Copyright© 1976 American Society for Microbiology Printed inU-SA.
Properties of the Scrapie Agent-Endomembrane Complex
from
Hamster Brain
J. S. SEMANCIK,* R. F. MARSH, J. L. M. C. GEELEN, AND R. P. HANSON
Departments ofPlant Pathology and CellInteraction Group, University of California, Riverside, California 92502,*and Department of Veterinary Science, University of Wisconsin, Madison, Wisconsin 53706
Received for publication 2 September 1975
Subcellular fractionation of scrapie-infected hamster brain indicatedthe
asso-ciation of the scrapie agent with a component of the endomembrane system.
Characterization by equilibrium density gradient centrifugation, electron
mi-croscopy, and markerenzymessuggestedaprimary association with rough and
smooth endoplasmic reticulum and a possible incorporation into the plasma
membrane. DNA polymerase activity demonstrated a direct correlation with
regions of scrapie activity from thegradientfractions. Ascrapie-related product
wasdetected after [3H]TMP incorporationand analysison 2.2% polyacrylamide
gels. Analysis of nucleic acid species extracted from subcellular fractions
re-sulted in a greater quantity fromhealthybrain; however, noqualitative
distinc-tions weredetected.
Thetransmissible agent of scrapie disease of
sheep has unusualproperties ofthermal
stabil-ity (8) and a small radiation target size of 2 x 105 daltons (2). Similar characteristics have also been reportedfor"viroid"agentsproducing infection ofplants (5, 18-20). No nucleic acid
species analogousto the low-molecular-weight
pathogenic RNA, characterized as the causal
agentofviroid diseases (21), has beenisolated
from scrapie-infected tissue or any other
ani-mal system (16). Nevertheless, high titers of
biologicallyactivescrapieagent canbe demon-strated ie membranepreparations (17). The
as-sociation of the citrus exocortis viroid with a
componentof theendomembranesystem (J. S.
Semancik, D. Tsuruda, L. Zaner, J. L. M. C. Geelen,and L.G.Weathers,Virology,inpress)
further extended the comparative features of these diseases. The characterization of the
scra-pie agent-membrane complex from hamster
brain, employing procedures comparable to
those utilizedinviroid characterization, is the subject of this report.
MATERIALS AND METHODS
Scrapie agents. The scrapie agent usedin these
studies originated from the Chandler strain of
mouse-adapted scrapieasdescribedpreviously (11).
Fractionationexperiments wereperformedonpools
(5 g each) of fresh brain tissue representing the
agent's4thand 5th passagesinoutbred(LVG:LAK) Syrian hamsters with intermittent passages in
inbred hamsters (LHC/LAK), both obtained from the Lakeview HamsterColony, Newfield,N.J.
Ad-ditional pools, containing 50 g each offresh brain
from the 6th scrapie passage or from normal
ani-mals, wereused for fractionation followed by nucleic acid extraction.
Extraction and subcellular fractionation proce-dure. Hamsters, 4monthsold, either uninoculated
orshowing early signs of disease, were killed with chloroform, and their brains were rapidly removed and placed on wet ice. Healthy or scrapie-infected brains (2.5 g) were quartered and immediately ho-mogenized on ice (Polytron blender, 15 s, no. 4 set-ting)in10to 15 ml of a medium composed of 0.4M
sucrose,0.01 Mtricine (pH 7.5), 0.002MMgCl2, 0.02
MKCl,0.1%bovine serumalbumin, 0.001 M dithio-threitol, and 0.04Mmercaptoethanol. The extracts were pooledand centrifuged at 250 xg for 10min, and thepellet was discarded.The250-gsupernatant
wassequentially centrifuged at 1,000 xg(fraction 1 pellet) and 13,000 xg (fraction2pellet)inaSorvall RC-2B for10mineach. The 13,000 xgsupernatant
wasthen centrifuged at 80,000xg for 30min (frac-tion3pellet)inaBeckman SW27 rotor. Finally, the supernatant wascentrifuged at 100,000 x gfor2h (fraction4pellet) in a Beckman 35 rotor. The high-speed supernatant constituted fraction5.Allpellets were suspended by three strokes in a Vitro Teflon homogenizer in a medium containing 15% (wt/wt) sucrose,0.01Mtricine(pH 7.5),0.002 MMgCl2,0.02 MKCl,and 0.001 M dithiothreitol.
Density gradient centrifugation. Sucrose
solu-tions were prepared in the resuspension buffer. Samples (4to6ml)werelayeredover alinear(25to
40%, wt/wt) sucrosegradientover 4 mlof50% (wt/ wt) and2mlof60% (wt/wt)sucroseand then
centri-fuged for21 hat 25,000 rpm ina Beckman SW27
rotor at 4 C.Fractions (1to 2ml)werecollectedby needle-puncturing the bottom of the tube, and the density of theconsecutive sampleswasdetermined withanAbberefractometer. Aliquotsof these
sam-pleswereemployed for the enzyme assay,analytical analysis, and scrapiebioassay.
693
on November 10, 2019 by guest
http://jvi.asm.org/
Analytical and enzymaticdetermination. RNA, DNA, and protein were quantitated by theorcinol, diphenylamine, and Folin tests, respectively (22). The plasma membrane (PM) marker enzyme (ATP-ase) and endoplasmic reticulum (ER) marker
en-zyme (reduced nicotinamide adenine dinucleotide [NADH] cytochrome c reductase) were determined by the method of Hodges and Leonard (9). DNA and RNA polymerase activities were monitored by [3H]TMP and [3H]UMP (J. L. M. C. Geelen, L. G. Weathers, and J. S. Semancik, Virology, inpress) incorporation.
Nucleic acid extraction and gel electrophoresis. Nucleic acid preparations were extracted and parti-tioned as reported previously (17) and analyzedon 5
and 2.2% gels with 0.5% agarose (3). Gels were scanned at 260 nm in a Beckman Acta II spectropho-tometer.
Bioassay of scrapie infectivity. Coded fractions were tested for scrapie infectivity by intracerebral inoculation (0.05 ml) of weanling outbred hamsters. Four animals were inoculated with each dilution, beginningat10-5,and end pointswerecalculatedat
the end of 15 weeksbythe method of Spearman and Karber (7).
Electron microscopy.Gradientsamples of subcel-lular fractions were pelleted and immediately placed in 2% glutaraldehyde in 0.01 M potassium phosphate buffer, pH 7.2. After a 2-h fixation, the pelletswererinsed well with buffer andpostfixedin 1%buffered osmium tetroxide for 90 min.After sev-eralbuffer andwaterrinses, specimens were dehy-drated through anacetone series andembedded in
Epon (14). Thin sections were stained with uranyl
acetateand leadcitrate.Specimenswereviewed and photographed with a Hitachi HU-11 microscope.
RESULTS
Subcellular fractionation of hamster brain.
A differential centrifugation sequence similar
tothat employed for detecting theassociationof
thecitrus exocortis viroidwitha componentof
theendomembranesystem (Semanciketal.,in press) was utilized to process polytron
homoge-nized brain tissue. After the250 x g (10 min)
crude pellet was discarded, the series of
frac-tions wastaken, as presented in Table 1, and
assayed for scrapie activity (SA). The SA
de-TABLE 1. Associationof SA with subcellular fraction from infected hamster brain.
Total recoverableSAa Pellets
Expt1 Expt2 Expt3
(1) 1,000 xg, 10 min 10925 10W75 109 (2) 13,000 xg, 10min 109 108.75 109 (3) 80,000 xg,30 min
109i25
108T75
109 (4) 100,000 xg,2h 108'5 108.25 108.75(5)Supernatant 10775 108-25 108
a Mean lethaldose per total volume of the
resus-pended pellet.
tected in the three fractions collected between 250 and80,000 xg, containingthe bulk of the
endomembrane system, was usually 1 log
greaterthan the final high-speed pelletor su-pernatant. Whensubjectedtoequilibrium
cen-trifugation ina25 to60% (wt/wt) sucrose
den-sity gradient, characteristic light-scattering
patterns were observed in the three fractions,
indicating the presence ofaPM-likezone (37%)
in the 1,000 x g pellet, a heavy mitochondria
zone (39%) in the 13,000 xgpellet, anda more
diffuse ER-like zone at lower densities (30 to
34%) in the 80,000 x g pellet. The latter two
fractions also displayedmorefaintzonesinthe
PM region. Since the levels of SA in the three
fractions were comparable, yet the 13,000 x g
pelletwasgreatly enriched inmitochondria, we
chose to focus on the characterization of the scrapie-associated component in the 1,000 and
80,000 xgpellets, thus using themostdistinct
subcellular membrane fractions not contami-nated with excessive levels of mitochondria.
Characterization of membrane components ofsubcellular fractions. Subcellular fractions
of the 250 to1,000 x g and 13,000 to80,000 x g
pellet, designated H-1 and H-3, respectively, were prepared from Polytron-homogenized,
healthy hamster brains.Afterequilibrium
sed-imentation in a 25 to 60% (wt/wt) sucrose
gra-dient, sampleswerecollected andanalyzed for
density (Fig. 1A) protein concentration (Fig.
1B), NADH cytochrome c reductase activity
(Fig. 1C) as a mitochondria and ER marker,
ATPase activity (Fig. 1D) as a PMmarker, and
[3H]TMP incorporation into an acid-insoluble product as an indication of DNA polymerase
activity (Fig.
1E).
Since these experimentswere designed to localize the fractions in the gradient enriched in either ER or PM, the
marker enzyme activities arepresentedas
val-ues relative to other gradient samples. The [3H]UMP incorporatedacrossgradientsamples
wasinsufficientto serve as amarkerfor RNA
polymeraseactivity.
Thehigh level of cytochromec reductase
ac-tivity (Fig. 1C) in sample 4 (-42%)from both
the H-1 and H-3 preparations indicates that
residual mitochondria, as well as aggregated
debris,collect on the high-density(-55%)
cush-ion. The second peak of cytochrome creductase
activityat -37%intheH-1gradient could
rep-resent either larger sheets of rough ER, since
Mg2+ is present in the gradient, or
mitochon-drial membrane fragments. The areaof
reduc-taseactivity at 30 to 35% in the H-3 gradient is
compatible with the density properties of smooth ER.
The prominent ATPase activity (Fig. 1D) at
37% inthe H-1 gradient, indicating the
on November 10, 2019 by guest
http://jvi.asm.org/
VOL. 18, 1976
4 8 12 16 20
F R ACT 0N
FIG. 1. Distribution ofprotein (B), NADH cyto-chromecreductase(C),ATPase(D), and[3H]TMP incorporated (E) by gradient (A) samples after
equi-librium sedimentation (21 h at 25,000 rpm) ofa 1,000 xg,10-min(H-1) pellet( )andan80,000 x g, 30-min (H-3) pellet (-) from healthy
hamster brain.
ble sedimentationposition of PM, demonstrates
that the250to1,000 xgpellet comprisesa
PM-rich fraction. This suggests that the PM-rich
zone ofthe H-1 preparation might display an
equilibrium density comparable withaportion
of the rough ER (Fig. 1C). Other regions of
ATPase activity at 42% andthe disperse 30 to
35% region probably correspond to
mitochon-dria-rich and smooth ER-rich regions,
demon-strating nonspecific phosphatase activities.
In-PROPERTIES OF THE SCRAPIE AGENT 695
corporation of [3H]TMP is principally confined
to the mitochondria region (Fig. 1E) and is in
good agreement with the density indicated by
the cytochrome c reductase and ATPase levels. The equilibrium sedimentations of the 250 to
1,000 xg pellet (S-1) and 13,000 to 80,000 x g
pellet(S-3) from scrapie-infected brains present similar distributions of subcellular components (Fig. 2). Mitochondria and aggregated debris are indicated at about 42%, a PM-rich region is indicated at about 36.5% as noted by the high ATPase activity (Fig. 2D), and an ER-rich
re-gion isindicated atabout 30 to 35% as notedby
the diffuse cytochrome c reductase activity
(Fig. 2C). The[3HITMPincorporation profile of
theS-1 preparation is distinct from the healthy
profile, with a minor peak of activity localized
at about 36 to 37%, corresponding to the
PM-rich region described by the ATPase activity in
Fig. 2D. This putative DNA polymerase
activ-ity might result from mitochondrialfragments;
however, no corresponding cytochrome c
reduc-tase activity was detected (Fig. 2C).
The distribution ofSA inthe S-1preparation
demonstrates coincidence with the peak ATP-ase and [3H]TMP activity at 37%, suggesting
the possible associationofa PM-like constitu-ent with DNA polymerase activity. Asimilar
regionofSA isobservedinthe S-3preparation;
however, significant levels of SA are also
dis-played in the 42% and 30 to 35% regions.
There-fore, the DNA polymerase activity does not
totallyrepresentscrapie-specific activity. Since
the S-3 pellet did not resuspendwith
homoge-neousconsistency, theactivity at 42% may
re-flect aggregated debris. Nevertheless, the
membranes sedimenting at 30 to 35% appear to
be well dispersed and presumably represent a
smooth ER-rich region. Since it is difficult to
envision such ahigh concentration of PM-like
membranesat 30 to 35%after equilibrium
sedi-mentationfor21h, the datasuggest an
associa-tionof thescrapie agentwith smooth ERat30
to 35%.Itmightfollow then that the SAatthe
centerof thegradient (37%)could be relatedto
thepresenceofroughER.
Evidence for thePM-rich1,000 xgpellet and ER-rich80,000 xgpellet, aswellasthe
cosedi-mentation of PM androughERmembrane, is
provided by electronmicroscopeexamination of
fractions 6 through 10 from the S-1 and S-3
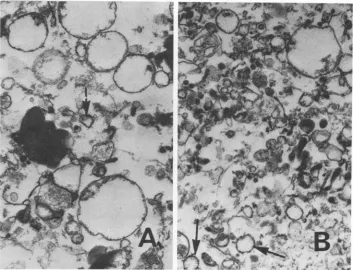
gradients (Fig. 3). It canbe observedthat this
ATPase-rich region of the S-1 contains
large
PM-likevesicles (Fig. 3A). Nevertheless, even
thoughnocytochromec reductase
activity
wasdetected in those samples, a trace amount of rough ER-like vesicles couldbe observed. The
presenceofmitochondrial fragments might
ex-plainthe [3H]TMP incorporationintothis
on November 10, 2019 by guest
http://jvi.asm.org/
[image:3.503.58.246.50.483.2]tion.
Conversely,
examination of the S-3prepa-3;A ration (Fig. 3B) indicates smaller vesicles,
3 40 some identifiable as rough ER, with a minor
quantity
oflarge
PM-like vesicles.30
_ Assuming that the scrapie agent might bea 41.8% 36.5% 32.6% a comprised, in part, of a nucleic acid component,
LO 4 4 the detection of the [3H]TMP incorporation
co-20
inciding
with
SA
lends credence
to
the
possible
o / % B involvement of nucleic acidmetabolism in the
o 4 ' synthesis of a scrapie agent-membrane complex
40 _ orthe
pathogenic expression
of the scrapiedis-rL
VA/
Vease. A similar interaction wassuggested
pre-Z
viously
for thepathogenic
(viroid) RNAofexo-l
\cortis
disease (Semancik etal.,
inpress).
With0
these considerations, a 13,000 to 80,000 x g
resuspended pellet
wassubjected
toequilib-a5
c rium sedimentation in a lighter gradient (-20to40%)asindicated in
Fig.
4A. In thisway, an- A
attempt
was madetopellet
theheavy
scrapie-> 3 A
containing
fraction (42%)whileachieving
abet-ter definition of the two
lighter
regions,
-37%1 $
l\and
30 to 35%(Fig.
4E). When thegradient
C <
samples
z_ t > \were_
assayed
for 13H]UMP(Fig.
4C),0
12m
4 ,,
and[3H]TMP
(Fig.
4D)incorporation,
distinct12 D profiles emerged, suggesting that
contamina-10 - tion by nuclear
fragments
was probably notF 8_ in v_ ^ _
prevalent.
The distribution of[3H]TMP
incor-X@6_ s \ | 5 \ _ poration coincided with SA, possibly
implicat-ing a DNA-related phase in scrapie
pathogen-4 -
esis.
2 m Totest for the presence ofa
scrapie-specific
product, gradient fractions from healthy and
infected brains were allowed to incubate over-600 E
night
in[3H]TMP
followedby
nucleicacidisola-tion (by phenol extraction). When analyzed by
EO
electrophoresis
on2.2%polyacrylamide
gels,
the400-
incorporation
product
couldbe detectedprinci-pally
inhigh-molecular-weight
DNAfrombothhealthy
andscrapie-infected preparations (Fig.
200 - 5). However, avery low,
reproducible
profile ofincorporation
intolow-molecular-weight
DNAcouldbedetectedinboth theS-1andS-3
prepa-,'7r;N
DF
'ration but notin thecomparable
fraction from109
healthy
tissue.
Nucleic acid
species
recovered
fromsubcel-i [////
A/////lular
( s 2fractions.Quantitative
andqualitative
o r determinations of nucleic acid
species
extractedfrom subcellular fractions were undertaken.
About 50 brainseachfrom healthy and
scrapie-infected hamsters were
Polytron homogenized
o / FIG. 2. Distribution ofprotein (B), NADH
cyto-LLI
///// \\\\ \\\\ ///_ chromec reductase (C), ATPase (D),[3H]TMPincor-poration (E), and SA (F) by gradient (A) samples
vr
//g;\\\\\\\\/// after equilibriumsedimentation(21hat25,000 rpm)0 7 / ofa 1,000 x g, 10-mm (S-i) pellet ( ) and an
2 4 6 8 10 12 14 80,000 x g,30-min (S-3) pellet(-) from scrapie-F RA CT 0N infected hamster brain.
696
SEMANCIK ET AL. J. VIROL.on November 10, 2019 by guest
http://jvi.asm.org/
[image:4.503.87.227.49.658.2]PROPERTIES OF THE SCRAPIE AGENT
697
FIG. 3. Electron micrographs ofgradient fractions (7through9)ofa1,000 xg,10-minpellet(A) andof gradient fractions (6 through 10) of an 80,000 xg, 30-minpreparation (B) from scrapie-infectedhamster brain (see Fig. 2). Arrows indicate rough ER vesicles. x39,000. N
and fractionated as described above, followed by nucleic acidisolation by phenol extraction.
Ethanol-precipitable nucleic acids were
parti-tioned by2 M LiCl precipitation of
ribosomal-like RNA and recovery of
high-molecular-weight DNA after addition of 3 volumes of
ethanoltothe 2MLiClsupernatant. Determi-nations of DNA and RNA by diphenylamine and orcinol tests were made on each of the nucleic acid fractions.
A significantly higher level of extractable
nucleic acids was detected in all subcellular fractions from healthy, ascompared with
scra-pie-infected, braintissue. Thismayreflect the widespread cellular degeneration associated
with thescrapie disease process. The
distribu-tion ofnucleic acidspecies reflects the distinct quality of the subcellular fractions. The 1,000 and 13,000 x g pellets contain relatively high concentrations ofDNA (50 and 35%),
presuma-bly resulting from nuclei and mitochondria, whereas the80,000 and100,000 xgpelletsare
rich (80 and 95%) in RNA, reflecting the
pres-ence ofmembrane-bound and free
ribosomes,
respectively. When aliquots fromhealthy and
scrapie-infected preparationswereanalyzed by
electrophoresis on 2.2 or 5% polyacrylamide
gels, no differences in thequality of the various
RNA species wereevident.
DISCUSSION
Previous attempts to describe an infectious
scrapie nucleic acid species, analogous to the
pathogenic (viroid) RNA, have not been
suc-cessful (16, 23). Renewed interest in the
com-parative pathology ofscrapie disease with vi-roidinfectionsinplants wasstimulated by the
finding of amembrane-relatedphase in
exocor-tisdisease (Semancik et al., in press) and the
observation of acytopathic effect onthe
endo-membrane system (Semancik and
Vander-woude, Virology, in press).
Therefore,
pro-ceduresemployed in the characterization of the
exocortis disease agent have been adapted for
use inexaminingthe scrapieagent membrane
complex
andpossibly
a putative nucleic acidcomponentofthe scrapie agent. These studies
have been facilitated by the use of the
ham-ster-scrapie model, providingthehighesttiters of scrapie
infectivity
of any known bioassaysystem (15).
The concentration of SA in the subcellular
VOL. 18, 1976
on November 10, 2019 by guest
http://jvi.asm.org/
[image:5.503.80.433.67.337.2]IV 2 4 6 8 10 12 14 16
FRACTION
FIG. 4. Distribution ofprotein (B), RNA polym-eraseactivityasindicated by [3H]UMPincorporation
fractions
pelleted
inthe 250 to80,000
xg rangegenerally
supports
thescrapie-membrane
hy-pothesis
(8), i.e.,
thepossible
association ofasmall
pathogenic
nucleicacid,
asdeduced fromthe
target
size of -105daltons(2),
withamem-brane component. The primary association
with the PM
(17)
cannot befully
reconciledwith the present data.
Samples
taken fromequilibrium density
gradients,
even afterpre-fractionation of subcellular components, are not
sufficiently homogeneous
toascribeSA toasingle
membranespecies.
Thepresent
datain-dicatethat
portions
oftherough
ERcosedimentwith the PM-rich
regions. Therefore,
thepre-ferred
explanation
ofthesedimentationhetero-geneity
ofthescrapie
agentin13,000
to80,000
x g
(S-3)
pellets
suggests an association withrough
ERataboutadensity
of 1.17g/cm3
andwith smooth ER ata
density
of 1.13to 1.15g/
cm3.
Alternatively,
thedensity heterogeneity
of theSAcomplex
mayresultfromthespecialized
nature ofthe brain
tissue,
since thesynaptic
vesicles and external synaptic membranes demonstrate
characteristically
differentsedi-mentationproperties (24).
Also,
theassociationof the presumptive nucleic acid component in
the
scrapie-membrane
complex may result inan alteration in the
density
of normal host membranes.When consideringthe high specific
infectiv-ity
ofthe scrapie agent inthe absence of anyrecognizableagent,onemustbe extremely cau-tious in
ascribing
aspecific
association to asingle
component of the endomembrane sys-tem.Furthermore,
iftheagentbecomesassoci-ated with theendomembrane system at
early
developmental periods,
it may be possible forthe agent to retain biological activity
during
andafterthe process ofmembrane
differentia-tion,
thusresulting
insomeSAintegratednotonly
with the ER but also with the PM. Afurther considerationcenters ontheassociation
ofthe SA with some scrapie-directed de novo
membranestructure orpossibleartifacts of
sub-cellularfractionation which cosediments with
either PM or ER but is not discernible as a
distinct component ofthe membrane popula-tion.
The data are compatible with the
hypothesis
presentedfor the pathogenesis ofthecitrus
exo-cortis viroid (Semancik et al., in press), in
which both a nuclear (19) and an
endomem-brane
phase
havebeenpostulated
forthepossi-(C), DNA polymerase activity as indicated by
[3H]TMPincorporation (D), and SA (E) bygradient
(A) samples after equilibrium sedimentation (21 h
at25,000 rpm) ofan80,000 xg,30-min (S-3)pellet fromscrapie-infected hamster brain.
J. VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:6.503.70.251.44.622.2]AGENT
I I
B
50~~~~~~~~~~~~
0 10 20 30
F R ACTI0N
FIG. 5. Electrophoresis of[3H]TMPincorporation products of subcellular fractions 1,000 x g,10 min (A) and80,000xg,30min(B) from healthy (0)and
scrapie-infected (@) hamster brain in 2.2% poly-acrylamide gels with 0.5%agarose after4 h and6
mA/gel. Migrationwasfromlefttoright, with3-mm
slices treatedinOmnifluor-toluenewith 3% Protosol
for48hat40C.
ble siteof synthesis and site of accumulation of
the pathogenic RNA. The scrapie-membrane
complex may reflect a synthetic end point or
accumulation ofthe scrapie agent. Sincewedo
not know the molecular nature of the scrapie
agent, much less the template and pathway by
which the agent is synthesized, the most
cau-tious interpretation of the present data would
be thatthescrapieagentappearstobe
primar-ily associated with ER-like, and possiblysome
PM-like, membranes which may represent
sitesofaccumulation of scrapieprogeny.
The correlation of [3H]TMP incorporation or
DNApolymerase activity with SAcanbe taken
onlyas averypreliminary indication that this
site of accumulationmayalsorepresentasite of
some synthetic activity of the scrapieagent. A
low-molecular-weight DNA species has also
been reported in scrapie-infected mousebrain
after the intracerebral inoculation of
[3H]thy-midine (10). These data, together with the
dynamics of [3H]thymidineturnoverin
scrapie-infectedmousebrain (1,12)and the correlation
between cell division and scrapie infectivity
(4), are compatible with the suggestion of a
DNAcomponentinthe scrapie agent.
The characteristics ofscrapiediseaseand its transmissible agent present interesting paral-lels to the plant disease caused by the citrus exocortis viroid. The association of the
patho-genic(viroid) RNA of the exocortisdisease with
the endomembrane system appears to result in
a vesicular proliferation or disruption of PM
(Semancik and Vanderwoude, in press). Since the overt pathology of scrapie disease also in-volves aberrations of the PM (13), these cellu-lar phenomena, coupled with the estimates of
molecular size and stability, reinforcethe
possi-bleviroid nature of the scrapie agent (6).
Com-parative aspectsof the two diseases project the
possibility of a nucleic acid component in the scrapie agent, as well as a nuclear phase
in-volved in the replication process. The hamster
scrapie model presents an excellent system to exploit in the study of membrane-"viroid"
inter-actions,eventhoughamoleculardescriptionof
the scrapie agent, fundamental to the
under-standing of the disease, has yet to be estab-lished.
ACKNOWLEDGMENTS
These investigations weresupported byaNational Sci-enceFoundation grant (GB 39605) to J. S. Semancik andby Public Health Service grant AI 11250 fromthe National Institute of Allergy and Infectious Diseases.
Wewish to acknowledge theexcellenttechnical contri-butions of Deanna Tsuruda, Kazue Matsumoto, and D. A. Reynolds,and thankW.VanDerWoudeandR.Leonard for advice and consultation.
LITERATURE CITED
1. Adams, D. H. 1972. Studies on DNA from normal and scrapie-affected mouse brain. J. Neurochem. 19:1869-1882.
2. Alper, T., D. A. Haig, and M. C. Clarke. 1966. The
exceptionally small size of the scrapie agent. Bio-chem.Biophys. Res. Commun.22:278-284.
3. Bishop, D. H. L., J. R. Claybrook,and S.Spiegelman, 1967.Electrophoretic separation of viralnucleic acids
onpolyacrylamide gels. J. Mol. Biol. 26:373-387.
4. Clarke,M.C.,and D. A.Haig. 1970.Multiplicationof scrapie agentincell culture. Res. Vet. Sci. 11:500-501.
5. Diener, T.0. 1971. Potato spindle tuber "virus."IV. A
replicating low molecular weight RNA. Virology 45:411-428.
6. Diener, T.0.1972. Isthe scrapie agent aviroid? Nature (London) 235:218-219.
7. Dougherty, R. M. 1964. In R. J. C. Harris (ed.), Tech-niques in experimental virology, p. 183. Academic PressInc.,NewYork.
8. Gibbons,R.A., and G. D. Hunter. 1967. Nature of the scrapie agent.Nature(London) 215:1041-1043.
9. Hodges, T. K., andR.J. Leonard.1974.Purification of
a plasma membrane bound adenosine triphosphate from plant roots, p. 392-406.InS.P.Colowick andN.
0. Kaplan (ed.), Methodsinenzymology, vol. 32, part
B.AcademicPressInc., New York.
10. Hunter, G. D., R. H. Kimberlin, S.Collis, and G. C. Millson. 1973. Viral and non-viral properties of the scrapie agent. Ann.Clin. Res. 5:262-267.
11. Kimberlin, R.H., andR. F.Marsh.1975. Comparison ofscrapie and transmissible minkencephalopathyin
VOL. 18, 1976
on November 10, 2019 by guest
http://jvi.asm.org/
[image:7.503.57.237.54.311.2]hamsters.I. Biochemicalstudiesofbrainduring
de-velopment of disease. J.Infect. Dis. 131:97-103. 12. Kimberlin, R. H.,D. B. Shirt,andS. C.Collis.1974.
Theturnoverof isotopicallylabelled DNAin vivo in
developing, adult and scrapie-affectedmouse brain.
J.Neurochem.23:241-248.
13. Lampert, P., C. J. Hooks, Jr., and D. C. Gajdusek.
1971. Altered plasma membranes in experimental scrapie.ActaNeuropathol. 19:81-93.
14. Luft,J. M.1961. Improvementsinepoxyresin
embed-ding methods.J.Biophys. Biochem.Cytol.9:409-414.
15. Marsh,R.F.,andR.H.Kimberlin.1975.Comparison of scrapie and transmissible mink encephaolopathy
inhamsters.II.Clinical signs, pathology, and
patho-genesis. J.Infect. Dis. 131:104-110.
16. Marsh, R. F., J. S. Semancik, F. C.Medappa, R. P.
Hanson, and R. R. Rueckert. 1974. Scrapie and transmissible minkencephaolopathy: search for
in-fectious nucleic acid. J. Virol. 13:993-996.
17. Millson, G. C., G. D. Hunter,and R. H. Kimberlin.
1971. An experimental examination of the scrapie agentincellmembrane mixtures.II.Theassociation
ofscrapie activity withmembrane fractions. J. Comp. Pathol.81:255-265.
18. Semancik, J. S., T. J. Morris, and L. G. Weathers.
1973. Structure andconformation of low molecular weightpathogenicRNA from exocortisdisease.
Virol-ogy53:448-456.
19. Semancik,J.S.,and J. L. M. C. Geelen.1975.Exocortis disease: detection of DNAcomplementaryto
patho-genic (viroid)RNA. Nature(London)256:753-756. 20. Semancik,J. S.,and L. G. Weathers. 1972. Exocortis
disease: an infection free-nucleic acid plant virus
with unusualproperties.Virology47:456-466.
21. Semancik,J.S., and L. G.Weathers. 1972. Exocortis disease: evidence foranewspeciesof"infection" low
molecular weight RNA inplants. Nature (London) New Biol.237:242-244.
22. Shatkin, A. J. 1969. Colorimetric reactions forDNA,
RNA, andprotein determination, p. 231-237. In K. Habel and N. P. Salzman(ed.), Fundamental tech-niquesinvirology. Academic Press Inc., New York.
23. Ward, R. L., D. D. Porter, andJ. G. Stevens. 1974.
Nature of thescrapieagent: evidenceagainsta vi-roid. J.Virol. 14:1099-1103.
24. Whittaker, V. P. 1966. Some properties of synaptic membranesisolated from the centralnervoussystem.
Ann. N.Y. Acad. Sci. 137:982-998.
J. VIROL.
![FIG.1.1,000incorporatedlibriumhamsterchromex Distribution of protein (B), NADH cyto- c reductase (C), ATPase (D), and [3H]TMP (E) by gradient (A) samples after equi- sedimentation (21 h at 25,000 rpm) of a x g, 10-min (H-1) pellet () and an 80,000g,30-min(H-3)pellet(-)fromhealthy brain.](https://thumb-us.123doks.com/thumbv2/123dok_us/1559628.108582/3.503.58.246.50.483/incorporatedlibriumhamsterchromex-distribution-protein-reductase-gradient-samples-sedimentation-fromhealthy.webp)
![FIG.2.poration80,000chromeafterofinfected a Distribution of protein (B), NADH cyto- c reductase (C), ATPase (D), [3H]TMP incor- (E), and SA (F) by gradient (A) samples equilibrium sedimentation (21 h at25,000 rpm) 1,000 x g, 10-mm(S-i) pellet () and an x g, 30-min (S-3) pellet (-) from scrapie- hamster brain.](https://thumb-us.123doks.com/thumbv2/123dok_us/1559628.108582/4.503.87.227.49.658/poration-chromeafterofinfected-distribution-reductase-atpase-gradient-equilibrium-sedimentation.webp)
![FIG. 4.erase Distribution of protein (B), RNA polym- activity as indicated by [3H]UMP incorporation](https://thumb-us.123doks.com/thumbv2/123dok_us/1559628.108582/6.503.70.251.44.622/fig-erase-distribution-protein-polym-activity-indicated-incorporation.webp)
![FIG. 5.productsforslicesscrapie-infectedacrylamide(A)mA/gel. Electrophoresis of[3H]TMP incorporation of subcellular fractions 1,000x g, 10 min and 80,000 x g, 30 min (B) from healthy (0) and (@) hamster brain in 2.2% poly- gels with 0.5% agarose after 4 h and 6 Migration was from left to right, with 3-mm treated in Omnifluor-toluene with 3% Protosol 48 h at 40 C.](https://thumb-us.123doks.com/thumbv2/123dok_us/1559628.108582/7.503.57.237.54.311/productsforslicesscrapie-infectedacrylamide-electrophoresis-incorporation-subcellular-migration-omnifluor-protosol.webp)