Vol. 45, No. 1 JOURNALOFVIROLOGY,Jan. 1983,p.251-263
0022-538X/83/010251-13$02.00/0
Polypeptide Structure and Encoding Location of the
Adenovirus Serotype
2 Late,
Nonstructural
33K
Protein
ELIZABETH A. OOSTEROM-DRAGONt AND CARL W. ANDERSON* BiologyDepartment,Brookhaven NationalLaboratory, Upton,New York11973
Received 19July1982/Accepted4October 1982
Radiochemical microsequence analysis of selected tryptic peptides of the
adenovirustype233K nonstructuralprotein has revealed the precise region ofthe
genomicnucleotide sequencethat encodes this protein. The initiation codon for
the 33Kprotein lies 606 nucleotidestotheright of theEcoRI restriction site at 70.7
mapunits and 281 nucleotidestotheleft of the postulatedcarboxyterminalcodon
of the adenovirus 100K protein. The coding regions for these twoproteins thus
overlap; however, the 33Kprotein is derived fromthe +1 frame with respect to
the postulated 100K reading frame. Our results contradict an earlierpublished
report suggesting that these two proteins share extensive amino acid sequence
homology (N. Axelrod, Virology 87:366-383, 1978). The published nucleotide
sequence of the Ad2 EcoRI-Ffragment(70.7 to 75.9mapunits)cannot
accomo-dateinasingle reading frame thepeptide sequencesof the 33K protein that we
have determined. Sequence analysis of DNA fragments derived from virus has
confirmedthepublishednucleotide sequence in all criticalregionswithrespect to
thecoding region for the33Kprotein. Consequently,ourdataareonly consistent
with the existence of an mRNA splice within the coding region for 33K.
Consensus donor and acceptor splice sequences have been located that would
predict the removal of 202 nucleotides from the transcripts for the 33Kprotein.
Removal of these nucleotides would explain thestructureofapeptidethat cannot
otherwise be directly encoded by the EcoRI-F fragment. Identification of the
precise splice points by peptide sequencing has permitted a prediction of the
complete amino acidsequence for the 33Kprotein.
Adenoviruses havenonenveloped, icosahedral
virions composed of a linear double-stranded
DNA ofabout 23 x 106 Mr encased in a shell
constructed from 10 to 12 different
virus-en-coded proteins (29). In addition to these
struc-turalproteins,lateviral transcriptsarebelieved
to encode at least three nonstructural proteins
thatare believed tofunction in virion assembly
and maturation. A maturation proteinase
activi-tyhasbeen described (9) thatprocesses atleast
four virion polypeptides andthatis believed to
be encoded within late mRNA region L3 (see
Fig. 2) (1, 17, 21), but the polypeptide
corre-sponding to this activity has not yet been
ob-served. Twononstructural proteins are
synthe-sized in large amounts late afterinfection from
mRNAs produced from late region L4. The
100,000-molecular-weight(100K)polypeptide is
believedtofunction in hexoncapsmer assembly
andtransport(14-16,26, 27).A33Kpolypeptide
tPresentaddress:CODON, Brisbane,CA 94005.
is also encodedby this region, but its function is
notknown, and its possible relationship to the
100Kproteinsequencehasbeensuggested.
Be-fore functions can be definitively ascribed to
either the 100Kor33Kpolypeptides, their
rela-tionship to each other and to their
genome-encoding nucleotide sequences must be
deter-mined.
Althoughabundantatlate times afterinfection
of HeLacells, the 33Kproteinwasoverlooked
byearly studies ofadenovirus-specific proteins
(4, 36),probably because of its low methionine
content. Thefirstdescription ofaca. 33,000Mr
polypeptide specific to extracts of
adenovirus-infected cellswasby Russell and Blair (31), who
described amajorvirus-specific phosphorylated
polypeptide of39,000 Mr found in the nuclei of
adenovirus 5-infected cells late after infection. This componentwassubsequently observedby
Levinsonetal.(22) andby Axelrod (6). Axelrod
compared the tryptic peptides from the three
major adenovirus-specific nonstructural
phos-251
on November 10, 2019 by guest
http://jvi.asm.org/
252 AND ANDERSON
phoproteins, the early 72,000-Mr DNA-binding
protein, the lOOK protein, and the 33K protein
and concluded that the lOOK protein and the
33K protein wereclosely related in amino acid
sequence. Lewiset al. (23) mapped by in vitro translation of hybridization-selected adenovirus
mRNA a 38,000-Mr polypeptide to the EcoRI-F
(70.7 to75.9 mapunits [m.u.]) and -D (75.9 to
83.4m.u.)restriction fragmentsofadenovirus2. But notuntil the much later
hybridization-trans-lation studies of Ricciardietal. (30)andMilleret
al.(25)wasthe33Kpolypeptideshown to be an
independently codedadenovirus product. Even
these studies didnotshow that the in vivo and in
vitro 33K products were necessarily related in sequence.
Recently Gambke and Deppert (14, 15)
pre-pared antiserum to purified adenovirus 2 33K
protein and found that thisserumdid not
cross-react with the lOOK protein. This result led
Gambke and Deppert to reexamine the
conclu-sionof Axelrod that theseproteinswereclosely
related; their own tryptic mapping studies
showed that theseproteinswereprobably
unre-latedinaminoacidsequence. Using their
antise-rum we have shown that the in vitro product
mappedtothe EcoRI-Ffragment of the
adenovi-rus2 genomeis indeed the sameasthe in vivo
33K productdescribed byAxelrod, Gambke and
Deppert, and others. Radiochemical
microse-quenceanalysis of the major tryptic peptides of
33Kprotein isolated from adenovirus 2-infected
HeLa cells has allowed us to predict the
com-plete amino acid sequence of this protein by
comparing partial peptide sequences with the
knownnucleotidesequenceof the EcoRI-F
frag-ment (13). Surprisingly, this analysis showed
that the 33K protein is encoded by an mRNA
that is spliced within the coding region. This
result explains why the 33K protein could not
previously be relatedcorrectlytothenucleotide
sequence.
MATERIALSANDMETHODS
Cells and virus.TheoriginsofourHeLacells and of serotype 2adenovirus stocks and the propagationof these have been described previously (2, 32). The adenovirus stock used fortheseexperimentsis direct-ly related to those usedby the Cold Spring Harbor Laboratory for DNA sequence studies(38). Adenovi-rus DNA was prepared from purified virions as de-scribedpreviously(32).
Purification of radioactively labeled 33K protein. Nearly confluent 90-mm culture dishes ofHeLacells wereinfected with 250PFUof adenovirus per cell.At 24 to26 hafterinfection, the infected cultureswere
placed into medium containing the appropriate 3H-amino acid(1 mCi per dish) or [35S]methionine (250
,uCiperdish). After incubationat37°C for4 to 6h, the radiolabeled cultures were fed with 5 ml of normal Dulbecco modifiedEaglemediumsupplementedwith 2% calfserumperdish,and incubationwascontinued
until approximately 48 h after infection. Harvested cultures were washed with ice-cold phosphate-buff-ered saline and then with0.01 MTris-hydrochloride (pH 7)-0.5 M NaCl-0.002 M MgCl2 (TSM buffer). Aftersuspension in 1.0 ml of ice-cold TSM buffer per dish, thecultureswerelysedby the addition of Noni-det P-40 to0.5% and phenylmethylsulfonyl fluorideto 300,ug/ml.Thenuclearpellet, obtained by centrifuga-tion, was washed once inTSM buffer containing0.5% Nonidet P-40 suspended in sodium dodecyl sulfate (SDS) sample buffer (4), and heated for 5 min in a boiling-water bath. Nuclearextracts werefractionated byelectrophoresis through slab-type SDS-polyacryl-amidegels (4). Proteinswerelocated bystaining with Coomassie brilliant blueorby autoradiographyof the dried gels. A dried gel slice corresponding to the position of the 33K protein was excised and eluted electrophoreticallywithanISCOsampleconcentrator
(3,33).
Digestion with trypsin and peptide fractionation by reverse-phase(C18) high-pressure liquid chromatogra-phy. Samples to be digested with trypsin were first extracted with acetone-acetic acid-triethanolamine (90:5:5)threetimes andacetonetwicetoremoveSDS (18). After the addition of 1 mg of apomyoglobin,
samples wereperformicacid oxidized (4) and
lyophi-lized. For digestion with trypsin, samples were dis-solved in 0.9 ml ofwaterand 0.1 mlof1MNH4HCO3; 10 ,ug oftosylamido 2-phenylethyl chloromethyl ke-tone-treated trypsin (Worthington Diagnostics, Free-hold, N.J.)wasadded.After 2 h of incubation at 37°C, asecond 10 ,ug oftrypsinwasadded, and incubation wascontinued foratotal of 6 h. Samples were then frozen andlyophilized.
Fractionation by reverse-phase chromatography wasperformed byusinga250-by 4.6-mm,10-p.m RP-18 column (E. M. Reagents, Inc.) equilibrated with 95%buffer A(10 mMammonium acetate, pH 5.4) and 5% buffer B (100% acetonitrile). Samples were dis-solved in 0.3 mlof20%formic acid andcentrifugedat 12,000 x g for 15 min before application to the column. The columnwasdevelopedwitha discontinu-ouslineargradientof acetonitrile(seeFig.3)at aflow rateof 2.0ml/min; 1.0-mlfractionswerecollected. The position ofapomyoglobin peptideswas monitoredat 214 nm; these positions were used to facilitate the alignmentofradioactivepeptideprofilesfrom separate experiments. In general, separations varied by less than +1 fraction. A portion of each fraction (0.05to 0.15ml) wasassayed forradioactivity with 3.5 mlof scintillation fluid (Aquasol; New England Nuclear Corp., Boston, Mass.). Appropriate fractions were pooled,1mgofapomyoglobinwasadded, and pooled fractionswerelyophilizedbefore amino acid sequence analysis.
Amino-terminal sequence analysis. Radiochemical microsequenceanalysis wasperformed witha Beck-man 890 C sequencer and a 0.1 M Quadrol buffer programasdescribedpreviously(3, 5, 33).Polybrene (3 mg) was included with all peptide samples. Se-quencer performance was routinely monitored by analysisof carrierapomyoglobin.
DNAsequenceanalysis. DNA sequenceanalysiswas performed bythe method of Maxamand Gilbert(12, 24). Appropriate restriction enzyme-generated frag-ments werelabeled with[32P]ATPandpolynucleotide kinase.
on November 10, 2019 by guest
http://jvi.asm.org/
ADENOVIRUS 2 33K PROTEIN 253 Immunoprecipitation. Immune complexes were
ad-sorbedtoFormalin-fixedStaphylococcusaureuscells
(The Enzyme Center, Boston, Mass.) and collected by centrifugation (19). Complexeswerereleasedby
boil-ing in 2% SDS-0.1 Mdithiothreitol sample buffer as
previously described(26).
RESULTS
Identification of the adenovirus 33K protein. The adenovirus 33K protein isamajor nonstruc-tural virus polypeptide produced at late times afterinfection; itcanreadily beidentified in the
pattern of Coomassie brilliant blue-stained bandsafterSDS-polyacrylamidegel electropho-resisof infected cell nuclearextracts(Fig. 1A). To confirm the identity of the 33K protein,
radioactively labeled nuclearextracts were
im-munoprecipitated with antiserum specific for the adenovirus 33K protein (the kind gift of W. Deppert), and the precipitateswereanalyzed by
SDS-polyacrylamidegelelectrophoresis. Figure
1B shows that theonly product
immunoprecipi-tated from [35S]methionine-labeled or3 P04-la-beled nuclear extracts of infected cells is a
product of ca. 33,000 Mr that corresponds in
mobilitytothemajor nuclearcomponent identi-fiedasthe33Kprotein in Fig. 1A. Figure 1B also shows that theelectrophoreticallypurified prod-uct used for the peptide and sequence analysis described here is radiochemically pureand can
be immunoprecipitated by 33K protein-specific
antiserum.
Invitro 33Kproduct labeled with
[35S]methio-ninewasprepared by translation of RNA
select-edby hybridizationtothe EcoRI-F(70.7to75.9 m.u.) orHindIII-H (73.6to79.9m.u.) fragment ofadenovirus 2 DNA(28, 30). Among the prod-uctsfrom thesetranslationswas amajor
compo-nent withamobilityby SDS-polyacrylamide gel
electrophoresis essentially identical to that of 33K protein from infected cells (data not shown). This component was excised from a
preparative SDS-polyacrylamide gel, digested with trypsin, and compared with similarly
pre-pared 33K protein from infected cells by
re-verse-phase liquid chromatography (data not shown). Both preparations had single major
me-thionine-containingpeptides that eluted from the
column atidentical positions. Subsequent stud-ieshave shown that this elution position
corre-spondstothat ofthesingle methionine-contain-ing peptide predicted to be present in the 33K protein (see below and Table 1). Thus, the in vivo 33K product must be encoded within the region from 70 to 80 m.u. on the adenovirus 2 genome(Fig. 2).
Inspection of the nucleotide sequence
corre-sponding to late mRNA within the EcoRl-F fragment of adenovirus 2 (13) reveals four mod-erately long open reading frames that could
potentially encode the 33K protein. The first
open reading frame begins at the left-hand
boundary of the EcoRI-Ffragment (70.7m.u.),
is 296 codons long, contains one methionine
codon, and is believed to encode the
carboxy-terminal portion of the adenovirus 100K late,
nonstructural protein (13). The second open
frame begins at nucleotide 307 (N307; to the
right of70.7), is 295 codons long, overlaps the
first openframe, and contains four methionine
codons. The thirdopenframe begins atN1006,
partially overlaps the second open frame, and
containsnomethionine codons. The fourthopen
frame begins at N1519 and has been found to
encode the precursor to virioncomponent VIII
(Oosterom-Dragon and Anderson, unpublished data).
33K protein has ablocked amino terminus.The
33Kprotein is knownto map tothe leftof virion
component VIII (25). The five methionine
co-dons located in the firsttwoopenreading frames
are each followed within 20 codons by one or
more leucine oralaninecodons. Consequently,
if the amino terminus of the 33K protein were
notblocked, direct amino-terminal
radiochemi-cal sequence analysis of [3H]leucine- or
[3H]al-anine-labeled 33K protein wouldyield
radioac-tivity inacharacteristicpattemof residues that
wouldidentify theencodinglocation of the
ami-noterminus of the protein. Several such
experi-ments were performed with [35S]methionine-,
[3H]leucine-, or[3H]alanine-labeled33Kprotein
isolated as described above, and no significant
releaseofradioactivitywasobtainedfromanyof
the first 20 Edman cycles. Analysis of carrier
apomyoglogin included with the radioactive
samples assuredusthatthesequencer was
func-tioningproperly. Since wehaverarely failedto
obtain sequenceinformationbythis methodfor
proteins without blocked amino termini (3, 5,
33),weconcludethatthe 33Kprotein,likemost
of the latevirionproteins, hasa blockedamino
terminus that precludes direct sequence
deter-mination.
Sequence analysis of 33K protein tryptic
pep-tides. Adenovirus 2 33K
?rotein,
labeledsepa-rately with different H-amino acids or
[35S]methionine,
was purified bygelelectropho-resis, performic acid oxidized,anddigested with
trypsin as described above. The 20o formic
acid-soluble peptides wereappliedto a
reverse-phasecolumn forfractionationbyhigh-pressure
liquid chromatography. Examples of the
pat-terns ofradioactivetryptic peptides obtainedare
givenin Fig. 3. Eachdifferent amino acid label
producedauniquepattern of well-resolved
pep-tides; some peptides appeared to contain only
one of the radioactive amino acids used, and
othersweredetectedby several different amino
acid labels. Appropriate fractions from
individ-VOL.45, 1983
on November 10, 2019 by guest
http://jvi.asm.org/
C C ...'C
_ N) .
4..
...i .-s X
-Z
()>(" illT
Y),
)3
mzi (. /z (
.-r. CDr Cl)
DBP-33:K _
. 4W2 *
8 9
[image:4.489.51.446.71.419.2]_
FIG. 1. Identificationof the adenovirus33Kprotein.(A)Coomassiebrilliant blue R-250-stained 17.5% SDS-polyacrylamide geluponwhichwereelectrophoresedthefollowing: 1,nuclearextractof mock-infectedHeLa
cells; 2, nuclearextractofadenovirus 2-infectedHeLacellsharvested32h after infection; 3, the corresponding cytoplasmicextractofadenovirus 2-infectedHeLacells. Thepositionoftheadenovirus33Kproteinaswellas
thepositions ofseveral other viral proteinsareindicatedatthe left of the figure. A cellular protein withamobility
similarto theadenovirus33Kprotein is found in nuclear extractsof uninfected HeLa cells, butthis cellular protein isnotrecognized by antibodytotheviral33Kprotein.Cellular protein synthesis wouldhave been largely
inhibited atthetimeinfected cellswerelabeledfor the preparationof radioactive33Kprotein.(B)
Autoradio-gramofanSDS-polyacrylamide geluponwhichwerefractionated immunoprecipitates obtained with antibody
specific for the 33K protein. The samples were as follows: 1, [j5S]methionine-labeled nuclear extract of adenovirus2-infected HeLa cellslabeledca.26to32hafter infection;2,immunoprecipitate of the [I5S]methio-nine-labelednuclearextractobtained with normal rabbitserum;3,asin lane2,butobtained withserumspecific
fortheadenovirus33Kprotein; 4, nuclearextractof adenovirus 2-infectedHeLacetlslabeled with32Pi26to32h afterinfection;5,immunoprecipitateof the
32Pi-labeled
nuclearextractwithnormalrabbitserum;6,asin lane5,butwith antiserumspecific for the adenovirus33Kprotein (Fig. 3); 8, immunoprecipitate of [3H]alanine33K
protein withnormal rabbit serum; asin lane 8, but withantiserum specific for the adenovirus 33Kprotein. Immune complexes were collected with Formalin-fixed S. aureus cells as previously described (26). The
polyacrylamide gel wastreated with En3Hance(NewEngland NuclearCorp.) before fluorographyonKodak
AXR-5film.
ualexperimentswerepooled andlyophilizedfor
subsequentsequence analysis.
Individual peptides were applied tothe spin-ning cup sequencer for automated sequential
Edmandegradation. This analysis gives the
po-sition(s) of the radioactive amino acid with
re-specttotheamino-terminaltrypsin cleavage site
that produced the peptide. Examples of the analysis of several 33Kpeptidesaregiven in Fig.
4and 5. Thepatternsof[3H]leucine and [3H]pro-line released from peptides designated L12 and P10 are given in Fig. 4A and B, respectively.
Thesetwopeptides elutedatthe sameposition
during chromatography. Leucine was found in A
Hexon-100K
-D8P"
33K
P-f
-
p-Histones
Ii
VI
on November 10, 2019 by guest
http://jvi.asm.org/
VOL.45, 1983 ADENOVIRUS 2 33K PROTEIN 255
-fo3- 3
z
_m.
oF
=V
r
(AnE
n.O
-,;
r
m
O.
r'o-o0
9:
0.
CD
n
CD
0e
a
CO)
t-i
w w
;F
la
a,
0
00 08
r
<zz4
zz
z0
>
~
> > >vc 00
r
- W.
~->c::>r:>3r S :> :> ^3 > e
~>
> >3rOcr @t
@
>
>
>3
<3
>> 0tTl
Cr
,r
0c>
co
rs
IQt@o
oc@~ ~~(
crsH
tTi In >>ev@
wr>
0r
r
0>->
>>
Hr
>@
la
r-
E' 0c
>
@)
c-
ar>zC)0 >>> CQ)
co % -=r w ro P q cA
>~~~~v
>r
> D~~~~>
> > 0 03
> >> X
w> r>
3>r
2T
ro 0-4 cn <> > cnC
tTi~~r o
t-C: m
~
~~
>3 3> Q> ~ > >.q
> co
r
Coco~ co O q
;F
la
o0 .0
c.
0 0 a0CL
o-t #._.
3 5 .* 3 >
3 o o
_. n
3 o p
O r ^
t v _.
3
q cn
° - £
C U Qq
. r o
O o 3
3 cn. sL
. _
F3 O- °
o O t
r _, m
00 r W _. r .
O . t
3 ;
cn o_
1ON
0-oC
on November 10, 2019 by guest
http://jvi.asm.org/
V 23K
ma m P-VII P-VI 1I IOOK 33KP-VIII
LATE
IV J PROTEINS
L I L2 L3 L4 E3 L5
- 3
--O -- -It
- - - - M*
- -~~~~~- -
-}LATE mRNAs
L A Ba IFT D I E C 3
0 10 20 30 40 50 60 70' 80 90 100
E2A(DBP), E4
E28 Q 31 I
IVa2
70.7
IOOK
11
i
ItIII
I 4I I I80
400
75.9
33K
tL___
I
___-1200 1600
[image:6.489.67.433.62.329.2]DBP I St LEADER
FIG. 2. Mapof the adenovirus 2genome.The adenovirus 2genomeisrepresented bytwothinparallellinesin
themiddle ofthepicture. Mapunits aremarked below these lines;the positionsof EcoRIcleavagesitesare
indicated by solid trianglesabove theselines;the letterdesignationofthe EcoRIfragmentsisgivenbetweenthe lines. Lines above and below the genome representation indicate RNA transcripts or mRNAs; promoter
positionsareindicatedbyabracket. Individual late mRNAbodysequencesfrom the5 lateregionsLi throughL5
areindicated bythick lines(10).Theapproximatelocationencodingseveralproteinsis indicatednearthetopof
thefigure.The lowerpartof thefigure showsamapofjustthe EcoRI-Ffragment (70.7to75.9m.u.).Theregions encoding the 100K protein (13),the 33Kprotein (this manuscript), andtheprecursortovirioncomponentVIII
(Oosterom-DragonandAnderson,unpublished data)areindicated.The exactlocationof the amino terminusof
P-VIII hasnotbeen determined. Solid diamonds indicate thepositionsofHpaIIrestrictionsites;theopencircle designatesthepositionof theHindlllrestriction site at72.8m.u. Numbersalongthe bottom oftheEcoRl-F fragment representation indicate distance in nucleotides fromthe EcoRI site at 70.7m.u.
amino acid positions 1 and 3 of this peptide, whereas prolinewasfound inpositions 4, 5, 6, 7, and 8. Inspection of the EcoRI-F nucleotide
sequence revealedonly one location that could encode a peptide with this pattern of leucines
and prolines. The location begins 624 nucleo-tides fromthe EcoRI-Fcleavagesiteat70.7m.u.
in thesecond openreading frame. Immediately preceding the leucine codon at N624 are three
lysine codons,acondition consistentwith
cleav-age by trypsin at this location in the protein. Similar analysis ofthe peptide A7-P7 (Fig. 4C andD) shows itto beuniquelyencoded by the
sequence beginning at N813, also within the secondopenreadingframe.These resultsprove
thataportion of the 33Kproteinisencoded by the open reading frame between N307 and N1190. Since the lOOKprotein is believedtobe encodedbythe firstopenreading frame (13, 25),
this result strongly suggests that the 33K and lOOK proteins cannot be directly related by amino acid sequence. To confirm this
expecta-tion, we prepared [3H]leucine-labeled lOOK
trypticpeptides and chromatographed them
un-derconditions identicaltothose used for
separa-tion of the 33Ktryptic peptides. No significant coincidence of radioactiveprofileswasobserved (datanotshown).
Partial sequenceanalysis of the four peptides
for which dataarepresented in Fig. 5 revealed surprisingly that these peptidesare allencoded in the thirdopenreading frame. PeptideP4-V3is encoded beginning atN1258, peptide A3 is
en-coded beginning at N1219, peptide F8 is
en-codedbeginningatN1303,andpeptideL5-A5 is encodedbeginningatN1435(Table 1).The data presented in Table 1, therefore, prove thatthe 33Kprotein is also partiallyencodedbythethird
open readingframe.
Peptide A6 was found to have alanines at positions1,2, 5,7, 8, 9, 10, 11, 12, 13, 15,and16
(Fig. 4). This pattern of 11 alanines cannot be encoded by a contiguous sequence within the EcoRI-Ffragment;however,the last 10 alanines
IX EIS *
EIA >
r
I
I
on November 10, 2019 by guest
http://jvi.asm.org/
ADENOVIRUS 2 33K PROTEIN 257
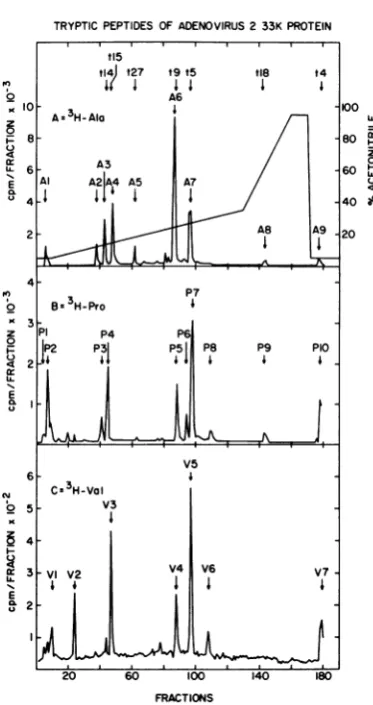
TRYPTICPEPTIDES OF ADENOVIRUS 2 33K PROTEIN
0
C-)
E
C-)
4
E
a:
E
N.
0
I?
:
E
[image:7.489.52.239.69.422.2]FRACTIONS
FIG. 3. High-pressure reverse-phase, liquid chro-matographyof thetryptic peptidesof theadenovirus 2 33K protein. SDS-polyacrylamide gel-purified 33K protein,labeled in vivo with[3H]alanine(A),
[3H]pro-line (B), [3H]valine (C), or several other amino acid isotopes ([3H]isoleucine, [3H]leucine, [3H]lysine,
[35S]methionine, [3H]phenylalanine, or [3H]tyrosine;
datanotshown)wasdigestedwithtrypsinasdescribed in the text, and thetrypticdigestswerefractionatedby reverse-phase high-pressure liquid chromatography. The programmed gradientproffle used for all experi-ments is shown in (A); the flowrate was2.0ml/min,
and 1.0-ml (30-s) fractions were collected. For the
threeexperiments shown, 100 Fl of each fractionwas countedwith 3.5 ml ofAquasolpersampleinaSearle Mark IIIscintillation counter. The countsper minute from these100-1Llsamples have been plotted directly without background subtraction. The effluent was also monitored at214nm to detecttryptic peptides derived from the apomyoglobin present as carrier in each sample (profiles not shown). Each radioactive peak has beenassigned a name derived from the isotope used (single-letter amino acid code) and the relative elution order for that peptide. Appropriatefractions were pooled, mixed with apomyoglobin, and lyophi-lizedinpreparation for radiochemical sequence analy-sis.Approximate positionsoftryptic peptides predict-edfrom the nucleotide sequence(Table 1)that have
exactly match thesequencepredicted beginning
at N1120 in the third reading frame.
Further-more, prolines found in peptide P5 and the
valine found in peptide V4, two peptides with
elution positions identicaltotheposition of A6,
also exactly matched this sequence (Table 1).
This result strongly suggested that the coding
region for the carboxy-terminal portion of the
33Kprotein beginsatleastasfar leftin the third
openframeasN1120, and thatan event
connect-ing the second and third open reading frames
mightoccurimmediately tothe leftof this point.
Analysis of peptides fromopenreading frame 2
indicatedthat the lastconfirmed residue in this
frame is the proline (residue 20) in peptide P7
(Fig. 4D) encoded by N870toN872. Therefore,
the connectingevent must occur to theright of
this site and within thenext 249nucleotides.
Two separate explanations for the above
re-sults are possible. An error might have been
made in the sequence between N872 and
N1120, orthemRNAforthe 33Kprotein might
bespliced within the coding region.
Partialnucleotide sequence analysis of the
ade-novirus 2 EcoRI-F fragment. To address the
possibility that a sequencing ambiguity may
have causedan apparentshift in the33Kprotein
reading frame, weindependently determined the
sequence of selected portions of the EcoRI-F
fragment. The sequence of 249 nucleotides
around the HindIIIsite at72.8 m.u. was
deter-mined by labeling HindIII-cleaved genomic
DNAwith[32P]ATPandpolynucleotide kinase,
isolating the relevant EcoRl subcut fragments
(70.7 to 72.8 m.u., 72.8 to 75.9 m.u.), and
treating these fragments by the method of
Maxamand Gilbert (12, 24). The 153 nucleotides
totheleft of N736 (to N583) and the 1%
nucleo-tidestotheright of N736 (to N932)werefoundto
be identicaltothepublishedsequence(13). Ina
similar way, except starting with the isolated
genomic EcoRI-F fragment, the 188 nucleotides
totheleft of the HpaII siteatN1554(to N1365)
and the 160nucleotides tothe leftof theHpaII
site at N1134 (to N975) were also found to be
identical to the published sequence. The
697-nucleotide sequence independently confirmed
by usinclude the sequence immediately before
the alanine codon at N1120. Thus the two
co-donsimmediately preceding this alaninemustbe
Ser-Thr- andnot(Lys/Arg)-Ala- asissuggested
by our direct peptide sequence analysis. Our
sequenceanalysishasconfirmedall but 43of the
nucleotides between N872 (the Pro in P7) and
N1120(thesecond Ala in A6).
beenidentifiedby sequence analysisare indicatedat
the top of(A). Predictedtrypticpeptidesarenumbered from the aminoterminus(Table1).
VOL.45,1983
on November 10, 2019 by guest
http://jvi.asm.org/
TRYPTIC PEPTIDES OF THE ADENOVIRUS 233K PROTEIN LQLPPPPPTDEEEYWDSQAEE LATVPSIATTSAPQAPPALPVR
peptide L12 C peptide A7
A 3H-Leu 1 4 3H-Ala
AAPTAPAAAAAAATAAVTQK
E peptide A6
140r
__. 3H-Ala;_ IV*_J v ;J IV v v v, Iv ;
RESIDUE RESIDUE RESIDUE
FIG. 4. Radiochemical microsequence analysisof adenovirus 2 33Kprotein tryptic peptides. Tryptic peptides ofradioactivelylabeled 33K proteinwerefractionated by reverse-phasechromatography as describedinthe
legendtoFig.3.Appropriate pooledfractionswereappliedtoaBeckman 890Csequencertogetherwith 2mgof apomyoglobinand 3mgofpolybrene.Panelpairs depictthetotalyieldofradioactivityin theamino acid fraction after eachEdmandegradation cycleforcorresponding peptides. Peptidesarenamedasdescribed inthelegendto
Fig. 3,and theisotopewith which eachpeptidewaslabeled is indicatedinthepanel. Peptideelutionpositions
canbe ascertained fromFig. 3orTable 1. The amino acidsequencepredictedfrom thenucleotide sequence
coffespondingtoeachpeptide pairisgivennearthe top of theupperpanelinthesingle-letteramino acid code. PeptidesA6 and P5(panelsE andF) correspondtothetryptic peptidethatspanstheintemalsplice junctionin
the33K mRNA(see text).Theamountofradioactivity appliedtothesequencerwasasfollows:(A) 3,900cpmof
[3H]leucineinL12; (B) 5,300cpmof[3H]prolineinP1o;(C) 1;520cpmof[3H]alanineinA7; (D) 13,500cpmof
[3H]prolineinP7; (E) 36,300cpmof[3H]alanineinA6;and(F) 6,650cpmof[3H]prolineinP5.
33K mRNA isinternally spliced.The structure ofpeptide A6 cannotreadilybeexplained by a
frameshifteventinthevicinity of N1120. There-fore, we examined the possibility that the 33K mRNA isspliced in amannerthatconnects the tworeadingframes andexplainsthe structureof peptide A6. The nucleotide sequence between
N1112 and N1120 (TCTCTACAG) conforms welltothe consensus splice acceptor sequence
YNYYYNCAG (34) (Fig. 6).Examination of the
sequencetothe left of N1120revealeda poten-tialsplice donor siteatN918. Asplicefrom this position tothe C at N1121 would result in the removal of 202 nucleotides from the transcript for the 33Kprotein, wouldpreservethe alanine
codon that appearedtobeencodedby N1120to N1122, and would cause this codon to be
pre-ceded in the mRNAbyanArg-Ala codon pair.
Thispredicted splice is precisely consistent with the observed structureofpeptide A6. No better potential splice donor sitewasfound by
exami-nation of the nucleotide sequencebetween N872
and N1120.
If the33K mRNA is indeedsplicedas suggest-edabove, the 202-nucleotide splice should have beenseenbyheteroduplexorSi nuclease
analy-sis ofregion L4 mRNAs, and certain peptides predicted tobe encodedby either the secondor
thirdopenreadingframe shouldnotbe identifi-able. In fact, a spliced L4 mRNA has been
observed both by Si analysis (8; S. Berget, unpublished data quoted in reference 25) and by heteroduplex analysis (10). Furthermore, we
havenotbeen abletofind severalpeptidesthat
we would have expected to observe during
reverse-phase chromatography of 33K tryptic peptides. For example, the proline-containing peptide Gln-Pro-Pro-Leu-Ala-Gln-Glu-Gln-Gln-Gln-Arg, predicted by thesecond openreading frame before the start of the third openframe,
has not been found among the proline-or ala-nine-labeledtryptic peptides.
N
x
0
Fn
w
E
06
0
x
3 0
w
E
U.
J. VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:8.489.52.447.74.339.2]ADENOVIRUS 2 33K PROTEIN 259
TRYPTIC PEPTIDES OF THE ADENOVIRUS 33K PROTEIN
N
0
x 0
Fn w
E
CL
0
0
n
w
a
w E
UL
10
RESIDUE RESIDUE RESIDUE
FIG. 5. Amino-terminal sequenceanalysis of adenovirus 2 33Kproteintryptic peptides. Sequenceanalysis wasasdescribedin the legend toFig.4.The amount ofradioactivity applied to the sequencer was as follows: (A) 8,300 cpm of[3H]prolinein P4; (B)4,550cpmof[3H]valineinV3; (e) 9,500 cpmof[3H]alanineinA3; (D) 1,200
cpm of [3H]phenylalanine in F8; (E) 5,450 cpm of [3H]leucine in L5; (F) 5,150 cpm of[3H]alanine in A5.
Additionalsequencingexperiments with other labels and otherpeptidesarenotshown;asummaryof all amino acidpositions directlyconfirmedbysequencingisgivenin Table 1.
To exclude the unlikely possibility that all
predicted peptides from the intervening
se-quence were lost during reverse-phase
chroma-tography, we prepared tryptic peptides of
[3H]isoleucine-labeled 33K protein and
subject-edthe complete mixture toamino-terminal
se-quenceanalysis (Fig. 7). The secondopen
read-ing frame can potentially code for five
isoleucines,threeof which wouldcomefrom the
predicted intervening sequence. The third open
frame could potentially encode six isoleucines,
threeof which also wouldcomefrom the
inter-vening sequence. The isoleucine codons in the
predicted intervening sequence are situated so
thatradioactivitywould beexpected in the 2nd,
3rd, and 14th residues from second reading
frame codons, and the 16th, 27th, and 31st
residues from third reading frame codons.
Ra-dioactivity would be expected in residues 1
(twice), 7, 8, and possibly3 from the remaining
isoleucine codons. Figure7 shows the presence
of isoleucine in residues 1,2, 7, andpossibly 8,
butnotsignificant radioactivityinresidues3,14,
and 16. Thepresenceofisoleucine in residue 2 is
explained by an unusual trypsin cleavage that
occursafter thetyrosineinpeptidet18(Table 1).
Trypsin is known to occasionally cleave after
tyrosine,and the existenceof the
carboxy-termi-nal fragment of this peptide has been
indepen-dently confirmed by reverse-phase
chromatog-raphy and partial sequence analysis. The
carboxy-terminal fragment elutes at
approxi-mately thesamepositionast27(fraction 62,data
not shown). Thus, at least two isoleucine
pep-tides from the expected intervening sequence
peptideswerenotfound,andallof theexpected
isoleucines from the peptides predicted to be
encoded beginning at N570, except for the
iso-leucineatposition3(seebelow),wereobserved.
Locations encoding the amino terminus and VOL.45, 1983
on November 10, 2019 by guest
http://jvi.asm.org/
[image:9.489.55.443.94.403.2]PREDICTED ADENOVIRUS 2 33K PROTEIN INTERNAL mRNA SPLICE POINTS
... Thr ThrGly Thr Arg Ala gly lys ser lys gln pro... Frame 3
... .ACCACT GGA ACC AGG GCC GGT AAG TCT MG CAG CCG... 1 strand (3' side)
901 911 921 931 nucleotide number
A GGU RAG 5' Splice consensus sequence
lice
... ile thr thr val ile ser thr Ala Pro Thr Ala Pro Ala Ala... Frame 1
... ATT ACT ACCGTC ATC TCT ACA GCC CCT ACT GCA CCG GCG GCA... 1 strand (3' side)
1101 1111 1121 1131 nucleotide number
Y- mYY -CA G 3' Splice consensus sequence
FIG. 6. Predicted internalsplice sites in the 33KproteinmRNA.Portions of the nucleotide sequence within the EcoRI-Ffragment are shown. The corresponding openreadingframe amino acid sequence is also given. Nucleotidesarenumbered from theEcoRIcleavage siteat70.7m.u.(13). Theconsensussplicesite sequences arefrom Sharp (34).
carboxy terminus of the 33K protein.The
amino-terminalandcarboxy-terminal sequences of the
33Kprotein have notbeen determined directly,
but several indirectarguments canbeappliedto
deduce their encoding locations. None of the
expected peptides predicted by the sequence
between N411 and N606 has been observed,
including the isoleucine-containing tryptic
pep-tide predictedby N570 to N587. Only one major
methionine-containing 33K tryptic peptide was
observed by reverse-phase chromatography,
andthis peptide elutedattheposition of peptide
t4(Fig. 3). This 33-amino-acid peptide is
expect-ed to contain two methionine residues at
posi-tions 29 and 30from the amino terminus (Table
1). Sequence analysis of the mixture of
[35S]methionine-labeled33Ktryptic peptidesdid
not reveal amethionine in the first20residues.
Sequence analysis of cyanogen
bromide-cleaved, [3H]alanine-labeled 33K mixed
pep-tides revealed no significant radioactivity in the
first 20 residues (datanotshown). Thus cleavage
after methionine residues produced no
un-blocked amino termini. These results allsuggest
that the 33K protein contains only the two
methionines expected in peptide t4. Cleavage
after these methionines by cyanogen bromide would leave an amino-terminal glutamic acid
residue which would be expected to cyclize
underthe strongacid conditions used. The
me-thionine codon atN606 to N608 isfollowed by
analaninecodon. Where this sequence occurs at
known eucaryotic protein synthesis initiation
sites (5, 37), the initiation methionine has been
found to be removed, and the amino-terminal
alanine becomes acetylated. Thus, our findings
arecompatible with initiation of33Ktranslation
at the N606-608 methionine codon. The se-quencesurroundingthis AUGalso matches the
mostfrequentlyfound eucaryotic translation
ini-tiation sequence, CANNAUGG (20). If
initia-tion were to occur at the only other available
methionine codon in the second open frame
(N411 to N413), we would notexpect it to be
removed post-translationally, and we would
have expected to find evidence for the
methio-ninepostulatedtobe encodedbyN606 to N608.
Furthermore, the sequences surrounding this
AUG codonsuggest that itmight not be a strong
initiation site for translation even if present in
the mRNA(20).
The last codonconfirmedby peptidesequence
analysis in the thirdopen frame is the Phe codon
ADENOVIRUS 33K PROTEIN
3H-Ile TRYPTIC PEPTIDES
'4
0
0
U,
0 5 10 15 20
RESIDUE
FIG. 7. Amino-terminal sequence analysis of the mixed [3H]isoleucine-labeled 33K tryptic peptides. Adenovirus 2 33Kproteinlabeled with[3H]isoleucine wasdigestedwithtrypsin. Thelyophilized digest con-taining 20,000 cpmwasdissolved in88% formic acid andapplied to the sequencer together with 2 mg of apomyoglobin and3mg ofpolybrene.Sequencingwas asdescribed inthetextandinthelegendtoFig.4.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:10.489.54.438.107.200.2] [image:10.489.254.445.388.603.2]ADENOVIRUS 2 33K PROTEIN 261
TABLE 2. Predicted amino acid composition of adenovirus 2 33K proteina
Predicted amino acidcomposition of proteins
Nonpolarb Noncharged, polarb Charged,polar"
Amino No. of % Amino No. of Amino No.of
acid residues acid residues acid residues
Gly 5 2.2 Ser 18 7.9 Asp 12 5.3
Ala 34 15.0 Thr 19 8.4 Glu 22 9.7
Val 7 3.1 Asn 4 1.8 Lys 14 6.2
Ile 4 1.8 Gln 11 4.8 Arg 17 7.5
Leu 20 8.8 Cys 2 0.9 His 1 0.4
Met 2 0.9 Tyr 4 1.8
Phe 4 1.8
Trp 3 1.3
Pro 24 10.6
aPredicted number of amino acids, 227; calculated polypeptide chain molecular weight, 24,892.
bNonpolar residues, 45.4%; noncharged, polar residues, 25.6%; charged, polar residues, 29.1%.
from t27atN1459toN1461. Elevencodons after this Phe codon, the sequencepredicts an UAG termination codon. The existence of this termi-nation codon was confirmed by our nucleotide sequence analysis. Because no obvious splice donor site islocated between N1461 and N1493,
we conclude that the 33K protein is likely to terminate with aspartic acidaspredicted by the
nucleotide sequence.
Composition of the 33K protein. The predicted
sequence of the 33K protein is given in Table 1
in the form of the expected tryptic peptide
sequences. Thepredicted amino acid
composi-tion is given in Table 2. The protein has ahigh
content of alanine (15%), proline (10.6%), and glutamic acid (9.7%). The 33K protein has previ-ously been showntocontainphosphoserine (6), and a preliminary analysis of the 32P04-labeled
tryptic peptides by reverse-phase chromatogra-phy indicates thatapeptide with the mobility of
peptide t4 is phosphorylated. This 33-amino-acid peptide contains sixserine residues;wehavenot yetdetermined whichone(s) is phosphorylated. Additional 32Pradioactivitywasobserved in the
column flow-through. We have not yet deter-mined whether this materialrepresentsasecond
phosphorylation site orphosphate released
dur-ing trypsin digestion. A potential cyclic AMP-dependent kinase phosphorylation site (37)
ex-ists in peptide tll (Table 1). No other peptide that is retarded during reverse-phase chromato-graphy was found tobe labeled by 32P04 (data notshown). At leastonepotential glycosylation
site(37) exists in the carboxyterminal portion of the 33K protein (Asn-Arg-Ser-); however, we
have no experimental evidence to suggest that this site is actually glycosylated.
DISCUSSION
We have shown that the adenovirus 2 33K protein is a unique, late, nonstructural virus
protein that is encoded bytwoseparateportions
of the EcoRI-F restrictionfragment (70.7to75.9
m.u.). Sequence analyses of radioactively
la-beledpeptides andcomparison withpredictions
made from thepublished nucleotidesequenceof
this region (13) have allowed us to deduce the
precise sites for an internal mRNA splice, the initiation site for translation, and the termination
sitefor 33Kprotein synthesis. Thisinformation
wasnot(13-15)and couldnothavebeenreadily
deduced from the nucleotide sequence alone
givenour presentunderstanding of the
molecu-larsignals required forgene expression.
The proposedsequenceof the 33Kproteinis
227 amino acids long; the calculated molecular
weight of this amino acid chain is 24,892, a
numberconsiderably smallerthanthe size
esti-matedfrom SDS-polyacrylamide gel
electropho-resis. Similarly anomalous behavior has been
noted for the adenovirus 2 ElA proteins (35).
We believe that the high proline and glutamic
acidcontentof theseproteins maycontributeto
theirlower-than-expectedmigrationratesduring
gel electrophoresis. In fact, we estimate the
molecular weight of the 33K protein by gel
electrophoresistobe closerto39,000Mrthanto
33,000 Mr.Toavoidconfusion,wehaveretained
the name "33K protein" until an appropriate
functionaldesignation canbegiventothis
prod-uct.
The amino acid sequence ofthe 33K protein
has some unusual features. The regionbetween
amino acid residues 103 and 118 (within t9,
Table 1) contains 12 alanines, 7 of which are consecutive. This sequence is encoded
immedi-ately after the internal mRNA splice and is
followedatresidue139 to 146by anotherrunof
five alanines and three glycines. Regions very
rich inproline (residues 9 to 13), glutamic acid
(residues16 to53) orbasic amino acids(residues
3 to5, 90 to 95, and 122 to 136)are alsofound.
Analysis oftheproteinsequencebythemethods
ofChou and Fasman(11)suggests the presence VOL.45, 1983
on November 10, 2019 by guest
http://jvi.asm.org/
OOSTEROM-DRAGON AND ANDERSON
of several short regions of probable a-helix.Two
of thesearein theamino-terminal portion
(resi-dues 22to38, and residues41 to 52)before the
mRNA splice point. The corresponding region
of the nucleotide sequenceprobably alsocodes
for the carboxy-terminal portion of the 1OOK
protein; 103 codons are predicted to
simulta-neouslycode for bothproteins. This is the only
region of the adenovirus 2 genome thus far
identifiedthatsimultaneously encodestwo
prod-ucts unrelated in amino acid sequence. The
carboxy terminus of the100Kproteinis
predict-edto occurjust before the 33K mRNA splice point.
Why is the 33K mRNA spliced? Asfar asis
known, neither strand of the sequencebetween
N918 and N1120 contains signals for other
eventsthatmight be incompatable witha
func-tional coding sequence for the 33K protein. A
promoter sequence for region E2 transcripts does occur onthe opposite strand (7), butat a
positioncorrespondingto coding sequencesfor
thecarboxyterminus of the 33Kprotein (Fig.2).
No termination codons in eitherreading frame
usedby the 33Kproteinoccurwithin theregion
removed by splicing. Thus we have found no
obvious explanation for the internalsplicein the
33K mRNA except to connect the two reading
frames andtoalter the amino acidsequencethat
would otherwise result.Perhaps the 33Kprotein
was once two separatefunctions,orperhaps the
regionbetween N918 and N1120performssome
function which has not yet been recognized.
WhilemanycellularproteinmRNAsarespliced
within coding sequences, theonly other
adeno-virus products with mRNAs knowntobespliced
withincodingsequences aretheearlyregion1A
products (35). It has been suggested, however,
that the mRNAsforcomponentIVa2andfor the
region E2Bproducts maybespliced justwithin
the amino-terminal coding region (Jeff Engler
and BruceStillman, personalcommunication).
The 33K mRNA presumably corresponds to
the L4 mRNA in which a splice has been
ob-servedby heteroduplexanalysis (10).The
struc-tureof this mRNA isnotknown precisely, but
presumably another splice also connects
se-quences to theleft ofN606(the initiationcodon)
tothetripartite leadersequence. The nucleotide
triplet CAG found in the spliceacceptor
concen-sus sequence occurs six times between N606 and N411; the codon for the first AUG in the
amino-terminal openreading frame is atN411.
Fourof these CAG sequencesare preceded by
pyrimidine-rich sequences whichare very
simi-lar to the concensus splice acceptor sequence. Incontrast, thefourCAGsequences thatoccur
betweenN606and thedouble-methionine codon
atN708 are preceded by purine-rich sequences
and donotappeartoresemble
splice
sites.The function ofthe 33Kprotein is unknown.
To ourknowledge,no mutantshavebeen
identi-fiedwhichmapwithinthe predicted33Kcoding
sequences. Severaltemperature-sensitive
muta-tions are known that map within sequences
encoding the lOOK protein. Two of these,
H5tsll5 andH5tsll6, havevery similar
pheno-types, belong to a single complementation
group,butmapabout 1 m.u. apart (27; E.
Oos-terom-Dragon, Ph.D. thesis, Yeshiva
Universi-ty, Bronx, N.Y., 1980) H5ts116 maps to
theleft of the EcoRI restriction siteat 70.7 m.u.,
whereasH5tsll5mapswithintheEcoRI-F
frag-ment(70.7to 75.9m.u.).The similar phenotypes
of these two mutants suggest that they are
defective in only the same function. Thus we
believe that the H5tsll5 mutationis likely to be
found before N606 and the start ofsequences
which alsoencode the 33Kprotein. Although we
havenotbeen abletodeduce the function of the
33K protein from knowledge ofthe nucleotide
sequenceswhich encode it, this knowledgemay
allow the construction of mutants which will
helptoelucidate its function.
ACKNOWLEDGMENTS
Wethank Medora Hardy and Sydell Lamb for excellent
technical assistance. We are grateful to John Dunn and William Crockett for assistance with the sequencing of DNA fragments, and to Marshall Elzinga, Nicholas Alonzo, and JeanneWysocki for assistance with peptide separation tech-niques and peptide sequence analysis. We thank Keith
Thompsonfor computer analysis of nucleotide and protein sequences.
Thiswork wassupported by Public Health Training grant
T32CA09121 fromthe NationalCancer Institute and by the
UnitedStatesDepartment of Energy. LITERATURECITED
1. Akusjarvi, G., J. Zabielski, M. Perricaudet, and U. Pet-tersson.1981.The sequenceof the 3' non-coding region of the hexon mRNA discloses a novel adenovirus gene. Nucleic Acids Res. 9:1-17.
2. Anderson, C. W. 1981.Spontaneousmutantsofthe adeno-virus-simian virus 40 hybrid, Ad2+ND3, that grow effi-ciently in monkey cells. Virology 111:263-269.
3. Anderson, C.W.1982. Partial sequencedetermination of
metabotically labeled radioactiveproteinsand peptides, p. 147-167. In J. K. Setlow and A. Hollaender (ed.), Geneticengineering:principles and methods, vol. 4. Plen-umPublishingCorp., New York.
4. Anderson, C. W., P. R.Baum,and R. F.Gesteland. 1973. Processing of adenovirus 2-induced proteins. J. Virol. 12:241-252.
5.Anderson, C. W., and J.B. Lewis.1980. Amino-terminal sequence ofadenovirus type 2 proteins: hexon, fiber, component IX, and early protein 1B-15K. Virology
104:27-41.
6.Axelrod,N.1978.Phosphoproteinsof adenovirus 2. Virol-ogy 87:366-383.
7. Baker, C. C.,T.Heriss6,G.Courtols,F.Galibert,and E. Ziff.1979.Messenger RNA for the Ad2DNAsequences
encodingthefirst leader andheterogeneityatthe mRNA 5'end.Cell18:569-580.
8. Berget,S. M.,and P. A. Sharp.1979. Structure of late adenovirus2heterogeneousnuclear RNA.J. Mol. Biol. 129:547-565.
J. VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
ADENOVIRUS 2 33K PROTEIN 263
9. Bhatti, A. R., and J. Weber. 1979. Proteaseof adenovirus type 2:partialcharacterization. Virology 96:478-485. 10. Broker, T. R., and L. T. Chow. 1979. AlternativeRNA
splicingpatterns and the clustered transcription and splic-ing signals of human adenovirus 2, p. 611-635. In R. Axel, T.Maniatis, and C. F. Fox (ed.), Eukaryotic gene regula-tion. Academic Press, Inc., NewYork.
11.Chou, P. Y.,and G. D. Fasman. 1978. Empirical predic-tions of protein conformation. Annu. Rev. Biochem. 47:251-276.
12. Dunn, J. J., and F. W. Studier. 1981.Nucleotide sequence from the genetic left end of bacteriophage T7 DNA to the beginning of gene 4. J. Mol. Biol. 148:303-330. 13. Galibert, F., J. Herisse, and G.Courtobs.1979.Nucleotide
sequence of the EcoRI-F fragment of adenovirus 2 genome.Gene 6:1-22.
14. Gambke, C., and W. Deppert.1981. Late nonstructural 100,000-and33,000-dalton proteins of adenovirus type 2. I. Subcellular localization during the course of infection. J.Virol. 40:585-593.
15. Gambke, C., and W. Deppert. 1981. Late nonstructural 100,000- and 33,000-daltonproteins ofadenovirus type 2.
II.Immunological and protein chemical analysis.J. Virol. 40:594-598.
16. Ginsberg, H. S., and C. S. H. Young. 1977.Genetics of adenovirus, p. 27-86. In H. Frankel-Conrat and R. R. Wagner (ed.), Comprehensive virology, vol. 9. Plenum PublishingCorp.,NewYork.
17. Hassell, J. A., and J. Weber. 1978. Geneticanalysis of
adenovirustype 2. VIII. Physical locations of
tempera-ture-sensitive mutations.J.Virol. 28:671-678.
18. Henderson, L. E., S. Oroszlan, and W. Konigsberg. 1979. Amicromethodforcompleteremoval of dodecylsulfate from proteins by ion-pair extraction. Anal. Biochem.
93:153-157.
19. Kessler, S. W. 1975.Rapidisolationofantigensfrom cells with astaphylococcal protein A-antibody adsorbent: pa-rameters of the interraction ofantibody-antigen complex-eswithproteinA.J. Immunol. 115:1617-1624. 20. Kozak, M. 1981. Possible roleofflanking nucleotides in
recognition of the AUG initiator codon by eukaryotic
ribosomes. Nucleic Acids Res. 9:5233-5252.
21. Krul3er,W., F. M. A. van Schaik, and J. S. Sussenbach. 1980.Nucleotide sequenceanalysisof aregion of adeno-virus 5 DNA-encoding a hitherto unidentified gene. Nu-cleic Acids Res. 8:6033-6042.
22. Levinson,A.D., E. H. Postel,and A.J. Levine. 1977. In vivoand in vitro phosphorylation ofthe adenovirus 5
single-strand specific DNA-binding protein. Virology
79:144-159.
23. Lewis,J. B., J. F. Atkins, C. W. Anderson, P. R. Baum, andR. F. Gesteland. 1975. Mapping of late adenovirus genesby cell-freetranslation of RNA selectedby
hybrid-ization tospecificDNAfragments.Proc.Natl.Acad. Sci. U.S.A. 72:1344-1348.
24. Maxam, A. M., and W. Gilbert. 1980. Sequencing end-labeled DNA with base specific chemical cleavages. MethodsEnzymol. 65:499-560.
25. Miller, J. S., R. P. Rkxiardi, B. E. Roberts, B. M. Paterson, and M. B. Mathews. 1980. Arrangement of messenger RNAs and protein coding sequences in the major late transcription unit of adenovirus 2. J. Mol. Biol. 142:455-488.
26. Oosterom-Dragon, E. A., and H. S. Ginsberg. 1979. Purifi-cation andpreliminaryimmunologicalcharacterization of the type 5adenovirus, nonstructural 100,000-dalton pro-tein. J. Virol. 33:1203-1207.
27. Oosterom-Dragon, E. A., and H. S. Ginsberg. 1981. Char-acterization of two temperature-sensitive mutants of type 5 adenoviruswith mutations in the100,000-daltonprotein gene. J.Virol.40:491-500.
28. Palmiter, R. D. 1977. Prevention of NH2-terminal acetyla-tion ofproteinssynthesized in cell-free systems. J. Biol.
Chem.252:8781-8783.
29. Philipson,L., U. Pettersson, and U. Lindberg. 1975. Mo-lecularbiology ofadenoviruses. InS.Gard and C. Hal-laver(ed.), Virologymonographs, vol. 14.
Springer-Ver-lag, New York.
30. Ricciardi, R. P., J. S. Miller,andB. E. Roberts. 1979. Purification andmappingofspecific mRNAsby
hybrid-ization-selection and cell-free translation. Proc. Natl. Acad.Sci. U.S.A.76:4927-4931.
31. Russell,W.C.,andG. E. Blair.1977.Polypeptide
phos-phorylation inadenovirus-infected cells. J. Gen. Virol. 34:19-35.
32. Schechter, N. M., W.Davies,andC. W. Anderson.1980.
Adenovirus coded deoxyribonucleic acidbinding protein. Isolation, physical properties, and effectsofproteolytic digestion.Biochemistry19:2802-2810.
33. Semler, B. L., C. W. Anderson, N. Kitamura, P. G. Rothberg, W. L. Wishart, and E. Wimmer. 1981. Poliovi-rus replicationproteins: RNA sequence encodingP3-lb
and thesites ofproteolyticprocessing.Proc.Natl.Acad. Sci. U.S.A.78:3464-3468.
34. Sharp, P.A. 1981. Speculationson RNA splicing. Cell 23:643-646.
35. Smart, J. E., J. B. Lewis,M. B.Mathews, M.L.Harter, and C. W. Anderson. 1981. Adenovirus type 2 early
proteins: assignment of the early region 1A proteins
synthesized in vivo and in vitro to specific mRNAs. Virology 112:703-713.
36. White,D.O., M. D. Scharff, and J. V. Maizel.1969.The
polypeptidesof adenovirus III.Synthesisin infected cells.
Virology38:395-406.
37. Wold, F. 1981. In vivochemicalmodifications ofproteins. Annu.Rev. Biochem. 50:783-814.
38. Zain, S., J. Sambrook,R.J. Roberts,W.Keller,M.Frled, and A. R.Dunn. 1979. Nucleotide sequenceanalysisof the leadersegments in acloned copy of adenovirus 2 fiber mRNA. Cell 16:851-861.
VOL.45,1983
on November 10, 2019 by guest
http://jvi.asm.org/


![FIG. 4.canapomyoglobinoflegendafterthecoffespondingFig.Peptides[3H]leucine[3H]proline radioactively Radiochemical microsequence analysis of adenovirus 2 33K protein tryptic peptides](https://thumb-us.123doks.com/thumbv2/123dok_us/1450994.97649/8.489.52.447.74.339/canapomyoglobinoflegendafterthecoffespondingfig-peptides-radioactively-radiochemical-microsequence-analysis-adenovirus-peptides.webp)


