JOURNAL OFVIROLOGY,Dec.1967,p.1158-1163
Copyright ( 1967 AmericanSociety forMicrobiology PrintedVol.in1, No.U.S.A.6
Analysis
of
Additional
Interference Occurring
After
the
Removal
of Interferon1
ROYCE Z. LOCKART, JR.
CentralResearchDepartment,ExperimentalStation,E. 1. du PontdeNemoursandCompaniy,
Wilmington,
Delaware 19898Receivedforpublication19June 1967
Cultures of chick cells exposedtointerferoncontinuedtodecreasein virus-produc-ingabilityduring incubationaftertheinterferonwasremoved. Therateof develop-mentofthe additionalinterference and the degree of viral interference finally mani-festedweredependentontheconcentration ofinterferonto which the cultureswere exposedandthe time ofexposure. Additionalinterference occurred also ininfected cells. Additional interference was inhibitedby actinomycin Dand puromycin. The best explanation ofadditional interference is that it results from interferon that is fixedtothe cellsduringtheir initialperiod ofcontact.
Vilcek and Rada (9) demonstrated that cells
exposedtointerferon at 4 C were not resistant if
challenged with virus immediately after the
removalof interferon. The cells became
resistant,
however, if they were incubated at 37 C for
several hours before challenge. More recently,
Levine (4) confirmedtheobservations mentioned
above and showed
additionally
that chick cellswhich had been briefly exposed tointerferon in
the presence of puromycin, and then infected
immediately with Western equine encephalo-myelitis (WEE) virus, produced less virus than
didcontrol cultures which hadnotbeenexposed tointerferon.If,afterthe interferonwasremoved,
the cultures were incubated
additionally
forseveral hours with
medium,
they produced
still less virus after their infection with WEE virus(4). Levine interpreted his data to indicate that the additional reduction of viral
yields
was duetoresidual interferon
activity,
probably
resulting
from cell-fixed interferon.
Theexperimental results reportedhere confirm and extend those
previously reported.
They
sup-port the contentionthat theadditional reductions of viralyields bycellsaftertheir briefexposuretointerferon result from residual bound interferon because both
deoxyribonucleic
acid (DNA)-dependent ribonucleic acid(RNA)
and protein synthesis in the cells arerequired
for the addi-tionalreductions inviralyields.
It is knownthat the antiviral effectresulting
from interferonre-quires cellular RNA and protein synthesis (2, 4, 6, 8).Also,residual interferon
activity
wasshowntotakeplaceininfectedcells.
1Thisworkwasdonewhile the authorwas a mem-ber of thefacultyoftheDepartmentofMicrobiology,
TheUniversityofTexas, Austin.
MATERIALS AND METHODS
Cell cultures, media, and solutions. Chick embryo cells were prepared from embryos 10 days of age. The cells weresuspended inEagle'smedium (1) containing 3% calf serum (EC medium) and distributed into 60-mm petri dishes. Cultures were used the day after their preparation, at which time each culture con-tained approximately 3 million cells. Phosphate-buffered saline containing bovine serumalbumin at a concentration of0.1% (PBSA) was used to wash the cell cultures and served as a diluent for viruses. Ac-tinomycin D was provided by Merck, Sharp and Dohme, West Point, Pa. It was dissolved in95%ethyl alcohol and further diluted in EC medium for use. Puromycindihydrochloride was purchased from Nu-tritionalBiochemicals Corp., Cleveland, Ohio.
Virus. The preparation of WEE virus stocks, their storage, and the assay method for virus titrations were previously described (5).
Interferoln.Thepreparation of chick interferon was described previously (6). Fluids containing nonpuri-fiedinterferon were used. For potencymeasurements,
thefluids werediluted in twofold steps in EC medium, and 2 ml was added to chick embryo cultures. The greatestdilution capable of preventing the appearance
ofcytopathological effects in the cultures when they were infected with a large multiplicity [>10
plaque-formingunits (PFU) per cell] of WEE virus 17 to 24 hrlater wasconsidered tocontain one protective unit ofinterferon.
RESULTS
Kineticsofinterferon action. The rate at which culturesof CEcellsexposed to interferon at 37 C losttheir ability toproduce virus varied directly with the concentration of interferon to which they wereexposed. This dependencyis illustrated in Fig. 1. The ratedifferences shown are readily reproducible and may serve as a sensitive way to 1158
on November 11, 2019 by guest
http://jvi.asm.org/
3
HOURS WITH INTERFERON
FIG. 1. Kineticsof interference resulting from
inter-feron. Cultures ofchickembryocellswereincubatedfor
1,3, and5hrat37Cwith2, 8,and 32 protective units
of interferon. At each interval of time, cultures were washed andinfectedby the addition ofmore than 10
PFUpercellofWEE virus. Virus yields ofthecultures
wereplottedrelative to thosefromcultures which had
receivednointerferon.
measure relative interferon concentrations. In the experiment illustrated, replicate cultures of
chickembryocells wereincubated with interferon
diluted in EC medium. Each culture received 2
ml. At the times indicated, each culture was
washed three times with3 ml ofPBSA,carebeing
taken to remove all fluid between washes. Each
culture was then infected by the addition of at least10PFUpercell ofWEEvirus. Thetitersof
virusin thefluidsremoved 16 hr postinoculation were determined and areplotted in Fig. 1
rela-tive to the titers found in control fluids. Cells
generally produced between 1,000and3,000PFU
per cell. Therefore, itcanbeseenthatcells were
rendered able to produce an average of only a
few PFU when incubated for 3 to 5 hrwith 32
units of interferon, but produced considerably morewhen incubatedwith less interferon.
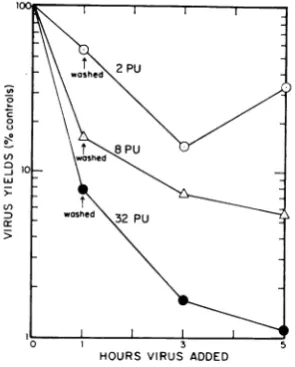
Additional interference after removal of
inter-feron. Replicate cultures of chick embryo cells were exposed to2, 8, and 32 units of interferon
for1 hrat37 C. Theinterferonwasremoved,and
the cultures were washed four times with warm PBSA. Two cultures exposedto each concentra-tion of interferonwereimmediatelyinfectedbythe
additionof >10PFUpercell of WEEvirus. Two
ml of warm EC medium was added to the re-mainder of the cultures. After 2 and 4 hr of additional incubation at 37 C in medium only, two cultures which had been exposed to each
concentration of interferon for the initial hour were removed and infected. The concentrations of virus in the fluids of the cultures were
deter-mined 20 hr after theirinfection.The viral yields obtained from the cultures exposed to interferon
were compared with those from the control cultures (Fig. 2). Acomparison ofthe viralyields
relativeto controlsafter1 hrof incubationinthis
experiment with those shown inFig. 1 reveals a closecorrespondenceofthedegreeofinterference
in the two experiments, thus illustrating the
reproducibility ofthe technique. The data show
that cellcultures incubated withinterferon for 1 hrcontinuedtodecline in their abilitytoproduce
virus,evenafter the interferonwasremoved. This
continued loss ofability toproduce virus will be
referred to as "additional
interference"
through-out the remainder of this paper. The additional interference was most pronounced in the 2 hrafter removal ofthe interferon and was almost
completeby4hrafter its removal. Themaximal degree to which the cultures were inhibited in
theirabilitytoproduce viruswasdependentonthe
concentration of interferon to which they were exposed. Note the ability of cells exposed to 2 units of interferon to produce more virus when
they were incubated longer than 2 hr after the removal oftheinterferon. The rapid recovery of
virus-producing ability after a briefexposure to
o
wshed3 PU
0
hd
0 3 5
HOURS VIRUS ADDED
FIG. 2. Demonstration of additional interference.
Culturesofchickembryo cellswereincubated with2, 8!
and32protectiveunitsofinterferonforIhrat37C.All cultures were washed,andsome wereinfected
immedi-ately.Otherswereincubated2and 4hrlongerinmedium
at37 Cpriortobeinginfected. Yieldsofvirusobtained 20 hrafter infection areplottedrelative to the yields
from cultures which hadnotbeenexposedtointerferon.
on November 11, 2019 by guest
http://jvi.asm.org/
[image:2.471.38.231.49.264.2] [image:2.471.264.412.388.572.2]LOCKART
uI
-I
n,
a:)
>r
TIME(hrs.)VIRUS ADDED
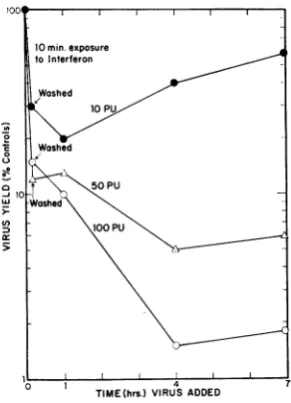
FIG. 3. Additional interferenceafter10minof
incu-bation withinterferon. Theexperimentwasthesameas
that illustratedin Fig. 2,except thathigher
concentra-tionsof interferonwereused and theperiod ofexposure
ofthe cellstointerferonwas10 min.
small amounts of interferon and further
incuba-tion hasbeen noticedrepeatedly.
Figure 3 illustrates anotherexperiment of the
samenature,but which differed from thatshown inFig. 2in that the concentrations ofinterferon
were higher and the period ofexposure was
re-ducedto10 min.Again,those culturesexposedto
to the two highest concentrations of interferon attained the maximal degree of inhibition when
they were incubated at 37 C withmedium for 3 hr and 50 min after the removal of interferon. The degree to which virus production was
in-hibitedat thattime again depended onthe
con-centration ofinterferon to which they had been
exposed. Also, as before, the cultures incubated
with the least concentration of interferon (10 protective units) showed an initial decrease in
their abilitytoproduce virus and thenagradual
increase in this ability. The time of initialcontact with interferon can be even shorter. Cells
in-cubated with 500 units of interferon for only 1 min were reduced in their ability to produce virusby 92.4 to 96.7% in threeseparate
experi-ments after an additional 4 hr of incubation in
warmmedium which contained nointerferon. Evidence that the additional interference was cell-fixed. The experiments just reported do not permitonetodecidebetween the alternatives that the additional interference was due to (i) cell-fixed interferonor(ii) the release into the medium
ofa small amount of reversibly adsorbed inter-feron. Therefore, the following experiment was performed. Six CE cultures were incubated with 16units of interferon for 1 hrat37C and subse-quently treated in the following three ways. (A) Twocultureswerewashed 12 times and immedi-ately infected withmore than 10 PFUpercell of
WEEvirus. (B) Twocultures were washed three
times after the removal of interferon and three times after each hour of incubation at 37 C in medium until they had received 12 washes. The cultures were then infected with virus. They had
been incubated with interferon for 1 hr and medium for 3 more hours after the removal of interferon. (C) Two cultures were washed 12
times after the removal ofinterferon, incubated additionally for 3 hr at 37 C, and infected with virus. Two cultures which hadreceived no
inter-feron were included in each of the scheduled
treatments (A to C) outlined above. Virus yields
ofthe cultures were determined 20 hr after they were infected. The results are shown in Table 1.
[image:3.471.63.209.73.275.2]Additional interference occurred during the 3-hr period of incubation andto verynearly thesame
TABLE 1. Cell-fixednatureof"additional" interference
Virus titer(PFU/ml)
Viralyields(per Treatment No previousincubation 1-hrincubation with16 cent of control)
withinterferon units of interferon
Expt1 Expt2 Expt1 Expt 2 Expt1 Expt 2
(A) Washed12 times, then infected
immediately... 1.0 X 109 7.7 X 108 5.7 X 107 5.4 X 107 5.7 7.0 (B) Washed three times
immedi-ately; three times after each of threehourly periods of
in-cubation; infected... 1.3 X 109 1.6 X 109 1.5 X 107 5.9 X 106 1.2 0.4
(C) Washed 12times; incubated 3
hr; infected... 1.7 X 109 1.3 X 109 1.3 X 107 6.3 X 106 0.8 0.5
1160 J. VIROL.
on November 11, 2019 by guest
http://jvi.asm.org/
[image:3.471.46.436.499.648.2]OCCURRING INTERFERON
O . Adhed
intE cutue ploteas clse cices an desinate
C ~~~3~ 5
HOURS
FIG. 4. Inhibition ofadditional interference by
ac-tiinomycinD. Additionalinterference wasdemonstrated
in the culturesplottedasclosed circles anddesignated
VA. Those cultures had been incubated withinterferon
for 1 hr andwere infectedwith WEE viruscontaining actinomycinD inthe inoculum eitherimmediatelyafter
the removalofinterferonorafter2 and 4 hrofadditionlal incubation with medium. The closed circle with a V
indicates theyield ofvirusfrom cultures incubatedfor
1 hrwithinterferon andinfected immediately afterits removalwith viruswhoseinoculum didnotcontain
ac-tinomycinD. Theprevention ofadditionalinterference
byactinomycinisshownby thepoints designatedA V. Afteraninitial hourofincubation withinterferon,itwas
removed and these cultureswere incubatedadditionally
with medium containing actinomycinD (I pg/ml). Cell
cultureswhich hadnotbeenexposedtointerferonwere incubatedfor2 and 4 hr withactinomycinDtoserveas
controls. Theyare represented by the open circles fol-lowedbyV.
extent,
regardless
of whether the cultures werewashed
12 timesinitially
or 3 times each houruntil they had been washed 12 times. It seems
unlikely thatthe additional interference was due
to
reversibly
adsorbed interferon.Inhibition of the additional interference by
actinomycin D. The effect of
actinomycin
D on the additional interferencejust
described wasdetermined. Two cultures were infected with WEE virus, and the
yield
of virus 20 hr later served as thecontrol value. Other cultureswereinfected after incubation with
actinomycin
D(1,ug/ml)for2and4 hr.The viral
yields
fromthe cultures incubated withactinomycin
served as a measure of the loss ofvirus-producing
capacity
bythecellsresulting
fromthedrug
(open
circles,
Fig. 4). Other cultures were incubated with 16
units ofinterferon for 1 hr. Four cultures were washed and infected
immediately.
Two of the cultures containedactinomycin
in the virusinoculum
(VA
inFig. 4);
two did not(V
inFig. 4). Thosecultures which had been incubated
withinterferon for 1 hr and which were infected
immediately with a virus inoculum containing
actinomycin produced only about 50% as much
virusasthe controls (whichalso were challenged with actinomycin in the virus inoculum). Those cultures incubated with interferon for 1 hr and
infected immediately with a virus inoculum
con-taining no actinomycin produced only 11% as much virus.Therefore, cultures of cells incubated
with interferon for 1 hr produced one-ninth as much virus when infected with virus alone, but virus production was reduced by only
one-half when actinomycin D was present in the
inoculum. The discrepancy in virus yields indi-cated that additional interference probably occurred after infection. Thispoint will be dealt with later. Four cultures which had been incu-bated with interferon for 1 hr were washed and incubated further at 37 Cwith medium. After 2 hr, andagain4 hr later,two of thecultures were
removed, washed, and infected with a virus inoculumcontainingactinomycinD. The yields of virus 20 hr after infection were determined.
Whereas the cultures which were infected with
virus in the presence of actinomycinimmediately after the removal of interferon produced 50% asmuch virus ascontrols, those culturesincubated
an additional 2 and 4 hrprior to infection
pro-duced 11 and 7.3%, respectively, as much virus asthe controls. Thisdemonstrationofadditional
0
-n
0
z 0
0 0
C-0 0~
C,)
5;
0 3 5
HOURS
FIG. 5. Inhibition ofadditionalinterference by puro-mycin. AfterI hrofincubation with 32unitsof
inter-feron, cultures wereincubatedadditionally foreither2 or4 hrwith mediumlacking (0) andcontaining (0)
puromycinat
50,Sg/ml.
Atthetimeperiods designated,they were infected with virus. The virus inocula
con-tained actinomycinD (5ug/ml).Theclosed circlewith
a Vindicates theyield ofvirusfrom cultures incubated
forIhr with interferonandinfectedimmediately after itsremoval with virus whose inoculum didnotcontain
actinomycinD.
on November 11, 2019 by guest
http://jvi.asm.org/
[image:4.471.60.206.70.243.2] [image:4.471.242.435.392.543.2]LOCKART
interference is shown in Fig. 4 (closed circles labeledVA).Fourothercultures which had been
incubated with interferon for 1 hr were washed and incubatedfurtherat37 Cwithmedium con-tainingactinomycin D (1 ,ug/ml) and 32 units of interferon.Theinterferonwasadded to show that the actinomycin D was working. Two cultures
were removed after 2 hr of
incubation,
and the finaltwocultures wereincubated for4hr. At thetime oftheir
removal,
they were washed and in-fected. The amounts of virus produced by the cultures at 20 hr after infection were deter-mined and are represented in Fig. 4bythe opentriangles. The action of the 32 units of added interferon and any additional interference were
preventedbythepresence ofthe actinomycin D. The slight decline of virus production found in
those cultures was mostprobablydue tothe loss
of
virus-producing
abilityresulting
from the presence of the actinomycin, as the yields of virus from control cultures(open circles, Fig.
4) decreased at the same rate when incubated
withactinomycin.
Inhibition of the additional interference by
puromycin.Theproceduresfollowedtodetermine
the effect ofpuromycinonadditionalinterference
were the same as described for the
experiments
in which
actinomycin
was used.However,
thecultureswereincubated with 32 units ofinterferon rather than 16, and noadditionalinterferon was
added during the additional incubation in the presence of
puromycin.
Puromycin
was usedatafinal concentration of 50
,g/ml.
Also,
all thosecultures incubated for additional
periods
afterthe removal of interferon were infected with virusinocula
containing
actinomycin
D(1 ,ug/ml).
As shownin
Fig.
5,puromycin,
likeactinomycin,
prevented the additional interference which
oc-curred in the presence of medium alone when cultures were incubated for 2 and 4 hr after the
removal ofinterferon.
Occurrence
of
additionalinterference
ininfected
cells.After brief
periods
of exposuretointerferon,
virusyieldswerenearly
10-foldgreaterwhenacti-nomycin D was present in the virus inoculum
(Fig.4and6)oraddedjust
prior
toinfection (6).Inlight ofthe
foregoing
results,themostprobable explanation is that the reducedyields
of virusproduced when the cultures are infected with virus alone area result, atleastin part, of addi-tional interference
taking
place in the infectedcells.
Actinomycin
Dwas usedtoshowthat this didoccur. Culturesofcellswere incubated for 1 hrwith 16 units ofinterferon.Theinterferonwasremoved and the cultureswerewashedwithwarm
PBSA. Two cultures (closed circle, Fig. 6) were
infected
immediately
with virus and two (opencircle,
VA, Fig.6)
with virus inaninoculumcon-0
D VA
0L
0 6ashe, Infect
Act.
v ~~~~~~~~Act.Act
-1 0 3 6 9 12
Time of addition of ACTINOMYCIN(Hrs. Post-infection)
FIG. 6.Additional interference in infected cells.
Cul-turesof CE cellswereincubated with16 unitsof
inter-feron forI hr, washed, and infected. To the medium, actinomycinD wasaddedtoaconcentration ofI,ug/ml
atthe timesindicated. The closed circle witha V
indi-catesthe yield ofvirusfrom cultures incubated forIhr
with interferon and infected immediately after its re-movalwithviruswhoseinoculum did not contain actino-mycin D. Viral titers were determined 20 hr after in-fection.
taining
actinomycin
D (5 ,ug/ml). An additional eight cultures which hadbeenincubated with in-terferon for 1 hr were washedandinfected,
andmediumwasadded to each.At2, 5, 8, and 11 hr afterinfection, sufficient medium with
actinomy-cinwasaddedsothatitwaspresentat a concen-trationof 1 ,ug/ml. The concentration ofvirus in thefluidsremoved 20 hrafterinfectionwas deter-minedforallthecultures. The results are plotted in Fig. 6 (open circles, Act.). Additional
inter-ference ininfected cells accounted for the
differ-ence in virusyields found between those cultures
whichwereincubatedwithinterferonfor 1 hr and
infected, withor withoutactinomycin. The addi-tional interference was reflected only gradually andwasinhibitedby the addition of actinomycin
totheinfectedcultures at time intervals prior to 9hrafter infection.
DISCUSSION
Thedata inFig.2and 3 show that cellcultures which have beenincubated with interferon only
briefly, sothat only a small amount of virus
in-hibition had occurred, continued to decrease in their abilitytoproduce virus whenincubated for
1162 J. VIROL.
on November 11, 2019 by guest
http://jvi.asm.org/
[image:5.471.255.448.66.291.2]additional periods of time. Additional
interfer-ence was described previously by Levine (4).
Several otheraspects of the additionalinterference
became apparent in the present work. The rates at which additional interference occurred were de-pendent on the concentrations of interferon to which the cultures were exposed, but were less than those observed if theinterferon was left on the cultures (compare Fig. 2 with Fig. 1). The final amounts by which virus yields were decreased were alsodependent on the concentra-tion of interferon to which the cultures were
initially exposed. Those cultures exposedto low
concentrations (Fig. 2), or morebriefly (Fig. 3),
were decreased in their virus-producing ability
upon additionalincubation, butregainedsomeof thatabilityon further incubation. Itwas demon-strated that the continued interference was
cell-fixedbyshowingthat12washes,whether applied
immediately upon the removal of interferon or intermittently fora 3-hrperiodoftime, failedto
affectsignificantlythe amount ofadditional ference that occurred after the removal of inter-feron. Having established that the additional interference was cell-fixed, several
possibilities
were considered. Additional interference
might
have resulted from (i) cell-fixed
interferon,
(ii)
messenger RNAwhich had accumulated during
theinitialperiodofexposuretointerferon, or
(iii)
some other process set in motion
by
the initial exposure to interferon. As the additionalinter-ference was stopped by both actinomycin and
puromycin, it almostcertainlywasnotaresultof anearlyaccumulationof interferon-induced
mes-senger RNA. And, by being sensitive to both
drugs, the additional interference reacts like the interference which occurs in the continuing pres-ence ofinterferon.
Finally, itwas
interesting
to seethatadditional interference occurred eventhough the cells wereinfected. Such a
finding
is consistent with the demonstration that interferon can inhibit arbo-viruses when added after infection(7, 10),
and it suggests thatWEE virus has very little effect oncellularRNA and
protein synthesis.
Otherexperi-ments
(Sreevalsan, personal
communication)
haveindicatedthat infection of chick cells with WEE
virus
only
slowly
inhibits cellular RNAand pro-teinsynthesis,with50%
inhibitionbeing
apparent at about 8 hrpostinoculation.
That additionalinterference occurs in infectedcells also
explains
why
approximately
10 times more virus ispro-ducedwhen cultures which have been incubated withinterferonareincubated with
actinomycin
Dat the time of or before their inoculation with WEEvirus (6).Thislarge difference in viral yields due to the presence ofactinomycinoccurred, how-ever, only duringrelatively short periods of incu-bation with interferon when the cultures were
infected priorto thetime that theinterferon pres-ent had exerted its maximalinhibitoryeffect.
Although the data presented still fall short of proving that interferon is taken up by the cell, they are most consistent with that explanation. Further, they indicatethat, once taken up, inter-feron can continue its induceractivity for several hours,eventhough the cellsareinfected.
ACKNOWLEDGMENT
Iwish toacknowledge the excellent technical assist-ance of Barbara Horn.
Thisinvestigation was supported byPublic Health Service grantsAI-03538 and5-K3-AI-19,385 from the National Institute of Allergy and Infectious Diseases.
LITERATURE CITED
1. EAGLE, H. 1955. Nutrition needs of mammalian
cells intissueculture. Science 122:501-504.
2. FRIEDMAN, R. M., AND J. A. SONNABEND. 1964. Inhibition of interferon action by
p-fluoro-phenylalanine.Nature 203:366-367.
3. LEVINE, S. 1964. Effect of actinomycin D and
puromycindihydrochloride on action of inter-feron. Virology 24:586-588.
4. LEVINE, S. 1966.Persistence of active interferon in cells washed after treatment with interferon. Proc.Soc.Exptl.Biol. Med. 121:1041-1045. 5. LOCKART,R. Z. 1963. Production of an interferon
byLcells infected with Western equine
enceph-alomyelitisvirus. J. Bacteriol. 85:556-566. 6. LOCKART, R. Z. 1964. Thenecessity for cellular
RNA andprotein synthesis for viral inhibition
resulting from interferon. Biochem. Biophys. Res. Commun.15:513-518.
7. LOCKART, R. Z., AND T. SREEVALSAN. 1963. The effect of interferon on the synthesis of viral nucleic acid. Viruses,Nucleic Acids, and Can-cer, 17th AnnualSymposium on Fundamental Cancer Research, University of Texas M. D. Anderson Hospital and Tumor Institute, Houston, p.447.The Williams and Wilkins Co., Baltimore.
8. TAYLOR, J. 1964. Inhibition of interferon action byactinomycin. Biochem. Biophys. Res.
Com-mun. 14:447-451.
9. VILCEK, J.,ANDB. RADA. 1962. Studies onan in-terferon from tick-borne encephalitis virus-infected cells (IF). III. Antiviral action of IF. Acta Virol. 6:9-16.
10. WAGNER, R. R. 1961. Biologicalstudies of
inter-feron. I. Suppression of cellularinfection with Eastern equine encephalomyelitis virus. Virol-ogy 13:323-337.