamide gel electrophoresis performed as
de-scribed(29),by using7.5%acrylamidegelat 5mA/gel for5h.Gels slicedinto 1-mmlengthswere
depolymer-izedin0.2 ml ofhydrogenperoxide (25%)at55Cfor4
h priortothe additionofBray scintillation fluid. For
theidentification ofviralproteins,complete reliance
was placed on the migration of intracellular [3H]proteins coelectrophoresed with authentic viral [4C] proteins extracted from purified virions (21).
Under these conditions it was regularly observed that intracellular [3HIprotein G migrates faster than the virion marker ["C ]protein G, as reported by Kang and Prevec (14). Radioactivity was measured in an IntertechniqueSL40 spectrometer assisted by a dou-ble-labeling computer program.
RESULTS
Viral RNA synthesis at 39.2 C. A first
approachto viral RNAsynthesis in
complemen-tation group IV has been done by using the prototype mutant tsIV100 (20). Parallel studies
of three mutants, ts IV62, ts IV111, and ts
IV100, now make it possible to generalize about some properties of group IV mutants. As
com-pared to wild type, the total amount of RNA
synthesized at 39.2 C is always less than 10%.
NodegradationofRNAalready synthesizedcan
be detected, and the rate ofsynthesis appears
constant without amplification throughout the
cycle.Moreover, thepattern ofthe RNA
synthe-sized at 39.2 C shows no change at any time
after infection. It is characterized by the
pres-ence oflight RNAs at the top ofthe gradient
(less than lOS); in addition, among specific
VSV RNApeaks,only 13to15S RNAisreadily
identified (Fig. 1, full-circle curves).
Neverthe-less, in the same experimental conditions the
largest proportions of 13 to 15S RNA are
ob-served with ts IV111 and the smallest with ts
IV100. Therefore, a minimal and maximal
re-sidual viral RNA synthesis amongthe mutants
studied is provided by ts IV100 and ts IV111,
respectively. Consequently,further studieswere
carried outwith thesetwo mutants only.
Viral RNAsynthesisat39.2 C in
cyclohex-imide-treated cells. Results stated above
showed thatsomestep(s)intranscription,
espe-cially for thesynthesis of28S, is(are) affected
inthegroup IV mutants. Inordertodetermine
whether any alteration could be detectedfrom
theearly stage ofprimarytranscription defined
by Huang and Manders (13), RNAsynthesisat
39.2Cwasstudiedinthepresence of
cyclohexi-mide whichwasaddedjustafter infection(Fig.
1, open-circle curves). Whatever the mutant
used, the RNA pattern obtained under these
conditions differsfromthatobtainedat30C in
the presence of cycloheximide (Fig. 1, star
curve) which shows all the characteristics of
VSV primary transcription (13). The striking
facts werethe lack of28SRNA andthe similar-ity of the twoprofiles withorwithout
cyclohexi-mide. Despite the apparent importance ofthe
RNA synthesis by ts IV111 under these
condi-tions, this mutant behaves as RNA-, because here wecompare the RNAsynthesizedat39.2C
VOL.13,1974 923
on November 10, 2019 by guest
http://jvi.asm.org/
4 C D39.2C, we observed a
depression
of all viralA tomI" \RNA peaks with bothmutants (Fig. 2,
full-cir-28s Iss cle
curves).
Thesingle-stranded
28S
RNA,
com-$ +I* plementary to the virion RNA synthesized at
30C, was not recovered in the upward shift
a lL l conditions. The small amount of28S still
de-tected was
essentially
RNase-resistant. On theother hand, the two profiles strikinglydiffered
4. ^\ *Iy \ A concerning 13 to 15Sand 38S RNAs. Actually,
after the
upward
shift, ts IV100 and ts IV111produced
both RNAssedimenting
in the 13 to15Sregion which,like truemRNAs,hybridized
x *. completely to virion genome (98%). However,
s44I when compared to the quantity of 13 to 15S
normallypresent at 30 C, the amount of 13 to
4. P / 15S was only decreased from 10 to 50% in the
case ofts
IV111,
whereas it was reduced lessthan50% for ts
IV100.
c , $ . The38S
peak,
inourupward
shiftconditions,
t.
m was never detected in the ts IV111 profile,4 whereas it was always present in that of ts
IV100, although in various proportions (10 to
90% of its 30-Cyield).
I10 20 30 40 50 _______________________
FRACTIONS 2 A
tslIIItN
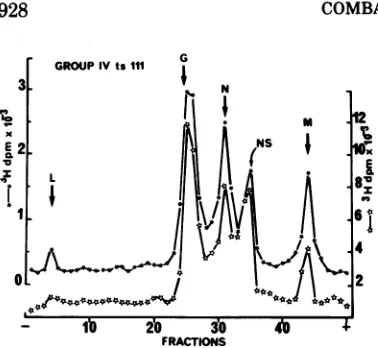
FIG. 1. Sucrosegradient separation ofRNAspecies
made by ts group IVmutants in the presence or ill|
absence of cycloheximide. Equivalent samples of
HeLa cellswereinfectedeitherbyts
IVI00,
tsIV62,orE.I
\ / I\
ts
IVI11
atthesamemultiplicity ofinfection (- 20). 28S 18sAfter 45 min of adsorption at room temperature,
/
actinomycinDwas addedto afinal concentrationof\
10
,g/ml.
Severalsamples (seesymbols)weretreated iwith100lAgofcycloheximideper ml.Then allsamples
werelabeled with40
,Ci
of[3H]uridineper ml. Cells /were harvested at 3 h p.i. Cytoplasmic RNAs were 0.5.
separated on 15 to 30% sucrose-SDS gradients, col- I lected into50fractions, andacid-precipitated by5% x
trichloroacetic acid. 0,RNA profile of infectedcells X I
at 39.2C; 0, RNA profile of infected cells treated I withcycloheximideat39.2 C;
4,
RNAprofile ofcells ts100 infected withtsIV111 and treated withcycloheximideat30C. Peaksofradioactive RNAidentifiableat30C 0.5
in infections with ts IV62 and tsIV100, identicalto 0. thosesynthesized bytsIV111, are notshown.
with the RNA obtained at 30C in primary __
transcription whichaccounts foronly10%ofthe lb1To io 40
total RNAsynthesis (13, 28). FRACTIONS
Effect of anupward temperature shift on FIG. 2. Sucrosegradient analysis of RNA species
RNA synthesis. As was already mentioned synthesized bytsIVI00-andtsIVIII-infectedHeLa
with ts IV100 (20), a rapid change in RNA cells at 39.2 Cafterprior incubation at 30 C.Infected
synthesis follows a temperature shift to 39.2 C HeLa cells with tsIVI00ortsIVIII werelabeled in
after 3 hat 30C.Hence,suchexperimentswere the presence of actinomycin D with 20 MCiof
[3H]uri-simultaneously conducted withts IV100 andts
dine
perml 3to4hp.i.RNAswereanalyzedasinFig.IV111. Immediately afterIVil.Imediaely fterthetheupward tempera-pwar temera-a t i,39.2CtsatMutant-infected3 h p.i.; 0, cells transferred from 30 C tots mutant-infected cells at
ture shift, RNAs werepulse labeledfor 1 hand 39.2C; *, infectedcellsmaintained at 30 C for 4 h.
compared with the corresponding control at As in Fig. 1,and for the same reason, only one profile
30C (Fig. 2, star curve). After transfer to isshown.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:3.494.65.251.65.324.2] [image:3.494.267.455.316.545.2]VSV COMPLEMENTATION GROUP IV TS MUTANTS
The rate of RNAsynthesis declined after shift
to39.2 C as shownbythegradual decay of RNA
labeling, without specific alteration of any one
peakduring the second and third hour after the
shift (not shown). Finally, the effect of prior
incubation at 30 C disappeared, because 3 h
after a shiftthe profile became the same as if it
wasobtained from infected cells maintained at
39.2 C throughout the experimental time course
(Fig. 2, open-circle curves).
It canbe seen from the control curve in Fig.
2that even at 30C all the mutants in group IV
induce a remarkable RNA profile, because 38S
isgenerally the highest peak, and 28S is
especi-allybroad. Inthis important and heterogeneous
28S region, additional RNA categories appeared.
One ofthem, 23S RNA, was previously
men-tioned (20) but was not regularly observed
either at 30 C or at 39.2C depending on
un-known contingencies.
Combined effect of cycloheximide
treat-ment and upward temperature shift. In
up-ward temperature shift experiments, viral
pro-teins synthesized during the first 3 h at 30 C
may interfere withormask someconsequences
ofthe mutation taking place at 39.2 C.
There-fore, to interpret the former results,
interven-tion of these proteins must be estimated by
preventing further protein synthesis after the
shift. Thus, cycloheximide was added at the
time of the upward temperature shift (3 h
postinfection [p.i. ]) in cells infected
respec-tively by the ts IV100 and ts IV111 mutants
(Fig. 3), whereas a sample was maintained in
parallel at 30C as a control (Fig. 3, square
curve). The transferto39.2C ledto adecrease
ofboth classes ofmRNAs, whichwasespecially
drastic atthe level of28S.
We verified withwild-typevirus thatnoneof
these facts was the result either ofthe direct
action ofcycloheximideorof itscombined effect
with theupwardtemperatureshift. First,asdid
otherauthors(19,30),wefound thatadditionof
cycloheximide atany timeduringan infectious
cycle at 30C was conduciveto ashutoffof38S
RNA synthesis (visible on the control curve),
whereastranscriptionof28S and13 to15S RNA
was unaffected oreven enhanced as compared
with cells without the drug. Second, the
com-bined effect of a temperature change and
cy-cloheximide treatment was shown to
merely
promotean extraamplificationoftranscription.
Now comparisonofthetwoprofilesobtained
for each mutant after an upward temperature
shift in the presence or absence of
cyclohexi-midepointedoutthe roleofnewly synthesized
proteins at 39.2 C. Although both types of
profiles have already been reported, but from
nonsimultaneous analyses, we preferred to
com-pare new curves resulting from another
experi-ment. De novo protein synthesis at 39.2 C
resulted for both mutants in a dramatic
reduc-tion of the amount of 13 to 15S and almost a
disappearance of 28S RNA (Fig. 4). Moreover,
38S RNA appeared when cycloheximide was
omitted, but only with ts IV100.
Protein synthesis by mutants ts IVIOO and
ts IVlll. The synthesis of viral proteins has
been investigatedinconditions similar tothose
used forthe RNAsynthesis. In our hands, CEF
supporting a much more active and rapid
pro-teinsynthesis than HeLa cells, without
exhibit-ing consistent differences, were preferentially
used.
Figure 5showsthe electrophoretic profiles of
the viral proteins extracted from CEF infected with the mutants ts IV100 and ts IV111 at 30 C,
39.2C, and inupward temperature shift
condi-tions. All five viral proteins were present in
normal amounts in cells infected at 30 C with
thetwomutantsand werelabeledfrom2to 3h
p.i. At nonpermissive temperature and during
the same labelingperiod, the level ofsynthesis
was markedly reduced, espeically with the
mu-28s ls
x~~~~~~~~
FRACYONS
FIG. 3. Sucrose gradient pattern of RNA synthe-sized by ts IV100 and ts IV111 after addition of cycloheximide in upwardshift experiments.Infected
andactinomycin D-treated HeLa cellswereincubated
3hat 30C.At3hp.i., cycloheximidewasadded(100 ug/ml). Fifteenminuteslater,cellswerelabeled with
20AtCiof[8H]uridineper mlfor1handanalyzedasin
Fig.1.0,tsIVI I I-Infectedcellstransferred from30C
to 39.2Cat 3hp.i. timeofadditionoftheantibiotic; 0,tsIV100-infectedcells treatedastsIV111-infected
cells; 0, control infected cells maintained at 30C after additionof thedrug.
VOL.13,1974 925
on November 10, 2019 by guest
http://jvi.asm.org/
[image:4.494.248.440.361.532.2]T.
FIG. 4. Sucrose gradient centrifugation of viral
RNA madebytsIV100-ortsIV111-infected cells,in
thepresence orabsenceofcycloheximide inupward
temperatureshift experiments. After infectionwith ts
IV100 ortsIVi11, actinomycinD-treated cells were incubated 3 h at 30C. At 3 h p.i., cells were transferred to 39.2C, and one of the duplicates
received cycloheximide (100 Ag/ml). At 3.15 h, all
cellswerelabeled with20uCiof[3Hjuridinepermlfor
1h.0,Infectedcells withcycloheximideadded at the
timeoftheshift;0,infectedcells without
cyclohexi-mide.
tant tsIV100;inthe lastcase,onlytheproteins N and NS emerged from the background. At
39.2C, thetsIV111-infectedcellsgavea
better-defined profile, but the N protein peak
ap-peared low as compared with the other ones.
With bothmutants, theLproteinwasvirtually
absent. The pattern of protein synthesis
ob-tainedwiththetwomutantsin theupwardshift condition are shown in panel B. The main
difference between thetwomutantswas inthe
behavior of the N protein. During the 1-h pulse
at39.2 C whichfollowed the 2-hpulse at30C, the N protein peak decayed drastically incells
infected withts IV111, but wasnot affected in
thecaseofthe mutanttsIV100.Thisresult has
been regularly observed in repeated experi-ments.Thedrop of theNproteinwasduetoan
absence of synthesis rather than to a rapid
destruction ofnewlysynthesized polypeptides, because in a chase experiment of labeled
pro-teins at 30C no drastic effect wasobserved at
39.2C atthelevelof anyviralprotein
(unpub-lished data). The L protein synthesis was
re-duced in a similarway forboth mutants.
A peculiar effect of the upward shift was
observed at the level of the G protein; its
synthesis was markedly increased in cells
in-fected with ts IV100 and ts IV111. This effect
wasnottransitoryand continuedtobeobserved
during successive 1-h pulses.
The transcription process in cells infected
withmutant ts IV100 or ts IV111 wasincreased
when upward shiftwas combined with a
treat-ment ofcycloheximide (Fig. 4). Toobservethe
effecton protein synthesis, HeLa cells or CEF
both infected with ts IV111 havebeen treated
bycycloheximide (100,ug/ml)atthetime ofthe
upward shiftto 39.2 C and extensively washed
before the 1-h amino acid pulse. Compared with
Fig. 5, Fig. 6 shows that the same profile of
proteins is obtained, but the proteinpeaks are
higher; this may result from the presence of
moremRNAinthat condition.Parenthetically,
the electrophoretic profiles were qualitatively
similar with proteins extracted from CEF or
HeLa cells.
DISCUSSION
Analysis of viral RNAs made at 39.2C after
infection with any one ofthegroupIVmutants
demonstrates that both the expected
transcrip-tionandreplication cannottake place.
Discuss-ing our previous data (20), we proposed that
transcriptionwasthestepdirectlyconcerned by
the mutation. In the present paper, shorter
labeling pulses and upward temperature shift
experimentsconductedtoamplify theeffects of
the mutation confirm that transcription is
un-doubtedly affected. Thisappearsfromthe early
phase of primary transcription during whichare
observed thealterationsinRNA synthesis
char-acteristic ofgroup IV mutants, i.e., lack of28S
and a lower degree of 13 to 15S transcription
(Fig. 1). In a recent study, Unger and
Reich-mann(28) reported the deficient RNAsynthesis
of one mutant of group IV at nonpermissive
temperature. They argued about the special
alteration of the long mRNA, but there is no
real discrepancybetween theirobservationand
ourown, because in conditions similar to theirs (5 h at 39.2 C) we actually found labeling in the 28S region, although it was not recovered as one
conspicuous peak (20). In addition, when they
examined the primary transcription of their
group IV mutant, theynoticed thesame quanti-tative change atnonpermissive temperature as we related. Because, bydefinition, among viral
on November 10, 2019 by guest
http://jvi.asm.org/
[image:5.494.72.253.73.333.2]VSVCOMPLEMENTATION GROUP IV TS MUTANTS
N
' I --
4Atd
4;=;RS-/V\f\
^A
lN
A
88
L G (P5 64(5
44
4
4
1 20 30 40 50 ¶0
io
30 4' 50 [image:6.494.38.443.76.335.2]FRACTIONS
FIG. 5. Electropherograms of virus-specific proteins extracted from CEF infected at a multiplicity of- 50
with ts IV100 or tsIViii underpermissive, nonpermissive, or upward shift conditions. After an adsorption period of 40 min at room temperature, three plates (60 mm in diameter) for each mutant were covered with 2 ml of1:10(vol/vol)-diluted minimal essential medium and incubated at 30 C (two plates) or 39.2 (one plate). Two hourslater, [3H]leucineand [3H]tyrosine (10gCi/mleach) wereadded to one culture at 30 C and to the other at 39.2C; cells were harvested after 1 h of labeling (2 to 3 hp.i.). The remaining plate at 30 C was brought to 39.2Cat2.30hp.i. and labeled from 3 to 4 h p.i. Proteins were extracted from cytoplasmic contents ofcells, disruptedwithaDouncehomogenizer, with 10% acetic acid, 0.5 M urea, 1% SDS, and 0.1%2-mercaptoethanol. Extracted [3Hlproteins were subjected to electrophoresis in 7.5% neutral SDS acrylamide gels along with marker[14C]proteinsextracted from purified virions. Vertical arrows show the peak positions of the four marker
[14C]proteins. Left hand side, ts IV100; right hand side, ts IV111. Panels A: 0, permissive temperature; 0
nonpermissivetemperature. Panels B: upward shift conditions.
proteins only those associated with the virion
are involved in primary transcription (13), it
follows that one of these structural proteins,
essential particularlyfor 28S RNAsynthesis, is
alteredin group IVmutants.Moreover, in vitro
transcriptase activity at 39.2C has been
dem-onstrated with this group ofmutants (20, 26),
whereas it failed for group I mutants; in the
lattergroupthe lesionseems tobemorelikelyin
the enzyme thanin the template (P. Printz et
al., in R. D. Barry and B. W. J. Mahy, (ed.),
Negative strandviruses,inpress; R. R.Wagner,
personal communication). Therefore, besides
eventual host factors (19), we mustemphasize
that another viral structural protein, which for
convenience will betermedprotein IV,is neces-sary to obtain a faithful in vivo transcription.
The ts event renders it rapidly inactive at
nonpermissive temperature.This is observed for
protein IV of the incoming virions in primary transcription (Fig. 1), as well as for the new
molecules made at 30 C and shifted to 39.2 C
when additional protein synthesis was
pre-vented by cycloheximide (Fig. 3). However,
whencells infected either by tsIV100or tsIVIli
werekept 3 h at thenonpermissive temperature
and then shifted downward to the permissive
one inthe presenceofcycloheximide,
transcrip-tion was completely recovered (not shown).
Therefore, the ts event does not seem to be
irreversible, and protein IV is not degraded at
high temperature.
Upward temperature shift experiments, with
and without cycloheximide addition, posed the
problem of the effect of the mutation in the
group IV mutants at the level of replication.
First, there is apersistant synthesis of 38S with
tsIV100 after anupwardshiftwithout
cyclohex-VOL. 13,1974 927
on November 10, 2019 by guest
http://jvi.asm.org/
(NS .
: ~ ~
- 1b 2b 3'0 FRACTIONS
FIG. 6. Electropherograms of virus-specific pro-teins extracted from CEF or HeLa cells infected at a
multiplicity of '-.50with ts IVIII in an upward shift
experiment combined with a cycloheximide
treat-ment. Afteradsorption, the plates (60 mm in
diame-ter) were covered wvith1:10 (vol/vol) dilutedminimal essential medium andincubated at 30 C for 2 h (CEF)
or 3 h (HeLa cells). After that period at 30 C, the
medium was drained, and new medium heated at
39.2 C and containingcycloheximide (1001lg/ml) was added. The plates, kept at 39.2 C for 50 min, were then washed extensively and covered with 2 ml of
medium devoid of cycloheximide, but heated at
39.2 C. Cells, maintained at 39.2 C, were pulse
la-beled with [3Hjleucine and [3H]tyrosine (10y.Ci/ml each) from 3 to 4 h p.i. (CEF) or from 4 to 5 h p.i.
(HeLa cells). Cytoplasmic extracts were treated as
described in Fig. 5. Vertical arrows show the peak
position7s of the four marker ['4C]proteins. *, CE fibroblasts; *, HeLa cells.
imide. Second, neither 38S nor any RNA species noncomplementary to virion genome are de-tected with ts WIVllunder the same conditions. Recent reports (19, 30; P. Printz et al., in press) gave evidence ofan equilibrium between tran-scription and replication and of a shift of this equilibrium towards transcription in the ab-sence of replication by devotion of the whole RNA polymerase activities to mRNA produc-tion. After an upward temperature shift, only the addition ofcycloheximide allowed this in-creased transcriptase activity in both mutants (Fig. 4). Accordingly, even with tsIVi11 in the absence of cycloheximide, transcription was still restricted as if equilibrium was preserved, arguing for a potentially present replicase func-tion although unable to finally lead to 38S or possible precursors (15). Whereas the recovera-ble transcriptase activity is nearly identical for both mutants in the presence of cycloheximide (Fig. 3), in upward shift experiments without the drug (Fig. 2) the balance between
replica-tion and transcription is notthesame. Lack of
28Sandaloweramountof 13 to15S RNA being
a common effect oftheir mutations, weascribe theapparentdiscrepancybetween the behavior ofthe two mutants to some subtle differences inprotein IV, relevanttothe site of the
muta-tion.
Study of translationwasundertakento
possi-bly reveal other differencesatanylevelevenin
transcription (undetected by RNA gradient
analyses). Analysis of protein synthesis at
39.2C merely reflects the respective deficiency
of RNA synthesis in ts IV100- or ts
IV111-infected cells (Fig. 5). Thisis in agreementwith
Wunner andPringle's results (31). Thus, as in
RNA study, upward temperature shift was
performed to sensitize the detection of the
mutational effects. In such experiments, three
particularities appear intheelectropherograms
(Fig. 5 and 6). The behavior of G protein, as
discussed elsewhere (P. Printz et al., in press),
is notgroupIV-specific and could be duetoits
particular biosynthesis (10). The reduced
syn-thesis of Lprotein, in the sameproportion for
both mutants, may well be ascribed to their
common deficiency of28S mRNA (25; D.
Bal-timoreetal.,in R. D. Barryand B. W. J.Mahy,
(ed.), Negative strand viruses, in press). If L
protein is responsible for the transcriptase
ac-tivity detected in vitro (7), absence of new
moleculesofL transcriptase,alongwithfailure
ofreplicationat39.2 C withoutpriorincubation
at 30C, would explain that no amplification
occurs in RNA synthesis with group IV mu-tants. Thus, we conciliate two explanations
suggested by Unger and Reichmann, namely,
lack of replication and block in additional
synthesis ofproteins required for transcription
(28). Finally, the major difference in protein
synthesisbetweenthetwo mutantsinvestigated
concerns the synthesis of N protein; they also
differintheir 38Sproduction (Fig. 2).Asanew
protein synthesis is required for 38SRNA
for-mation(19, 30; Fig. 3),acausalrelationshipcan
be drawn from these two observations. If the recovery ofintracellularvirion RNAwas
pack-aged primarily with Nprotein, unstabilization
of38S RNA might result from the lackofthis
protein. This would partially explain protein
requirement forreplication, becauseother viral
proteins also appear to be involved inthe 38S
RNAsynthesis (manuscript inpreparation).
Anotherexplanation forthe different
behav-ior, with regard to the replication process,
between the two mutants in the same
com-plementationgroupcould be given if protein IV,
differently affected by the various ts point
on November 10, 2019 by guest
http://jvi.asm.org/
[image:7.494.65.254.55.229.2]VOL.13, 1974 VSVCOMPLEMENTATION
mutations, is involved in severalfunctions. Our
results make it conceivable that protein IV is
concerned with some aspects of signal
recogni-tion and, thus, could intervene both in tran-scription andreplication. Effectively, transcrip-tion is supposedto set on the putative start or stop signals (18, 23) along the template. The presence ofintracellular38S RNA complemen-tary to virion RNA (D. Baltimore et al., in press; S.M. Perlman and A.S. Huang,
submit-ted for publication) suggested that replication
might also function by opening one initiation
site, with all others remaining masked.
In-teracting with either the template, the tran-scriptase, the putative replicase, or with any
complex between them, protein IV should be
devotedtosignals switch-on and -off orsimply
to their recognition, giving quantitative and
qualitativeefficiency to transcription or
replica-tion orboth. This hypothesis wouldaccount for
the observation thatingroup IV mutants RNA
synthesis in vivo at 39.2C occurs as if
tran-scriptasealonewas atwork, becauseas inthe in
vitro reaction (3) (i) input genomes of group IV
mutants arecompletely transcribed (8),and (ii)
thesemutantsgivemostly RNA speciesofshort
sizes (4, 12, 13, 20, 30). Inaddition,we
demon-stratedthat transcriptionof28S RNA and13 to
15S RNA may occur independently, which is
consistent with some sort of sequential
tran-scriptionofthe VSV RNA (22).
Protein IV cannot be identified by merely
studying in vivo protein synthesis, because,
besides the transcriptase L protein andgenome
RNA, the complete VSV transcription
appara-tuscomprises in vitro(5,6, 27), aswellas in vivo
(1), protein N,
NS,
and eventual minorpro-teins; protein IV could be any one ofthem. Its
proposedfunctions maybe modulated bysome
chemical transformation (24).
ACKNOWLEDGMENTS
ThisresearchwassupportedbytheCentre National de la Recherche Scientifiquethroughthe L.A. 136, theFondation pour laRechercheMedicaleFrancaise, and the Commissariat al'EngergieAtomique.Saclay, France.
We thankA.Berkaloffformanyhelpfuldiscussions. For twoofus,C. Martinet and C. PrintzAne,thisworkispartofa
thesis(Doctoratd'Etat).
LITERATURECITED
1. Baltimore,D. 1971.Virus-associatedpolymerase,p. 124. In G. E. W. Wolstenholme and M. O'Connor (ed.),
Strategyofviralgenome.ACibaFoundation
Sympo-sium.
2. Baltimore, D., A. S. Huang, and M. Stampfer. 1970. Ribonucleicacidsynthesisof vesicular stomatitisvirus, II. AnRNApolymeraseinthe virion. Proc. Nat. Acad. Sci. U.S.A.66:572-576.
3. Bishop, D. H. L. 1971. Complete transcription by the
3ROUPIVTSMUTANTS 929
transcriptase of vesicular stomatitis virus. J. Virol. 7:486-490.
4. Bishop, D. H. L., and P. Roy. 1971. Kinetics of RNA synthesis by vesicular stomatitis virus particles. J. Mol. Biol. 57:513-527.
5. Bishop, D. H. L., and P. Roy. 1972. Dissociation of vesicular stomatitis virus and relation of the virion proteins to the viraltranscriptase. J. Virol. 10:234-243. 6. Emerson, S. U., and R. R. Wagner. 1972. Dissociation
and reconstitution ofthe transcriptase and template activities of vesicular stomatitis B and T virions. J. Virol.10:297-309.
7. Emerson, S. U., and R. R. Wagner. 1973. L protein requirement for in vitro RNA synthesis by vesicular stomatitis virus. J. Virol.12:1325-1335.
8. Flamand, A., and D. H. L. Bishop. 1973. Primary in vivo transcription ofvesicular stomatitis virus (VSV) and temperature-sensitive mutants of five VSV com-plementation groups. J. Virol. 12:1238-1264. 9. Flamand, A., and C. R. Pringle. 1971. The homologies of
spontaneous and induced temperature-sensitive mu-tants ofvesicular stomatitis virus isolated in chick embryo and BHK21cells. J.Gen. Virol. 11:81-85. 10. Grubman, M. J., and D. F. Summers. 1973. In vitro
protein-synthesizing activity of vesicular stomatitis virus. J. Virol. 12:265-274.
11. Holloway, A. F., P. K. I.Wong, and D. V. Cormack. 1970. Isolationand characterization of temperature-sensitive mutants of vesicular stomatitis virus. Virology 42:917-926.
12. Huang, A. S., D. Baltimore, and M. A. Bratt. 1971. Ribonucleic acid polymerase in virons of Newcastle disease virus:comparison with the vesicular stomatitis viruspolymerase. J. Virol. 7:389-394.
13. Huang, A. S., and E. K. Manders. 1972. Ribonucleic acid synthesis ofvesicular stomatitis virus. IV. Transcrip-tion by standard virus in the presence of defective interferingparticles. J. Virol. 9:909-916.
14. Kang, C. Y., and L. Prevec. 1971. Proteins of vesicular stomatitisvirus. III. Intracellular synthesis and extra-cellular appearanceofvirus-specific proteins.Virology 46:678-690.
15. Kiley, M. P., and R. R. Wagner. 1972. Ribonucleic acid species of intracellular nucleocapsids and released virions of vesicular stomatitis virus. J. Virol.
10:244-255.
16. Martinet, C., and C. Printz Ane. 1970. Analyse de la synthese de l'ARN viral du virus de la stomatite vesiculaire(VSV). Utilisation de mutants thermosensi-bles. Ann. Inst. Pasteur Paris 119:411-419.
17. Montagnier, L. 1968. Presence d'un acideribonucleique
double chaine dans la cellule animale. C. R. Acad. Sci. Ser.D267:1417-1419.
18. Mudd, J. A., and D. F. Summers. 1970. Polysomal ribonucleic acidofvesicularstomatitisvirus-infected HeLacells.Virology 42:958-968.
19. Perlman, S.M., andA.S.Huang.1973.Ribonucleic acid
synthesisofvesicular stomatitis virus. V. Interactions between transcription and replication. J. Virol. 12:1395-1400.
20. Printz Ane, C., A. Combard, and C. Martinet. 1972. Study of the transcription and the replication of vesicularstomatitis virusby using temperature-sensi-tive mutants.J. Virol. 10:889-895.
21. Printz, P., and R.R.Wagner.1971.
Temperature-sensi-tive mutantsofvesicular stomatitisvirus:synthesisof virusspecific proteins.J. Virol. 7:651-662.
22. Roy, P., and D. H.L.Bishop.1972.Genomehomologyof vesicular stomatitisvirusanddefectiveTparticlesand evidenceforthesequential transcriptionofthevirion ribonucleic acid. J.Virol. 9:946-955.
23. Roy, P., and D. H. L. Bishop. 1973. Initiation and
4c
on November 10, 2019 by guest
http://jvi.asm.org/
direction of RNA transcription by vesicular stomatitis virus virion transcriptase. J. Virol. 11:487-501. 24. Sokol, F., and H. F. Clark. 1973. Phosphoproteins
struc-tural components of Rhabdoviruses. Virology 52:246-263.
25. Stampfer, M., and D. Baltimore. 1973. Identification of thevesicular stomatitis virus large proteinas aunique viral protein. J. Virol. 11:520-526.
26. Szilagyi, J. F., and C. R. Pringle. 1972. Effect of temperature-sensitive mutations on the
virion-associated RNA transcriptase of vesicular stomatitis virus.J. Mol. Biol. 71:281-291.
27. Szilagyi, J. F., and L. Uryvayev. 1973. Isolation ofan
infectious ribonucleoprotein from vesicular stomatitis virus containinganactiveRNA transcriptase. J. Virol.
11:279-286.
28. Unger, J. T., and M. E. Reichmann. 1973. RNAsynthesis intemperature-sensitivemutantsofvesicular stomati-tisvirus. J. Virol. 12:570-578.
29. Wagner, R. R., T. C. Schnaitman, and R. M. Snyder. 1969. Structural proteins of vesicular stomatitis
vi-ruses.J. Virol.3:395-403.
30. Wertz, G. W., andM. Levine. 1973. RNAsynthesis by vesicularstomatitisvirus anda smallplaquemutant: effects ofcycloheximide. J. Virol. 12:253-264. 31. Wunner,W. M., and C. R. Pringle. 1972. Protein
synthe-sis in BHK-21cells infected with vesicular stomatitis virus. I. ts Mutants of the Indianaserotype.Virology 50:829-840.
on November 10, 2019 by guest
http://jvi.asm.org/



![FIG. 5.periodof39.2withhours39.2disruptedExtractednonpermissivemarker[14C]proteins. 1:10 Electropherograms of virus-specific proteins extracted from CEF infected at a multiplicity of - 50 ts IV100 or ts IViii under permissive, nonpermissive, or upward shif](https://thumb-us.123doks.com/thumbv2/123dok_us/1585382.111184/6.494.38.443.76.335/withhours-disruptedextractednonpermissivemarker-proteins-electropherograms-extracted-multiplicity-permissive-nonpermissive.webp)