Copyright © 1971 American Society for Microbiology Printed in U.S.A.
Deoxyribonucleic Acid Replication in Simian
Virus
40-Infected Cells
IV. Two Different Requirements for Protein Synthesis During Simian
Virus
40
Deoxyribonucleic Acid Replication
H. S. KANG, T. B. ESHBACH, D. A. WHITE, AND A. J. LEVINE Department of Biochemistry, PrincetonUniversity, Princeton, New Jersey 08540
Received forpublication 8 September 1970
The replication of simian virus 40 (SV40) deoxyribonucleic acid (DNA) was inhibited by 99% 2 hr after the addition of cycloheximide to SV40-infected primary
Africangreenmonkey kidney cells. The levels of 25S (replicating) and 21S (mature) SV40 DNAsynthesizedafter cycloheximide treatment were always lower than those
observedinaninfecteduntreated control culture. This is consistent with a require-mentfor aprotein(s) or for protein synthesis at the initiation step in SV40 DNA
replication. Therelativeproportion of 25S DNA as compared with215 viral DNA
increased with increasing time after cycloheximide treatment. Removal of cyclo-heximide from inhibited cultures allowed the recovery of viral DNA synthesis to
normallevelswithin 3 hr. During the recovery period, the ratio of 25S DNA to 21S DNA was 10 timeshigher than that observed after a 30-min pulse with3H-thymidine
with an infected untreated control culture. The accumulation of 25S replicating SV40 DNA during cycloheximide inhibition or shortly after its removal is
inter-pretedto meanthat aprotein(s) orprotein synthesis is required to convert the 25S
replicatingDNAto21Smatureviral DNA. Further evidence of a requirement for proteinsynthesisinthe 25Sto 21S conversion was obtained by comparing the rate of this conversion ingrowingandrestingcells. Theconversion of25S DNA to 21S DNAtookplaceat afaster rate in infected growingcells than in infected confluent
monolayer cultures. A temperature-sensitive SV40 coat protein mutation
(large-plaqueSV40) had no effect on the replicationof SV40 DNA at the nonpermissive temperature.
Infection of primary African green monkey kidney(AGMK) cellswith simian virus 40(SV40) results in the synthesis of viral deoxyribonucleic
acid
(DNA)
and infectious virus (10,12).
SV40DNAreplication begins at about 15 hr after in-fection and reachesa maximum rate atabout 30 hr. When infected cells were
pulse-labeled
with 3H-thymidine for shortperiodsof time (less than tO min), littleor no mature viral DNA was de-tected eitherby neutraloralkalinesucrose gradi-ents or by cesium chloride-ethidium bromideequilibrium centrifugation. Instead,a viral DNA form that sedimented in neutralsucrose
gradients
at25S was observed. This DNAwas
completely
denaturable in alkali and banded at a lighter densitythanclosed circular matureviralDNA in ethidium bromide-cesium chloride equilibrium
gradients. The 3H-labeled 25S DNA could be chased into 21S mature viral DNA when
un-labeled thymidine was added to the culture medium (12).
Electron micrographs of 25S DNA showed
SV40replicating circles (two branchpoints, three
branches, and no visible ends) where90%O of the SV40 moleculehad completedits replication. At any one time, most of the replicating molecules
observed in the infected cell were of this 25S replicating type (75 % of thereplicating molecules were90% completed). This ledto thesuggestion
that therewas a sloworrate-limiting step late in the replication of an SV40 DNA molecule and thatthisresulted inanaccumulation of 255 DNA
(12).
In thepresentstudy,it is shownthat
cyclohexi-mide
(10
lg/ml) inhibits thereplicationof SV40 DNA. Thesedataarein agreementwith previousstudies with SV40 (11) and polyoma virus
(5).
The results indicate that protein synthesis is
re-quiredat twodifferent stages in the
replication
of SV40 DNA: (i) theinitiation ofDNAsynthesis,
and(ii)theconversion ofthe25Sreplicatingform to 215matureviralDNA
(finishing
step).
112on November 11, 2019 by guest
http://jvi.asm.org/
MATERIALS AND METHODS
Virus. The SV40 large-plaque mutant (16, 17, 19, 20) and wild-type strain (13) wereemployed in thesestudies.
Tissue culture. AGMK cells (Flow Laboratories, Inc.) were cultured in either plastic petri dishes (Falcon, 100 by 20 mm) or plasticflasks (Falcon,75 ml) in Dulbecco's modified Eagle's medium (Grand Island Biological Co.) supplemented with 10% calf
serum.
Infectivity assay. Virus stocks were prepared as describedpreviously (12). Infectiousviruswastitrated
onmonolayer cultures of AGMKor BSC-1 cellsby
useofaplaqueassayprocedure (6).
Measurement of the rate of DNA synthesis. The
rate ofviral DNA synthesis was measured by the addition of3H-thymidine (14 to 18 Ci/mmole, New England Nuclear Corp.) to infected cell cultures. After an appropriate exposure to the isotope, the cells were washed twice with ice-cold phosphate-buffered saline(PBS,0.01 Msodiumphosphatebuffer
at pH 7.2 and 0.15 M NaCl) and lysed with 0.6% sodium dodecyl sulfate (SDS) in 0.01 M sodium phosphate buffer, pH 7.2, 0.15 M NaCl, and 0.01 M disodium ethylenediaminetetraacetic acid (EDTA). Viral DNA wasseparatedfromcellularDNAbythe Hirt procedure (9). A portion of the 1 M NaCl-SDS soluble fraction (small-molecular-weight DNA) was
sedimentedthrougha5 to20%linearsucrosegradient for 3 hrat40,000rev/mininanSW 50.1 rotor.The 3H-labeled component that sedimented at 21S was
used to quantitate the levels of mature viral DNA synthesis. The quantity of 3H-labeled DNA that sedimented at 25S was employed to determine the
amounts ofreplicating SV40 DNA madeduring the
labeling period (12). The quantity of3Hcounts per minute in the 1 M NaCl-SDS precipitable fraction (large-molecular-weight DNA) was employed to
determine the rate of cellular DNA synthesis. Only the fraction ofthe 3H-label that was trichloroacetic acid-precipitableandresistanttoalkalinedegradation (0.3 MKOH,37 Cfor 18hr)wasusedto measurethe
rate of cellular DNA synthesis.
After a 1-hr exposure ofSV40-infected cells with 3H-thymidine, over 90% of the trichloroacetic acid-precipitable radioactive label in the 1 M NaCl-SDS soluble fraction is virus-specific (12). Greater than 90% ofthe 3H-labeled DNA in the 1 M NaCl-SDS precipitable fractioniscellularDNA(12).
Benzoylated-naphoylated diethylaminoethyl cellu-lose(BNC)chromatography. BNCwassynthesizedby the procedure of Gillam et al. (7) as modified by Sedat, Lyon, and Sinsheimer (18). BNC columns were prepared and utilized as described previously (12).
Centrifugationtechniquesandisotopemeasurements.
Sucrose density centrifugation was performed as
previously described (12). The details of each sedi-mentation run aregivenin thefigurelegends. Radio-active samples were prepared and counted in a
Beckman liquid scintillation counter as described previously (12).
Cycloheximide treatment. Cycloheximide was
obtained fromSigmaChemical Co. At the
concentra-tion employed (10 ug/ml), cycloheximide inhibited the incorporation of 3H-leucine (30 to 50 Ci/mmole, NewEngland Nuclear Corp.) into hot trichloroacetic acid-precipitable (100 Cfor45min) materialby96% during the first hour and 99% by the second hour of treatment. Infected and uninfected AGMK cells behaved in an identical manner.
RESULTS
Effectof cycloheximide onSV40 DNA replica-tion. To examine the effects ofcycloheximide on thereplication of SV40 DNA, thefollowing experi-ment was performed. Six monolayer cultures of AGMK cells were infected with SV40 large-plaque mutant, and 30 hr later five of these cultures were treated with 10,ug of cycloheximide per ml. One cycloheximide-treated culture and one untreated control culture were then labeled with 3H-thymidine (1,uCi/ml) for 1 hr at 36 C. A second cycloheximide-treated infected culture was labeled from 1 to 2 hr after theaddition of theantibiotic (31 to 32 hr after infection). Each additional culture was labeled in an identical manner during the 3rd, 4th, or 5th hr after exposure tocycloheximide.At theend of the 1-hr
pulse-labeling period, each culture was washed two times with ice-cold PBS, and the cells were lysed with0.6% SDS as described in the preceding
section. A 0.2-ml sample ofthe 1 M NaCl-SDS
soluble fraction was sedimented through a 5 to 20%linearsucrose gradient for 3 hr at 40,000
rev/
min. Theresults of this experiment are presented inFig. 1.
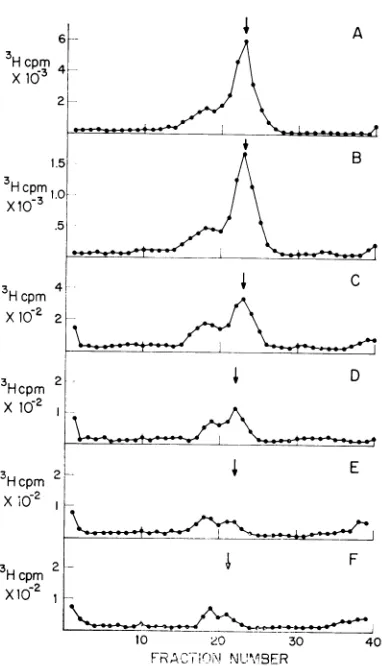
Theadditionofcycloheximidetovirus-infected
AGMK cells results in the cessation of SV40
DNAreplication. The synthesis ofSV40-specific
DNAwas inhibited by 99 % after 2 hr of exposure to this antibiotic. Cycloheximide inhibited the
production of both 25S (replicating) DNA and 21S (mature) SV40 DNA. However, after an initial 1-hr delay, the ratio of 25S to 21S DNA was observed to increase from 0.3 to 1.0. This
can be seen in the sucrose gradientprofile pre-sented inFig. 1 (compare 1A with 1E or IF).
Levine etal. (12) demonstrated that 25S viral DNAis aprecursor of 21S mature SV40 DNA. Theincreased levels of 25S DNA relative to 21S DNAobservedaftercycloheximidetreatment
sug-gestedthepossibility thatcycloheximidemay act
byblocking the conversion of 25S to 21S DNA
(as wellasinhibiting the initiation ofSV40DNA
synthesis). Because ofdifficulties in quantitating
therelativeamounts of 25S and 21S DNA when sucrose gradient centrifugation was used, a secondprocedure was also employed to separate these two forms ofSV40DNA. BNC chromatog-raphyhas been shown to fractionate replicating
(25S) and mature SV40 DNA (12). The
on November 11, 2019 by guest
http://jvi.asm.org/
KANG ET AL.
X10
Hcpm
Hcpm
X10-2
3H
cpm D3Hcpm2
X -2
xI
10 20 30 40
FRA1 N UMB R
FIG. 1. Sedimentation ofSV40 DNA synthesized after cycloheximide inhibitioii. Sixmontolayercultures
ofAGMK cells were iitected with SV40, and30 hr
laterfive ofthe cultures received10jig of
cyclohexi-mide per ml. One cycloheximide-treated culture (B)
andone untreatedculture (A) were labeled with
3H-thymidine (1 ,uCi/ml) for1hr.Theotherclulturzeswere
labeled (C) from I to 2hr, (D) from 2 to 3 hr, (E)
from3to4hr,anid(F) from4to5 h-after additionof
the drufg. A sample of the I M NaCI-SDS soluble fraction was sedimented for 3 hr at 40,000 rev/mim
in an SW50.1 rotor. (*) 3H collultspe- miniute; the
arrow represents thepeak of a 21S 92P-labeled SV40
DNA sedimentationi mnar-ker.
periment presentedin Fig. 1 wasrepeated except
that the 1 M NaCl-SDS soluble fraction was
phenol-extracted and chromatographed on BNC columns. Mature viral DNA (21S) eluted from the BNC resin with1 MNaCl. The 25SDNA
re-mained boundto the column under these condi-tionsandwaseluted ata later time with 2% caf-feine in 1 M NaCl. This procedure permit<> an
excellent separation of replicating and matui-e
[image:3.491.63.254.70.403.2]viral DNA (12).
TABLE 1. Inhibition of S V40 DNA synthesis by cycloheximide
Sucrose gradient BNCb Cyclohex-iiodeX |Labeling
iie timePecetPrcn
concn Per cent of Percent Per cent of Percent control DN2A control DNA2
,.g, nl hr
0 0-1 lOOc 24 lO0c 27
10 0-1 24 22 31 23
10 1-2 7.5 37 9.3 33
10 2-3 1.0 43 1.8 41
10 3-4 0.9 50 1.0 48
10 4-5 0.6 54 1.7 53
aPercentage of total DNA synthesized during
thetimeinterval indicated that sedimented at255
or was eluted with 26/a caffeine from a BNC
column.
bBenzoylated-naphoylated diethylaminoethyl cellulose.
,The100%value for the 3Hcountsperminuteas
measured by sucrose gradients is 29,000 counts/ minandby BNC is215,000counts/min.
Table 1 presents theresults of experiments de-signedto investigate theeffects of cycloheximide
onSV40DNA replication. Boththerateof viral DNA synthesis and the relative proportions of 25S and21S SV40 DNAweremeasured byuseof
BNC chromatography and sucrose gradient analysis. Within the first hour after the addition ofcycloheximide, viral DNAsynthesis decreased
to25to30%3oof thelevel observedin theuntreated control culture. During this time, the relative
amountsof25S and 21SDNAremainedthesame
in the antibiotic-treated and untreated cultures. After a 1-hr exposure to this drug, the rate of viralDNAsynthesis continuedto fall(from 10to
1 %), but an increasing proportion of the SV40
DNA remained in the replicating form (25S or
caffeine-eluted DNA). The percentage of 25S DNA synthesized in a 1-hr pulse-labeling period increased from 25cO in the absence of cyclohexi-mideto50c%Oby3to4 hr after the addition of this drug. The sameresults wereobtained with either sucrose gradient analysis or BNC chromatog-raphy.
Recovery ofSV40DNAsynthesis after the
re-movalof cycloheximide.Theexperimentsdescribed in the preceding section suggest that cyclohexi-mide might be inhibiting both the initiation of viral DNA synthesis and the conversion of 25S
DNA to mature viral 21S DNA. It is difficult
however, to make any conclusions about the
significance of the relative proportions of 255
and 21S DNA when the rate of viral DNA synthesis is only 1% of the normal levels. For
thisreason,itwasnecessarytoinvestigatewhether
114
J. VIROL.on November 11, 2019 by guest
http://jvi.asm.org/
[image:3.491.263.461.87.216.2]viral DNA replication could fully recover from
this cycloheximide treatment and to determine
whathappened during thisrecoveryperiodtothe conversion of 255 to 21S DNA.
Cycloheximide (10,g/ml) was added to five
AGMK monolayer culturesat 36 hrafter
infec-tion. After 3 hr,the cycloheximide was removed
fromfour of thesecultures by washing the cells four times with 10 ml ofwarm (37 C) medium.
Viral DNA synthesis was measured by
pulse-labeling these cultures with 3H-thymidine (1 ,uCi/ ml) for 30 min at different time intervals after the
removal of cycloheximide. The pulse-labeling
periods were stopped by washing the cultures
with ice-cold PBS and the cells were lysedwith
0.6%
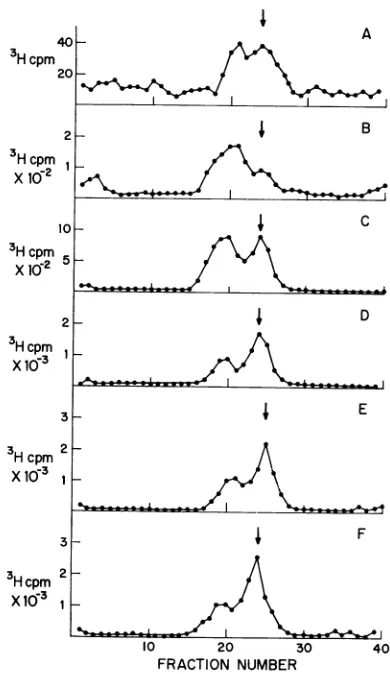
SDS. A sample of the 1 M NaCl-SDS soluble fractionwasanalyzed bysucrosegradient centrifugationaspreviously described. Theradio-active sedimentation profiles from these sucrose gradients are presented in Fig. 2. Figure 2Awas
obtained from an infected culture where cyclo-heximidewas notremoved. Figures2B, C,D,and E were obtained by pulse-labeling with 3H-thymidine at 0 to 0.5hr, 0.5 to 1 hr, 1.5 to 2hr, and 3to3.5 hr after the removal of
cycloheximide.
Figure 2F was obtained from a control culture
that neverreceivedcycloheximide. Itcanbeseen thatremoval of thisdrug allowsafull recovery of
therate of viralDNA synthesis. The viral DNA
synthesized
shortly
after the removal ofcyclo-heximide (0to 0.5hrand0.5to1
hr)
wasmostly
25Sreplicating DNA. Withincreasing
timeafterreversal ofcycloheximide
inhibition,
the ratio of25Sto21SDNA
changed
from 3.0to0.3.Table2presents theresults ofexperimentsthat
demonstrate the reversal of
cycloheximide
in-hibition of viral DNA synthesis. Both BNC chromatography and sucrose gradient
analysis
were employed to investigate the rate of viral
DNAreplication andthe relativeproportions of
25S and21SviralDNA. After 3 hrofexposureof infected cells to
cycloheximide,
the rate of viralDNA synthesis was 1%ofthatin anuninfected control culture. By 3.5 hr after the removal of this drug, the rate ofviral DNA
synthesis
was normal (87 to 100% ofthe control value), and the relativeamounts of25S and 21S DNA wereidenticaltothoseintheuntreated infected control culture(30to35% of 25SDNA).Between0and 2 hrafter removal ofthisdrug, the rate of SV40 DNA synthesis recovered from 20% to 70% of the untreated control. At that time, the levels of 25S DNAdeclined from 75% to 39 % of the total viral DNA synthesized during the 30-min
pulse-labeling period.
Effects of a "shift-up in cell growth" on SV40 DNAreplication. The results presented in the two previous sections suggest that the conversion of
20 30
FRACTION NUMBER
40
FIG. 2. Sedimentation of SV40 DNA synthesized during recoveryfromcycloheximideinhibition. Cyclo-heximide(10
,ig/ml)
wasaddedtofive infectedAGMK monolayer cultures for 3 hr. At that time, the drug was removed from four of these cultures. Viral DNA synthesis was measured by pulse-labeling with 3H-thymidine (I,uCi/ml)for30-min intervals. (A) Cyclo-heximide not removed; (B-E) cycloheximide was removedand the cultures were labeledat 0 to0.5 hr,0.5 to Ihr, 1.5 to 2hr, and 3 to3.5hr, respectively,
after reversalof the inhibition. (F) No cycloheximide added. Theprocedures forsucrose gradient
sedimenta-tion are presented in figure 1. (0) 3H counts per minute; thearrow
represenits
the peak ofa 21S 32p_ labeled SV40 DNA sedimentation marker.25S to 21S DNA requires a protein(s) that is
reduced in amount by cycloheximide treatment. At2 to 3 hrafter removal of thisdrug, the
pro-tein(s)ispresent in sufficientquantitiesto permit
thenormalrateof conversion of25S DNAto21S mature DNA.Thesedata, however, restsolelyon the use of an inhibitor (cycloheximide) and thereforerequireconfirmationbydifferent
experi-mentalprocedures.
on November 11, 2019 by guest
http://jvi.asm.org/
[image:4.491.244.439.76.413.2]TABLE 2. Recovery of SV40 DNA synthesis from cycloheximide inhibition
Sucrose gradient BNCb
Cycloheximideconcn Labeling time
Per cent Per cent Per cent Percent of control 25SDNAa ofcontrol 25SDNAa
,Ag/ml hr
10 (not removed) 1-1.5c 1.8 49 0.9 52
10 (removed) 0-0.5d 21 74 25 68
10 (removed) 0.5-1 54 60 50 58
10 (removed) 1.5-2 68 39 68 47
10 (removed) 3-3.5 87 35 100 39
0 (notadded) 3_3.5e lOOf 33 lOOf 35
aPercentageoftotalDNAsynthesizedduring the time intervalindicated that sedimented at
25S
or eluted with 2% caffeine from a BNC column.IBenzoylated-naphoylated diethylaminoethyl cellulose.
cLabeling time 1 to 1.5 hr aftercycloheximidide was added.
dLabeling time 0 to 0.5 hr after removal of cycloheximide.
e Nocycloheximidewaseveradded; 3-to 3.5-hrlabeling time refers to the same time interval as for the sample labeled3 to 3.5 hr after removal ofcycloheximide.
fThe 100% valuefor the 3H counts per minute as measured by sucrose gradients is 16,500
counts/
min andbyBNC is 136,000counts/min.To determine whether protein synthesis plays arole in the 25Sto 21S conversion, cultures of AGMK cells in two very different physiological
states
(growing
and resting cells) were infected with SV40.Growing AGMK cells(about 40% ofthe petri dish surface contained cells) were
in-fected with large-plaque SV40 in 2 ml of
Dul-becco's medium containing 2%calf serum. After 3hrofadsorption, theexcess virus wasremoved,
and 15mlof Dulbecco's medium containing 10% calf serum wasaddedtotheculture. Atthesame
time, monolayer cultures of AGMK cells (100%
of the
petri
dish surface containedcells)
wereinfected with
large-plaque
SV40 in 2ml of Dulbecco's medium containing 2% depleted (21) calfserum. Afterthe 3-hr adsorption period, theexcessviruswasremoved,and 15ml ofDulbecco's mediumcontaining 2% depleted calfserum was
added to the cultures. At 30 hr after
infection,
each of these cultures was
pulse-labeled
with 3H-thymidine(25,Ci/ml)
for a differentlength
of time
(4,
5,8, 12, 20,
or 25min) depending
on the experiment. Thepulse-labeling
period was terminatedby washing
thecell cultures withice-cold PBS, and the cells were
lysed
with0.6%
SDS. A 0.2-ml
sample
of the 1M NaCl-SDSsoluble fraction was
adjusted
to 0.1 M NaOH (finalconcentration)
and incubated at 37 C for 10min. Thissample
wasthencentrifuged
through analkaline sucrosegradient,
and thepercentageof the total 3H counts per minute that co-sedi-mentedwitha32P-labeledSV40 DNA sedimenta-tion marker was determined
(Fig.
3).
In the in-fectedmonolayer
cultures withdepleted
serummedium, therewas a 7- to8-min
delay
in thepro-so80h~~~~~~~~~~~~ ~ ~
%ofcpm60+
MATURE
SV40 /
DNA
40L
20t / /
4 8 12 16 20 24
TIME AFTERISOTOPE ADDITION
FIG. 3. Kineticsof formation of3H-labeledmature
viral DNA as afunction ofthelength of exposure to
3H-thymidine. At 30 hr after SV40 infection of
sta-tionaryorgrowingcultures ofAGMKcells, each cul-ture waslabeled with 3H-thymidine for the length of
time (inminutes) indicatedon the abscissa. A sample of the I M NaCI-SDS solublefractionwasadjustedto
0.1 M NaOH and incubatedat37Cfor10min.Alkaline
sucrose gradients (5 to20% sucrose in 0.1 m NaOH
and0.9 M NaCI) were runfor2 hrat40,000rev/min
in an SW 50.1 rotor. The ordinate is plottedas the
percentage oftotal counts per minute that co-sedi-mented witha 32P-labeledSV40 closed circular DNA sedimentation marker.(-,*, *) Stationarycultures; (0, O, i) growing cultures.
duction of 3H-labeled mature viral DNA. This
delayis duetothe time
required
tocomplete
one replication ofanSV40 genome(one doubling
ofa DNA
molecule).
It isclearly
not due to thedifficulty
ofgetting
theradioisotope
into thecell,
because the
incorporation
of3H-thymidine
into viral DNA(25S
and21S)
isapproximately
linear116 KANG ETAL. J. VIROL.
on November 11, 2019 by guest
http://jvi.asm.org/
[image:5.491.266.458.313.436.2]with timeovertheseshort timeperiods (12). The lag in the production of mature labeled viral DNA has been attributed, therefore,toaslow or ratelimitingstepin thereplicationofSV40DNA (12). This slowstephas beenshowntobe the
con-version of 25Sto21S DNA (12). In the infected growing cultures with 10% calfserumthelagin theproductionof labeledmatureviral DNAwas
4to 5 min (Fig. 3). It canalso beseen that the
rate ofproduction of mature viral DNA in the growing cultures (compare the slopes of thetwo curves) wasgreaterthan inthenongrowingcells. These results demonstrate that in infected growing
cells, where therateofprotein synthesisishigher than innongrowing infectedcells,theconversion of 25Sto215DNA takes less timethan in
mono-layer cultures. This is consistent with the idea that active protein synthesis is required for the 25S to 21S conversion and that this is a
rate-limitingstepin confluentmonolayer cultures.Itis important to point out that even in infected
growing cultures, where the rate ofprotein
syn-thesisisveryhigh,thereisstillalagordelayin the production of labeled mature viral DNA.
Effectsof SV40temperature-sensitive coat
pro-tein mutation on viral DNA replication. The pre-vious resultssuggestedthatprotein synthesiswas
required fortheproductionofmatureviralDNA (21S) from its255precursor.Ifthis is correct, the protein(s) required for this process could be coded forbythevirus,thecell,orboth. The fact thatafunctionalcoat protein is required for the synthesis of single-stranded 4X174 DNA (14) and the maturation of double-stranded linear lambda DNA (15) suggested the possibility that SV40coatproteins mightbeinvolved in the
con-version of 255to21S viral DNA.
TheSV40 large-plaquemutanthasbeenshown to possess a temperature-sensitive mutation in a
coatproteingene(16, 17, 19, 20). Todemonstrate the temperature-sensitive nature of the SV40 large-plaque mutant, the following experiment
was performed. Two cultures of AGMK cells wereinfected withlarge-plaque SV40andanother setoftwocultures with wild-type SV40.
Adsorp-tion wasallowedtoproceed for 3 hr at36C. At the end of that time, one culture infected with
large-plaque SV40 culture infected with wild-type SV40 wereshifted to 40 C. All four infected cell cultures were harvested at 72 hr after infection and the sonically treated celllysatesweretitrated
forinfectious virusat36 C (experiment A, Table 3). Wild-type SV40 replicated almost as wellat
40 C as at 36 C. On the other hand, the large-plaque SV40 was restricted in its replication
(greater than 99.9%) at 40C.
To determine the time course of the SV40
large-plaque temperature-sensitive restriction, the
same experiment was repeated but the infected
cultureswereshiftedupto40 Cat24hrafter in-fection. Infected cell lysates were prepared by
sonic treatment at 10, 24, and 72 hr after infec-tion, and infectious virus was titered at 36C (experiment B, Table 3). Wild-type SV40 was
made innormal quantities after the shiftto 40 C. Little or no increase in infectious virus was
ob-served when the SV40 large-plaque mutant was
shiftedto40 C at24hrafterinfection. This
indi-cates that the temperature-sensitive step occurs
afterviral DNAsynthesis normallybegins (16to
20 hr afterinfection at 36C). These data are in
excellentagreement with those of others (16, 17, 19, 20) and are consistent with the results that
indicate that thelarge-plaque SV40 isa
tempera-ture-sensitive mutant for a late function (coat
protein; 16, 17).
To observe whether the SV40 large-plaque mutation in the viral coatprotein had anyeffect onviral DNAreplication,twomonolayercultures of AGMK cells were infected with large-plaque
SV40 at 36C. After 24 hr ofincubation at the permissive temperature (36C), one of the
in-fectedcellcultureswas shiftedto40 Cfor 24 hr. 3H-thymidine (1,Ci/ml) was then added to a
cultureateachtemperature (36 and 40 C) for 45 min. Atthe end of this labeling period, the cells werewashedwithcold PBS andlysed with 0.6% SDS. Asample of the 1MNaCl-SDSsupernatant
fraction wassedimentedthrough asucrose
[image:6.491.46.440.555.625.2]gradi-entfor3 hrat40,000rev/min. The results of this
TABLE 3. Temperature-sensitive restriction of large-plaque SV40
ExptA:Gtiter of virusreplicatedat ExptB:btiter at time after infection Virus
36C 40C 10hr 24hr 72 hr
Wild-type SV40... 4.2 X 108 1.2 X
108
2.5 X 104 8.8 X 104 1.8 X108
Large-plaque
SV40... 1.0 X 109 5.0 X106
6.0 X 104 1.4 X106
3.0 Xl0r
a Viruswasadsorbed for 3 hrat36 C and thenshifted to 40 C. Virus production was titrated at 72 hr after infectionat36C.
bViruswasadsorbed andincubated for 24 hr at 36 C. Cultures were then shifted to 40 C, and virus production was titrated at 36 C. Samples were obtained at 10, 24, and 72 hr after infection.
VOL. 7, 1971
on November 11, 2019 by guest
http://jvi.asm.org/
KANG ET AL.
Hcpm
20-3
H cpm B
X10-3 2 z
10 20 30 40
FRACTIONNUMBER
FIG. 4. Sedimentation of large-plaque SV40 DNA synthesized at 36 or 40 C. Twomoniolayercultures of AGMK cells were infected with large-plaque SV40 at 36 C; 24 hr later, one culture was shifted up to 40 C. At 48 hr afterinfection, each culture was labeled with3H-thymidine (1juCi/ml)for45min. Asample of the IMNaCI-SDS soluble fraction was sedimented in
aneutral sucrose gradient as described in Fig. 1. (A) 40C; (B) 36C. (0) 3H-counts per minute; the arrow represents the peak ofa 32P-labeled 215 SV40 DNA sedimentationmarker.
experiment are presented in Fig. 4. SV40 DNA
was synthesized atboth thepermissive and non-permissive temperatures. About 1.9 times more
viral DNA was made at 40C than at 36C. In
bothcases,however,22 to23 %of theSV40DNA made in a 45-min pulse-labeling period was 25S DNA. The
only
detectable difference between thetwo cultures was thatabout twiceasmuch viral
DNA wasmadeat40Cas at36C.Twoadditional
experiments were
performed
todetermine whether this difference was duetothe absence ofa func-tionalcoatproteinat40Corjust
dueto adiffer-ence in the incubation temperature of the two cultures. First, the
experiment
described abovewas
repeated
with thewild-type
SV40instead ofthe large-plaque virus. In this
experiment,
the wild-typeviruswasfoundtosynthesize
1.7timesmore viral DNA at 40C than at 36C. This is consistent with the
interpretation
that thelarge
plaqueSV40synthesizesmoreviralDNAat40 C thanat36C because ofa temperature difference
and not becauseafunctional viralcoat
protein
is missing. Inasecondtype ofexperiment,
six mono-layer cultures of AGMK cellswereinfected with large-plaque SV40 at 36C. At 48 hr afterinfec-tion, half of these cultureswereshiftedto40 Cfor
1 hr.
3H-thymidine
was then added to eachin-fectedculture(at36and 40
C),
andthe amountof viral and cellular DNAsynthesized
during 4-,
7.5-, and 8-hr
labeling
periods
wasdeterminedasdescribed previously. In each case, the rate of viral DNA synthesis was 1.6 to 2.5 times greater at 40C than at 36 C. The rate of cellular DNA
synthesis inthese same cells was 1.5 to 2.3 times greater at 40 C than at 36 C. Since a functional
viral coatprotein is clearlynot required for
cel-lularDNAsynthesis,the higherlevels of host-cell DNA replication at 40 C must be due to the
difference in incubation temperatures. Thus, the rates of cellular and viral DNA synthesis were
increasedby aboutthe same amounts (1.5to2.5 times) at 40C. This is also consistent with the
interpretation that the absence of a functional
viral coatproteinat40 C is notresponsible for the observed increase in the rateof viral DNA syn-thesis.
DISCUSSION
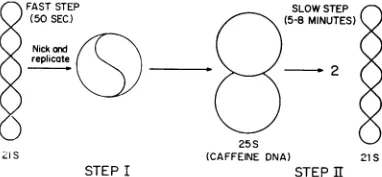
Figure 5 presents amodel forSV40 DNA repli-cation based ontheresults of Levine etal. (12).
The mature SV40closed circular and suprcoiled
DNA is first nicked and replication begins. Be-cause16S, nickedcircularDNAisneverobserved witheither shortor long
pulses
of3H-thymidine,
replicationmust startvery soon after theopening
ofthe SV40
polynucleotide
strand. At least onestrand ofthe SV40duplexmustbeopenatsome timeduringsynthesistopermitasemiconversative modeofreplicationto occur (8,22). Mostofthe
replicating molecules observed inaninfected cell have completed 90% of their
replication
(25S form; 12). This ledtothe suggestion that step Iofthis
replication
modelwasthefaststep(50 sec),
whereas step IIwas slower
(5
to8min). Thus,
a rate-limitingorslowstep(II)
occurs atthe endofthereplication ofanSV40circle
(12).
Mutants of
polyoma
virus(1-3)
have been isolatedthatdo notsynthesize
viralDNAatthenonpermissivetemperature. Ifthese mutantsare
shifted up to the
nonpermissive
temperature during polyoma DNAsynthesis,
thenpolyoma
DNA replication stops (4). This shows that the
FAST STEP
(50SEC.)
Nickand replicate
STEPI
SLOWSTEP
(5-8MINUTES)><
-~~~ -~~~2
25S
(CAFFEINE DNA) 21S
STEP E
FIG. 5. Modelfor the replication ofSV40 DNA.
Each line representsadouble helix. The 21SDNA is
closed circular and supercoiled DNA; 25S DNA is
thepredominantformofreplicatingmolecules observed
in aninfectedcell.
118 J. VIROL.
on November 11, 2019 by guest
http://jvi.asm.org/
[image:7.491.262.453.517.606.2]presenceofavirus-specific protein is
required
con-currently with the synthesis of
polyoma
DNA. Although it has not been proven, thisprotein
ismost likely required for the initiation step of polyomareplication(aviral DNAintermediate is
not accumulated; 4). The addition of
cyclohexi-mide to SV40-infected cells inhibits viral DNA replication (11) by 99% within2 hr. One
might
expect thatcycloheximide inhibitsthe
synthesis
ofa virus-specificprotein (SV40
protein
analogous
to that found with the polyoma
mutants)
that initiates the synthesis of SV40 DNA. Ascyclo-heximide inhibition of SV40 DNA
replication
proceeds (3 to 4hr after the additionofthe
drug),
thereis anincreasein theproportion of25SDNA synthesized (from one-fourth to one-half 25S
DNA) in a 1-hrpulse. Thiswould be expected if
theinitiation step wasinhibited at a slowerrate than thefinishing step (step
II).
This is not ob-served during the first hour of cycloheximidetreatment,indicatingthatforthis timeintervalthe
protein(s) needed for the finishing step is in-hibited or inactivated at the same rate as the
initiator
protein(s).
It isimportant
topoint
out that throughout thecycloheximide treatment (or recovery) lower than normal levels of 21S and 25S viralDNA wereobserved. Thesedata indicate that both initiation and finishing steps arein-hibitedbycycloheximide.
Removal of cycloheximide from infected cells allows the recovery of SV40 DNA synthesis to normal levels. The first viral DNA accumulated (labeled with 3H-thymidine) after removal ofthe
drug
(0
to 0.5hr)
ismainly 25S(75%).
This is consistentwith theexpectationthat initiatorpro-tein(s) recover from cycloheximide inhibition at a faster rate than the finishing
protein(s).
The presence of normal levels of finishing protein(s)canbeobserved
by
3hr after reversal of thisdrug. These data show that finishingprotein(s)
is in-activated or inhibited at a faster rate and re-coversfrom cycloheximide inhibitionat aslowerrate than theinitiator protein(s). By 0.5 to 1 hr after removal of cycloheximide from infected cultures, the rateofviralDNAsynthesisis 50 to
55% ofnormal, and thelevels of25S DNA are 40 to 45 %higherthannormal.
Theuseofaninhibitor likecycloheximide
suf-fersfromtheproblemthattheinfected
cell
ismostlikely in an abnormal state during the inhibition orrecovery stagesofthetreatment. It was
there-foredesirable toobtainanindependent confirma-tionofarequirement forproteinsynthesis for the 25S to 21S conversion step. To do this, stage II of SV40 DNA
replication
was measured in in-fectedgrowing and resting cells. The rate of pro-teinsynthesis wasfoundto bethree to five timesgreater in infected growing cells than in the
in-fected confluentmonolayer cells. The conversion of 25S to 21S DNA was also faster in growing
cells. There was both a decrease in the delay of the production of 3H-labeled 21S DNA and an
increase in the rate ofproduction of 21S DNA.
These data demonstratethat atleastaportion of
the rate-limiting step (step II) in SV40 DNA replication can be eliminated by using infected growing cells. It is important to note, however,
that alagordelayin theappearanceof3H-labeled 21SDNAisstillobserved withthese cells (4- to
5-min delay), and therefore some of the factors needed for step II arestillrate-limiting for SV40
DNAreplication.
There are several difficulties in comparing
events in growing and stationary cells. Virus
production and the rate ofincorporation of
3H-thymidineintoviralDNAarefasterin the growing
cell system. It is clear that 30 hr after infection
with growing cells may be analogous to a later
time period in infected monolayer cultures. To
eliminate theseobjections, therateofconversion
of 25Sto21SDNAhas been measuredatdifferent times after lytic infection of stationary cultures. At all timestested (24, 36,48, and60hr),therate
of conversion of 25S DNA to 21S DNA was
about the same (6to 8 min). Since the ratio of
replicating DNA to coat protein is probably differentat24and 48 hrafterinfection,thisresult
is consistent with thesuggestion that SV40 coat
protein has no effect on this step II conversion. Presumably, if one could look
early
enough in infection (when the replicating pool ofDNA isvery
small),
thedelay
in the 25Sto 21Sconver-sion would be eliminated.In
addition,
thesedata eliminate thepossibility that the time after infec-tion thatone measuresthe 25Sto21S conversionmay not be comparable in resting and growing cells.
The experiments presented in this paper sug-gest thata protein(s), orproteinsynthesis, is
re-quired at two stages in the replication ofSV40 DNAmolecules.Avirus-specificinitiator protein
is
probably
requiredto start around ofreplica-tion (step I). A second protein, or proteins, ap-pears to beneededeithertofinishthe last10%of
thereplicationor toclose the SV40 polynucleotide
strands,
or to do both. Atleast
one SV40 coatprotein, prescribed by the SV40 large-plaque mutation, is not involvedin DNA replication.
ACKNOWLEDGMENTS
Theexcellenttechnical assistance of Angelika K.Tereskyis gratefullyacknowledged.
Thisresearch wassupported byPublicHealth Service grant CA 11 049-02fromtheNational Cancer Instituteandby grant E-591 fromtheAmerican CancerSociety. We thank theWhitehall
VOL. 7, 1971
on November 11, 2019 by guest
http://jvi.asm.org/
Foundation in the Biology DepartmentatPrinceton University for thegenerous useof their equipment.
LITERATURE CITED
1. Di Mayorca, G., J. Callender, G. Marin, and R. Giordano. 1969. Temperature-sensitive mutants of polyomai virus. Virology 38:126-133.
2. Eckhardt, W. 1969. Complementation and transformation
by temperature-sensitivemutants ofpolyolmavirus.
Virol-ogy38:120-125.
3. Fried, M. 1965.Cell transformning ability ofa
temperaiture-sensitivemutant ofpolyomra virus. Proc. Nat. Acad. Sci.
U.S.A.53:486-491.
4. Fried, M. 1970. Characterizaton of atemperature sensitive mutantofpolyomavirus.Virology40:605-617.
5.Gershon, D.,and L. Sachs. 1964. Thetemliporal relationships ofproteinand DNAsynthesisinpolyoma virus
develop-ment.Virology 24:604-609.
6.Gilead, Z., andH. S.Ginsberg. 1965. Characterization ofa
tum-iorlike antigen in type 12 and type 18
adenovirus-infected cells. J. Bacteriol.90:120-125.
7.Gillam, I., S. Millward, D. Blew, M. vonTigerstroiimm,E Winsmer, and G. M. Tener. 1967. Theseparationof
sol-uble ribonucleic acids on benzoylated diethylaminoethyl
cellulose. Biochemistry 6:3043-3056.
8. Hirt, B. 1966. Evidence for semiconservative replication of
circular polyomiia DNA. Proc. Nat. Acad. Sci. U.S.A.
55:997-1004.
9. Hirt, B. 1967. Selective extraction of polyoma DNA from infected mouse cell cultures. J. Mol. Biol. 26:365-369.
10. Kit, S. 1967. Enzymie interactions in cell cultures during
productive and abortive infections by papovavirus SV40,
p.495-525. Iln J. S. Colter and W. Paranchych (ed.), The
molecular biologyofviruses.
11. Kit, S., T. Kurimura, R.A. deTorres, and D. R. Dubbs.
1969. Simian virus 40 deoxyribonucleic acid replication.
I. Effect of cycloheximide on the replication of SV40 deoxyribonucleic acid in monkey kidney cells and in
heterokaryons of SV40-transformed and susceptible cells.
J. Virol. 3:25-32.
12. Levine, A. J., H. S. Kang, and F. Billheimer. 1970. DNA
replicationinSV40 infected cells.1.Analysesof replicating
SV40DNA.J. Mol. Biol. 50:549-568.
13. Levine, A. J., and A. K. Teresky. 1970. Deoxyribonucleic acid replication in simian virus 40-infected cells. II.
De-tection and characterization of simian virus 40 pseudo-virions.J. Virol. 5:451-457.
14. Lindquist, B. H., andR.L.Sinsheimer. 1967. Theprocessof infection with bacteriophage4Xl74. XV. Bacteriophage DNA synthesis in abortive infections with a set of
con-ditional lethalmutations.J. Mol. Biol. 30:69-80. 15. MacKinlay, A.G.,andA. D.Kaiser.1968. DNA replication
in head mutants ofbacteriophage lambda J. Mol. Biol. 39:679-683.
16. Ozer, H. L., and K. K. Takemoto. 1969. Siteof host restric-tionofsimian virus40mutantsinanestablished African
greenmonkey kidney cell line. J. Virol. 4:408-415. 17. Ozer, H. L., K. Takemoto,R.L.Kirschstein, and D.
Axel-rod. 1969. Immunochemnical characterization of plaque
mutantsof simian virus40.J. Virol. 3:17-24.
18. Sedat, J.,A. Lyon, andR.L.Sinsheimner. 1969. Purification of Escherichia coli pulse-labeled RNA by benzoylated DEAE-cellulose chromatography. J. Mol. Biol. 44:415-434.
20. Takemoto, K., G. Todaro, and K. Habel. 1968.Recoveryof SV40 virus with genetic markers of original inducing virus fromSV40 transformedmousecells.Virology 35:1-8. 19. Takemoto, K., R. L. Kirschstein, and K. Habel. 1966.
Mu-tantsof simian virus 40 differing in plaque size,
oncoge-nicity, and heat sensitivity. J. Bacteriol. 92:990-994. 21. Todaro, G., Y. Matsuya, S. Bloom, A. Robbins, and H.
Green. 1967. Stimulation of RNA synthesis and cell di-visioninresting cells byafactorpresent inserum, p. 87-101. Iz V. Defendi and M. Stoker (ed.), Growth regulating
substances for animal cells in culture. Wistar Inst. Press, Monographno.7.
22. Vinograd, J., andJ. Lebowitz. 1966. Physical and topologica properties of circularDNA.J. Gen.Physiol. 49:103-125