JOURNAL OFVIROLOGY, June 2009, p. 5339–5352 Vol. 83, No. 11 0022-538X/09/$08.00⫹0 doi:10.1128/JVI.00232-09
Copyright © 2009, American Society for Microbiology. All Rights Reserved.
Dimerization and a Novel Tax Speckled Structure Localization Signal
Are Required for Tax Nuclear Localization
䌤
Kimberly A. Fryrear, Sarah S. Durkin, Saurabh K. Gupta, Jessica B. Tiedebohl, and O. John Semmes*
Department of Microbiology and Molecular Cell Biology, Eastern Virginia Medical School,700 W. Olney Road, Norfolk, Virginia 23507
Received 2 February 2009/Accepted 2 March 2009
The human T-cell leukemia virus type 1 oncoprotein Tax has pleiotropic activities, a subset of which likely leads to immortalization of T cells. Tax is expressed and known to function in both the cell nucleus and the cytoplasm. Tax has defined nuclear localization (NLS) and nuclear export signals that enable shuttling between the two compartments. In this study, we identified a novel region in Tax that targets the protein to discrete nuclear foci that we have previously termed Tax speckled structures (TSS). We demonstrated that the identified region is both necessary and sufficient for directing proteins to TSS. This novel TSS localization signal (TSLS), spanning amino acids 50 to 75, is separable from and adjacent to the NLS of Tax. Coexpression of a Tax NLS mutant and a Tax TSLS mutant rescued the nuclear entry and subnuclear TSS targeting of both proteins, demonstrating that these signals are independent domains. Our analysis also revealed that Tax proteins deficient for dimerization fail to localize to the nucleus. Consequently, when we restored dimerization via induction of a heterologous “dimerizer” domain, nuclear localization was rescued. Thus, we defined additional domains in Tax specific for nuclear localization and subnuclear targeting. Our results reveal a more complex network for regulation of Tax subcellular localization and subsequent function.
Human T-cell leukemia virus type 1 (HTLV-1) is the etio-logic agent of adult T-cell leukemia as well as subneoplastic disease states, the most prevalent being HTLV-1-associated myelopathy/tropical spastic paraparesis (22, 44, 47, 63, 68). Tax is a potent transcriptional transactivator of both viral and cel-lular genes and can physically associate with a number of cellular proteins, thus altering their functions (10). Expression of Tax results in the dysregulation of critical cell functions, including cell cycle regulation, proliferation, DNA damage rec-ognition and repair, and apoptosis. Furthermore, although the pathogenesis of adult T-cell leukemia is not completely under-stood, the transformation of T cells infected with HTLV-1 can be logically attributed to genomic instability resulting from the pleiotropic activities of the viral protein Tax (30, 41).
Of particular note for this study, Tax has documented nu-clear and cytoplasmic functions; thus, the regulation of Tax localization to subcellular compartments is a critical aspect of overall Tax function (11, 23, 52). Tax contains a nuclear local-ization signal (NLS) sequence in the N-terminal region of the protein that is necessary and sufficient for nuclear localization (23, 53, 55). We previously showed that Tax “shuttles” between nuclear and cytoplasmic subcellular compartments and identi-fied a consensus nuclear export signal (NES) sequence defined by amino acids 190 to 203 (11). Subsequently, a fine-mapping mutational analysis of this region by Alefantis et al. (1) clearly demonstrated a functional NES at this site. These signals pre-sumably mediate interactions between Tax and karyopherins in the nuclear pore complex to allow Tax to translocate through the nuclear membrane (58, 64). The specific importins and
exportins involved in Tax transport have yet to be identified, but recent studies indicate that Tax can both import and export from the nucleus without the aid of carrier proteins or energy and can interact directly with the FG nucleoporins within the nuclear pore complex (67). Both the NLS and the NES of Tax are atypical. The NLS is large, involving the first 48 amino acids of Tax, and lacks the basic residues that define classical NLS (23, 53, 55). The NES of Tax contains a classic leucine-rich region between amino acids 188 and 202 that is capable of mediating export of Tax via the CRM-1 pathway but displays notably slower kinetics (1, 11). This NES is believed to be masked in the native Tax protein, and there is strong evidence that ubiquitylation serves to “unmask” the NES, in response to DNA damage, for instance (21). In fact, “nuclear” Tax is pre-dominately sumoylated whereas “cytoplasmic” Tax appears to be primarily ubiquitylated (37).
While in the cytoplasm, Tax activates transcription via the NF-B pathway (61) by directly binding to the IB kinase
␥/NF-B essential modulator, leading to the phosphorylation and degradation of IB and the release of NF-B (38, 43, 62). In the nucleus, Tax interacts with cellular transcription factors to acti-vate or repress transcription of cellular and viral genes via the ATF/CREB (42), AP-1 (19), and SRF (29) pathways (reviewed in reference 33). The list of nuclear proteins that bind to Tax and are functionally modulated is growing and includes APC (39), CBP (7, 48), Cdk4 (25), Chk1 (45), Chk2 (26), DNA protein kinase (15), MAD1 (32), c-Myc (50), and retinoblastoma protein (34). While the NLS and NES provide one level of regulation for Tax localization, it is likely that there are other mechanisms at work regulating Tax subcellular and subnuclear localization and asso-ciated functions. In addition to interaction with other proteins, Tax is also capable of dimerization/oligomerization, and studies indicate that optimal transcriptionaltrans-activation by Tax re-quires that it be in a dimeric/oligomeric form (5, 23, 31, 65).
* Corresponding author. Mailing address: Department of Microbi-ology and Molecular Cell BiMicrobi-ology, Lewis Hall, 700 West Olney Road, Norfolk, VA 23507. Phone: (757) 446-5904. Fax: (757) 446-5766. E-mail: [email protected].
䌤Published ahead of print on 25 March 2009.
5339
on November 8, 2019 by guest
http://jvi.asm.org/
In previous studies, we and others have demonstrated that Tax enters the nucleus and is directed to discrete nuclear foci that we termed Tax speckled structures (TSS) (6, 52). TSS coincide with interchromatin granules and consist of multipro-tein complexes that partially overlap with subnuclear regions involved in splicing and transcription. We have also shown that Tax recruits cellular proteins involved in the DNA damage recognition and repair response into the TSS as well (15, 24, 26). The formation of TSS and the colocalization of spliceo-some component 35 (SC35), DNA protein kinase, Chk2, and 53BP1 within the TSS places Tax near cellular machinery for transcription, splicing, DNA damage response, and checkpoint activation (26). This may explain how Tax is able to affect multiple cellular functions simultaneously (41). Although the NLS for Tax has been previously described, the domain in Tax that dictates TSS localization has not yet been defined.
In this study, we identified the region in Tax containing the sequence that targets the protein into TSS. We fine mapped the TSS localization signal (TSLS) to a region containing amino acids 50 to 75. This sequence lies adjacent to but is completely separable from the Tax NLS. We also demon-strated that dimerization of Tax is required for nuclear local-ization. We showed that deletion of the dimerization domain of Tax prevents nuclear accumulation and results in the cyto-plasmic expression of the protein. The induction of dimeriza-tion in a Tax dimerizadimeriza-tion mutant restored nuclear localiza-tion. These results elucidate two novel mechanisms in the regulation of Tax subcellular localization and the subsequent function.
MATERIALS AND METHODS
Plasmids.Generation of the pSTaxGFP construct has been described previ-ously (16). STaxGFP mutants were created with PCR-based deletion mutagen-esis, using a QuickChange XL mutagenesis kit (Stratagene). The primer se-quences used for cloning and mutagenesis are available upon request. pSNLSTaxGFP constructs were created by inserting the simian virus 40 (SV40) T antigen NLS in frame between the S tag and thetax-gfpfusion, using a PCR-based ExCite mutagenesis kit (Stratagene). pSTax1-75GFP and pSNL-STax50-75GFP were constructed by PCR-based mutagenesis with a Phusion site-directed mutagenesis kit (Finnzymes, Inc.), using pSTaxGFP and pSNL-STaxGFP as the templates. A Myc peptide-tagged Tax construct (pMycTax) was a kind gift from Ralph Grassmann. A pMycTax(⌬30-52) construct was created by site-directed mutagenesis of pMycTax, using a QuickChange XL mutagenesis kit (Stratagene). The inducible dimerization construct pSTax(⌬100-150)-Fv-GFP was created by amplifying the Fv domain of pC4-Fv1E (Ariad Pharmaceuticals)
and inserting it into the ClaI site within pSTax(⌬99-150)GFP.
Cell culture and transient transfection.HEK 293 cells were maintained at 37°C in a humidified atmosphere of 5% CO2in air in Iscove’s modified
Dulbec-co’s medium supplemented with 10% fetal bovine serum and 1% penicillin/ streptomycin (Invitrogen). Transfections were performed by the standard cal-cium phosphate precipitation method (11). The cells were washed 16 h posttransfection and incubated at 37°C until harvest. Cells were harvested 48 h posttransfection following a single wash with 1⫻phosphate-buffered saline (PBS). Transcriptionaltrans-activation assays.HEK 293 cells were transfected with Tax-expressing plasmids and either HTLV-luciferase or the NF-B-luciferase reporter, using the calcium phosphate method. Cells were lysed in 400l of reporter/lysis buffer (Promega), and lysates were immediately frozen at⫺80°C. Samples were allowed to thaw on ice, collected, and quantified, using a Bradford assay (Bio-Rad). A total of 1g of protein from each sample was applied to 100 l of luciferase substrate (Promega), and a luciferase assay was immediately read in a Turner BD 20/20 luminometer. Transcriptional activation was analyzed and expressed as the level of activation over that of the reporter alone (level of activation⫽1).
Immunofluorescence confocal microscopy.HEK 293 cells were seeded at 1⫻ 105
cells/well on ethanol-washed coverslips in 6-well plates. The cells were sub-jected to transient transfection to express the desired proteins. After 48 h, the
cells were fixed by the paraformaldehyde/methanol method as previously de-scribed (52). The primary antibodies used were anti-SC35 mouse monoclonal S4045 (Sigma), anti-Myc rabbit polyclonal ab9106 (Abcam), and anti-Tax rabbit polyclonal (52) at a 1:1,000 dilution. The secondary antibodies were species-appropriate Alexa Fluor 594-conjugated antibodies (Molecular Probes) at dilu-tions of 1:1,000. Nuclei were counterstained with To-Pro-3 iodide (Molecular Probes) diluted 1:1,000 in 3% bovine serum albumin–PBS. The cells were then mounted on glass slides, using Vectashield with 4⬘,6-diamidino-2-phenylindole (Vector Laboratories). Confocal fluorescent images were acquired on a Zeiss LSM 510 confocal microscope (Carl Zeiss), using argon (488 nm), HeNe1 (543 nm), and HeNe2 (633 nm) lasers, and were imaged with LSM Image Browser software.
STaxGFP purification.HEK 293 cells expressing the STaxGFP fusion protein were lysed in 400l of M-Per mammalian protein extraction reagent (Pierce) with protease inhibitor cocktail (Roche Applied Science) and immediately fro-zen at⫺80°C. The lysates were assayed for total protein concentration using a Bradford protein assay and were normalized using a bovine serum albumin standard curve. The protein solution was brought to a total volume of 500l with M-Per for each sample, and 150l of S-protein agarose beads (Novagen) were applied to the sample. S-tagged proteins were purified from lysates as described previously (16). Supernatants containing isolated proteins were loaded onto an 8%- to 12%-gradient sodium dodecyl sulfate (SDS)-polyacrylamide gel and sep-arated by electrophoresis, transferred to Immobilon-P membranes (Millipore) by semidry transfer, and subjected to immunoblot analysis.
Immunoblot analysis.Proteins separated by electrophoresis were transferred to Immobilon-P membranes using the semidry transfer method with 400 milli-amps applied for 50 min. The membranes were then blocked for 1 h at room temperature in 1⫻Odyssey blocking buffer (Li-Cor Biosciences). Primary anti-bodies diluted in 1⫻Odyssey blocking buffer were applied to the membranes and allowed to interact at 4°C overnight on an orbital shaker. Membranes were washed four times for 5 min with PBS-1% Tween. Li-Cor Odyssey secondary antibodies were diluted to a concentration of 1:20,000 in 1⫻Odyssey blocking buffer containing 0.5% SDS and 0.5% Tween and then incubated for 1 h at room temperature on an orbital shaker while protected from light. The membranes were washed four times for 5 min with PBS-1% Tween and then stored in PBS and protected from light until analyzed. Blots were scanned and analyzed with a Li-Cor Odyssey scanner and software.
Dimerization assay. Full-length MycTax protein was coexpressed with STaxGFP or mutant Tax protein in HEK 293 cells. The Tax-expressing cells were subjected to lysis and subsequent S-bead purification as described above. The isolated proteins were separated by SDS-polyacrylamide gel electrophoresis and subjected to immunoblot analysis. STaxGFP and the corresponding mutants were detected with mouse monoclonal anti-green fluorescent protein (GFP) primary antibody (Santa Cruz) at a 1:1,000 dilution followed by Odyssey goat anti-mouse 488 (Li-Cor Biosciences) secondary antibody at a 1:20,000 dilution. Copurified MycTax was detected by polyclonal Myc peptide primary anti-body (Abcam) at a dilution of 1:1,000 followed by Odyssey goat anti-rabbit 680 (Li-Cor) secondary antibody at a dilution of 1:20,000.
Complementation analysis.Full-length MycTax was coexpressed with “test” mutant proteins in the STaxGFP backbone. The resulting cellular localization of the coexpressed proteins was visualized with immunofluorescence confocal mi-croscopy as previously described. MycTax and Tax mutant proteins were de-tected by staining with rabbit polyclonal anti-Myc peptide primary antibody (Abcam) at a dilution of 1:1,000 followed by goat anti-rabbit secondary antibody conjugated to Alexa Fluor 594 (Molecular Probes) at a 1:1,000 dilution.
Induced dimerization assay.STax(⌬100–150)-Fv-GFP was transiently trans-fected in HEK 293 cells as described above. Forty-eight hours after transfection, the cells were treated with AP20187 from an Argent homodimerization kit (Ariad Pharmaceuticals) at concentrations of 0 nM, 0.01 nM, 0.1 nM, 1.0 nM, 10 nM, 50 nM, and 100 nM for 24 h. Cells were then washed, fixed with 4% paraformaldehyde, permeabilized with methanol, and processed as previously described for confocal microscopy analysis. Images were analyzed for nuclear accumulation using MetaMorph image analysis software. Briefly, 100 cells from each condition were analyzed for nuclear fluorescence expressed as a percentage of total cell fluorescence.
RESULTS
Deletion-scanning mutational analysis of Tax. Previous structure/function studies of Tax involving mutational analysis involved amino acid substitution or refined scanning-linker approaches so as to preserve the transcriptional activity of the
on November 8, 2019 by guest
http://jvi.asm.org/
protein (53, 56). In this study, we constructed a series of con-secutive 25- to 50-amino-acid deletions that covered the length of the Tax protein as depicted in Fig. 1A. Our experimental objective in generating these larger deletions was to alter
[image:3.585.98.483.67.576.2]cel-lular localization. The parental construct, which we have pre-viously described (16), provided an N-terminal S tag for pro-tein purification and a C-terminal GFP fusion to facilitate localization studies and displays activity comparable to that of
FIG. 1. Tax mutants and corresponding transcriptional activity. (A) Diagram of STaxGFP depicting the deletion-scanning series of Tax mutants. Shown are the relative positions of the S peptide (S Tag), NLS, leucine zipper region (LZL), NES, activation domain (ACT), and GFP fusion (GFP). Also indicated are the CREB-interacting, zinc finger, SH3 binding, PDZ binding, dimerization, and P/CAF binding domains. (B) Efficient expression of Tax deletion mutants is comparable to that of wild-type protein. Shown is an immunoblot analysis of whole-cell lysates from 293 T cells transfected with the appropriate plasmids for expression of the indicated proteins. Rabbit polyclonal anti-GFP antibody was used to evaluate protein expression. (C) Transcriptional activity of Tax mutants on the HTLV-1-LTR-Luc and NF-B-Luc promoter/reporter constructs. HEK 293 cells were transiently transfected with the indicated reporter construct and either STaxGFP or the indicated deletion mutant, and lysates were analyzed for activity. The reported level of activation is relative to that of the control in which the indicated reporter construct was cotransfected with empty vector expression plasmid.
VOL. 83, 2009 NOVEL Tax SPECKLED STRUCTURE LOCALIZATION SIGNAL 5341
on November 8, 2019 by guest
http://jvi.asm.org/
wild-type Tax. When examined for protein expression levels via Western analysis, the deletion mutants generated comparable amounts of protein (Fig. 1B). We next analyzed each of the mutants for transcriptionaltrans-activation activity and subcel-lular localization. Nearly all of the deletion mutants displayed no transcriptional trans-activation for either an HTLV-1-re-sponsive or an NF-B-responsive promoter/reporter (Fig. 1C), consistent with previous observations that mutations within Tax ablate transcriptional activation (56). The one exception was the⌬323–353 mutant, which deletes the C-terminal por-tion of Tax downstream of the sequences mutated in the classic M47 mutant.
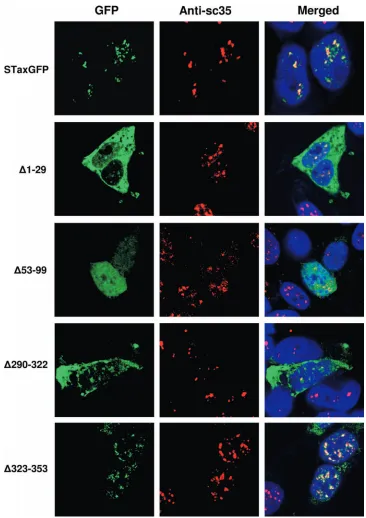
Confocal microscopy studies revealed that the mutants dis-played one of three phenotypes: localization to discrete TSS foci, diffuse nuclear localization, or cytoplasmic localization (Fig. 2). One mutant protein with the Tax NLS deleted in part, the⌬1–29 mutant, showed cytoplasmic expression with weak nuclear expression in TSS foci, while the second NLS mutant, the⌬30–52 mutant, was completely confined to the cytoplasm. Our results confirm previous studies that define the Tax NLS between amino acids 18 and 52 (55). Unexpectedly, mutants with the midregion of Tax deleted, including the⌬100–150,
⌬151–202,⌬203–254, and ⌬255–289 mutants, were also con-fined to the cytoplasm. One C-terminal deletion mutant, the
⌬290–322 mutant, was expressed in the cytoplasm but also displayed nuclear expression and foci formation. The extreme C-terminal deletion mutant, the⌬323–353 mutant, localized to the nucleus and formed nuclear foci. The⌬323–353 mutant protein displayed unusually high steady-state expression levels compared to the wild-type protein. One mutant, the⌬53–99 mutant, displayed a diffusely nuclear localization with no ap-parent foci. It should be noted that between 10% and 20% of cells that express wild-type Tax protein show cytoplasmic ex-pression in addition to the classic nuclear TSS localization. In this context, the⌬53–99 mutant protein was observed to have cytoplasmic localization in 43% of expressing cells in addition to the diffuse nuclear localization. In all cases, both wild-type protein and the⌬53–99 mutant displayed nuclear expression. We next determined whether the foci formed by the nucleus-expressed Tax mutants were TSS, as defined by colocalization with the cellular protein SC35. Three of the four Tax mutants, which had displayed at least partial nuclear expression, were able to form TSS (Fig. 3), including the⌬1–29 mutant and the two C-terminal Tax deletion mutants, the⌬290–322 and⌬323– 353 mutants. Again the⌬53–99 Tax mutant was able to enter the nucleus but clearly did not form TSS. In addition, we observed that deletion of amino acids 53 to 99 does not impact NLS function but disrupts TSS localization.
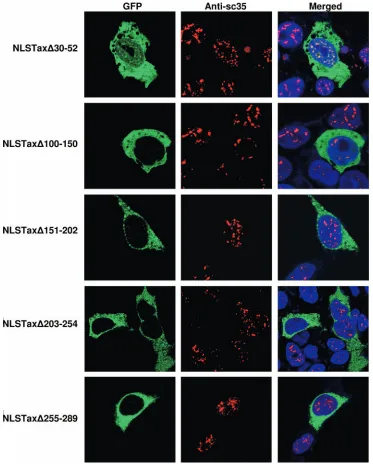
Determining loss of function for cytoplasm-expressed Tax mutants.In the scanning series of deletion mutants of Tax, there were five Tax mutants that were confined to the cyto-plasm. One mutant (the⌬30–52 mutant) has a deletion of the Tax NLS, and the remaining four mutants have deletions in the midregion of Tax. We hypothesized that if these deletions affected the functioning of the Tax NLS, the addition of an exogenous NLS would restore nuclear expression of the mu-tants. To test this hypothesis, we added the SV40 Tag NLS in frame at the N-terminal of each of the five cytoplasmic Tax mutants. As expected, the presence of an exogenous NLS was able to restore nuclear expression for the mutant that had a
deletion of the Tax NLS, the⌬30–52 mutant (Fig. 4). In fact, the NLSTax⌬30–52 protein was competent for TSS formation. However, all of the mutations in the midregion of Tax failed to be rescued by fusion to an exogenous NLS. This finding sug-gests that these mutant proteins were deficient for a function other than that fulfilled by a classic NLS sequence. Implied in this conclusion is that Tax nuclear localization is dependent on more than the presence of an NLS.
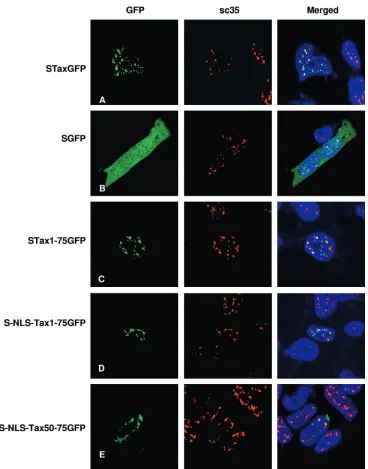
A novel domain within Tax amino acid residues 50 to 75 is sufficient for localization to TSS. We have shown that the deletion of amino acids 53 to 99 allows the retention of nuclear expression but disrupts TSS formation. We also showed that the addition of a heterologous NLS to the⌬30–52 Tax NLS mutant rescues nuclear expression and also rescues TSS for-mation, implying that the region required for TSS formation resides in the NLS mutant. These observations indicate that amino acids 53 to 99 contain the Tax TSS localization signal. In order to more fully characterize this putative TSLS, we de-signed a construct containing the first 75 amino acids of Tax fused to an N-terminal S tag and a C-terminal GFP. This construct, which contains the Tax NLS, was able to target GFP into TSS in a fashion that was indistinguishable from that of wild-type Tax (Fig. 5A and C). In comparison, a construct containing only the S tag and GFP was expressed diffusely throughout the cell, with no targeting to nuclear foci (Fig. 5B). The NLS for Tax is contained in the first 50 amino acids (23, 53, 55), and deletion of this region ablates nuclear localization. In order to determine if the TSLS is distinct from the NLS, we inserted an exogenous NLS from the SV40 T in frame and N-terminally to Tax1-75GFP. This insertion did not interfere with the targeting of S-NLS-Tax1-75GFP to TSS (Fig. 5D). We then effectively deleted the first 49 amino acids of Tax by creating S-NLS-Tax50-75GFP and observed that this construct was also able to target the GFP fusion into the TSS (Fig. 5E). This result established that the TSLS is outside of the Tax NLS and resides within amino acids 50 to 75 of Tax.
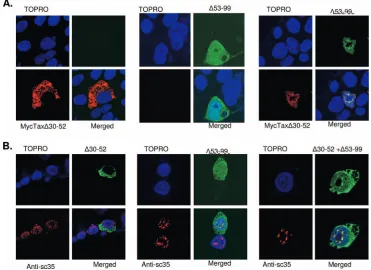
Complementation analyses of NLS and TSLS mutants.We then hypothesized that if the Tax NLS and TSLS are indepen-dent sequences, then coexpression of the two mutant proteins should result in dimerization and rescue of Tax localization to TSS. We therefore coexpressed the MycTax⌬30–52 mutant with the ⌬53–99 mutant to test whether the mutants could rescue nuclear localization and foci formation. The My-cTax⌬30–52 mutant failed to localize to the nucleus, and the
⌬53–99 mutant failed to form TSS, as expected (Fig. 6A). However, coexpression of the MycTax⌬30–52 and⌬53–99 mu-tants resulted in normal nuclear expression and TSS formation (Fig. 6A). To confirm that the foci formed by the coexpression of the mutants were TSS, we expressed the GFP fusion version of each mutant either alone or in combination (Fig. 6B) and observed colocalization of each with SC35. As was the case for the previous experiment, expression of either the⌬30–52 or
⌬53–99 mutant resulted in Tax protein that failed to localize to the nucleus or to form TSS, respectively. Coexpression of the
⌬30–52 and⌬53–99 mutants resulted in nuclear localization and the formation of TSS that overlap with the expression of SC35. Complementation by these two different domain mu-tants clearly demonstrated that the Tax NLS and TSLS are separate and sufficient for directing specific subcellular local-ization.
on November 8, 2019 by guest
http://jvi.asm.org/
Full-length Tax rescues nuclear localization of Tax midre-gion mutants.Previous studies have established that Tax forms dimers and that optimal transcriptionaltrans-activation by Tax requires Tax dimerization (5, 31). The described dimerization domain spans approximately 150 amino acids in the midregion of Tax. Our observation that Tax mutants with deletions in this
region are expressed in the cytoplasm suggests a possible role for dimerization in nuclear localization. Three of our four midregion mutants of Tax that were nuclear excluded con-tained deletions within the Tax dimerization domain. This large domain reportedly contains three subdomains, I, II, and III, that are involved in Tax dimerization, but the
interdepen-FIG. 2. Localization of Tax deletion mutants. Confocal microscopy images of Tax mutants transiently expressed in HEK 293 cells, fixed, and counterstained with To-Pro-3⬘iodide (1:1,000). Tax expression is detected via the fluorescence of the GFP fusion (green). In the top left panel, an “untagged” version of Tax (HpX) was expressed and fixed as above but visualized via indirect immunofluorescence using a rabbit polyclonal anti-Tax primary antibody and a goat anti-rabbit Alexa Fluor 488 secondary antibody. For each protein examined, 50 expressing cells were observed. Shown is a representative of the expression pattern observed in⬎85% of cells.
VOL. 83, 2009 NOVEL Tax SPECKLED STRUCTURE LOCALIZATION SIGNAL 5343
on November 8, 2019 by guest
http://jvi.asm.org/
dence of these three subdomains for efficient dimerization has never been evaluated (5).
In our original mutational analysis, we created constructs that were each missing one of these subdomains of the Tax dimerization region. The⌬100–150 mutant is missing the first subdomain, the⌬151–202 mutant is missing the second sub-domain, and the⌬203–254 mutant is missing the third
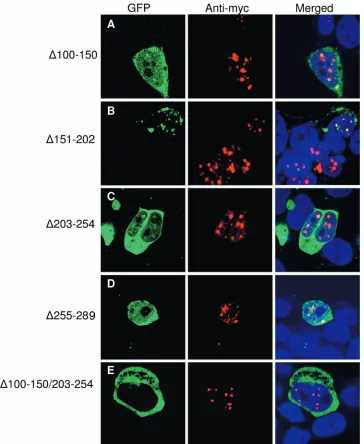
[image:6.585.108.474.65.583.2]subdo-main. In addition, the double mutant, the⌬100–150⌬203–254 mutant, is missing the first and third dimerization subdomains. As shown in Fig. 7, immunofluorescence confocal microscopy studies revealed that coexpression of full-length Tax was able to partially restore nuclear localization of those mutants miss-ing only one dimerization subdomain (Fig. 7A to D) but not of the mutant missing two dimerization subdomains (Fig. 7E).
FIG. 3. Colocalization of Tax deletion mutants with TSS. Shown are confocal microscopy images of fixed HEK 293 cells transiently expressing “nuclear” Tax mutants. Tax proteins (green) were observed via fluorescence of the GFP fusion. SC35 (red) was detected with mouse anti-SC35 primary antibody (1:5,000) and goat anti-mouse Alexa Fluor 594-conjugated secondary antibody (1:1,000). The nuclei were counterstained with To-Pro-3⬘iodide (1:1,000). The overlap (white) was displayed by merging the two images.
on November 8, 2019 by guest
http://jvi.asm.org/
Mutual complementation by coexpression of different single-subdomain mutants revealed that none of the mutants could be rescued for nuclear localization (data not shown). These re-sults suggest that the midregion Tax mutants are impaired for dimerization and that presentation of a dimer interface is a prerequisite to nuclear localization.
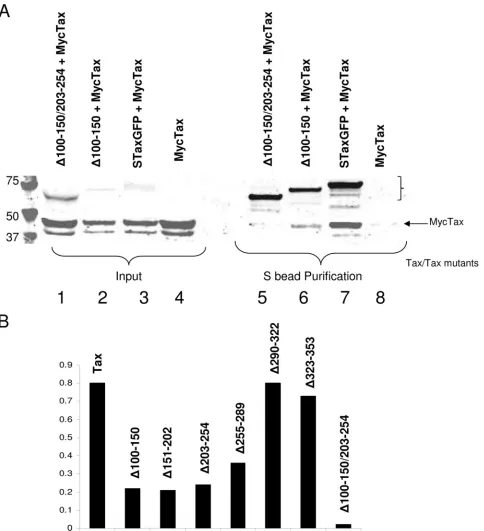
Dimerization subdomain Tax mutants are deficient for Tax binding.To directly test whether the subdomain mutants were deficient for dimerization, we coexpressed MycTax with STaxGFP or a Tax mutant missing one or two dimerization subdomains, the ⌬100–150 or ⌬100–150 ⌬203–254 mutants, respectively. We then assessed dimerization function by
puri-fication of STaxGFP, the⌬100–150 mutant, or the⌬100–150
⌬203–254 mutant, followed by immunoblot analysis for the presence of coprecipitated MycTax (Fig. 8A). Full-length STaxGFP was able to bind to MycTax, demonstrating dimer-ization between the two full-length proteins (Fig. 8A, lane 7). When the⌬100–150 mutant was tested, a measurable but re-duced level of MycTax was detected (Fig. 8A, lane 6), consis-tent with weak dimerization with full-length Tax. However, when the⌬100–150 ⌬203–254 mutant was examined, no evi-dence of dimerization with full-length Tax could be detected (Fig. 8A, lane 5). MycTax did not bind to the S beads alone (Fig. 8A, lane 8). We quantitated the dimerization by
deter-FIG. 4. Rescue of Tax NLS mutant by exogenous NLS tagging. An NLS sequence from SV40 Tag was added to all nucleus-excluded Tax mutants, and the constructs were transiently expressed in HEK 293 cells. Tax proteins (green) were observed via fluorescence of the GFP fusion. SC35 (red) was detected with mouse anti-SC35 primary antibody (1:5,000) and goat anti-mouse Alexa Fluor 594-conjugated secondary antibody (1:1,000). Nuclei were counterstained with To-Pro-3⬘iodide (1:1,000). The overlap (white) was displayed by merging the two images.
VOL. 83, 2009 NOVEL Tax SPECKLED STRUCTURE LOCALIZATION SIGNAL 5345
on November 8, 2019 by guest
http://jvi.asm.org/
[image:7.585.106.479.68.532.2]mining the ratio of MycTax that coprecipitated with STaxGFP or the STaxGFP-derived deletion mutant. When this analysis was performed, the single deletions between amino acids 100 to 289 resulted in similarly reduced dimerization compared to that of the wild-type protein (Fig. 8B). The double deletion that removes two dimerization subdomains resulted in a fur-ther reduction in binding. These results confirm that this re-gion is required for dimerization and that deletion of the dimerization subdomains has an additive effect.
Nuclear localization requires Tax dimerization activity.We have shown that deletion of Tax dimerization subdomains
im-pairs protein-protein interactions in a dimerization assay and results in cytoplasmic expression of Tax protein. Clearly, these results strongly support that dimerization is a prerequisite to nuclear localization of Tax. However, we wanted to determine if restoration of dimerization activity is sufficient to rescue nuclear localization of Tax. To accomplish this, we designed a Tax dimerization mutant protein that contains an inducible dimerization domain, STax(⌬100–150)-Fv-GFP. The inducible dimerization cassette is based on the binding domain of the human protein FK506 binding protein (FKBP) and its ability to bind to the immunosuppressive drugs FK506 and
rapamy-FIG. 5. Tax peptide comprising amino acid residues 50 to 75 is sufficient for targeting to TSS. HEK 293 cells transiently expressing STaxGFP, SGFP, or N-terminal regions of Tax fused to GFP were assayed for TSS formation as confirmed by colocalization with SC35. Tax proteins (green) were observed via fluorescence of the GFP fusion. SC35 (red) was detected with mouse anti-SC35 primary antibody (1:5,000) and goat anti-mouse Alexa Fluor 594-conjugated secondary antibody (1:1,000). The nuclei were counterstained with To-Pro-3⬘iodide (1:1,000). The peptide protein (green) and SC35 (red) and merged images (white) are also shown.
on November 8, 2019 by guest
http://jvi.asm.org/
[image:8.585.110.478.64.533.2]cin. Fv is a modified version of the FKBP binding domain containing a phenylalanine-to-valine substitution that in-creases the affinity of a rapamycin derivative, AP20187, for the Fv fusion protein by 1,000-fold over that for the wild-type protein. Thus, AP20187 is able to associate any two proteins containing the FKBP binding domain by inducing dimeriza-tion. The expression of STax(⌬100–150)-Fv-GFP in the ab-sence of the chemical dimerizer AP20187 resulted in cytoplas-mic localization of the protein. Upon the addition of increasing concentrations of the dimerizer, the Tax protein accumulated in the nucleus (Fig. 9A). The nuclear form of the protein dimer does not target TSS because the Fv region was inserted into the TSLS region using the unique ClaI site within the Tax open reading frame. Semiquantitative analyses of the percentages of Tax expression in the nucleus revealed dose-dependent re-sponses of from less than 1% to more than 40% nuclear ac-cumulation (Fig. 9B). This result clearly shows that dimeriza-tion funcdimeriza-tion is required for Tax nuclear localizadimeriza-tion.
DISCUSSION
Compartmentalization is a well-established mechanism for regulating protein function (14, 60). Clearly, this critical rela-tionship between cellular targeting and function goes beyond nuclear-versus-cytoplasmic accumulation. For instance, a for-midable number of nuclear proteins target specific subnuclear domains, many of which denote specific cellular functions (13,
35, 36). It is also understood that since localization is dynamic (for instance, proteins may exhibit compartmental “shuttling”), the stoichiometry is critical to overall protein function and an ideal target for protein regulation. In the case of the HTLV-1 Tax protein, localization is influenced by the presence of both NLS and NES sequences, as well as by the action of posttrans-lational modifications such as sumoylation and ubiquitylation (1, 11, 21, 23, 37, 53, 55). Our results from the current study provide an additional regulatory sequence, the TSLS, which allows for accumulation of Tax within the subnuclear structure TSS. We have further shown that dimerization of Tax is re-quired for overall nuclear accumulation of this important viral protein.
[image:9.585.106.477.71.341.2]Although nuclear localization is delineated from cytoplasmic localization by a clear nuclear membrane, subnuclear localiza-tion is largely dictated by interaclocaliza-tions with protein complexes present in the nucleus, with membership in these complexes defining the specific structure. All of the well-known sub-nuclear structures, represented by sub-nuclear speckles (36), paraspeckles (36), Cajal bodies (36), gems (20), and ND10/ PML bodies (4, 59), are dynamic structures that contain char-acteristic sets of nuclear proteins and protein complexes that reside in distinct subnuclear regions (46). Nearly all of these nuclear structures contain subpopulations of nuclear factors, but each is distinguished by the presence of a nuclear pro-tein(s) unique to each structure (36). One specific class of
FIG. 6. Complementation of Tax NLS and TSLS mutant proteins. (A) Confocal microscopy images of the expression of MycTax⌬30–52 (left panels) and⌬53–99 (middle panels) expressed alone or in combination (right panels). MycTax⌬30–52 (red) was detected with rabbit anti-Myc primary antibody at a dilution of 1:1,000 and Alex Fluor 594 goat anti-rabbit secondary antibody (1:1,000).⌬53–99 (green) was detected via fluorescence of the GFP fusion. The nuclei were counterstained with To-Pro-3⬘iodide (1:1,000). (B) Colocalization with SC35 of⌬30–52,⌬53–99, or⌬30–52 coexpressed with⌬53–99 (right panels). Tax proteins (green) were observed via fluorescence of the GFP fusion. SC35 (red) was detected with mouse anti-SC35 primary antibody (1:5,000) and goat anti-mouse Alexa Fluor 594-conjugated secondary antibody (1:1,000). The nuclei were counterstained with To-Pro-3⬘iodide (1:1,000).
VOL. 83, 2009 NOVEL Tax SPECKLED STRUCTURE LOCALIZATION SIGNAL 5347
on November 8, 2019 by guest
http://jvi.asm.org/
structures, called nuclear speckles, are interchromatin granule clusters that contain the pre-mRNA splicing machinery, in-cluding small nuclear ribonucleic proteins, non-small nuclear ribonucleic protein splicing factors, and spliceosome subunits such as SC35 (59). We previously observed that Tax localized to specific TSS, so named because of their overlap with nuclear speckles, colocalized with SC35, did not contain promyelocytic leukemia protein, and did not colocalize with nucleoli (3). However, TSS are distinct from nuclear speckles in that they contain various other nonsplicing cellular proteins. These pro-teins include DNA protein kinase, Chk2, 53BP1, and␥H2AX, proteins involved in DNA damage recognition and repair (15, 24, 26). The colocalization of Tax with cellular machinery for
transcription/splicing, DNA damage response, and checkpoint activation suggests that targeting to TSS is an important inte-grating event for the varied functions of Tax.
[image:10.585.112.476.67.511.2]Subnuclear protein targeting to nuclear speckles appears to be mediated by at least two separable trafficking signals: one for nuclear import (the NLS) and one for mediating inclusion into speckles (12, 28, 49, 54). As might be expected, the iden-tified domains are structurally variable and include examples such as ankyrin repeats in the IBL, FF domains in CA150, or interaction with RNA and are believed to represent modalities for protein-protein interactions. In our study, we found that the TSLS that is required for localization to TSS was physically distinct from the NLS and could function independently to
FIG. 7. Complementation of deletion mutants with full-length Tax. Full-length MycTax was coexpressed with the indicated Tax midregion mutants deficient for nuclear localization. Shown are single-subdomain mutants (A through D) and one double-subdomain mutant (E). Tax proteins (green) were observed via fluorescence of the GFP fusion. MycTax was observed by using mouse anti-Myc primary antibody (1:500) and goat anti-mouse Alexa Fluor 594-conjugated secondary antibody (1:1,000). Nuclei were counterstained with To-Pro-3⬘iodide (1:1,000).
on November 8, 2019 by guest
http://jvi.asm.org/
direct Tax to its specific intranuclear site. There are no remark-able features of the 25-amino-acid TSLS except that it is en-riched for proline residues, a characteristic shared by many nuclear speckle proteins, but it does not contain an
[image:11.585.53.537.67.598.2]arginine-serine (RS) motif common to targeting signals of splicing speckle components (8, 9, 17, 18, 27). This may reflect the unique nuclear address of TSS and the ability of Tax to form novel protein complexes.
FIG. 8. Midregion Tax mutants are deficient for dimerization. (A) MycTax was coexpressed with STaxGFP or Tax mutants missing one (⌬100–150) or two (⌬100–150,⌬203–254) dimerization subdomains. Dimerization of STaxGFP and Tax mutants with MycTax was assayed by S-bead purification of protein followed by immunoblot analysis for coprecipitated MycTax. Starting levels of each protein are shown in the left panel (input). Analyses of the isolated Tax complexes are shown in the right panel (S-bead purification). Protein complexes were normalized to levels of GFP expressed by Tax fusions (lanes 5, 6, and 7). Corresponding levels of coprecipitated MycTax are indicated. (B) Quantitation of the dimerization assay. Following the isolation of the Tax complexes shown in the right panel of panel A above, the ratios of MycTax and the test proteins were determined for relative quantitation of dimerization ability. The bar graph shows the percentages of MycTax relative to those of the isolated STaxGFP or mutant proteins as indicated (ratio).
VOL. 83, 2009 NOVEL Tax SPECKLED STRUCTURE LOCALIZATION SIGNAL 5349
on November 8, 2019 by guest
http://jvi.asm.org/
It is also important to point out that the 25-amino-acid sequence comprising the TSLS is sufficient for targeting to TSS when fused to either the native Tax NLS or a heterologous NLS. This implies that sumoylation of Tax is not a requirement for either nuclear localization or the formation of TSS. Likely, the role for sumoylation is in the context of the whole protein and may involve the removal of a block to entry or facilitate protein-protein interactions that enable nuclear accumulation. Such a model is reminiscent of that established for the tumor suppressor Wilms’ tumor gene (WT1) in which modification by
SUMO proteins is separable from nuclear speckle localization (57). As with Tax, although sumoylation is required for nuclear entry of WT1, an independent sequence is sufficient for tar-geting to nuclear speckles in the absence of a SUMO protein target. It may also be that sumoylation serves to mask the function of the NES, as has been suggested previously (37).
Our results also demonstrate that the dimerization of Tax is a necessary prerequisite for nuclear localization/accumulation. There are numerous examples of proteins that must dimerize or oligomerize prior to nuclear translocation, including the
FIG. 9. Induced dimerization of a dimerization-deficient Tax mutant restores nuclear localization. (A) Confocal microscopy images of cells expressing STax(⌬100–150)-Fv-GFP treated with 0 nM, 1 nM, 10 nM, or 100 nM of the chemical inducer of dimerization AP20187. Cells were fixed and counterstained with To-Pro-3⬘ iodide (1:1,000). STax(⌬100–150)-Fv-GFP (green) was visualized via fluorescence from the GFP fusion. Overlapping expression of stained nuclei and GFP fusion protein is shown (merged). (B) Quantitation of the nuclear accumulation of STax(⌬100– 150)-Fv-GFP with increasing concentrations of dimerizer. The amounts of nuclear fluorescence are expressed as percentages of total fluorescence.
on November 8, 2019 by guest
http://jvi.asm.org/
human cytomegalovirus protein ppUL44 and the cellular pro-tein p53 (2, 66). Other propro-teins, such as those of the AP-1 family, including c-Jun, JunD, JunB, and c-Fos, enter the nu-cleus as monomers but require heterodimerization in order to remain in the nucleus (40). In our studies, we were able to link the inability of Tax mutants to dimerize with a failure to ac-cumulate in the nucleus in spite of a competent NLS. In fact, the addition of a heterologous NLS failed to rescue this defect. The relationship between dimerization efficiency and nuclear accumulation is linear in that weakly dimerizing proteins were able to weakly accumulate in the nucleus. Specifically, we gen-erated a deletion of two subdomains within the Tax dimeriza-tion domain as well as subdomain mutadimeriza-tions. The single-subdomain Tax mutant retained a slight dimerization ability, whereas the Tax mutant in which the larger dimerization do-main is removed showed no dimerization. The rescue of single-subdomain dimerization mutants by wild-type Tax protein sup-ports a complementation phenotype, and our observation that none of the subdomain mutants could complement distal sub-domain mutants implies a requirement for a dimer interface. Finally, we showed that induction of dimerization in a previ-ously cytoplasm-restricted Tax dimerization subdomain mu-tant resulted in restored nuclear accumulation. Although our studies definitively link dimerization as a prerequisite to nu-clear accumulation, additional studies are required to deter-mine if this dimerization occurs in the cytoplasm as a prereq-uisite to nuclear entry or within the nucleus as a means of retaining the protein within its subnuclear address.
Previous studies on the dimerization of Tax have suggested that the N-terminal zinc finger domain (51) is important for Tax self association (5, 31). These findings were based on yeast two-hybrid assays where an N-terminal deletion of Tax failed to interact with wild-type Tax. In mammalian cells, as used in our system, deletion of the zinc finger region (⌬30-52) was rescued for nuclear localization by the coexpression of wild-type protein, demonstrating the ability to interact. Our func-tional complementation assay between the NLS mutant, the
⌬30-52 mutant, which is missing the zinc finger domain, and the TSLS mutant, the⌬50-75 mutant, demonstrated that the zinc finger domain is not required for dimerization, since a complementation phenotype requires interaction. Likewise, the fusion of a heterologous NLS to the⌬30-52 mutant was able to restore nuclear localization and subsequent TSS accu-mulation, providing further evidence that the zinc finger do-main is dispensable for Tax self association in our system.
Our results also allow for some inferences regarding the stoichiometry of Tax self association. The complementation assays between the NLS mutant and TSLS mutant suggest that at the minimum, a single copy of each, NLS or TSLS, is re-quired for nuclear accumulation of Tax in TSS. Thus, Tax exists at least in a one-to-one dimer state. In addition, the inability of the dimerization subdomain mutants to comple-ment distal subdomain mutants implies that at least two intact molecules of Tax are needed to form the required dimer in-terface. This finding is supported by findings in the induced dimerization assay. The design of the heterologous “dimer-izer” domain is such that only dimers, and not nonpaired higher-order oligomers (for instance, trimers), are formed upon the addition of the chemical dimerizer. Although in-duced dimerization was able to increase the nuclear
accumu-lation of the mutant protein, it did not result in a total resto-ration of nuclear accumulation to wild-type levels. This suggests the necessity of the presentation of specific dimer interfaces for normal Tax nuclear accumulation. Perhaps these specific interfaces mediate interaction with specific proteins. Interestingly, the ability of wild-type Tax to rescue dimeriza-tion subdomain mutants suggests the existence of a higher-order (⬎2) oligomeric structure. This conclusion derives from the reasoning that if dimer interfaces are required for nuclear accumulation and the mutants are incapable of forming inter-faces, then wild-type Tax must provide them. In order for this to occur, the rescued Tax complex must contain at least 2⫻ molar equivalents of the wild-type protein. Thus, it is likely that Tax is capable of forming a homotetrameric complex. Higher-ordered Tax oligomerization could provide for more-complex regulation of Tax functions. Oligomerization could generate new intermolecular interfaces to improve stability, control the accessibility and specificity of active sites, and increase the number of cellular binding partners for Tax. The complexity of such a model helps explain how a single protein displays such a wide range of activities.
ACKNOWLEDGMENT
This work was supported by Public Health Service grant CA76959 (to O.J.S.) from the National Cancer Institute.
REFERENCES
1.Alefantis, T., K. Barmak, E. W. Harhaj, C. Grant, and B. Wigdahl.2003. Characterization of a nuclear export signal within the human T cell leukemia virus type I transactivator protein Tax. J. Biol. Chem.278:21814–21822. 2.Alvisi, G., D. A. Jans, and A. Ripalti. 2006. Human cytomegalovirus
(HCMV) DNA polymerase processivity factor ppUL44 dimerizes in the cytosol before translocation to the nucleus. Biochemistry45:6866–6872. 3.Ariumi, Y., T. Ego, A. Kaida, M. Matsumoto, P. P. Pandolfi, and K.
Shimo-tohno.2003. Distinct nuclear body components, PML and SMRT, regulate the trans-acting function of HTLV-1 Tax oncoprotein. Oncogene22:1611–1619. 4.Ascoli, C. A., and G. G. Maul.1991. Identification of a novel nuclear domain.
J. Cell Biol.112:785–795.
5.Basbous, J., A. Bazarbachi, C. Granier, C. Devaux, and J. M. Mesnard.2003. The central region of human T-cell leukemia virus type 1 Tax protein contains distinct domains involved in subunit dimerization. J. Virol.77:13028–13035. 6.Bex, F., A. McDowall, A. Burny, and R. Gaynor.1997. The human T-cell
leukemia virus type 1 transactivator protein Tax colocalizes in unique nu-clear structures with NF-B proteins. J. Virol.71:3484–3497.
7.Bex, F., M. J. Yin, A. Burny, and R. B. Gaynor.1998. Differential transcrip-tional activation by human T-cell leukemia virus type 1 Tax mutants is mediated by distinct interactions with CREB binding protein and p300. Mol. Cell. Biol.18:2392–2405.
8.Bickmore, W. A., and H. G. Sutherland.2002. Addressing protein localiza-tion within the nucleus. EMBO J.21:1248–1254.
9.Birney, E., S. Kumar, and A. R. Krainer. 1993. Analysis of the RNA-recognition motif and RS and RGG domains: conservation in metazoan pre-mRNA splicing factors. Nucleic Acids Res.21:5803–5816.
10.Boxus, M., J. C. Twizere, S. Legros, J. F. Dewulf, R. Kettmann, and L. Willems.2008. The HTLV-1 Tax interactome. Retrovirology5:76. 11.Burton, M., C. D. Upadhyaya, B. Maier, T. J. Hope, and O. J. Semmes.2000.
Human T-cell leukemia virus type 1 Tax shuttles between functionally dis-crete subcellular targets. J. Virol.74:2351–2364.
12.Calado, A., and M. Carmo-Fonseca.2000. Localization of poly(A)-binding protein 2 (PABP2) in nuclear speckles is independent of import into the nucleus and requires binding to poly(A) RNA. J. Cell Sci.113:2309–2318. 13.Cardoso, M. C., and H. Leonhardt.1998. Protein targeting to subnuclear
higher order structures: a new level of regulation and coordination of nuclear processes. J. Cell. Biochem.70:222–230.
14.Chubb, J. R., and W. A. Bickmore.2003. Considering nuclear compartmen-talization in the light of nuclear dynamics. Cell112:403–406.
15.Durkin, S. S., X. Guo, K. A. Fryrear, V. T. Mihaylova, S. K. Gupta, S. M. Belgnaoui, A. Haoudi, G. Kupfer, and O. J. Semmes.2008. HTLV-1 Tax oncoprotein subverts the cellular DNA damage response via binding to DNA protein kinase. J. Biol. Chem.283:36311–36320.
16.Durkin, S. S., M. D. Ward, K. A. Fryrear, and O. J. Semmes.2006. Site-specific phosphorylation differentiates active from inactive forms of the human T-cell leukemia virus type 1 Tax oncoprotein. J. Biol. Chem.281:31705–31712.
VOL. 83, 2009 NOVEL Tax SPECKLED STRUCTURE LOCALIZATION SIGNAL 5351
on November 8, 2019 by guest
http://jvi.asm.org/
17.Eilbracht, J., and M. S. Schmidt-Zachmann.2001. Identification of a se-quence element directing a protein to nuclear speckles. Proc. Natl. Acad. Sci. USA98:3849–3854.
18.Fu, X.-D.1995. The superfamily of arginine/serine-rich splicing factors. RNA 1:663–680.
19.Fujii, M., H. Tsuchiya, T. Chuhjo, T. Akizawa, and M. Seiki.1992. Interac-tion of HTLV-1 Tax1 with p67SRF causes the aberrant inducInterac-tion of cellular immediate early genes through CArG boxes. Genes Dev.6:2066–2076. 20.Gall, J. G.2000. Cajal bodies: the first 100 years. Annu. Rev. Cell Dev. Biol.
16:273–300.
21.Gatza, M. L., T. Dayaram, and S. J. Marriott. 2007. Ubiquitination of HTLV-I Tax in response to DNA damage regulates nuclear complex forma-tion and nuclear export. Retrovirology4:95.
22.Gessain, A., F. Barin, J. C. Vernant, O. Gout, L. Maurs, A. Calender, and G. de The´.1985. Antibodies to human T-lymphotropic virus type-I in patients with tropical spastic paraparesis. Lancetii:407–410.
23.Gitlin, S. D., P. F. Lindholm, S. J. Marriott, and J. N. Brady.1991. Trans-dominant human T-cell lymphotropic virus type ITAX1mutant that fails to
localize to the nucleus. J. Virol.65:2612–2621.
24.Gupta, S. K., X. Guo, S. S. Durkin, K. F. Fryrear, M. D. Ward, and O. J. Semmes.2007. Human T-cell leukemia virus type 1 Tax oncoprotein pre-vents DNA damage-induced chromatin egress of hyperphosphorylated Chk2. J. Biol. Chem.282:29431–29440.
25.Haller, K., Y. Wu, E. Derow, I. Schmitt, K. T. Jeang, and R. Grassmann. 2002. Physical interaction of human T-cell leukemia virus type 1 Tax with cyclin-dependent kinase 4 stimulates the phosphorylation of retinoblastoma protein. Mol. Cell. Biol.22:3327–3338.
26.Haoudi, A., R. C. Daniels, E. Wong, G. Kupfer, and O. J. Semmes.2003. Human T-cell leukemia virus-I tax oncoprotein functionally targets a sub-nuclear complex involved in cellular DNA damage-response. J. Biol. Chem. 278:37736–37744.
27.Hedley, M. L., H. Amrein, and T. Maniatis.1995. An amino acid sequence motif sufficient for subnuclear localization of an arginine/serine-rich splicing factor. Proc. Natl. Acad. Sci. USA92:11524–11528.
28.Herrmann, A., K. Fleischer, H. Czajkowska, G. Mu¨ller-Newen, and W. Becker.2007. Characterization of cyclin L1 as an immobile component of the splicing factor compartment. FASEB J.21:3142–3152.
29.Jeang, K. T., R. Chiu, E. Santos, and S. J. Kim.1991. Induction of the HTLV-I LTR by Jun occurs through the Tax-responsive 21-bp elements. Virology181:218–227.
30.Jeang, K. T., C. Z. Giam, F. Majone, and M. Aboud.2004. Life, death, and tax: role of HTLV-I oncoprotein in genetic instability and cellular transfor-mation. J. Biol. Chem.279:31991–31994.
31.Jin, D.-Y., and K.-T. Jeang.1997. HTLV-I Tax self-association in optimal trans-activation function. Nucleic Acids Res.25:379–387.
32.Jin, D.-Y., F. Spencer, and K.-T. Jeang.1998. Human T cell leukemia virus type 1 oncoprotein Tax targets the human mitotic checkpoint protein MAD1. Cell93:81–91.
33.Kashanchi, F., and J. N. Brady.2005. Transcriptional and post-transcrip-tional gene regulation of HTLV-1. Oncogene24:5938–5951.
34.Kehn, K., L. Fuente Cde, K. Strouss, R. Berro, H. Jiang, J. Brady, R. Mahieux, A. Pumfery, M. E. Bottazzi, and F. Kashanchi.2005. The HTLV-I Tax oncoprotein targets the retinoblastoma protein for proteasomal degra-dation. Oncogene24:525–540.
35.Lamond, A. I., and J. E. Sleeman.2003. Nuclear substructure and dynamics. Curr. Biol.13:R825–R828.
36.Lamond, A. I., and D. L. Spector.2003. Nuclear speckles: a model for nuclear organelles. Nat. Rev. Mol. Cell Biol.4:605–612.
37.Lamsoul, I., J. Lodewick, S. Lebrun, R. Brasseur, A. Burny, R. B. Gaynor, and F. Bex.2005. Exclusive ubiquitination and sumoylation on overlapping lysine residues mediate NF-B activation by the human T-cell leukemia virus Tax oncoprotein. Mol. Cell. Biol.25:10391–10406.
38.Li, X. H., K. M. Murphy, K. T. Palka, R. M. Surabhi, and R. B. Gaynor. 1999. The human T-cell leukemia virus type-1 Tax protein regulates the activity of the IB kinase complex. J. Biol. Chem.274:34417–34424. 39.Liu, B., S. Hong, Z. Tang, H. Yu, and C. Z. Giam.2005. HTLV-I Tax directly
binds the Cdc20-associated anaphase-promoting complex and activates it ahead of schedule. Proc. Natl. Acad. Sci. USA102:63–68.
40.Malnou, C. E., T. Salem, F. Brockly, H. Wodrich, M. Piechaczyk, and I. Jariel-Encontre.2007. Heterodimerization with Jun family members regu-lates c-Fos nucleocytoplasmic traffic. J. Biol. Chem.282:31046–31059. 41.Marriott, S. J., and O. J. Semmes.2005. Impact of HTLV-I Tax on cell cycle
progression and the cellular DNA damage repair response. Oncogene24:5986– 5995.
42.Mesnard, J. M., and C. Devaux.1999. Multiple control levels of cell prolifera-tion by human T-cell leukemia virus type 1 Tax protein. Virology257:277–284. 43.Nicot, C., F. Tie, and C. Z. Giam.1998. Cytoplasmic forms of human T-cell leukemia virus type 1 Tax induce NF-B activation. J. Virol.72:6777–6784. 44.Osame, M., K. Usuku, S. Izumo, N. Ijichi, H. Amitani, A. Igata, M. Matsu-moto, and M. Tara.1986. HTLV-I associated myelopathy, a new clinical entity. Lanceti:1031–1032.
45.Park, H. U., J. H. Jeong, J. H. Chung, and J. N. Brady.2004. Human T-cell leukemia virus type 1 Tax interacts with Chk1 and attenuates DNA-damage induced G2arrest mediated by Chk1. Oncogene23:4966–4974.
46.Phair, R. D., and T. Misteli.2000. High mobility of proteins in the mam-malian cell nucleus. Nature404:604–609.
47.Poiesz, B. J., F. W. Ruscetti, A. F. Gazdar, P. A. Bunn, J. D. Minna, and R. C. Gallo.1980. Detection and isolation of type C retrovirus particles from fresh and cultured lymphocytes of a patient with cutaneous T-cell lymphoma. Proc. Natl. Acad. Sci. USA77:7415–7419.
48.Ramírez, J. A., and J. K. Nyborg. 2007. Molecular characterization of HTLV-1 Tax interaction with the KIX domain of CBP/p300. J. Mol. Biol. 372:958–969.
49.Sa´nchez-A´lvarez, M., A. C. Goldstrohm, M. A. Garcia-Blanco, and C. Sun˜e´. 2006. Human transcription elongation factor CA150 localizes to splicing factor-rich nuclear speckles and assembles transcription and splicing com-ponents into complexes through its amino and carboxyl regions. Mol. Cell. Biol.26:4998–5014.
50.Semmes, O. J., J. F. Barret, C. V. Dang, and K.-T. Jeang.1996. Human T-cell leukemia virus type I Tax masks c-Myc function through a cAMP-dependent pathway. J. Biol. Chem.271:9730–9738.
51.Semmes, O. J., and K. T. Jeang.1992. HTLV-I Tax is a zinc-binding protein: role of zinc in Tax structure and function. Virology188:754–764. 52.Semmes, O. J., and K. T. Jeang.1996. Localization of human T-cell leukemia
virus type 1 tax to subnuclear compartments that overlap with interchroma-tin speckles. J. Virol.70:6347–6357.
53.Semmes, O. J., and K.-T. Jeang.1992. Mutational analysis of human T-cell leukemia virus type I Tax: regions necessary for function determined with 47 mutant proteins. J. Virol.66:7183–7192.
54.Semple, J. I., S. E. Brown, C. M. Sanderson, and R. D. Campbell.2002. A distinct bipartite motif is required for the localization of inhibitoryB-like (IBL) protein to nuclear speckles. Biochem. J.361:489–496.
55.Smith, M. R., and W. C. Greene.1992. Characterization of a novel nuclear local-ization signal in the HTLV-I tax transactivator protein. Virology187:316–320. 56.Smith, M. R., and W. C. Greene.1990. Identification of HTLV-Itax
trans-activator mutants exhibiting novel transcriptional phenotypes. Genes Dev. 4:1875–1885.
57.Smolen, G. A., M. T. Vassileva, J. Wells, M. J. Matunis, and D. A. Haber. 2004. SUMO-1 modification of the Wilms’ tumor suppressor WT1. Cancer Res.64:7846–7851.
58.Sorokin, A. V., E. R. Kim, and L. P. Ovchinnikov.2007. Nucleocytoplasmic transport of proteins. Biochemistry (Moscow)72:1439–1457.
59.Spector, D. L.2001. Nuclear domains. J. Cell Sci.114:2891–2893. 60.Strouboulis, J., and A. P. Wolffe.1996. Functional compartmentalization of
the nucleus. J. Cell Sci.109:1991–2000.
61.Sun, S. C., and D. W. Ballard.1999. Persistent activation of NF-B by the Tax transforming protein of HTLV-1: hijacking cellular IB kinases. Onco-gene18:6948–6958.
62.Sun, S. C., and S. Yamaoka.2005. Activation of NF-B by HTLV-I and implications for cell transformation. Oncogene24:5952–5964.
63.Takatsuki, K.2005. Discovery of adult T-cell leukemia. Retrovirology2:16. 64.Terry, L. J., E. B. Shows, and S. R. Wente.2007. Crossing the nuclear envelope: hierarchical regulation of nucleocytoplasmic transport. Science 318:1412–1416.
65.Tie, F., N. Adya, W. C. Greene, and C.-Z. Giam.1996. Interaction of the human T-lymphotropic virus type 1 Tax dimer with CREB and the viral 21-base-pair repeat. J. Virol.70:8368–8374.
66.Trostel, S. Y., D. L. Sackett, and T. Fojo.2006. Oligomerization of p53 precedes its association with dynein and nuclear accumulation. Cell Cycle 5:2253–2259.
67.Tsuji, T., N. Sheehy, V. W. Gautier, H. Hayakawa, H. Sawa, and W. W. Hall. 2007. The nuclear import of the human T lymphotropic virus type I (HTLV-1) tax protein is carrier- and energy-independent. J. Biol. Chem. 282:13875–13883.
68.Yoshida, M., I. Miyoshi, and Y. Hinuma.1982. Isolation and characterization of retrovirus from cell lines of human adult T-cell leukemia and its implica-tion in the disease. Proc. Natl. Acad. Sci. USA79:2031–2035.