JOURNALOFVIROLOGY, July 1978,p. 56-73 0022-538X/78/0027-0056$02.00/0
Copyright© 1978 AmericanSocietyforMicrobiology
Vol.27,No.1
Printedin U.S.A.
Mapping Early
Transcripts
of
Herpes
Simplex
Virus
Type
1
by
Electron
Microscopy
JAMES R. STRINGER, LOUIS E. HOLLAND, ANDEDWARD K.WAGNER*
Departmentof Molecular Biology andBiochemistry, University ofCalifornia, Irvine, California92717
Receivedfor publication23September1977
RNA displacement loop patterns in intact
herpes
simplex
virus DNA andherpessimplexvirus DNA restrictionfragmentsindicate that viral RNA
associ-ated with polyribosomes early after infection is transcribed from three major
areas of thegenome. One area ofearlytranscriptionis in the shortsegment of the
viralDNA and isroughlydelineatedby the invertedrepeat sequencesbounding
thissegment.The othertwo areasofearly mRNA
transcription
map in thelongsegment. Each ofthe three major areas ofearly mRNA
transcription
can befurther resolved into several regions offrequent loopingbordered by regionsin
which RNAdisplacement loopsare rare.Theseregionsrange in size from about 1.5kilobasestoabout9kilobases witha meansize ofabout 3.5kilobases.Although
the data do not allow precise assignmentofindividual earlygenelocations,it is
seen, even atthe lowest level ofresolution,thattheearlygenesare notcompletely
contiguous butaredistributedalong the
length
of theherpessimplex
type1viral genome.Theexpression ofherpessimplex virustype1
(HSV-1) during productive infection is
tempo-rallyregulated (14, 25, 26,29).Thefactors
gov-erning the control of HSV-1geneexpressionare
largely unknown.Animportant considerationin
studying such factors isa
knowledge
of thear-rangementofcoordinately expressed viralgenes.
PriortoviralDNAsynthesis(early),twoclasses of viralRNA,differinginabundancebyatleast 10-fold,are present onpolyribosomes. The abun-dant class of early RNA ishomologousto20 to
25% of the viral DNAsequences, and contains
two components, "a" or "immediate early"
RNA, and
",B"
or"delayed
early" RNA. Thesecomponents are experimentally distinguishable
by the fact that a or immediate early HSV-specific RNA is found in the cytoplasmin cells treated with cycloheximide during infection, while the,B,delayed early RNA isnot (6,9, 14,
26, 29). The amount of the DNAhomologousto
a-immediate earlyHSV-specific RNAhas been
reported to be in the rangeof10 to15%, i.e., half
orlessof the totalearlyRNA (14).
Thelow-abundance class of viral RNA found incellsprior toviral DNAreplicationis
homol-ogous to a further 20 to30% of HSV DNA (14,
25, 26, 29). This class of HSV RNA is present at much higher concentrations
following
viral DNA replication and is the "y" or"late" viral RNAencodingstructuralproteins (12, 14, 25, 26, 29).There are two fundamentally different
tech-niquesfor mapping viral transcripts onto viral
genomes. The first involves hybridizing viral
RNA to restriction endonuclease fragments
us-ing either solution hybridization or the blot
method of Southern(22). Thesecondtechnique,
termed RNA displacement loop (R-loop)
map-ping (27), utilizes the electron microscope to
directly visualize loops formed induplex DNA
through displacement of one DNA strand by
RNA moleculeshomologous to the other DNA
strand.
Bothmethods offer their ownunique
advan-tages.Hybridizationtorestrictionfragments
of-fersrelativespeed and,atleastpotentially,
quan-titation of the abundance of classes of RNA
transcriptshomologoustothefragmentin
ques-tion.R-loopingcanbeutilizedathighresolution
tovisualize individualtranscripts, thedirection
of the transcripts on the DNA, and unusual
structuressuch as"spliced" mRNAreported in
adenovirus and simianvirus 40 (2, 4, 5, 15-17).
R-looping at moderate resolution can resolve
areasoftranscriptional activity and itsabsence
withinregionsofthegenomeunresolved by
re-striction endonucleasecuts.Together,the meth-odsprovidecomplementary,aswellas
reinforc-ing, information concerningthe transcriptional activityofviral genomes. Both have been used
toresolvetranscriptional maps of the adenovirus
andsimian virus 40 genomes (1, 5, 15).
Biochemicalmapping by hybridization to
re-striction fragments has been used to locate
a-immediate early HSV transcripts. These studies showthatthe a-immediate earlygenes are not 56
on November 10, 2019 by guest
http://jvi.asm.org/
Voil. 27,1978
contiguous in the viral genome (6, 13). Since
total early HSV RNA has been shown to
hy-bridize to everyrestriction fragment examined
(6), there has been no definition in the total
earlyRNA transcriptionmap.
We have applied the R-loop mapping
tech-nique todeterminethe arrangement of the genes
codingfor totalearly HSV-1 mRNA. The data
presentedinthisreportprovidea
moderate-res-olution map of total early HSV-1 transcripts
which demonstrates areas of high, moderate,
and low or notranscriptionalactivity.
MATERIALS AND METHODS
Celisand virus. HeLa cellsweregrown in
mono-layer culture in Eagle minimal essential medium (8)
with Earle salts, 10% calf serum, and no antibiotics.
Growth conditions and assay formycoplasmic
contam-inationwere asdescribed(24, 29). TheKOS strain of
HSV-1was agift from G. CohenattheUniversity of
Pennsylvania. Virus was grown in HeLa cells at a
multiplicity of infection of0.1 PFUpercell and
rou-tinely plaque purified after four to six passages as
described (31).
RNA. Cells were infected at a multiplicity of30
PFU per cell using conditions described previously
(25, 26).Time after infection was measured after virus
absorption (0.5h). Cellswere harvestedat 3hafter
infection.
Details ofcell fractionation, polyribosome isolation,
and polyadenylated RNA purification were as
de-scribed(23). Briefly,theprocedure appliedthe
mag-nesium ionprecipitation method of Palmiter(19) to
obtainpolyribosomes. The Mg2+-precipitated
polyri-bosomepelletwassuspendedin 0.01 MNaCl-0.01 M
Tris-0.0045 M MgCl2, pH 7.4 (RSB), and briefly
di-gestedat roomtemperature with 200,gof
electropho-reticallypurified DNase(SigmaChemicals, St. Louis,
Mo.) per ml. RNAwasextracted from the
polyribo-somefractionbyproteinase K (Merck
Pharmaceuti-cals, Elmsford, N.Y.) digestion, followed by phenol
extraction as described (23). Radioactive RNA was
preparedasabove, butcellswerelabeled from2to 3
hafterinfection with15,uCiof[3H]uridineper ml(27
Ci/mmol; Schwarz Bio Research, Inc., Orangeburg,
N.J.).
DNA.HSV DNAwaspreparedbothas areagent
tobeused in the isolation of HSV RNA andasintact
genome-sizemolecules for restriction andR-loop
anal-ysis.Partially purifiedvirionswereobtained from
in-fectedcellhomogenatesbyequilibrium centrifugation
inadiscontinuoussucrosegradientasdescribed (28,
30, 31). DNA for preparative hybridization of HSV
RNAwaspreparedfrom thepartially purifiedvirions
asdescribedandassayedforpurity byanalytical
iso-pycniccentrifugation (23, 30).
Forisolation ofintactHSV DNA molecules,
par-tially purified virions (30, 31) were obtained as
de-scribed aboveand thenfurtherpurified by
homoge-nization in RSB containing 0.5% Nonidet P-40 and
differential centrifugation. Purified virions from 4x
108to5x 108 cellsweresuspendedin 0.9 ml of 0.05 M
Tris-0.02 M EDTA(pH8.0), andthe DNAwasgently
released by the addition of0.1 ml of 10% Sarkosyl.
MAPPING EARLY HSV-1 TRANSCRIPTS 57
The DNA wasdeproteinized by digestion with 200
jig
of proteinase K per ml for 6 h at 45°C, then loaded on
anagarose A-150column (2 by 35 cm; BioRad,
Rich-mond, Calif.) equilibrated with 0.5 M NaCl-0.1 M
PIPES [piperazine-N,N'-bis(2-ethanesulfonic acid)]
-0.005 M EDTA (pH 7.8). The column eluant was
collected in 1-ml fractions, and DNA was monitored
by absorbance at 260nm. Viral DNA collected from
the agarose column was between 20 and 50 ,ug/ml and displayed a single component thermal denaturation
profile with aTm of 88°C in 0.1x SSC (SSC is 0.15 M
NaCl-0.015 M sodium citrate). This material was com-posed primarily of intact HSV DNA molecules as
determined by electron microscopy. Nocellular DNA
was present as determined by analytical isopycnic
centrifugation.
Restriction endonuclease digestion of HSV
DNA. Restriction enzymes, EcoRI, HindIII, and Xba
I, werepurchased from New England Biolabs
(Bev-erley, Mass.).Digestion conditions were: 0.10 M
Tris-hydrochloride (pH 7.5)-0.05 M NaCl-0.005 M
MgCl2-100 ,tg of bovine serum albumin per ml, for
EcoRI; 0.06 M NaCl-0.007 M MgCl2-0.007 M
Tris-hydrochloride (pH 7.4), for HindIII; and 0.15 M
NaCl-0.006 M MgCl2-0.006 M
2-mercaptoeth-anol-0.006 MTris-hydrochloride (pH7.9), for Xba I.
DNA was digested at a concentration of 25 ug/ml.
Aliquots of1to 3ugof restricted DNA were layered
onto0.7% agarose (Sigma) gels, poured in Plexiglas
tubes(0.6x 14cm).DigestedDNAwassubjected to
electrophoresis in gel buffer (0.04 M Tris-hydrochlo-ride-0.005 M sodium acetate-0.002 M EDTA, pH 7.8)
at 0.5 mA per gel for48 to 60h at 4°C. Gels were
stained with 0.5,gof ethidiumbromide per ml of gel
buffer, and bandswerevisualized by fluorescence at
360nm.
DNAwasrecovered from gels bydissolving the gel
slicecontaining the band of interest in saturated KI,
dialyzing exhaustively against 0.10 M NaCl-0.01 M
Tris-0.001 M EDTA(pH7.4), andremoving the
solid-ified agarose by centrifugation at 15,000 rpmfor 30
min.From40to50%of the DNA is released into the
supernatant by suchtreatment. The DNAwasthen
extracted with isopentyl alcoholtoremove ethidium
bromide.
TheHindIII, Xba I, and EcoRI restriction sites in
the DNA of HSV-1 (KOS) have been mapped (21).
Whendigested by either Xba I orEcoRI, ourDNA
yielded digestionproducts which,onagarosegels,were
identicaltothoseexpectedfrom thepublishedmaps.
The identity of individual Xba I digestionproducts
wasconfirmed byredigestionof each agarosegel band
with EcoRI. When digested by HindIII, our strain
produced an agarose gel band pattern that lacked
bands corresponding to fragments I and 0 of the
published map.There ispresentabandaslargeasthe
sumofI and 0 instead. SinceI and 0 are adjacent
pieces of DNA, thesimplest explanation is thatthe
cleavage site between I and 0ismissinginourstrain.
Redigestionofindividual HindIIIdigestionproducts
withEcoRI showedthis to be the case. TheHindIII,
XbaI, and EcoRI restriction sitesinthe DNA of the
virus strain usedin theseexperiments are, therefore,
the same asthose publishedfor HSV-1 (KOS) (21)
except foramissingHindIII sitebetweenfragmentsI
on November 10, 2019 by guest
http://jvi.asm.org/
58
STRINGER, HOLLAND, AND WAGNERand0(seeFig. 5).
Preparative hybridization of HSV RNA.
Sev-eral preparations ofHSV-specific polyribosomeRNA
were prepared for R-loop hybridization as follows.
Infected-cell polyribosomalRNAlabeled with
[3H]ur-idinefrom2to 3hafter infection and isolatedatthe
end of the labeling period was added in eightfold
(wt/wt) excesstonative HSV DNA.32P-labeledHeLa
cellribosomal RNAwasaddedas aninternalmarker,
andthemixturewasethanolprecipitated. The
precip-itate was dissolved in 98% formamide and 0.1 M
NaCl-0.003 MPIPES-0.0003 M EDTA (pH 7.0) and
incubated at 72°Cfor 10minto denature the DNA.
Thesolutionwas nextadjustedto93%formamideand
0.3MNaCl-0.01MPIPES-0.001 M EDTA and
incu-batedat480Cfor24h. Under theseconditions,
rean-nealing of HSV DNA doesnotoccurand only
RNA-DNAhybridsareformed(3; L. E. Holland and E. K.
Wagner, unpublished data). At the end of the
incu-bationperiod, thehybridizationsolution wasdiluted
by the addition of2volumesof0.15Msodiumacetate
(pH 5.0) and precipitated with3volumesof ethanol.
The precipitate was suspended in 1 ml of2x SSC
containing0.020 M EDTA and treated with RNase
(10 ,ug of pancreatic + 10units ofTi per ml) for 10
minat 37°C. RNase wasremovedby digestion with
500,ugofproteinase K per ml in the presence of 0.5%
sodium dodecyl sulfateat 37°Cfor20min, followed
byphenolextraction.
Hybridswereseparated from unhybridized RNA by
gelfiltrationonSephadexG-100(Pharmacia,Uppsala,
Sweden).AG-100column (1.5by50cm),equilibrated
in0.1MNaCl-0.01 M Tris-0.001 M EDTA (pH7.4),
wasloaded with the aqueousphase fromphenol
ex-traction, and 1-ml fractions were collected. Hybrid
RNA(15% of the total 3H radioactivity) eluted in the
voidvolume well before any 32P-labeled rRNA.
Frac-tionscontaining hybridswerepooled and ethanol
pre-cipitated.Theprecipitatedhybridsweresuspended in
80% formamide and 0.2 M NaCl-0.02 Tris-0.001 M
EDTA(pH 7.4) and denatured by heatingat72°C for
10min. Formamide was removedby passage overa
Sephadex G-25 column equilibrated in 0.01 M
NaCl-0.01 M Tris-0.0045 MMgCl2(pH 7.4), and the
DNA wasdigestedat370C for 5 min with 100Mgof
electrophoreticallypurified DNase I per ml. The
mix-turewasthen made 0.5% sodium dodecyl sulfate 0.005
MEDTA, phenol extracted, and passed over a
Seph-adexG-100columnequilibratedin0.1MNaCl-0.01M
Tris and0.001MEDTA(pH 7.4). Excluded RNA was
collectedandethanolprecipitated. RNA obtained by
this method had aspecific radioactivity of
approxi-mately 50,000 cpm/yg and was free of 32P-labeled
rRNA. Electron microscopy showed no evidence of
DNA contamination. Rehybridization ofRNAusing
DNA excesshybridization (23) demonstratedthat it
wasatleast85%viral.
R-loop hybridization. R-loop hybridization was
performed in 75% formamide, 0.2 M NaCl-0.1 M
PIPESand 0.005 M EDTA(pH 7.8)at56°C for 20h.
Intact, native HSVDNAconcentrationswerefrom5
to10,ug/ml.Concentrationsof individualDNA
frag-ments generated by digestion withrestriction
endo-nucleaseswerenotdirectly determined, but were at
leastfivefold less than that of intact viral DNA.
Ex-,J. Viimu.
periments with total infected cellpolyribosomalRNA
wereperformedatRNA concentrations from50 to 100
times greater than the concentration of DNA.
RNA-to-DNAconcentration ratioswereapproximately2in
experiments withpurifiedHSVRNA and7in
experi-mentsusingpolyadenylatedRNA(basedon aspecific
activity of 50,000 cpm per,g ofRNA). DNAannealing
controlsweredone under thesameconditionsas
de-scribed above forR-loop experiments.Digestionswith
S1 single-strand nucleasewereperformedasdescribed
previously (19).
Electronmicroscopy. R-loop hybridization
mix-tureswerediluted 30-fold into 100
pl
of 50%formam-ide-0.1 M Tris-0.01 M EDTA(pH 8.5) containing, per
ml,50 ugofcytochrome c and0.2Mgof the relaxed
replicative form of4X174DNA(4XRFII DNA).This
mixturewasimmediately spreadover ahypophaseof
either10%formamide plus0.01MTris-0.001 M EDTA
(pH 8.5)or20%formamideplus0.01 MTris-0.001 M
EDTA (pH 8.5). Parlodion-coated microscope grids
were touched to the cytochrome-DNA monolayer,
stained withuranyl-acetate, and rotary shadowed with
platinum-palladium. Molecules were visualized and
photographed in either the Siemens 1Aor theJeol
100C electron microscope. Intact genome-size DNA
molecules werephotographed at anominal
magnifi-cationofx2,600.DNAfragmentswerephotographed
atmagnifications ranging fromx3,000toxlO,000.
Measurement of molecules. Negatives of
R-looped HSV DNA molecules and restriction fragments
wereprojected and traced to produce
high-magnifi-cation (x20to X30)replicas of the photographic
im-ages.ThesetracingsweremeasuredonaTektronics
4956 Graphics Tablet and Tektronics 4051 computer
(Tektronics Corp., Beaverton, Ore.). All HSV DNA
wasmeasured inreferencetothe average length of at
least104XRFIIDNAmolecules present in the same
field. HSV DNA molecules measuring 28±14XRFII
DNA unitswere scoredasintactviral genomes and
normalized to a standard length (28 4XRFII DNA
units). HSV DNA restriction fragments which fell
within 5%of themeanmolecularlength for that
frag-ment wereconsideredfullsize andwerenormalized to
themeanlength. Themeanlengths of the restriction
fragments analyzed agreed quite closely with those
expected fromanalysisonagarosegels.
RESULTS
R-loop
reaction. When duplex DNA is in-cubated with homologous RNA in highconcen-trations offormamide, RNA can hybridize to its
complementarysequence in the DNA, thereby
displacing the other DNA strand and forming an R-loop. R-loops form because RNA:DNA
hybridsare morestable than DNA duplexes in
high-formamidesolvents (3, 27). The rate of the
R-loop reaction is strongly dependent on the
incubation temperature,proceedingfastest near
the melting temperature of the DNA duplex
(27). The Tmfor HSV-1 DNA under our
hybrid-ization conditions was
570C,
as deternined byanalysis with
Si
endonuclease. Ourhybridiza-tionswereperforned at
560C.
on November 10, 2019 by guest
http://jvi.asm.org/
Voi.. 27, 1978
R-loops formed in the viralDNAupon
incu-bation witheither total infectedHeLa cell
po-lyribosomal RNA, polyadenylated infected
HeLa cell polyribosomal RNA, or HSV RNA
purified by preparative hybridization. The high
concentration oftotal infected cell
polyriboso-mal RNA required to form R-loops in HSV
DNAseverely impaired thevisualization of the
DNA. Most of the data presented hereare
there-fore derived from DNA molecules hybridized
with purified HSV RNAorwith polyadenylated
RNA. R-loops on representative intact HSV
DNA molecules are shown in Fig. 1A.
Repre-sentative R-loops showing good contrast
be-tween the double-stranded DNA:RNA duplex
and the displaced single-stranded DNA are
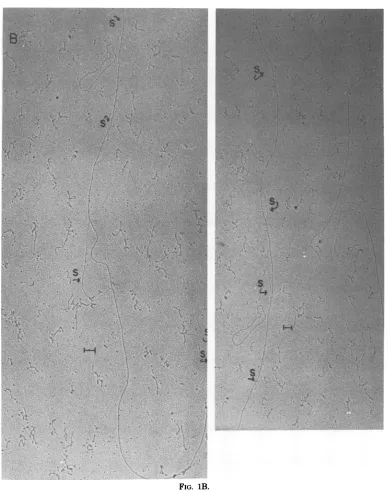
shown in Fig. 1B.
TheR-loop reaction isabsolutely specific for
viral RNA. No loops occurred when viral DNA
was incubated alone (Fig. 1C), with
mock-in-fected HeLa cell RNA, with
ribonuclease-treated infected-cellRNA,orwith
ribonuclease-treated HSV RNA. ThatnoDNA:DNA
anneal-ing occurred under R-loopanneal-ing conditions is also
shown in Fig. 2. No DNA:DNA duplex was
fornedevenwhen the reactionwasdriventoa
DNA Cotgreaterthanfive times that attained
in R-loop experiments.
The orientationproblem. Tovisuallymap
structuressuchasR-loopsonlinear DNA
mol-ecules, the molecules must be arranged in a
standard orientation.Orienting HSV DNA
mol-eculesisaproblem exacerbated bytheintemal
rearrangements which occur in the genome
yielding four differentarrangementsof the DNA
in approximately equimolar amounts (11, 20).
This property of the HSVgenome precludeda
simple best-fit R-loop alignmentorientation
pro-cedure,and itwasthereforenecessaryto
deter-mine the R-loop patternsofdefinedregions of
the DNA to aid in orienting intact R-looped
HSV DNA.
Due to the technical difficulties of handling
molecules the size of HSVDNA,wehave found
it impracticable to work with molecules
R-loopedinalarge portion of their length.
There-fore,wehave examinedalarge numberof intact
HSV DNAmoleculesbearingalimited number
ofR-loops permolecule. Since only molecules
bearing alimited number ofR-loopsper
mole-cule couldbe obtained ingood yield,itwasbest
toemploy regionsdevoid ofR-loopsas
orienta-tion aids. Regionsof the genomewhichare
in-frequently R-looped by early polyribosomal
RNAwererevealedby (i)analysisof theR-loop
frequency distribution ofa randomly arranged
population of intact HSV DNA molecules and
(ii) theR-loop patterns onpositionally defined
restrictionfragmentsof viral DNA.
MAPPING EARLY HSV-1 TRANSCRIPTS 59
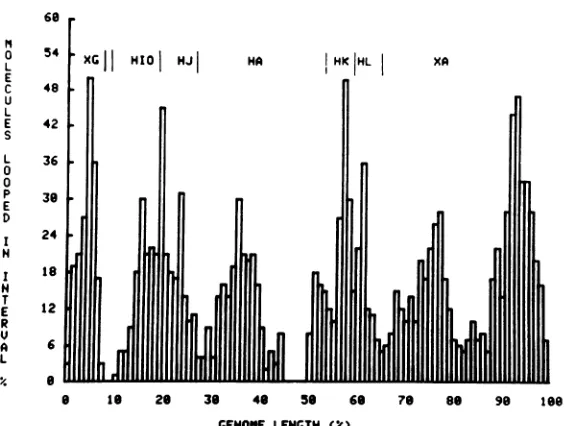
Random-order R-loop pattern. The
fre-quencydistribution of R-loops in51intactHSV
DNAmolecules, linedupwithoutregard to ori-entation, is shown in Fig. 3. As expected, the
histogram is complex and shows symmetry
around the middleof the distribution. Because
of the internal rearrangements in HSV DNA, mostpositions in the random-order histogram
represent four different physical loci on the
DNA (see Fig. 4); therefore, the absence or
presenceofR-loopsat mostDNAloci isdifficult
toascertainfrom the random-order histogram.
The center of the histogram is an exception
because
only
twophysical
loci on the DNA contributetothis region of the histogram. Since there isalow level ofR-loops around thecenterofthe random-order histogram,R-loops must be
rare orabsentatthetwolociin the DNAwhich
contribute to this region of the histogram.
In-spection of Fig.4shows that thetwoDNA loci whichwill contribute to the centerof the
ran-dom-orderhistogramoccur at 30and50%ofthe
genomelength in from the long-segment end of
the DNA.
R-loops
inselected HSV DNA restriction fragments. As discussed in Materials andMethods,
theHinduI,
XbaI,
andEcoRI restric-tionsites in the DNA of the HSV-1 (KOS) line used in theseexperiments
arethesame asthose published for HSV-1 (KOS) by Skare andSum-mers (21) exceptforamissing HindIII
cleavage
sitebetween
fragments
Iand0 (Fig. 5). Three Xba Irestrictionfragments, A, F, and G, wereselected for R-loop analysis. These fragments
accountfor 58% of theDNA and can beobtained
as
single
bandsonagarose gels.As seenin Fig. 5, thesefragments
coverthe distal 8% of thelong
segmentofthe HSV-1genome
(fragment
G);
asegment 16% of thegenome in
length
nearthecenterof thelongsegment(fragment F); and the
terminal 36% of the HSV-1genome,
encompass-ingboth arrangements of the shortsegment of
the molecule
(fragment A). Selected fragments
bearingR-loops
areshown inFig.
6.Individual molecular histograms for 25molecules ofeach of these fragments are shown inFig.
7A. Acomposite histogram
ofR-loops
of thesefrag-ments
corresponding
to theirposition
on theintactHSV-1genomeis shownin
Fig.
7B.Inspection of selected individualAfragments (Fig. 7A) showed thatloopsmayoccur onboth
ends ofanindividualfragmentbutare rare near
the center. Since any arrangement of A
frag-ments will
produce
asemi-symmetrical
histo-gram, the orientation problem is obviated. On
the other
hand,
sinceloops
donot occur onbothends of every Afragment, thelack of any
align-ment criteria means the relative
peak heights
within the histogram of the A
fragments
areon November 10, 2019 by guest
http://jvi.asm.org/
60 STRINGER, HOLLAND, AND WAGNER J. Viui-.
=~~~~
(fS;
X
-P
--{..-V~~~~~~~~~~V
FIG. 1. Electronmicrographs ofHSV-1 (KOS)DNA molecules bearing R-loops formedwith earlyRNA
Hybridizationwasfor24hat56°Cin 75%Formamideasdescribed in thetext. Thesmall circularmolecules
areOXRFIIDNA,includedas asize marker.(A)AnHSV DNA moleculeR-loopedwithpurifiedHSV RNA. The HSV DNA molecule is27.7
0XRMI
unitslong (47.1tun).
ThebarindicatesItun. (B)R-loopsformedin HSVNAb totl ealy RA islatedfrom polyribosomes. Arrows indicate loops, Sdesignatessingle-strandedDNA. The barindicates 0.1 ,um. (C)HSV DNA molecule incubatedunder conditionsidentical to
thoseforA and B but in theabsenceofRNA.Thelengthofthemolecule is 28.0
OXRFII
units(47.4 ,um).The bar indicates 1.0 ,un.on November 10, 2019 by guest
http://jvi.asm.org/
[image:5.499.52.452.61.567.2]MAPPING EARLY HSV-1 T'HRANSCRIIPT'S
v~~~~~
.t~~~~~~~~~~~4
r~~~c
'-"-V..,.;~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~
[image:6.499.59.448.70.561.2]~ @. ;|> w
FIG. 1B.
somewhatarbitrary.The AfragmentsinFig.7A
have been aligned to produce approximately
equivalent R-loop frequencies at both ends of
thehistogramsectioncorrespondingtofragment
A(Fig. 7B).
XbaIfragmentsFand Gboth showed
exten-siveR-loopingandcould not beunambiguously
orientedtoproduce uniquedistributions.
There-fore,inorientingR-looppatterns of intact
mol-ecules fragmentsF andGwereutilized onlyto
the extent that they established extensive
R-looping in the region of thegenomerepresented
bythesefragments.
Orientation ofR-loopedintact HSV DNA.
TheR-looppatternofXbaIfragmentA
unam-biguously established a high incidence of
R-Voi.. 27, 1978
61
on November 10, 2019 by guest
http://jvi.asm.org/
62 STRINGER, HOLLAND, AND WAGNER 1Vri.
C
t
*
C
4
1'
t
FIG. 1C.
loops in DNAlocatedoneither side ofthelong segment-short segment junction and very low levels ofR-loopingin theregionof thejointitself
(Fig. 7B). Analysis of the fragment, therefore,
revealed a defined non-R-looped region in the
DNA, which could be used to orient intact
R-loopedHSV DNA.
The existence ofat least one large region in whichR-loopsarescarcein thelongsegmentof the DNA was suggested by the random-order .4
J. vilml'.
11
11 I'' .:
I
'R
I
I I A
III, * .b
I
I
I
- F
11
I i
10 .
on November 10, 2019 by guest
http://jvi.asm.org/
[image:7.499.61.450.61.588.2]MAPIPING EARLY HSV-1 T'RANSCHIPI'TS
-1 -0.7 -6.4 -6.1 6.2 0.5 6.8 1.1 1.4 1.? 2
Log Cot
FIG. 2. Lack ofDNA renaturation under R-loop hybridization conditions. (0 ---0) 50,000 cpm of
'H-labeled HSVDNA(specificactivity, 300,000cpm/p) wasmixed with20pgofunlabeled HSVDNA which had
been shearedtoanaveragesizeof14kilobases(30). TheDNAwasprecipitated and dissolvedin 200,ulof
R-loopbuffercontaining75%formamideand0.2 MNa+.This mixturewasthenheatedto78°Cfor10minand
incubatedattheR-loopingtemperature(56°C) forvariousCot values (1Cais100pgofDNAperml incubated
for 1 h).Aliquots(10-,ul) wereremoved, diluted into 0.35
Al
ofSI digestion mix(25), and digested withSlnuclease todetermine theamountofDNArenaturation. (X-X) Aparallel experimentwascarriedout at 30°Ctoshow thattheHSVDNAdoes,infact,renature attheproperreannealingtemperature.
26 m
0
L
E
c u L
s
L
0
0 P
E D
N
T
E R
A
L %'
18
16 14 12
18
8
6
4
2
e 16 20 36 46 56 66 76 86 90 166
[image:8.499.105.397.75.307.2]GENOME LENGTH (M)
FIG. 3. Random-order histogram ofthe positions of R-loops formed in HSV-1 (KOS) DNA by early
polyribosomal RNA. Fifty-one R-looped, intact HSV-1 DNA molecules were aligned without regard to
orientationtoproducethishistogram(see text).Intactmoleculeswerethosemeasuring28+ I XRFIIDNA unitsinlength.
VoL. 27, 1978
188 99
63
s
I N
G L E S T
R
A H D E D D
N
A
88
78
60
58 40 30
20
16
6
on November 10, 2019 by guest
http://jvi.asm.org/
[image:8.499.106.405.394.616.2]64 STRINGER, HOLLAND, ANI) WAGNER
5 1 5 25 35 45 50 55 65 75 85 95
95 85 75 65 55 SO 45 35 25 1 5 5
75 65 55 45 35 4o 25 1 5 5 85 95
95 85 5 15 25 4O 35 45 55 65 75
5 1 5 25 35 45 4O 55 65 75 95 85
85 95 75 65 55
JO
45 35 25 1 5 575 65 55 45 35 30 25 1 5 5 95 85
[image:9.499.57.450.56.296.2]85 95 5 15 25 70 35 45 55 65 75
FIG. 4. Distributionof regions of the HSV-1genome inarandom-orderarrangement. Theeight possible
arrangementsofthe HSV-1genomeareshowndiagramatically.Numbers denote percentageofgenomelength
ofaprototypical arrangement. Cross hatches denote thelong segment-short segment junction. The dotted line
shows thetwolociofaprototypical arrangement of HSVDNA which will contributetothecenterofa
random-orderhistogram.
C;I HIIO | HJ HA HK HL HC HN HG
XG xc XF XE XA
HH HIO HJ HA dSK HL HB HN HM
-LONG SEGMENT _SHORT SEGMENT.
HD HL HK HA HJ HIO HF HN HG
XD XE XF XC XB
HlD HL HK HA HJ HIO HE HN HM
0
R D E R
1
0R D E R
FIG. 5. Sitesfor cleavage of HSV-1 (KOS) DNA by the restriction endonucleases Xba I and HindIIIas
determinedbySkare and Summers(21). Thefour arrangements oftheHSV-1genomearearrangedinto two
ordersonthe basisofthelong-segmentorientation. Thepositions of HindIIIsitesaremarkedbythe vertical
solid lines, those ofXba Ibyvertical dashed lines. The individualfragments (HA, XA, etc.) are named
followingtheterminology ofSkareand Summers (21). The sitesoftheHSV-1 (KOS) strain used hereare
identicalwith those describedbytheseauthorsexceptforthelackofaHindIIIsitebetweenfragmentsI and 0;theresultingcombinedfragmentis termed "HIO"(seethetext).
histogram.ThishistogramshowedR-loopstobe
scarce in DNA loci mapping approximately 30
and/or50% in from thelong-segmentend. Since
XbaIfragmentF could be orientedtoproduce
a non-R-looped region between 40 and 45% in
from thelong-segment end, andsince no early
polypeptides have been found to map in this
area (L. Morse, L. Pereira, B. Roizman,and P.
Schaffer, in F.Rappetal., ed.,ThirdInt.Symp.
onOncogenesisandHerpesviruses, inpress;N.
M.Wilkie,N. D.Stow,H. S.Maroden,V.Brown,
R. Cortini, M. C. Timbury, and J. H.
Subak-Sharpe, in F. Rappetal., ed., ThirdInt. Symp.
onOncogenesisandHerpesviruses,inpress),it
wasreasonabletotentativelyassumetheregion
of thegenome mappingfrom 40to 45% infrom
0 R D E R 1
0 R D E R 2
,1. viumol.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:9.499.59.448.360.506.2]A~~~~~~~~~~~~~~~~~~~~~~~~~~
-S~~~~~~~~~~~~~~~~~~~~~~~~.
o.%j7
41~~~~~~~~~~~~~~~~~~~~~~~b "49
FIG. 6. HSV-1 DNA restriction fragments R-looped with earlypolyribosomal RNA. The small circular
moleculesare
4LXRFII
DNA. The barindicates0.1 Lm.(A)Xba Ifragment F; this fragment is4.54XRFII
units(7.8gm)inlength. (B) HindIII fragmentA; thisfragmentis7.11XRFIIunits (12.3
pmn)
in length. (C)XbaIfragment G; this fragment is2.4
O.XRFII
units (4.15pin)inlength. Loops are shown with arrows indicatingthesingle-strandedDNA(5).
65
on November 10, 2019 by guest
http://jvi.asm.org/
[image:10.499.45.473.49.617.2]G F A
s * ~* ***muU 3
| |
'
|
-
:
-~~~~
*S
_________
mu*
EU.*- I s * **
]_~~~a
_________
_*LLJLL
33,s 3: - ml
sB '
a
lsU
* **
*
msa
G F
20 30 46 59 66
GENONE LENGTH (M) FIG. 7.
A
79 86 96 166
A.
1
5
16
15
26
25
B.
SS
54
46
42
36
30
24
18
12
6
6 N 0 L E C U L E S L 0 0 p E
D
I
I N T E R U
A L
X1
on November 10, 2019 by guest
http://jvi.asm.org/
MAPPl'ING
EARLY HSV-1 T'RANSCRIPTS 67 the long-segment end of the molecule to belargely devoid of R-loops. The validity of this
assumptionwasascertained from the results of orientationoperations based on the assumption asdiscussed further below.
If theorientationof the short segment of the
HSV-1genome is ignored,full-lengthHSV DNA
moleculescan bearranged intotwoorders
cor-responding to the two possible orientations of
the longsegment of the genome. These
proto-typicalarrangements areshowninFig.5asorder
1 and order 2. We undertook to sort out
R-looped intact HSV DNA molecules with regard
to the orientation of the long segment on the basis of (i) theR-loop distribution in the region of the DNA
represented by
Xba Ifragment Aand (ii) the strong suggestion ofa lack of R-looping in aregion nearthemiddle ofa
proto-typicalarrangementof the HSV DNA molecule.
A rarely R-looped region near the joint is
ex-pected to occurin all intact HSV DNA mole-cules,since thesequences nearthe
long
segment-shortsegment
joint
are the sameregardless
of the relative orientations of the two DNAseg-ments. The region of low
R-looping
whichap-peared tobe nearthemiddle ofa
prototypical
arrangementof the HSVDNAmolecule
would,
in anygiven
molecule,
occur oneither side ofthe center of rotation of the
long
segment,i.e.,
from 45 to50% for order 1molecules and from
30 to35% for order2molecules.
Moleculeswerescreened and
preliminarily
as-signedto eitherorder1 or order2onthe basis of their fitto
templates
whichdesignated
alack ofR-loopsintheregions
of theDNAdescribed above.R-loop
distributionhistograms
of thetwopopulations
wereprepared
andjuxtaposed
sothat the
regions
inthehistograms
representing
the DNA moleculelong
segments werecoli-nearly
aligned.
The success of thepreliminary
assignments
of molecules to order 1 or 2 wasdetermined
by
inspection
ofjuxtaposed
histo-grams for
mirror-image
symmetry. Thesym-metry was strong but could be
improved
by
realigning
molecules which couldbeorientedin morethan onewayonthe basisof theoriginal
template trial. The finalassignment
of HSV-1 molecules into orders 1and2is shown inFig.
8.Histograms of the
R-loop
frequency
in these moleculesareshown inFig.
9. The existence ofwell-definedmirror symmetry in Fig. 9 indicates
that the template-aided orientation procedure
hassuccessfully sortedoutthe differentgenome
arrangementsand oriented thesegroups of
mol-ecules with respect to the long and short
seg-mentsof the DNA. The histograms display
max-imum symmetry when the internal end ofthe
longsegment is placed 18% in from the
short-segmentend. This locationof the jointbetween
the long and short DNA segments is in good
agreementwith direct measurements of the joint
position (11, 20).
R-loop mapof the early genes of HSV-1
(KOS) determined from restriction
frag-ments. The distributions of R-loops produced
in seven restriction fragmentswere determined
toconfirm the R-loop dataobtained with intact
molecules. Fragments were chosen to cover as
much ofthe HSV-1genome as possible. HindIII
fragmentsA,
1-0,
J, K, and L and Xba Ifrag-ments A and G were used. As seen in Fig. 5,
these fragments together represent 99% of the
HSV-1genome.
All fragmentswith the exception of HindIII A
run assingle-component bandson agarosegels.
HindIIIfragmentA isamolar
fragment
whichcomigratesonagarosegels withaquarter-molar
fragment, HindIII fragment B. Preparations of
HindIII fragment A are therefore 80% pure.
Analysis of30 R-looped fragments showed 25
fragments which fit a pattern consistent with
thatexpected from analysis of (i)anoverlapping restrictionfragment,Xba F, and (ii) intact HSV DNAmolecules. The five HindIIIfragments in
whichR-loops occurredinarrangements
differ-entfromthe other25 weredesignatedasHindIII fragment B andwere not included in the
com-posite histogram shown in Fig. 10. Xba I
frag-mentsAandGwerealignedastheywereinFig.
7B.TheHindIII
fragments
werealignedtobest fit the loop distribution obtained from intact DNA in order1 (Fig. 9).Sinceonly fragments which contained R-loops
were scored when restriction fragment R-loop
patterns were determined, the relative peak
heights betweenfragmentsarenotindicative of the relative heights ofcorresponding peaks in
thehistogram derived fromintact DNA. There-fore, the composite histogram of R-loop
fre-quencyonthe HSV-1 genome shown inFig. 10
FIG. 7. (A)Graphicdisplay ofindividualXba Irestrictionfragments ofHSV-1(KOS)DNAR-loopedwith
earlypolyribosomalRNA. The restrictionfragmentsXba IA, F,and Gwererecoveredafterelectrophoretic
separationonagarosegelsandhybridizedwithpurified early HSVpolyribosomalRNA. Individual molecules
foreachfragmentwerealignedasdiscussed in thetext. The solid boxesrepresent thepositions ofR-loops.
The average lengthsforXba Ifragments A, F, and G were 9.6, 4.2, and2.4 4XRFIIunits, respectively,
corresponding to molecularweightsof33.8 x H06, 14.9x 106, and8.5 x 106(see thetext). (B) TheR-loop
frequency distributionsoftheHSVDNArestrictionfragmentsin(A).Theputative jointbetween thelongand
shortregionin XbaIfragmentAisat apositionnear orat82%genomelength. Thispartial histogramwas
usedtoalignintact HSV-1 DNA molecules andtoassignthemtoorder1ororder 2(see text).
Voi.. 27,1978
on November 10, 2019 by guest
http://jvi.asm.org/
68 STRINGER, HOLLANI), AND WAGNER
a a
.
-'
a
a
a
I
.
-.
U
Ia
a
N
OD 0. IV
COPl( OP
,,o\j ^r, 0% 0%
OD
n
N
.
-1U
(D w -4 ob
MNomCmN ~Co-SooB I
I .
1-40
:
:
a
a
I
a
. so
a
1-I
V MN -0
4C N N aSN
a -.a-.
-40
I
a
I)
M -
I
a
a a
a
.
N N _ _N _
_J _
-x
x)
x
x
1-Is
.
.
w
w
x
.
IV
Mv
M
x
W
JZ
X
w
x
La-0
x
m
Jz
Z
J. Viliol'.
.L.
C0.ti
CY
t4
*o
o6
00
.-
o
b*
L;qc
zt
0 0.
*) s
ci:to~
0. <
~
O+1E
n
;
on November 10, 2019 by guest
http://jvi.asm.org/
MAPP'ING EARLY HSV-1 ThANSCHIITS 69
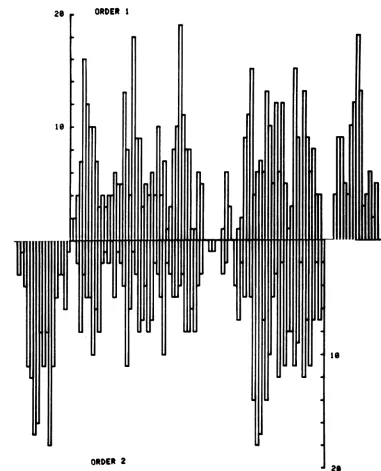
28 ORDER 1
Ie
I
ORDER 2
is
J
FIG. 9. Frequency distributionofearlypolyribosomalR-loopsformedin intactHSV-I(KOS) DNA. The
R-loopfrequency histograms ofHSV-1moleculesoforder1 andorder 2arearrangedsothatthe long regions
arecolinear. Theputative jointbetweenthelongandtheshortregionisattheverticalaxes.
mayonlybecompared with that for full-length corresponding to the order 1 non-R-looped
re-molecules of order 1 (Fig. 9) withrespect tothe gionmappingfrom 45to50% inFig. 9.
general pattern of peaks and gaps, not with DISCUSSION
respect to relative peak heights. The general
patterns are comparable. It is most significant We have obtained histograms detailing the
that HindIlI fragment A can be oriented to frequencydistributionalong thelengthofa
pro-produceanR-loop frequencydistribution which totypicalarrangement oftheHSVgenomeof
R-shows a lack ofR-loops in the region of DNA loops formed by polyribosomal RNA isolated
0
L E C
U
L E S
L
0 0 p E
D
N
N
T
E
R U
A
L
69
Vo1.. 27,1978
on November 10, 2019 by guest
http://jvi.asm.org/
[image:14.499.67.448.65.536.2]70 STRINGER, HOLLAND, AND WAGNER
60
II
L 54 XGI
HIO
|HJ|
HA
HK HL XA EC 48 ,
U L
E 42
S
L 36
0
0
P 38
E D
I 24
N
T 1
E 1
R
U
A 6
L
,% 8
8 Is 26 38 46 56 68 78 86 98 188
[image:15.499.112.394.67.280.2]GENOME LENGTH (IC)
FIG. 10. Composite R-loop distributioncompiled from R-loop patternsof HSV-1 DNA restriction fragments
hybridized to early polyribosomal RNA. Purified early polyribosomal HSV RNA was hybridized with
restriction endonucleasefragments,and25individualmoleculesofeachfragmentwereanalyzedasdescribed
in thelegendtoFig. 6 and in thetext.Data derivedfrom the R-loop distribution of Xba Ifragments A and G
and HindIIIfragmentsA, J,IO, K,andL,werecombinedtoformthecompositedistribution. The boundaries
ofthefragmentsareindicatedbythe solid lines. Thejoint region between the short and long segments is at
apositionnear or at82% genomelength.
early after infection
(Fig.
9and10).
Thealign-mentofintact
R-looped
HSVDNAwasaccom-plished by an iterative, subjective
procedure
aidedbytemplates
whichdesignated regions
of the DNAin whichR-loops
are rare.Thesetem-plates
wereconstructedonthe basis of(i)
infor-mationunambiguously
derived fromanalysis
of restrictionfragments
and(ii)
the inference from therandom-orderR-loop
distribution thatasec-ond
non-R-looped region
existednearthemiddle of theprototypical
arrangement of the HSVDNAmolecule. Thesuccessof this
approach
tocategorizing
andaligning
thedifferentarrange-ments of
R-looped
HSV DNA molecules wasestablished
by
three criteria:(i)
theR-loop
his-tograms of the two orders of HSV-1 molecule
show
mirror-image
symmetry when thehisto-grams arealignedsothatregions
corresponding
to the DNA long segments of each order are
colinear (Fig. 9); (ii) the junction between the
longand short DNA segments can beaccurately
located by analysis ofthe symmetry; and (iii)
the R-loop distribution of a prototypical
ar-rangement ofHSVDNAcanbe reproduced by
analysisofR-loops generatedonrestriction
frag-ments which unambiguously represent defined
portionsof the DNA(Fig. 10).
Ourspecificaim inthisstudyistodetermine
the arrangement ofearly viral genes from the
position ofR-loops. In a qualitative sense, the
positions ofearly genes are delineated by the general pattern of peaks in the R-loop
histo-gram.However,there are several factorswhich
complicate the interpretation of R-loop
histo-grams. First, relative peak heights inanR-loop
histogramare afunctionofmore than the
rela-tive concentrations ofdifferent RNAspeciesin
the reactionmixture. TherateofR-loop
forma-tionat any one sitedepends solelyon the
con-centration ofcomplementaryRNA (27); butthe
relative rates ofR-loop formation at different
sites are dependent not only on the relative
concentrations of thetwocomplementaryRNA
species,but also on theirbase composition.The rateofR-loop formation declinesrapidly asthe
reaction temperature deviates from the
Tm
ofthe sequence. Sites which differ by 6% inguanine
plus cytosine (G+C) content will exhibit a two-folddifferenceinR-looplevelseven thoughthe
concentrations of their complementary RNAs
are the same (27). Thus, the relative peak
heightsinFig. 9 cannotbe interpreted as
com-pletely indicative of the relative abundance of
different viral RNAspecies. Around 90% of early
viral RNA isolated frompolyribosomes
hybrid-izes to viral DNA withsingle-component
kinet-ics (23). Since RNA classesthat differ two- to
threefold in abundance would have exhibited
readily detectable multicomponent
hybridiza-tion kinetics (25), the two- to threefold
differ-,J. Viliol'.
on November 10, 2019 by guest
http://jvi.asm.org/
MAPPING EARLY HSV-1 TRANSCHIPT'S 71
encesin themajor R-loop peak heightsseen in
Fig.9 areprobably primarilydue todifferences
in base composition and not to differences in RNAspecies abundance.
While base composition can definitely influ-enceR-loopfrequencies, its effect is insufficient toexplain all thegapsandareas of rareR-loops
which appear in the R-loop histogram. This
conclusion is basedonthefollowing. (i) Notall regions of the HSV genome with higher than
average G+C content exhibit low R-loop
fre-quencies. Comparison of Fig. 9with partial de-naturation mapsof HSV DNA (7, 10) reveals that whilesomehigh-G+C regions exhibitalow frequency ofR-looping, R-loops occur athigh frequency in thehigh-melting regions at 5 and 67% of thegenomelength. (ii) The region
with-outR-loops which extends from43 to50% of the genomelength in Fig.9includes low-meltingas
wellashigh-meltingsequences (7). (iii) Russell and Wilkie (personal communication) have
re-cently directly determined the G+Ccontent of
many HSV-1 DNA restriction fragments by nearest-neighbor
analysis.
The DNA fragment correspondingtotheregion running from46 to50% of thegenomelength lacks R-loops (Fig.9),
but exceeds theaverageG+Ccontentof HSV-1 DNA by only 2%. According to the data of Thomas et al. (27), if significant amounts of RNA
complementary
tothisregion of theHSV-1 genome were present onpolyribosomes early
after infection, we would expect to see 70% as many
R-loops
in this region as occur inse-quencescontaining theaverage amountofG+C
forHSV-1 DNA.
Thereis, however,oneregion of HSV-1DNA
lacking
R-loops
in whichG+C contentmay be asignificant
factor.Nearest-neighbor analysis
(Russell
andWilkie, personal
communication) has showntherepeated
sequenceswhichbound the shortunique
segmenttocontain thehighest
amount ofG+C in the HSV-1 genome, 72.8%.
The dataof Thomas et al. (27)
predict
that, ifcomplementary
RNA concentrations wereequal, R-loops
intherepeated
sequencesofthe short segment would be 30% ashigh
as the frequency ofR-loops
formed in sequencescon-taining the averageamount ofG+C for HSV-1
DNA. Therefore,
R-loops
in theshort-segmentrepeated
sequences may notaccurately
reflectthe amount of early mRNA transcribed from
this part of the viralgenome.
Just as R-loop histogrampeak heights must
be
interpreted
as afunctionofmultiple
factors, R-loop histogram peak widths must beinter-preted in the
light
of thefollowing
considera-tions. The major component of early viral
mRNA is
homologous
to20 to 25%of the viralDNA,with the best value on the orderof 22 to
23% (23, 25). Since the length ofduplex DNA
involved in R-loops shouldbe twice the amount
ofviral DNA driven into hybridbyearlyRNA,
thisRNA should produce a histogram with
ma-jor R-loop peaks covering40 to50% of itslength.
However, measurement inaccuracies tend to
broadenR-loop peaksand increase the portion
of the histogram covered by R-loops. The
pri-mary source of measurement inaccuracies in
these
experiments
is thelength uncertaintyin-herentinworking with large linearDNA
mole-cules.Apopulation of linear DNA molecules
will
exhibit a rangeof lengths, evenin referencetointernal
length standards. Sincethere is noab-solute length criterion, molecules falling within
a narrow range of lengths are accepted a
full
size. HSV-1
(KOS)
DNA hasameanmolecular length of28.1±0.74XRFII
DNAunits (30). Inthis study,wehave acceptedasintactany mol-ecule measuring between 27 and 29
4XRFII
DNA units. Mostof these moleculeswereintactHSVgenomes, andanyvariation inlengthwas
corrected by
normalization
to a standard size. However,someof themoleculesacceptedasfull size may in fact contain less than a full HSV genome. TheR-loops
inthesemolecules willbelaterally
displaced from their true positions, thereby tendingtobroaden thepeaksin the R-loop histogram.The degreeto whichmeasuring inaccuracies broaden R-loop peak widths can be estimated from
R-loop
studieson adenovirus hexonmes-senger RNA. The size of
R-loops produced
bypurified hexonmessageis known (2). Individual R-loop sizes correlate well with the size of the
message, butthepeak inthe R-loop histogram
corresponding to the hexon gene is two times widerthan the hexonmessagewhen the
peak
is measuredattheabscissa. The hexonpeak width isequaltothatexpected
from the hexonR-loop
size when thepeak
ismeasuredatone-third its height (17). It is therefore reasonabletoregard
thepeak
widths atone-third theirheights
as amore accurate measure of the amount of the
genome
represented
asR-loops
than thepeak
widthsatthehistogram abscissa.
Whenthepeak widths of the order1 ororder
2molecules of HSV-1 in
Fig.
9aremeasuredatone-third
peak height,
the totalamount ofthegenome
represented
asR-loops
is about 50%ofthelength of the HSV-1genome.Thismaybe a
high
estimate
of theactuallength
of thegenomeabletoform R-loopswith the
major
componentofearly
polyribosomal
RNA, since the width of theR-looppeakin theshort segment is inflatedbythe short segment
being
represented
inbothitspossible orientations.With this
qualification,
the extent of the HSV-1 genome
forming
R-loopswith the
high-concentration
classofearly
Vo.. 27,1978
on November 10, 2019 by guest
http://jvi.asm.org/
72 ST'RINGER, HOLLAND, AND WAGNER
polyribosomal HSV RNA is ingood agreement
with valuespredicted by DNA-RNA
hybridiza-tionexperiments.
Apart from a loss ofresolution, the factors
that complicate the interpretation of R-loop
datain quantitative termsdo notinfluence the
qualitativesignificance of the data. The resolu-tion reported here issufficient to establish the
distribution ofearly viral genesand tosuggest
theirspecific locations. The patterns generated
by R-looping intact HSV DNA with
polyribo-some-associated viral RNA present early after
infection indicate that this class of viral mRNA is transcribed from genes distributed through
the genome.
Inthe short segment of theR-loophistogram,
there appears a single peak which is roughly
symmetrical about the centerof the short
seg-mentand extendsslightly into the inverted
re-peat sequences which bound this part of the
DNA. This symmetry isexpected,sincewehave
notdiscriminated betweendifferentorientations
of the short segment. It is apparent that the
unique sequences in the short segment are
ex-tensively transcribed. Sincewe are not dealing
with asingle orientation of the short segment,
theR-loop histogramdoesnotreveal thedetails of the distribution of the early genes in this
segment.However, wecaninfer fromanalysisof
the individual molecules shown inFig.8thatthe
earlygenesintheuniquesequencesof the short
segmentarenotcompletely confinedtooneend
of the segment, but are distributed across its
length.
The distribution ofR-loop peaks in the long
segment of the DNA is complex. Atthe lowest
level ofresolution, the segment is divisible into
twolargeareasoftranscriptionalactivity. These
regions span the length of the long segment,
indicating that the early genes which occur in
this segmentaredistributedalongitslength and
are notcompletely contiguous. The resolution of
thelong-segment R-loop pattern is increased if
the pattern is examinedatone-third the
individ-ualpeakheightsorifloopsoccurring inonly one
molecule (4%) are considered a "background"
,1. Vilmlwo.
level. Severallarge R-loop peaks arediscernible,
two ofwhich fall in the long-segment repeated
sequences while the restoccurin theunique part
of thelong segment. Some of these peaks
prob-ably represent more than oneearly gene, since
they are much larger than the average viral
message size, which is 20S, corresponding to
2,000 nucleotides in length (18, 23). Although
theearly genesare clearlynot completely con-tiguous, they may be clustered in groups. Broad
R-loop peaks may indicate where tightly
clus-teredearly genesoccur.
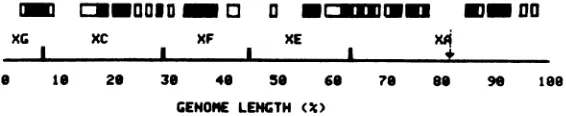
The R-loop data ofFig. 9 are interpreted in
Fig. 11 as an early transcription map of the
prototypical order 1 of HSV DNA. Fourteen
regions of the DNA are R-looped at relatively
high frequencies. Theseregionsrepresent35% of
the total DNAlength; R-loopsoccurwith
mod-erate frequency in regions representing an
ad-ditional 15% of the HSV-1 DNAlength. Of the
50% of the DNA length in which R-loops are
infrequent, 35% show some R-looping and 15%
show no R-loopsat all. Some of the
contribu-tions tothe regions with low numbers ofloops
areduetopositional uncertainties,while others may well be a result of the low-concentration speciesof viral RNAfoundearlywhich
hybrid-izes to more viral DNA than does the major
componentofearly RNA.
While the transcription map in Fig. 11 does
not resolve the locations of individual early
genes, it definitely differentiates regions of the
DNA that are notrepresented as major
compo-nentsofearlymRNAfrom regions of DNA that
are represented as early mRNA in high
abun-dance. As a moderate-resolution transcription
map,then, R-looping definitely establishes that
theearlygenesarescattered about the genome. This conclusion agrees with results obtained by
liquid hybridizationofasubset of the totalearly
viral transcripts (a HSV RNA) to different
re-strictionfragments(13).Furthermore, Clements
and co-workers(6),using the blot hybridization
technqiues ofSouthern (22), have shown that
early HSV transcripts hybridize to regions
throughout the HSV-1 genome. Our resultsare
IuOOuO
oD Omm
mm
noxc xc
1 XF I XE I
Oe
6 is 26 36 40 56 60 76 86 96 168
GENOKE LENGTH (Ic)
FIG. 11. Schematic summary ofthe early polyribosomal RNAR-loopdistribution on the order 1prototypical
arrangement ofHSV DNA. The positions of Xba I endonuclease cleavage sites are indicated. The arrow marks theposition ofthelongsegment-short segmentjunction. Closed and open boxes delineate regions of
highandmoderateR-loopingasdetermined fromanalysis of the orderIhistogram displayed in Fig. 9. (-)
Regionsin which greater than 8% of themolecules bearR-loops; (O) regions inwhich4 to8% of the molecules
bearR-loops.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:17.499.112.395.544.602.2]MAPPING EARLY HSV-1 TRANSCRIPTS
also consistent with the results of polypeptide
mapping studies which have shown the early
genes to be distributed throughout the HSV
genome (Morseetal., inpress; Wilkieet al., in
press). When the histogram is interpreted as a
higher-resolutiontranscription map,significant
regions of the DNA are revealed as not being
represented as large amounts of mRNA early
after infection.
ACKNOWLEDGMENTS
This research was supported by Public Health Service
grantCA-11861fromtheNational CancerInstitute andan
institutionalgrantfromtheAmericanCancerSocietytoJ.R.S. J.R.S.wassupported bypredoctoral traininggrant CA-09054 from theNational CancerInstitute.
Wethank S. Malan forprovidingcomputerexpertise,M. Rice for excellent technical assistance, and N. Wilkie for communicatingnot-yet-publisheddata. Thehelpof L. Howell is alsogratefully acknowledged.
LITERATURE CITED
1. Aloni, Y., R. Dhar, 0. Laub, M. Horowitz, and G.
Khoury.1977.Novel mechanisms forRNA maturation. Theleadersequences ofSV40 mRNAare not
tran-scribedadjacentto thecoding sequences. Proc. Natl. Acad.Sci. U.S.A.74:3681-3690.
2. Berget,S. M., C. Moore,and P. Sharp.1977.A
noncom-plimentarysequenceatthe5'-terminus of late adeno-virus 2 mRNA. Proc. Natl. Acad. Sci. U.S.A. 74:3171-3175.
3. Casey,J., andN.Davidson. 1977. Rateofformation andthermal stabilities of RNA:DNA and DNA:DNA duplexesathigh concentrations of formamide. Nucleic Acids Res.4:1539-1552.
4. Chow, L., R. E. Gelinas, T. R. Broker, and R. J. Roberts.1977.Anamazingsequencearrangementat
the5'endsofadenovirusmessengerRNA.Cell 12:1-8. 5. Chow, L., J. M. Roberts, J. B. Lewis, and T. R. Broker.1977. AmapofcytoplasmicRNAtranscripts from the adenovirus type 2, determined by electron microscopyofRNA:DNAhybrids. Cell 11:819-836. 6. Clements,J.B., R. J. Watson, and N.M.Wilkie.1977.
Temporal regulation of herpes simplex virus type I transcription: location of transcripts on the viral ge-nome.Cell12:275-285.
7. Delius, H., andJ. B. Clements. 1976. Apartial denatur-ation mapofHSV-1DNA.J.Gen.Virol.33:125-133. 8. Eagle,H. 1959. Aminoacid metabolism inmammalian
cellculture. Science130:432-437.
9. Frenkel, N.,and B.Roizman. 1972. Ribonucleic acid synthesis incellsinfected withherpessimplex virus: controls oftranscription andRNA abundance. Proc. Natl.Acad. Sci.U.S.A.69:2654-2658.
10.Hayward, G. S.,N.Frenkel, andB. Roizman. 1975. Anatomy ofherpessimplexvirusDNA: strain
differ-encesandheterogeneityinthe locations ofrestriction endonuclease cleavage sites. Proc. Natl. Acad. Sci. U.S.A.72:1768-1772.
11. Hayward, G. S.,R.J.Jacob,S. C.Wadsworth, and B. Roizman. 1975. Anatomy ofherpessimplexvirus DNA:evidence for fourpopulationsofmolecules that differin therelativeorientations oftheirlongand short components. Proc. Natl. Acad. Sci. U.S.A. 72:4243-4247.
12. Honess, R.W., andB. Roizman. 1973.Identification andrelative molarratesofsynthesisofstructuraland nonstructural herpesviruspolypeptidesintheinfected
cell.J.Virol.12:1347-1365.
13.Jones, P. C., G. S.Hayward, and B.Roizman. 1977. Anatomy of herpes simplex virus DNA:aRNA is ho-mologous tononcontiguous sites in both L and S com-ponents of viral DNA. J. Virol.21:268-276.
14. Kozak, M.,and B.Roizman. 1974. Regulation of herpes virus macromolecular synthesis: nuclear retention of nontranslated viral RNAsequences. Proc. Natl. Acad. Sci.U.S.A.71:4322-4326.
15. May, E., J. V. Maizel, and N. P.Salzman. 1977. Map-pingtranscription sites of simian virus 40-specific late 16S and19SmRNA byelectron-microscopy.Proc. Natl. Acad.Sci. U.S.A. 74:496-500.
16. Meisner, H. C., J. Meyer, J. V. Maizel, and H. West-phal. 1977. Visualization and mapping of late nuclear adenovirus RNA.Cell10:225-235.
17. Meyer, J., P. D.Neuwald, S.P.Lai, J. V. Maizel, Jr., and H.Westphal. 1977.Electron microscopy of late adenovirus type 2 mRNAhybridized todouble-stranded
viralDNA.J. Virol. 21:1010-1018.
18.Moss, B., A.Gershowitz,J.Stringer,L.Holland,and E.Wagner. 1977. 5'-Terminal andinternal methylated
nucleosides in herpes simplex virus type 1 mRNA. J. Virol.23:234-239.
19. Palmiter,R. D. 1974. Magnesiumprecipitation of
ribo-nucleoprotein complexes. Expedient techniques for the isolation ofundegraded polysomes and messenger ri-bonucleic acid. Biochemistry13:3606-3614.
20.Sheldrick, P., and N. Berthelot. 1974. Inverted repeti-tions inthechromosome of herpessimplexvirus.Cold Spring Harbor Symp.Quant. Biol. 34:667-678. 21. Skare, J., and W. Summers. 1977. Structure and
func-tion ofherpes virus genomes:EcoRI,XbaIand HindIII
endonuclease cleavage sites on herpes simplex virus type 1 DNA.Virology76:581-595.
22. Southern,E. M. 1975. Detection ofspecificsequences among DNAfragmentsseparatedbygel electrophore-sis. J. Mol.Biol. 98:503-517.
23. Stringer, J., L.Holland,R.Swanstrom, K.Pivo, and E.Wagner.1977.Quantitationofherpessimplexvirus type1RNA ininfected HeLacells.J.Virol. 21:889-901. 24. Sutherland,B., M.Rice, and E. Wagner.1975. Xero-dermapigmentosumcellscontainlowphotoreactivating
enzyme levels. Proc.Natl. Acad. Sci. U.S.A. 72:103-107. 25. Swanstrom,R., K. Pivo, and E. Wagner. 1975. Re-strictedtranscription of the herpessimplex virus ge-nomeoccurringearlyafter infection and in thepresence ofmetabolic inhibitors.Virology66:140-150. 26. Swanstrom, R., and E. Wagner. 1974. Regulationof
synthesisofherpessimplextype1virusmRNAduring
productive infection.Virology60:522-533.
27. Thomas,M., R. L. White, and R. W. Davis. 1976.
Hybridizationof RNA todoublestranded DNA: for-mation of R-loops. Proc. Natl. Acad. Sci. U.S.A. 73:2294-2298.
28. Wagner, E., B. Roizman,T. Savage, P. Spear, M. Mizell, F. Durr, andD.Sypowicz.1970. Characteri-zation of the DNA of herpesviruses associated with Lucke adenocarcinoma of the frog and Burkitt
lym-phoma of man.Virology42:257-261.
29. Wagner, E., R. Swanstrom, andM. Stafford. 1972.
Transcription of theherpessimplex virusgenome in humancells. J. Virol.10:675-682.
30. Wagner, E.,K.Tewari,R.Kolodnor,and R.Warner. 1974. Themolecularsizeoftheherpessimplexvirus type1genome.Virology57:436-447.
31. Wagner,E.K.,R.I.Swanstrom,M.Rice. LHowell,
andJ.Lane. 1976.Variationinthemolecularsize of the DNA fromcloselyrelated strains oftype 1herpes simplexvirus.Biochim. Biophys.Acta 435:192-205.
VOL. 27,1978 73