0022-538X/81/010080-12$02.00/0
Differences
in
the
Subpopulations
of the Structural Proteins
of
Polyoma
Virions and Capsids: Biological
Functions of
the
Multiple
VP1 Speciest
JOSEPH B.BOLEN, DAVID G. ANDERS, JANINETREMPY,AND RICHARD A. CONSIGLI* Section of Virology and Oncology, Division of Biology, Kansas State University, Manhattan,Kansas 66506
The structural proteins of polyoma virions and capsids were analyzed
by
isoelectricfocusingand sodium
dodecyl
sulfate-polyacrylamidegel
electrophore-sis. Polyoma virion
VP,
wasfoundtobecomposedof six distinctspecieswhichhad pI'sbetween pH 6.75 and 5.75. Polyoma capsid
VP,
was found tocontainfourspecieswithpI'sbetweenpH6.60and5.75.Thedifferent formsofvirionand
capsid
VP,
appeared to be generated by modifications (phosphorylation andacetylation) oftheinitial translationproduct. The most basic of thevirion
VP,
species (pI, pH 6.75) was absent in capsids and was found to be exclusively
associated with the viral
nucleoprotein complex.
Three of the virionVP, species
and three of the
capsid
VP, species
were found in capsomerepreparations
enriched forhexonsubunits.Two
VP, species
werespecifically
immuneprecipi-tated from virionswithhemagglutination-inhibiting antibodies. These two
VP,
species were common to both virions and capsids. Polyoma virions, but not
capsids, possessed a
single
VP,
species which was immune precipitated withneutralizing antibodies. Bothvirionand capsid VP2werefound tohave pI'sof
approximately pH5.50.Virion VP3 hadapIofapproximatelypH 7.00, whereas
capsid VP3had apIofapproximately pH6.50.
The late region ofpolyoma virus DNA
con-sistsof2,366 basepairswhich code for thevirus
structuralproteins
VP,,
VP2,and VP3 (9). Themajor capsid protein
VP,
is encoded at the 3'end of the lateregion and is translated froma
16SmRNA,whereas the minorcapsidproteins
VP2 and VP3 are encodedat the 5' end ofthe
lateregion andaretranslated from 19S and18S
mRNA's
(10).
These threeproteins comprise thestructural unitswhichareresponsiblefor
main-taining thestructuralintegrityofthevirus,but
theyarealsoessentialasreceptor sites for
infec-tionofhost cellsandashemagglutininsfor
ag-glutination of erythrocytesand areimportant in
DNApackagingand virusassembly.
In a previous report from this laboratory it
wasdemonstratedthatpolyoma capsids,which
are devoid of DNA and histones, did not com-pete with polyoma virions for specific binding sites on thesurfaceof mousekidneycells(MKC)
and were unable to inhibit virion infection of
MKC. Capsids could,however, block virion
ad-sorption toguineapig erythrocytes (GPE). Vir-ion adsorption to and infection of MKC were found to beindependentof theability of virions
toagglutinateGPE (2).Morerecently,wehave
been able to separate neutralizing antibodies
t Contribution no.81-60-j from theKansas Agricultural ExperimentStation,KansasStateUniversity.
from
hemagglutination-inhibiting
antibodies inantisera directed
against
themajor
structuralproteinVP1 (3).
Itis clear that themajorvirusprotein
VP,
ofpapovavirusesdoesnotrepresent asingle
poly-peptidebutrather is composed ofanumberof
polypeptide species
which have different pI's(11, 16, 17). However, thefunctionsofthe var-ious
VP1 subspecies
have remained obscure.Throughthe useof theantisera described above,
along with our understanding of the
biological
and biochemical differences between polyoma
virions andcapsids, we have nowbeen able to
assign functionstothedifferent species of
poly-oma
VP,.
MATERIALS AND METHODS
Celland viruspropagation. The preparationof
primary cultures ofMKC has been described (18).
Wild-type large-plaque polyoma virus was used to
infectcellsat amultiplicityofinfection of 10. Infected
cultures were maintained in serum-free
Dulbecco-modifiedEaglemedium(13).
Viruspurification. Virionswerepurifiedfrom the
infected-celllysateasdescribedpreviously(13). CsCl
gradients used to purify the virus were prepared as
describedby BrunckandLeick (8)and were described in greater detail previously (6, 7). Polyoma capsids
were purified from infected-cell lysatesasdescribed
previously (3).
80
on November 10, 2019 by guest
http://jvi.asm.org/
FUNCTIONS OF POLYOMA VP, SPECIES 81
Preparation of radioactively labeled virus. Purified polyoma virions orcapsids were labeled in vitrowith'25I,usingchloramine-T or the Enzymobead
method asdescribed previously (3). The preparation
of'H-amino acid-labeled polyoma virus has been de-scribedpreviously (14), as has the preparation of '2P-labeled virus (4). [14C]acetate-labeled virus was pre-pared by maintaining the infected cell cultures in serum-free culture media with 5pCi of['4C]acetate
per ml.Methyl-3H-labeledviruswas prepared by main-taining the infected cell cultures in serum-free, methi-onine-free Eagle medium with 20uCi of S-[methyl-'H]adenosyl-L-methionine.
Isolation of 48S DNA-protein complex and 18S, 12S, and 5S capsomere subunits. Radioactively labeled virions or capsids were incubated with
ethyl-ene glycol-bis(l-aminoethyl ether)-N,N'-tetraacetic acid (EGTA) and dithiothreitol (DTT), and the virus corecomplex and capsomere subunits were recovered from 5 to 20% or 10 to 30% sucrose gradients as describedby Bradyetal.(7).
GuHCI chromatography. Radioactively labeled polyoma virions were treated with 6 M guanidine-hydrochloride (GuHCl) and0.01 Mmercaptoethanol
asdescribedpreviously (4). The virion proteinswere
thenseparated bygelfiltrationthroughSephacryl
S-300 atpH5.0.
Immuneprecipitation. '25I-labeled,
GuHCl-puri-fied VP1 (100pl) wasincubated with25p1of
antihe-magglutinin,antireceptor, ornormal rabbit immuno-globulin G,allat 10pg/ml,for4hat room
tempera-ture. Afterincubation, 25p1of normal rabbit immu-noglobulin G (10 mg/ml) wasadded, followed by50
pl of goat (immunoglobulin G fraction) anti-rabbit
immunoglobulin G (20 mg/ml). The mixtures were
then incubated overnight on ice, and the immune precipitates were pelleted by centrifugation and washed three times in 10mM Trisbuffercontaining 0.25% TritonX-100and 0.15M NaCl(pH 7.8).This
protocolwasdevelopedfor immuneprecipitation re-actions in which both the antigen and antibodyare present inlowquantitiesand avoids theuseof
Staph-ylococcus aureus protein A, which leads to large
amountsofnonspecific precipitationofpolyomaVP,
(J.B.B.,unpublished data).
Two-dimensionalIEFand SDS-PAGE. Electro-phoresiswas done bythe method of O'Farrell (15). Radioactively labeled virions or capsids were
dis-rupted in2% sodium dodecyl sulfate (SDS) and 5%
mercaptoethanol for2minat90°C.Aftercoolingto roomtemperature,solidultrapureurea wasaddedto 10M, Nonidet P40wasaddedto 2%, pH 3.5 to 10
ampholines wereaddedto 2%, andmercaptoethanol
wasaddedto5%. Theampholinesusedfor the isoelec-tricfocusing(IEF;combiningpH3.5to10andpH5to 8ampholines)yieldedveryshallowpH gradients be-tweenpH5and8.pI'sweredeterminedbyslicingthe
focusing gel immediately after electrophoresis and
measuringthe pH of the individual gelsegments in distilledwater. FortheSDS-polyacrylamide slab
gel
electrophoresis (PAGE) (when second-dimensional analysiswasrequired),the
equilibrated
IEF tubegel
wasembeddedinagarose abovethestacking gel.
Al-ternatively, the focusinggels containing
l"I-labeled
samples were first sliced into 2-mm segments, the
radioactivity of each segment was determined by counting in a gamma counter, and the peak segments were then placed in wells of a conventional SDS-polyacrylamide slab gel as described previously (7).
Quantitative assays. Cesium chloride densities were determined bythe refractive index with a
refrac-tometerand calculated by using the equation of Vin-ograd and Hearst (19). Protein concentrations were determinedby theBio-Rad protein assay, using bovine
serumalbuminas astandard. Radioactivity was quan-titated in atoluene-Triton (3:1)scintillation fluid with
aBeckmanLS-233 liquid scintillation counter. When
'Hand14C or'Hand32P isotopes were counted in the samesample, the results were corrected for the overlap of'4Cand'2Pinto the'Hchannel and for quenching. Samples labeled with fl-emitters were prepared for counting by incubating the slices with Protosol (New England Nuclear Corp.) scintillation fluid (1 liter of toluene,4g ofOmnifluor, 30 ml of Protosol; 5 ml per vial) for24 hat37°C. The vials were cooled to room temperature beforecounting.
RESULTS
IEF of
"2'I-labeled
virions and capsids.IEF ofradioactively labeledpolyoma virions and
simianvirus40(SV40)virions has been reported
(11, 16, 17).However, in these reports the virus
proteins werelabeledby allowinginfected cells
toincubate in the presence of14C-or
:H-amino
acids,
""Pi
or [;5S]methionine. We were inter-ested in determining the pattern of IEF of in vitrochloramine-T-'251I-labeled
polyoma virionsand comparing the polypeptide distribution of
thevirionswithsimilarly labeled polyoma
cap-sids. This type of in vitrolabeling, whichlabels
all of the virion andcapsid
polypeptides,
yieldedspecific activities generally in the range of5 x
106
to 10 x 106 cpm/,ug of virus protein usingliquidscintillation spectroscopy and 1 x
108
to5 x
10'
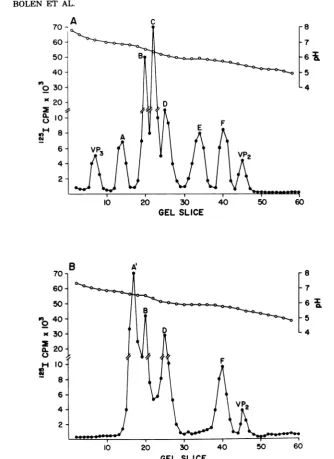
cpm/,ugofvirus protein using a gammacounter to quantitate the radioactivity. Figure
1A shows the IEF distribution of
'25I-labeled
virions. Atotalofeight polypeptide specieswere
resolved by
slicing
the IEF tubegels, countingthe segments inagamma counter,andresolving
the portions corresponding to the peaks in an
SDS-polyacrylamide
slabgel.
VirionVP3
wasfoundtohave pI ofpH 7.0, and virionVP2had
apI of5.5.Sixspecieswithmolecularweightsin the VP, range (42,000 to 45,000 [42Kto 45K])
werefound andwere
assigned
letterdesignationsA through F. The most abundant virion VP,
species (C) had a pl of pH 6.3, which is in agreementwith the results of Hewicketal. (11).
Our study revealed that the pI's of the other
VP, specieswere as follows: A,pH 6.75; B, pH
6.5; D, pH6.1; E,pH5.9; F, pH 5.75. Figure 1B
shows the IEF distribution of
'25I-labeled
cap-sids. Only five polypeptides species werere-solved. Two-dimensionalanalysisof these
poly-peptidesrevealed thatcapsid VP2hadapI
iden-VOL. 37,1981
on November 10, 2019 by guest
http://jvi.asm.org/
82 BOLEN ET AL.
10 20 30 40 50 1
GEL SLICE
A
10 20 30 40 50 60
GEL SLICE
FIG. 1. IEFof1251-chloramine-T-labeled virions andcapsids. Equalconcentrationsof iodinated polyoma virions (A) orcapsids (B)weresubjected toIEFasdescribed in the text. Thegelsweresliced into2-mm segments,and theradioactivityin each slicewasquantitatedinagammacounter. Themolecularweights of eachof the peak fractionswereestimatedbysubsequent electrophoresisinanSDS-polyacrylamide gel, using
iodinated virionproteinsasstandards.
tical tothat ofvirion VP2, whereas capsid VP:}
hadapI ofapproximately 6.5.FourcapsidVP,
species were found (designated A' through F)
with thefollowingpI's: A', pH 6.6; B, pH6.5;D, pH 6.1; andF, pH5.75.Theletter designations
for the capsid VP1 specieswere given as those
corresponding to species found in virions. The only capsid VP, species which did not have a
correspondingvirion VP1 species was found at
pH 6.6 and was designated A'. These results
show thatpolyoma capsidslackthree oftheVP1
species found inpolyoma virions (A, C, and E)
andpossessaVP3withan alteredpI.Itshould also be noted that under thepHrangesused for these experiments, polyoma histones did not enterthe IEFgels.
[8
-61
_5L4
14
60
-8
-67
-5
L4
60 50 -40 O 30
X 20
2 I
X 0-U
in 8
6
4.
2-
70-60
50
-00
40X
30-a.
20'H 10
C
8 6
4,
2
B
on November 10, 2019 by guest
http://jvi.asm.org/
[image:3.496.76.411.64.524.2]VOL.37,1981~~FUNCTIONSOF POLYOMA
VP,
SPECIES 83Two-dimensional
analysis
of"'I1-labeled
virions and
capsids.
Conventional O'Farrell(15)
two-dimensionalelectrophoresis
of"'M5-a-beled virion and
capsid
preparations
is showninFig.
2.Whentheautoradiographs
wereexposed
forshort
periods (Fig.
2AandC),
theVP,
sub-species
could be observed with some success.However,
it is clear thatduring equilibration
ofthe IEF
gels,
diff-usion of the focusedspecies
occurred which
prevented
the observationof thedistinct
patterns
observed whenthesingle
IEFdimension was
analyzed.
Thisproblem
wasun-doubtedly
complicated by
the1"5I labeling
of theproteins,
whichtends tofog
the filmsurrounding
theactual
polypeptide
spot.
L-onger
exposureofthe
autoradiographs clearly
showed themigra-tionofthe minor
proteins
VP2
andVP3(Fig.
2Band
D).
Both virion andcapsid
VP2 and VP:3migrated
as doublets in theautoradiographs
shown.However,
thedegree
ofseparation
of theVP2
andVP3
doublets variedconsiderably
be-tween
experiments. Again,
thealteration in themigration
ofcapsid
VP3 in IEF dimension canbeseen.
Differential
labeling
ofvirion andcapsid
proteins
with"'I1. Experiments
wereper-formedto compare theIEF
patterns
of virionsand
capsids
labeled in vitroby
the chloramine-T andEnzymobead
techniques.
In vitro iodina-tion of virions andcapsids
with chloramine-T labels both internal and extemnalproteins.
Invitro iodination of these virus
particles
withEnzymobeads,
however,shouldonly
labelexter-nal
proteins
withexposed tyrosine
residues.In-deed,
preliminary experiments
showed thatio-dinationof virions withchloramine-T
efficiently
labeled virioncapsomeresand the
DNA-protein
core, whereasiodination withEnzymnobeads
la-beledonly
the virion capsomeres (data notshown). We were therefore interested to see
whateffect thesetwotypesofiodinations would haveonthe IEF distribution of virion and
capsid
proteins.
Comparing Fig.
3A and B, it can beseen that virion
VP,
species
Awasnot labeled withEnzymobeads.
Notethat in thisfigure only
the distribution of VP1
species
is shown.Fur-thermore,theratio of
species
Ctotheremaining
species
wasdrastically
decreased afterEnzymo-bead iodination, whereas the relative levels of
species
B, D, E, and F were notsubstantially
altered. When thesame
experimnents
werecon-ducted with
capsids, only
minor alterations in the ratios between the VP1species
were ob-served(Fig.
30 andD).Theseresultssuggested
ISOELECTRIC
fOCUSING*lo
A a
0
vo VP1 C
0
yp3 VP3~*VP3
FI..IEadSD-PE
f
hormie T idnaedvrin ncpsdprten.
ocsngwan0hhoriontldmenionwit acdit
inreaing romlef torigt.
SS-PGE as nteveticl dmenionAferth
irt-ienioafcuin,th gl
wreeuiibatdaneetrphrse
i teseon imnsoasdscriedyO'arrll
(5).(A)
utordioraphof olyoa cpsid aftr
a4-hxpoure;(B)autoadi
ogrphofolom casdSfe
4hepsr;()atrdorp
foym
iinfe
xoue(D)autradogaphofplyma
irinsaftr
a24heposre VOL. 37,1981on November 10, 2019 by guest
http://jvi.asm.org/
[image:4.496.103.389.363.599.2]84 BOLEN ET AL.
i
a
!r4
IL
*1
L
S
45
.4
SOI
ao 30
9-r as d%
GEL SLICE GL SLIU
FIG. 3. IEFof chloramine-T- andEnzymobead-iodinated virionsandcapsids. Virionsand capsids were
labeled in vitro with either the chloramine-TortheEnzymobead technique. Thesamples were then analyzed
byIEF and SDS-PAGE as described in the text. (A) Chloramine-T-iodinated virions; (B)
Enzymobead-iodinatedvirions;(C)chloramine-T-iodinatedcapsids; (D) Enzymobead-iodinated capsids.Only the positions
of virion and capsidVP,speciesareshown.
that virionVP, subspeciesAandpossiblysome
ofspeciesCwereinternal in the virionstructure.
Since suchdiscrepancieswerenotobservedwith
capsids,itwasthoughtlikelythatpolyoma
cap-sids harbornointernal proteins. Capsid VP2 and
VP:i,
likewise, exhibitedno changes in their de-gree of iodination. (The extent of capsid VP:3labeledwithEnzymobeadswascalculated from
single-dimension SDS-polyacrylamide gels, and
all structural proteinratios in virion and capsid
experiments were calculated in an analogous
fashion[data notshown].) Invirions, however,
we were able to calculate that approximately
20% of the virion VP3 and 2 to5% of the virion VP2 was internal. These calculations were in agreement with the results ofBrady et al. (7),
who observed small amounts of VP2 and VP:3
associated with thepolyoma DNA-protein
com-plex after virion dissociation with EGTA and DTT.
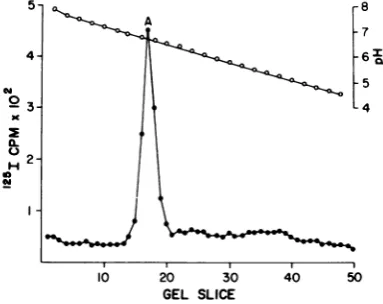
Isolationof virionDNA-proteincomplex;
virion and capsid capsomere subunits. Since thepriorexperiments suggested that
vir-ionVP, speciesA andperhapssome ofspecies
Cwere
internal,
we wereinterested in determin-ing which of the virionspecieswere associatedon November 10, 2019 by guest
http://jvi.asm.org/
[image:5.496.57.458.56.470.2]FUNCTIONS OF POLYOMA VP, SPECIES 85
with the DNA-protein complex. T
was obtained by dissociation of
(chloramine-T) virions with EGT. followedby velocity sedimentation ciated materialon5to20%sucrose
separatethecorecomplex from the
(7). The iodinatedcores werethen
SDS-PAGE and by IEF. The resu
the core are shown in Fig. 4. TI
species associated with the DNA-I plex was species A. Small amount:
VP3werefound when theSDS-pol
gels were autoradiographed for pi
riods, butwerenotobserved in the
to their low concentration. These gested that a single species of p
(species A)wasassociatedwith th(
Furthermore, these experimentsle'
ulation that perhaps the tertiary
5-A
4-3
2.
H
10 20 30
GEL SLICE FIG. 4. IEF of polyoma virion 48S complex. Chloramine-T-iodinated vir sociated in vitrobytheEGTA-DTTme et al. (7), and the 48S DNA-protein
isolatedafter velocitysedimentationi,
crosegradients. Thecomplexwasthe,
IEF and SDS-PAGEasdescribedint,
'his complex virion
VP,
species C prevented theexposureof'25I-labeled some of tyrosine residues of this polypeptide,
A and DTT therebyresulting inalowerdegree of
Enzymo-of the disso- bead iodination sincethis specieswasclearlynot
gradientsto associated with thecorecomplex.
acapsomeres Previous results from this laboratory have
analyzed by shown that, after EGTA-DTT dissociation of
dts of IEF of virions, three groups of capsomere subunits
he only VP, could be resolved by velocitysedimentation of
protein com- the dissociatedvirus(7).Thesethreegroupshad
s of VP2 and sedimentationvalues of18S, 12S,and5S.
Simi-lyacrylamide lartreatmentof capsids also yieldssimilar
cap-rolonged pe- somere sedimentationpatterns(J.B.B.,
unpub-IEFgel due lished data). The interest in these virion and
results sug- capsid capsomeresubunitsstemsfrom the fact
)olyoma VP1 that the 18S and 12S capsomeres contain
pri-avirus DNA. marily VP,. Electron microscopic analysis re-Itothespec- vealed distinct hexon arrangements (12S) and
structure of hexon aggregates (18S). No distinct structures
could be observed in the virion or capsid 5S
material, which contains primarily structural
[8 proteins VP2 and VP3 (7). The distribution of
-7 the various virion and capsid VP1species in the
-6
18S,
12S,
and 5Scapsomeres is shown in Table5 1.Note that inthese
experiments only
the virionandcapsid preparations labeled with125Iin vitro 4 thatyielded protein distributions closely
resem-bling in vivo 3H-aminoacid-labeled virions and capsidswere used (see below). Virion 12S cap-somerescontainedprincipally VP1 species B,
C,
andD, and capsid 12S material was composed
mostly of species A', B, and D. The same VP,
species also madeupthemajorityof virionand
capsid 18S capsomeres, but were enriched in
40 50
species
B. Virion5Scapsomereswerecomposed
principally
of VP,species
E andF,
whereasDNA -protein
capsid
5S capsomerescontainedmostly
species
ions
DNAproem
d D and F. These resultssuggestthatthe hexonions were dis-*
ethod
of Brady capsomeres of polyoma virions are made up ofcomplex was
VP1
andspecies
B, C,
and D and those ofcapsids
n 5 to 20% su- aremade up of VP1 species A', B,and D. These-nanalyzed by experiments lead us tospeculate thatpolyoma
hetext. capsid speciesA'
(pl,
pH 6.6) playsaroleequiv-TABLE 1. DistributionofVP,species incapsomeresubunits"
Virion-s()" Capsids(si.`
Sample
Total A B C D E F Total A' B D E F
18S 21.3 48.2 30.1 20.0 0.8 0.9 20.5 35.2 56.3 18.5
12S 66.3 30.4 40.0 27.6 0.5 1.5 65.3 48.3 26.1 22.3 3.3
5S 12.4 4.5 1.2 0.6 42.3 51.4 14.2 9.3 41.3 49.4
"Capsomeresubunitswereisolated fromsucrosegradientsafterEGTA-DTTdissociation of chloramine-T-iodinatedvirions andcapsids.
"Theareaof thepeakfor eachVP, speciesisexpressedasthepercentageof the totalareaoccupiedbythe speciespresent in thesample. Each value is the average ofanumber ofseparate determinations ofpooled samples.
'The amount ofradioactivityin each capsomere speciesis expressed asthe percentage of the total VP, radioactivityrecovered fromthegradients.
VOL. 37,1981
on November 10, 2019 by guest
http://jvi.asm.org/
[image:6.496.47.239.282.434.2]86 BOLEN ET AL.
alent to that of virion
species
C(pl, pH
6.3)sincetheirrelative distributionsinthecapsomere
sub-units were similar. The results may also shed
somelightonthenatureof thepolyomapenton.
Since distinctpenton capsomeres couldnotbe
observed in the 18S and 12S
subunits,
it hasbeensuggested that the
building
blocks of thepentonsreside in the5S
region
(7). Thus,it ispossible that virion
VP,
species E and F andcapsid
VP,
speciesDandFareinvolvedinthepenton structure.
Post-translational modifications of the
virion and
capsid
VP1species.
We nextsoughtto determine the nature ofsomeofthe
modifications of thevirion andcapsidVP, spe-ciesby labeling polyoma-infected cellswith
dif-ferentradioactiveprecursors. The infectedcells
were labeled with
:2pO4,
['4C]acetate,
orH-amino acids, and the virions and
capsids
werepurifiedthereafter. Equalamountsof32P-or
'4C-labeled virions and
capsids
and 3H-labeledviri-ons andcapsids were
mixed,
and the mixtureswereexamined by IEF,
SDS-PAGE,
andIEF-SDS-PAGE. The results of IEF of 3H-amino
acid-labeled virions and
capsids
are shown inFig.5. Itisimportantto notethat the number of
polypeptide speciesobserved and theirpI'swere
the same as the results obtained when
chlora-mine-T-'251-labeled
virionsandcapsids
wereex-aminedinthismanner
(see
Fig. 1).
These resultssuggested that in vitro iodination of
polyoma
virionsandcapsids didnot
appreciably
alter thepl's of the virus
proteins. However,
these resultsalso indicated thattherelative amounts ofthe
individualspecies couldbe altered to some
ex-tentduring theinvitro
labeling
procedure. Thus,to quantitate the protein
distribution,
only invivo 3H-amino acid-labeled virion and capsid
proteinswereused.
The results of the mixing experiments are
shown inTable 2 andagain pointout the
dra-matic differences between polyoma virion and
capsid proteins.Fourof the
polyoma
virionVP,species (A, D,E, and F) and two of the capsid
VP1
species (D and F) were foundto bephos-phorylated. OftheVP, speciescommonto both
virions andcapsids which were also
phosphoryl-ated,virionspeciesD and Fexhibitedtwice the
level of
:p2p
incorporationasdid capsid species D and F. Two of the virionVP,
species (C andD) and three of thecapsid VP, species
(A', D,andF)werefoundtobe
acetylated.
The minorstruc-tural proteinsof both virions and capsids (VP2
and
VP:j)
werefound to bephosphorylated butnotacetylated. Ourattempts to label virions and
capsids with
S-[methyl-:H]adenosyl-L-methio-nine to determine whetherany of the virus
pro-teinswere
methylated
have been unsuccessful.Although asmallamountof label has beenfound
associated with purified virions and slightly
more has been found associated with capsids,
thedegree ofincorporationwassufficientlylow
thatanalysis by SDS-PAGEorIEFyielded
in-conclusive results.
Immune
precipitation
and IEF ofGuHCIcolumn-purified VP1. In a previous report
from this laboratory, polyoma virion proteins
VPI,
VP2,andVP:3
and histones were separatedby using GuHCl gel
filtration
(4). Weutilized
this technique toisolate
'25I-labeled
virionVP1
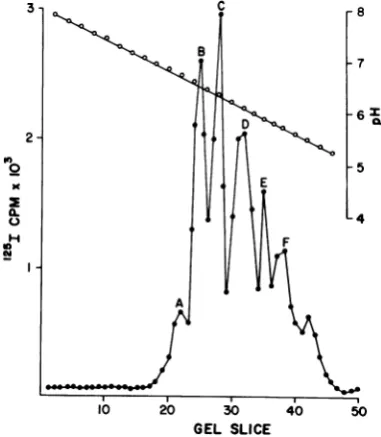
todetermine whether theGuHCl
column-puri-fiedVP, showedanIEFpattern similartothat
foundpreviouslywithpolyoma virions. The
re-sults shown inFig. 6clearly demonstrate that,
indeed,allofthe
VP,
species foundduring IEFofvirions were found after IEF of the
GuHCl
column-purified
virionVP,.
Samples ofthe samepooled VP, preparation were then used in
im-mune precipitation experiments with rabbit
anti-polyoma hemagglutinin and anti-polyoma
receptor. In anearlierreport (3), it was
demon-stratedthat these two antibody activities could
beseparated by polyomacapsid affinity column
chromatography from antiserum prepared
againstproteolytic cleavageproducts of the
ma-jor structural protein
VP,
inSDS-polyacryl-amide gels. In immune precipitation experi-ments,itwasfoundthat, after EGTA-DTT dis-sociation ofpolyoma virions (conditions known
to lead to the breakdown of
VP,),
theanti-polyoma
hemagglutinin specifically precipitated
a16K-daltonVP1
cleavage product,
whereastheanti-polyoma receptor
specifically
precipitateda14K-dalton
VP,
breakdownproduct. Whenthesame virion
preparation
wasdissociated underconditions which minimized VP1
degradation,
both
anti-polyoma
hemagglutinin and anti-re-ceptorantibodiesimmuneprecipitated 44K-dal-ton VP1. Thegoal
of the experiments withGuHCl
column-purified
VP1wasthus todeter-mine whether any one or a group of the
VP,
species would berecognized by oneor both of
these
antibody preparations.
The results of these immuneprecipitationexperiments are shown inFig. 7.Twoof the VP1specieswere
specifically
immuneprecipitatedwithanti-polyoma
hemag-glutinin (Fig. 7A). These two species
corre-sponded to
VP1-D
andVP,-F.
Onlyone oftheVP1
species(species E)wasimmuneprecipitatedwithanti-polyoma receptor immunoglobulin G
(Fig. 7B). These results suggest that the
VP1
speciesassociated with thehemagglutinating
ac-tivityofpolyomavirionsconsist of
VP1
speciesDand F. It isinterestingthatthese two species
are also foundon
polyoma
capsids which have been shown to compete for the samecellular
on November 10, 2019 by guest
http://jvi.asm.org/
FUNCTIONS OF POLYOMA VP, SPECIES 87
0 lo.
A, 8. o8
0
O 6-o6
,C E
a
.X
4-GEL SLICE
8
I-6
XC
5 460
a -8
-6:
-5 -410 20 30 40 50 60
GEL SLICE
FIG. 5. IEFof3H-aminoacid-labeled virions andcapsids.Polyoma virionsandcapsidswerepurified from
thelysates of infectedcells grown in the presenceof3H-aminoacidsduringtheperiodofvirusinfection.
3H-aminoacid-labeled virions(A)andcapsids(B)werethenanalyzed byIEFandSDS-PAGEasdescribed in thetext.
receptors asvirions on thesurface of GPE (2). Theseresults furthersuggestthatVP, speciesE
isresponsible for the virion-associatedreceptor
activityofpolyomavirions which isrequiredfor
specific adsorption and infection of primary
MKC since the anti-polyoma receptor anti-bodiesspecifically inhibitedpolyomavirion
ad-sorptionto MKC butnot toGPE (3).
Further-more, VP,-E is found only on polyoma virions and is absentin polyoma capsidswhich fail to
compete with virions for receptor sites on the surface ofMKC (2).
DISCUSSION
The central reason for comparing the IEF
patternsofpolyomavirion and capsidproteins
stems from previous studies conducted in our
laboratory which demonstrated distinct
biologi-cal,biophysical,andbiochemicaldifferences
be-tween these two populations of virusparticles
(2, 3, 12,20).MackayandConsigli (12) reported
thatthe fate ofpolyomacapsidswithin cellswas
distinct from the small
population
ofpolyomavirionswhichwereresponsibleforcellular infec-VOL. 37,1981
on November 10, 2019 by guest
http://jvi.asm.org/
[image:8.496.131.379.61.496.2]88 BOLEN ET AL.
TABLE 2. IEFofpolyomavirion andcapsid structuralproteins
Protein Species pI totalo P/'H" '"'C/'H' Virions
VP, A 6.75 8.6 0.2
B 6.50 18.0
C 6.30 37.8 0.05
D 6.10 13.2 0.40 0.25
E 5.90 6.6 0.60
F 5.75 5.8 0.80
VP2 5.50 4.8 1.00
VP:, 7.00 5.2 1.10
Capsids
VP, A 6.60 34.6 0.20
B 6.50 18.5
D 6.10 20.9 0.20 0.40 F 5.75 16.0 0.40 0.30
VP2 5.50 5.4 0.80
VP:, 6.50 4.6 1.00
"Theareaof thepeak foreach of theproteinsand
proteinspecies isexpressed asthepercentage of the
totalareaoccupied by the protein or protein species present inthesample.Eachvalue isthe average of a number of separate determinations of pooled
'H-aminoacid-labeledsamples.
'Ratio of "Pi-labeled proteins to :'H-amino
acid-labeled proteins.
'Ratio of["4C]acetate-labeledproteinsto 'H-amino acid-labeledproteins.
c
p
87
613
2-2 5
0
-U~~~~~~~~~~
10 20 30 40 50
[image:9.496.61.252.77.283.2]GEL SLICE
FIG. 6. IEF of GuHCI-chromatographed virion VP,. Chloramine-T-iodinated virion proteins were
chromatographedon aGuHClcolumn asdescribed
inthetext.TheVP, fractions from the column effluent
werepooled, dialyzed againstTrisbuffer (pH7.4)to
removeexcessguanidine, and analyzed by IEF and
SDS-PAGEasdescribed in thetext.
5
4
3
3
5-
2-0
a.
C. 5.-H
4.-
3.-2
--8
-6
1L
5
-4
-8
-6X 5 4
10 20 30 40 50
GEL SLICE
FIG. 7. IEF of GuHCl column-purified VP,
im-muneprecipitation. Samples ofthe Chloramine-T-iodinated VP, isolatedasdescribed in thelegendto Fig.6wereused in immuneprecipitationexperiments withantihemagglutinin andantireceptorantibodies
as described in the text. The immuneprecipitates
wereanalyzed byIEF. Thefigureshows thefocusing patternofGuHCIVP,plusantihemagglutinin (A)or antireceptor(B). Forclarity,thebackgroundcounts obtained by incubatingnormal rabbit
immunoglob-ulin Gwith theGuHCIVP,have beensubtracted.
tion. After virion adsorption to cells, a small percentage ofthe original inoculum was
trans-ported to the nucleus, where the virions were
uncoated (12, 20), and early transcriptional
ac-tivitieswereinitiated (20).Themajority of the
originalvirionswerefound in cellularlysosomes,
wheretheyweredegraded. Aftercapsid
absorp-tiontocells, nosignificant nuclear transport of
these"empty" virusparticleswas detected.
In-stead, almost the entire capsid population was
sequestered into cellular lysosomes and
de-graded(12). Subsequently, itwasreported that
thereasonfor this differential fate of virions and
capsidswithin cells resulted fromthemannerby
which virions and capsids adsorbed to the cell surface (2). Although capsids efficiently
com-petedwith virions for receptorsitesonthe
sur-face of GPE, capsids could not compete with virions for receptor siteson MKC, which were
requiredforsuccessful infection. This study also
demonstrated that thereason only 7 to 10% of
theinput virions ever entered thenucleus was
on November 10, 2019 by guest
http://jvi.asm.org/
[image:9.496.61.252.384.603.2]FUNCTIONS OF POLYOMA VP, SPECIES 89
because the number of specificsurface receptors to which virions, but not capsids, could bind was
limited.Recently, the maximum number of
spe-cific virion receptor sites on the surface of quies-cent primary MKC has been estimated to be
approximately 10,000 per cell (J.B.B.,
unpub-lished data). In the same report, it was reported that anadsorption mutant of polyoma (Py 235),
originallydescribed by Basilico and DiMayorca
(1), which lacked the ability to adsorb to and
agglutinate GPE, nevertheless possessed an
equalabilitytospecificallyadsorb to and infect
MKC at 32°C as did wild-type virions. These
results demonstrated that specific adsorption to
MKC and infection of these mouse cells was independent of the ability ofpolyoma virions to
adsorb to and agglutinate GPE. In a recent
report (3), we were able to demonstrate the
separation of neutralizing antibodies from
he-magglutination-inhibitingantibodies in antisera
directedagainst cleavage productsof virionVP1
which migrated in the histone region of
SDS-polyacrylamide gels. Itwasfound that the
anti-genic determinantsresponsible for
hemaggluti-nation-inhibiting activity of the serum were
present on a 16K-dalton polypeptide, whereas those antigenic determinents responsible for
neutralizationactivity of theserum were present
on a 14K-dalton polypeptide. These two
poly-peptideswerebothderivedfrom 44K-dalton
vi-rion
VP,.
Theseresults strongly suggested thatthemajority of the diverse biological functions
observed were dependent on the major
struc-turalprotein VP1.
In this report we have shown distinct differ-encesbetween thesubpopulations of the
struc-tural proteins ofpolyoma virions and capsids.
The results presented here demonstrate that
polyoma virionspossess sixspeciesofthe major
structural protein
VP,
in addition to the twominorstructuralproteins VP2and
VP:). Polyoma
capsids were found to contain only four VP,
species in addition to VP2 and VP:3. Multiple
species of
polyoma
virionVP, differing
in theirpl'shave beenreported previously (11, 17). How-ever, the number ofspeciesand thefunctionsof these
multiple species
were not clear. It is ofconsiderable interest that O'Farrell and
Good-man(16)
reported
thatSV40virionsalso possess six distinct VP1 species. Such results suggest thatthispattern ofsixVP,
species(eventhough the pl's of these species vary considerably be-tween polyoma andSV40) may be commontoallpapovaviruses.Our
experiments
haveallowedustotentatively assignfunctionstothe various
speciesofVP,and have aidedin the
comprehen-sion of howVP1possesses thepotentialfor such
biological diversity.
The major structural protein (VP,) of
poly-oma virions consists of six species which have pI's between pH 6.75 and 5.75. On
two-dimen-sional analysis these species all had molecular
weights in the 42K to 45K range. The molecular-weight diversity observed was not due to con-tamination of our standard wild-type virions with defective virus or deletion mutants, as freshly plaque-purified virus samples yielded
identical electrophoretic patterns. The
differ-ences in pI's of the VP1 species conceivably
result from the different degrees and types of modifications observed (Table 2). Four of the
VP,
species were differentially phosphorylated (A, D, E, and F) and two of the species wereacetylated (C and D). In our analysis to date,
only speciesBappearstobeunmodified. Similar
experimentswithcapsids revealed only four
VP,
species withpI's betweenpH 6.60 and 5.75. Two
of the capsid
VP,
specieswere phosphorylated(D and F). It is interesting that the degree of
phosphorylation as determined by the ratio of
32P/3H
ofthesetwocapsid VP1 specieswasonlyhalfthatobservedwith virion
species
D andF,
although the significance of this remains
ob-scure.
Similarly,
theimportance ofthree ofthecapsid
VP, species
A',C, andDbeingacetylated
isalsounclear. Aswithvirions, capsidVP1
spe-ciesB does notappeartobemodified via
ace-tylationor
phosphorylation.
Analysis
ofthemi-nor virion and
capsid
structuralproteins VP2
and VP3 also revealed that differences exist.
VirionVP3had apl of pH 7.00,whereas
capsid
VP3 had anapparent pl of pH 6.50.Again,
thesignificance of this observationisnotunderstood
atthepresent time.
Capsid
and virionVP2
had a commonpI ofpH
5.50.Furtheranalysis
ofall ofthe virion andcapsid
VP, species, VP2,
andVP3 using other
analytical techniques,
suchastryptic peptide
mapping,
anda widevariety
oftemperature-sensitive
and deletion mutants iscurrently
under way in thislaboratory.
It isanticipated
thatsuchanapproach
will behelpful
in
answering
manyofthequestions
raised inthisinvestigation.
The variety of
presumably
host-contributedpost-translational
modifications of the virusstructural
proteins,
particularly VP1,
suggeststhat suchmodifications couldallowanumber of different functions to be expressed by a
single
protein.Atthe present
time,
this appearstobethecase withpolyomavirus andrepresents
an-otherexampleof the virus'abilitytoefficiently
utilize and expand its limited genetic coding
capacity.Our results suggest thatpolyomavirion
VP, may serve atleast four distinct functional roles. The first is a structural function. The
experiments in which the various VP1
species
weredifferentiallyiodinated with
Enzymobeads
orchloramine-T
(Fig.
3)demonstrated thatVP,
VOL. 37,1981on November 10, 2019 by guest
http://jvi.asm.org/
90 BOLEN ET AL.
species A was an internal protein whereas the
remainingVP, specieswere available for
exter-nal labeling. Analysis of the EGTA-DTT-de-rived capsomere subunits (Table 1)
demon-strated that virionVP,species B, C, andDwere
presentincapsomerepreparations enriched with
hexon subunits, whereas species A, E, and F
wereabsent. These resultssuggestthatpolyoma virion hexonsarecomposed ofVP,species B, C,
and D.They alsosuggestthatVP,species E and F may be involved in the penton capsomere
subunitsof the virion. Thus, potentiallyfive of
the six VP1species performastructural function
for the virus. Similar analysis of capsids
sug-gested thatVP, speciesA' incapsids likely
per-forms thesamefunctionasvirion VP1species C
(Tables 1and2). In capsids, therefore, the
hex-onsareapparently composedofVP, speciesA',
B, and D. Since capsids lackaVP,species which
physically corresponds to virion species E, it
appears that capsid species D, along with F, mightcompensatestructurally for the lack of E. Minorproteins VP2 and VP3werefoundboth in
capsomerefractionsbeingaccessible forsurface iodination withEnzymobeads andinassociation with the DNA-protein core (7; J.B.B.,
unpub-lisheddata).At thistime, the roles ofVP2 and VP;3asstructural entitiesorregulatory proteins, orboth, unfortunately remain unresolved.
The second role thatpolyoma VP, possesses
is exemplified by virion VP, species A. This species,asnotedabove,couldnotbelabeled in vitro by the Enzymobead technique (Fig. 3B) andwasfoundtobe theonlyVP,species firmly associated with the virion DNA-protein core
(Fig. 4). Furthermore, VP, species A was not
found in capsids (Fig. 1A, 3C and D, and 5; Tables 1 and 2) or in significant amounts in
capsomere subunits (Table 1). The function whichthis DNA core-associatedVP,species
ful-fillsisnotclear at this time. ThatVP,species A, themostbasic (pl,pH 6.75) oftheVP,species, is apparently exclusively associated with the DNA-protein complex of the virus implies that its role may not be structural in nature. One function VP, species A may serveissuggested
by the recent experiments by Brady et al. (5; personal communication). They demonstrated that 110S SV40 nucleoprotein cores derived
from intact240S SV40 virions by EGTA-DTT dissociationcontained VP,and asmallamount ofVP2inadditiontotheproteinsnormallyfound
associated withtheSV40 55Sminichromosome (i.e., VP.,and histones). In additiontokeeping the 110Scorecompact,theassociatedVP1-VP2
significantly modified the transcriptional activ-ity of the core in the presence of Escherichia
coliRNApolymerase. The presenceofthelate
virusproteinsresulted intranscription of 16to
18SRNAwhichhybridizedtothe"early" SV40
DNA region. In the absence of VP1-VP2, the
RNA transcription product was only 4 to 6S.
These results lead to the speculation that the same events may take placewith polyoma
nu-cleoproteincomplexes. Experimentstotestthis
hypothesisarecurrentlybeingconducted in our
laboratory. Nevertheless, the experiments of
Brady et al. clearly point out the probability
that VP1 andperhaps VP2,
normally
consideredonly structural entities, alsopossess regulatory
functions.
The remaining two functions which can be
attributed topolyoma
VP,
deal with the virus'ability to adsorb to mammalian cells such as
GPE andprimary MKCor mouse
embryo
cells.The datapresentedinthisreportindicate that
threeofthe virion VP1
species (D, E,
andF)
areinvolvedinsuchactivities.
VP,
speciesDand Fappear to be common to both virions and cap-sids, although their distribution among
capso-mere subunits, degree ofmodification, and
ab-solute concentrationdifferto some extent
(Ta-bles 1 and 2). Our data suggest, however, that
whereasthecapsidD and Fspeciesappeartobe
slightly different from theirvirion counterparts,
theymayfulfillsimilarfunctions. These results
alsopoint out the possibility thatsome ofthe
modifications observedmay notnecessarily
dic-tatefunctionalchangesotherthan
allowing
con-formational diversity of the
VP,
moleculesre-quired for assembly of virusparticles. The
ad-ditional biological functions attributedtosome
of thesespecies could thus be dependentonthe
presence of other unidentifiedmodifications or in many instancesthey couldbeindependent of
suchalterations. The functionalidentity of
vir-ion species D and F wassuggested by the
im-mune precipitation experiments (Fig. 7).
Anti-bodieswhoseonlyknownfunctionwastoinhibit virion orcapsid absorption to and agglutination
of GPE could specifically recognize only
VP1
species D and F. Thus, it appears that virion
VP1
speciesDandF areinvolved withthe virus'abilitytoadsorbtoandagglutinateGPE.These
results further suggest thatcapsid VP, species Dand Fprobably
fulfill
the samefunction,since virions andcapsids have an equal ability to causehemagglutinationwhich canbe equally inhibited
bythehemagglutination-inhibitingantisera (3).
Virion VP, species E appears to represent the virion protein which is required for successful virus adsorption to specific cellular receptors, which results in infection of MKC and mouse
embryo cells. Thus,
VP,
species E apparentlyfulfillsthe role of thepolyomavirion-associated
receptor. Two lines of evidence support this
on November 10, 2019 by guest
http://jvi.asm.org/
FUNCTIONS OF POLYOMA VP, SPECIES 91
contention. Primarily, VP1 species E is found
onlyonvirions,never on capsids.Since, as
dis-cussed above,capsids donot compete with
viri-ons forspecific cellular receptors (2), it seems
logical that capsids would lack the correspond-ing protein present on virions. Similarly, since
capsids andvirions share the same
cell-associ-ated receptor on GPE (2), one would expect these two typesofvirusparticlestoshare
com-mon hemagglutinins. As discussed above, this
appears tobethecase.Second, antibodieswhose
onlyknownactivitywas toinhibit virion
adsorp-tionto mousecells,but not toGPE,specifically
recognizeddistinct antigenicdeterminantsonly
on
VP,
species Einimmuneprecipitationexper-iments (Fig.7B).
An additional point of interest concerns the
precursor-product relationshipbetween the
VP,
species. Thepulse-chase experiments performed
byO'Farrell andGoodman (16) suggestedthat
themostacidic of theSV40
VP,
species actedasaprecursor to twoadditional
VP,
species whichfocused
immediately adjacent
totheprecursor.If the same relationship is also present with
polyoma, then species F would represent the
precursorforspeciesDandE.Sucha
relation-ship would indicate that the
VP1
speciesin-volved in cellular adsorption activities are
de-rivedfromacommonprecursor which is distinct
from other
VP1
species, possibly throughde-phosphorylationof theprecursor(Table2).
Thisstudyhasallowedus toassign functions
tothe VP1species ofpolyoma virus. In the future
we anticipate being able to better understand
the molecular interactions which result in the
modified forms of
VP1
and to determine whatother functions
VP,,
VP2,and VP3mightexpressduring thecourseof
polyoma
infection.ACKNOWLEDGMENIS
Thisinvestigationwassupportedby Public Health Service grant CA-07139fromtheNational Cancer Institute. R.A.C.is aSenior Scientist with the Mid-America Cancer Center Pro-gram.
We expressourappreciationtoDiane Potts and Viola Hill forexcellent technical assistance andtoSueStefanskiforhelp inpreparationof thismanuscript.We also thank M. S. Center forhelpful discussions.
LITERATURE CITED
1. Basilico, C.,andG.DiMayorca. 1974.Mutant of poly-omavirus withimpaired adsorption toBHK cells. J. Virol. 13:931-934.
2. Bolen, J. B., and R. A. Consigli. 1979. Differential adsorption ofpolyoma virions andcapsidsto mouse kidney cells andguineapig erythrocytes.J. Virol.32: 679-683.
3. Bolen, J. B., and R. A.Consigli.1980. Separationof neutralizingandhemagglutination-inhibiting antibody activitiesandspecificityofantiseratosodiumdodecyl sulfate-derived polypeptides ofpolyomavirions. J. Vi-rol. 34:119-129.
4. Brady, J.N., andR.A.Consigli.1978.Chromatographic separation ofthepolyoma virus proteins and renatur-ation ofthe isolatedVP,majorcapsidprotein. J. Virol. 27:436-442.
5. Brady, J.N., C. Lavialle, andN. P.Salzman. 1980. Efficienttranscription of a compact nucleoprotein com-plex isolated from purified simian virus40virions. J. Virol. 35:371-381.
6. Brady, J. N.,V. D.Winston, and R. A.Consigli.1977. Dissociation ofpolyomavirusbythe chelation of cal-cium ions found associated with purified virions. J. Virol.23:717-724.
7. Brady, J.N., V. D. Winston, and R. A.Consigli.1978. Characterization ofaDNA-protein complexand cap-someresubunits derivedfrompolyoma virusby treat-mentwithethyleneglycol-bis-N-N'-tetraacetic acid and dithiothreitol. J. Virol. 27:193-204.
8. Brunck,C. F., and V. Leick.1969.Rapidequilibrium isopycnicCsCl gradients. Biochim. Biophys. Acta 179: 136-144.
9. Deininger, P., A.Esty, P. LaPorte, and T. Friedmann. 1979.Nucleotide sequence andgenetic organization of thepolyoma lateregion:features common tothe poly-omaearly region and SV40. Cell 18:771-779. 10. Fried, M., and B. Griffin. 1977. Organization of the
genomes ofpolyoma virus and SV40. Adv. Cancer Res. 24:67-114.
11. Hewick,R.M., M.D.Waterfield,L K.Miller,and M. Fried. 1977.Correlationbetweengenetic loci and struc-tural differences in thecapsid proteinsofpolyomavirus plaquemorphologymutants.Cell11:331-338. 12. Mackay, R.L.,and R. A.Consigli.1976.Earlyevents
in polyoma virus infection: attachment, penetration, and nuclearentry.J.Virol.19:620-636.
13. McMillen, J., M. S.Center,andR.A.Consigli.1976. Origin ofpolyoma virus-associated endonuclease. J. Vi-rol. 17:127-131.
14. McMillen,J., and R. A.Consigli. 1974.In vitro radio-isotopic labeling of proteins associated with purified polyomavirions. J. Virol.14:1627-1629.
15. O'Farrell,P.H. 1975.Highresolutiontwo-dimensional electrophoresis of proteins. J. Biol. Chem. 250:4007-4021.
16.O'Farrell, P. Z., and H. M. Goodman. 1976. Resolution ofsimian virus 40 proteins in whole cell extracts by two-dimensionalelectrophoresis: heterogeneity of the major capsid protein. Cell9:289-298.
17.Ponder, B. A. J., A. K. Robbins, and L. V. Crawford. 1977.Phosphorylation of polyoma and SV40 virus pro-teins.J. Gen. Virol. 37:75-83.
18. Smith, G. L, and R. A. Consigli. 1972. Transient inhi-bition of polyoma virus synthesis by Sendai virus (parainfluenzaI). I.Demonstration and nature of the inhibition by inactivated virus. J. Virol. 10:1091-1097. 19. Vinograd, J., andJ.E.Hearst. 1962. Equilibrium
sed-imentation ofmacromolecules and viruses in a density gradient. Prog. Chem. Org. Nat. Prod. 20:372-422. 20. Winston, V.D., J. B.Bolen, and R. A.Consigli.1980.
Isolation and characterization of polyoma uncoating intermediatesfromthenuclei of infected mousecells. J. Virol.33:1173-1181.
VOL. 37,1981