0022-538X/79/04-0157/09$02.00/0
Structural Studies of Retroviruses: Characterization of
Oligomeric Complexes
of Murine and
Feline
Leukemia Virus
Envelope and Core Components
Formed
upon
Cross-Linking
ABRAHAM PINTER* AND ERWIN FLEISSNER
MemorialSloan-KetteringCancer Center, New York, New York 10021 Received forpublication13September1978
Toexamine the protein proximity and subunit organization of type C
retrovi-ruses,preparations of AKRmurine leukemia virus were treated with bifunctional
cross-linkingreagentsandanalyzedby sodium dodecylsulfate-polyacrylamidegel
electrophoresis (SDS-PAGE).Thecross-linkedcomponentsobtained were
char-acterized by imnmunoprecipitation with monospecific antisera against purified
viral proteins,followed by SDS-PAGE analysis both before and after cleavage of
the cross-links.Withtheseprocedures, complexes of both viral envelope and core
components were identified. The major envelope subunit obtained was a large
(apparent molecular weight of 450,000 to 500,000), glycosylated complex,
com-posedoffourtosixgp7O-pl5(E)subunits. Thiscomplex was detected over a
100-fold range of cross-linker concentration and thus seems to represent a particularly
stable viral substructure. The cross-linked complexes of the core proteins
con-sisted of oligomers of p30 dimers, suggesting that the p30 dimer is a basic
structural unit of the viral core. When virionpreparations, which had previously
beendisruptedwith the nonionicdetergent Nonidet P-40, were cross-linked, the
envelope complexwasstill observed, indicating that this structure is stable in the
presence of Nonidet P-40. A similar envelope structurewasobserved for feline
leukemia virus,suggesting that such a complex may be aconserved feature of
oncornavirusstructure.
The type C oncornaviruses possess distinct
core and envelope morphological structures
which have been visualized in the electron
mi-croscope. The presence of surface "knobs"
ap-proximately8.0to10.0nmin diametercovering
theenvelopeof intactvirionshas been
demon-strated afternegative stainingof murine
leuke-mia virus(MuLV) preparationswithuranyl
ace-tateorbyacombination ofphosphotungsticacid
staining followedby freeze-drying (5, 22). Reg-ularly arranged subunits of viral cores, which
possess a diameter of approximately 6.0 nm,
havealsobeenvisualized afterfreeze-dryingand
shadowingof viral coresreleased after various
treatments(22). The MuLVenvelopehas been
showntocontaintwomajor proteincomponents,
a large glycosylated protein of
approximately
70,000daltons, gp70 (1), and asmaller,
nongly-cosylated hydrophobic protein,p15(E)(13);both
arederivedbyproteolytic cleavageofa common
precursor (8, 15, 26, 28). Thetwoenvelope
com-ponents can, underappropriate conditions,form
adisulfide-linked complex, referredto as[gp9O]
(23, 24), demonstrating an integral association
between gp7O and p15(E). The major intemal
structural components of the virus consist of four proteins-p30, p15, p12, and plO-which
are derived by proteolytic processing of the
"gag"geneproduct (7).Tobettercorrelateviral
proteins with the morphological substructures
seen in virions, we have studied the protein proximitiesandspatialarrangements ofproteins inviral substructures by treating virions with
two chemical cross-linking reagents,
dithiobis(succinimidyl proprionate) (DSP) and
1,5-difluoro-2,4-dinitrobenzene (DFDNB).Both
reagents contain two functional groups which
react readily at physiological conditions with
lysine amino groups, thus allowing the cross-linkingofneighboring proteins. DSP possesses
an internal disulfide bond; thus cross-links
formed withthis reagentcanbereadilycleaved
by reduction, facilitating the analysis of the
cross-linked products. These studies have
re-sultedintheidentificationofspecificcomplexes
of both viral envelope and viral core
compo-nents; these complexes may be related to the
morphological substructures which have been
visualizedin the electronmicroscope.
MATERIALS AND METHODS
Viruses.AKRMuLVwasobtained fromproducer
SC-1celLs,andfelineleukemia virus (FeLV)ABwas
obtained from aninfectedfeline lungfibroblast cell 157
on November 10, 2019 by guest
http://jvi.asm.org/
line. Viruses were radiolabeled by culturing in the
presence of either "C-labeled mixed amino acidsor
3H-labeled glucosamineprecursors,ataconcentration
of 100 uCi/ml, for 18 h, and purified by banding directlyon15to60%sucrosegradients. Concentrated
AKR MuLV, obtainedfrom Electro-Nucleonics Lab-oratory Inc., was labeled with [3H]NaBH4 by the method of Gahmberg and Hakomori (10).
Electrophoresis. Sodium dodecyl sulfate-poly-acrylamide gelelectrophoresis (SDS-PAGE)was
per-formed in 7to21%gradientslabgels by thesystem described by Laemmli (18). Gels were analyzed by
fluorography, accordingtotheprocedure described by Bonner andLaskey (3).Samples analyzed under
non-reducing conditionswereboiledinthepresenceof 1%
SDS for 1 min. Samples analyzed under reducing conditionswereboiled in thepresenceof1% SDS and 1%mercaptoethanol for1min.
Alkylation of virionswith NEM. A stocksolution of10%N-ethylmaleimide (NEM) in acetonitrile was
diluted 1:10 with phosphate-buffered saline (PBS). One volume of the PBS solutionwasthenaddedto9 volumes of the virus sample and incubated atroom
temperaturefor 15min.
Cross-linkingprocedure.Stock solutions of the cross-linking reagentswere prepared in Me2SOata
concentrationof 20 mg/ml. Thesewerethendiluted
with PBStoaconcentration fivefoldgreaterthan the desiredfinal concentration, then further diluted 1:5 with the virussample. DSP reactionswereperformed
in the presence of TN buffer (0.01 M Tris-0.10 M
NaCl); for DFDNB reactions, virus samples prepared inPBSwereused. Reaction timesweretypically 15
minatroomtemperature.Occasionally, unreacted
re-agentwasquenched by the additionof0.1 M
ammo-niumacetatebeforelysis of virions; however, similar results were obtained if thisstep wasomitted. The
concentration ofradiolabeled virions used in these studies wasless than 20 ig/ml. Similar patterns
re-sulteduponcross-linkingof virions ataconcentration of1mg/ml.
Radioimmunoprecipitations. Virionswerelysed
by the addition of NonidetP-40(NP-40) and NaClto
final concentrations of 0.5% and 0.5M, thenincubated with antiseraatafinal concentration of 1:50for1hat 37°C. Immune complexeswerethenpelleted by
treat-mentwith50
id
ofa10%solution ofstaphylococcus A(Pansorbin, Calbiochem) prepared as described by
Kessler (17). Afterpelletingat3,000rpmfor 10mi,
thestaphylococcus Awaswashedwith 10 mlofhigh salt(1MNaCl,0.5%NP-40,0.02MTris) and10mlof low salt(0.004 M Tris, 0.5% NP-40) buffers, pelleted, and then extracted with 0.1 ml ofSDS-containing bufferat100°Cfor 1min.
Reagents.Goat anti-Rauschergp7O andanti-p30
sera were obtained from R. Wilsnack; rabbit anti-p15(E) was prepared as described (13). DSP and
DFDNB were obtained from Pierce Chemical Co.,
SDS(specially pure)wasobtained from BDH
Chem-icalsLtd.,NEM wasfrom Sigma Chemical Co., and
radiochemicalswereobtained from NewEngland
Nu-clearCorp.
RESULTS
Cross-linking of AKR MuLV with DSP. We havepreviously shown that themajority of
the viral envelope proteins, gp70 and p15(E),
exist in native virionsinthe formofa
noncova-lently linked complex. Upon disruptionofvirions
with NP-40, a covalent disulfide linkage can
forn spontaneously between thetwo proteins,
resulting in the stable gp7O-p15(E) complex
re-ferred to as [gp9O]. In addition, activation of
appropriatethiol residues ofgp70 and p15(E) by
treatmentwith thethiol-specificreagentsNEM
ordithiobis(m-nitropyridine) (DTNP)results in
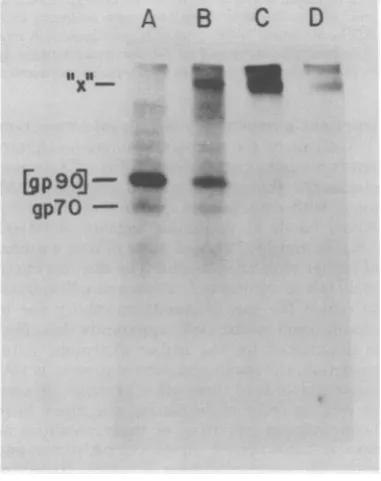
formation of[gp9O]inintact virions(23, 24) (Fig.
1). Samples of 14C-amino acid-labeled AKR
vir-ionsanalyzedby SDS-PAGE under nonreducing
conditionscontainedgp70andp15(E) (Fig. 1A);
aftertreatmentwithNEM, the gp70 bandisnot
observed, the p15(E) band diminishes in
inten-sity, and the [gp9O]bandappears(Fig. 1B). To
determine whether [gp9O] waspart ofalarger
envelope structure, virions pretreated with
NEMwerereacted with DSP andanalyzed
un-dernonreducingconditions.Atafinal DSP
con-centration of 10,g/ml,newbands with apparent
molecular weightsof 60,000 (a) and 120,000 (b)
wereobserved, along witha more diffuse band
atamolecularweightofapproximately 180,000
(c),andaband ofhigher molecular weight ("x")
(Fig. 10).When theconcentration of
cross-link-A B C D
S
RT
-
gp7O-MD
A
_0
x
- c
-b
0-
P30-p15
(E)-FIG. 1. Cross-linking of "C-amino acid-labeled AKRvirions with DSP. Virus samples were analyzed
bySDS-PAGE under nonreducing conditions after thefollowing treatments: (A) untreated virus; (B) +NEM; (C) +NEM, +DSP (10pg/ml); (D) +NEM, +DSP (100pg/ml). "x" represents the cross-linked envelope component.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:2.501.265.457.355.611.2]VOL. 30, 1979
mgreagent wasir
icant decrease in
viral protein ban bandsa,b,and c, inmaterialrunnix
To ascertain u
tained gp7O, the
peated with virio
had been specific
dase oxidation,
[3H]NaBH4
(10). tern obtained foinonreducing con
served
correspond
ofgp7O and only
ular-weight comj
show thepatterns
10, 100,and1,000
At the lowest DE
ponentwith the
withsome smalli 2B).At 100,ugof the
[gp9O]
has beE itant increase in bpercentageof the
component"x" ai
DSP per ml, alti
mostof theprotei
itX'
bp9g-gp7O
-FIG. 2. Cross-lin Virions, for which NaBH4reductionc (10), were analyze
ducingconditionsc +NEM; (B) +NEA +DSP(100pg/ml);
CROSS-LINKED COMPLEXES OF MuLV AND FeLV 159 icreasedto100l,g/ml,asignif-
larger
aggregateswhichcan nolonger
enterthe i intensity of the monomeric gel(Fig. 2D).
Theseexperiments
indicate thatds was observed, as well asof band "x" containsacross-linked
complex
oftheand therewas a
large
increase viralglycoprotein;
the observations that it isng atposition "x"(Fig. 1D). detectedover a100-foldconcentration range of
vhich cross-linked bands con- the
cross-linking
reagent and thatwell-charac-above experiments were re- terized
complexes
of smaller sizeare notreadily
)ns in which the glycoprotein observedsuggest that the
large complex
repre--ally labeled by galactose oxi- sents a
particularly
stableviral substructure.followed by reduction with The size of the
glycosylated complex
wases-Figure 2A shows the gelpat- timated
by comparison
of its SDS-PAGEmo-r NEM-treated virions under
bility
with that of the LETSmonomeranddimerditions. The major band ob- purifiedfromJLSV9cells
by
immunoprecipita-dsto[gp9O], withafaintband tion with
monospecific
antiserumprepared
trace amounts ofhigh-molec-
against
human cold insolubleglobulin (25).
Theponents.
Lanes B, C, and D LETSprotein
monomerhasamolecularweight
3obtainedaftertreatmentwith of 220,000 and has been shown to exist as ajug
ofDSPperml, respectively. disulfide-linked dimer(16).
Such ananalysis
3P concentration, a new com- showed that the cross-linkedcomplex
has amobility of "x" appearsalong
slightly
lowermobility
than that of thepurified
Dr, more diffuse material (Fig. LETS dimer
(data
notshown;
seeFig.
5), andDSP perml, essentially allof thuswehave
assigned
itanapparentmolecularancross-linked,witha concom-
weight
of450,000to500,000.and "x"(Fig.2C). Asignificant Characterization of DSP-cross-linked
radioactivity isstillpresent as components by immunoprecipitation. To
ftertreatmentwith
1,000,ug
of determine thecomposition
of the cross-linked iough under these conditions, components obtained after DSPtreatment,
inhas been cross-linkedtostill cross-linkedvirionswere
lysed
withNP-40,andthecross-linked
complexes
wereisolatedbyim-A
B
C D munoprecipitation with monospecific antiseraagainstindividual viralproteins.Thisprocedure
hasthe
advantage
thatit allows theanalysis
of3 -^"': individual
complexes
evenincaseswhere theremay beseveraldifferentcross-linked
complexes
of similar size. Thecomponentsof the
complexes
arethusidentifiedby their
antigenic reactivity
as well as by their SDS-PAGE mobility after
reduction.
Figure
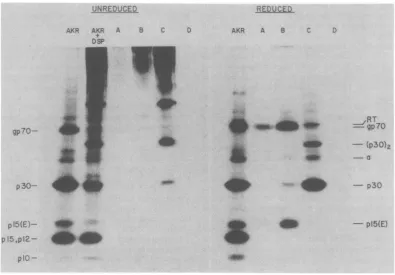
3 shows the SDS-PAGEpattern
ob-tained for
'4C-amino
acid-labeled AKR virionsbeforeandafter
cross-linking
with 100 ,Lgof DSPperml. In this
experiment,
the NEMtreatmentwasomitted. The
pattern
obtained aftercross-linking
wassimilartotheonedescribedearlier; almost all of the gp70 and most ofthep15(E)
have beencross-linked,
andthereisasignificant
reduction in theintensityof the
p30
band. The60,000-and
120,000-molecular-weight
bandsare readily apparent, and there is a considerableamount of
high-molecular-weight
materialwhich is morediffuse.Uponreaction with
anti-gp70 serum, very little radioactivity is
precipi-tated; this suggests that the DSP treatment
kingofAKRMuLVglycoproten3s.
resulted in a considerable modification of the gp70 had been labeled by[3H]-*zfter
galactoseoxidaseoxidationgp7O
moleculewith
aconsequent loss ofimmu-,d by SDS-PAGE under nonre- noreactivity. A faint band is observed for the
zfterthefollowingtreatments: (A) anti-gp70
precipitate (Fig.
3A). In the unreduced, +DSP (10pg/mI); (C) +NEM, sample, it has the
mobility
of"x";
afterreduc-(D) +NEM, +DSP
(1,OX
pg/ml).
tion, the disulfide bonds of thecross-linking
on November 10, 2019 by guest
http://jvi.asm.org/
[image:3.501.55.246.366.607.2]UNREDUCED REDUCE AKR AKR A
DSP
a,.
gp70- £
p30--p 15(E)-p15,p12 -p'0
-AitA.
B C D AKR A B c D
f.
_w
a
a
[image:4.501.60.456.72.347.2]S
FIG. 3. Immunoprecipitation of DSP cross-linkedcomponents. "C-aminoacid-labeledvirionswere
cross-linked with100pg/mlof DSP, lysed with 0.5% NP-40 and0.5MNaCI, and treated with: (A) anti-gp7O serum;
(B) anti-p15(E) serum; (C) anti-p30serum; (D) normalgoatserum.Immunecomplexeswerecollected with
staphylococcusAand, after washing, weredissolved inSDS-containing buffer and analyzed both with and
withoutadding mercaptoethanol. The bandlabeled RT has been characterizedas reversetranscriptaseby virtueof its identical mobility with that of thepolypeptideimmunoprecipitated with monospecific antiserum
topurifiedreversetranscriptase.
reagentarecleaved, andaband withthe
mobil-ity of free gp7O is observed. The sample istoo
dilutetodetectanyadditionalcomponents
pres-ent.
Reactionwith antisera top15(E) resulted in the immunoprecipitation ofconsiderablymore
material(Fig. 3B); apparently the immunoreac-tivity of p15(E) ismoreresistanttomodification. Upon analysis under nonreducing conditions,a
large diffuse band, similartothat recognized by the anti-gp7O serum, is observed. After
reduc-tion, this band is showntoconsist of onlytwo
majorcomponents, gp70and p15(E), indicating thatalarge complex of gp7O and p15(E) is
sta-bilized by DSP treatment. This complex does
not contain any p15, p12, or pLO; however a
slightamountof p30 isobservedafter reduction (Fig. 3B). It isnotclearwhether this is dueto
some trapping of cross-linked p30 aggregates with the cross-linked envelope complexes or
whether this reflects actual cross-linking
be-tweenp30 and the envelopecomponents.Inany
case,the relativeamountof p30 observed in this sample isso low that this does not appear to
representastructurally significant interaction.
Analysis of the anti-p30 immunoprecipitate undernonreducingconditions(Fig. 3C) demon-stratesthe presence ofasmall amount ofp30, along with considerable amounts of the
cross-linked bands at molecular weights of 60,000, 120,000, and 180,000, andmore diffusematerial
ofhigher molecular weight. The disproportion-ately low recognitionofmonomericp30appears
to reflect the loss ofimmunoreactivity due to modificationbytheDSP; apparentlythiseffect
is diminished for the higher oligomers. After
reduction, the major component presentisp30; minoramountsof three otherproteinsareseen as well. In order ofdecreasing size, these have
been assigned by virtue of their mobilities as
reversetranscriptase, unreduced p30 dimer,and
actin. Theco-precipitationofreverse
transcrip-tasewithp30has beenpreviouslyobserved(14), and thus isnotanindicationthat the two
com-ponentswere cross-linked. Actin has also been
observedtobea contaminantof viral
immuno-precipitates, and thus it is not clear whether there isanactualp30-actincross-linkedcomplex
_RT
- gp7O -(P3)2
-a
- p30
- p15(E)
on November 10, 2019 by guest
http://jvi.asm.org/
CROSS-LINKED COMPLEXES OF MuLV AND FeLV 161 formed by DSP treatment.
Treatment with nonimmiune serum did not
result in the precipitation of any labeled com-ponents (Fig. 3D), indicating that nonspecific adsorption of the cross-linked components to staphylococcus A is not occurring.
DSP treatment of NP-40-solubilized vir-ions. Recent studies have indicated that certain
MuLV morphological structures can survive
NP-40 treatment (9, 31). To study the MuLV
substructures which exist in the presence of
non-ionic detergent, AKR virions were disrupted
with 0.5% NP-40 and subsequently cross-linked
with 100
fig
of DSP per ml. Cross-linking ofintact virions results in the pattern described
earlier; gp7O and p15(E) are cross-linked into a large complex, and p30 oligomers are formed (Fig. 4B). DSP treatment of NP-40-lysed virions still results in the cross-linking of gp7O and
p15(E); however the p30 oligomers do not form,
thusallowingtheclearresolution of the envelope
complex (Fig.4C). A similar effect is observed
for virions pretreated with NEM (Fig. 4C').
These results indicate that the large gp7O-pl5(E)
A B C A' B' C'
_-, _f_ a _ Sto
_~~~~~~i
[image:5.501.53.244.326.560.2]--dslo.91
[gp9go]
gp70
-p15(E)
FIG. 4. DSP cross-linking of AKR virions dis-rupted with NP-40. "4C-amino acid-labeled virions analyzed bySDS-PAGE undernonreducing condi-tions after the following treatments: (A)
uncross-linkedvirions; (B) cross-linked with 100jpgof DSP perml; (C) disruptedwith 0.5% NP-40for15minat
roomtemperaturefollowedby cross-linking with 100
pgofDSPperml. Samples A',B' and Cwere
pre-treated with0.1% NEM before theother manipula-tions to convert gp7O to thegp7O-pl5(E) disulfide-linkedcomplex [gp90].
complex is quitestable in the presence of non-ionic detergent, whereas the p30 complexes are not.Itisbelievedthatnonionicdetergentsbind
primarily to hydrophobic regions of proteins
(12); thus thedissociation of the p30 complexes
by NP-40 suggests that the p30 associationsare
duemainly tohydrophobic interactions.
Cross-linking of AKR virions with DFDNB. To further characterize the subunit composition of native oncornaviruses, intact preparations of AKR MuLV were treated with DFDNB, a bifunctional cross-linking reagent
which reactsreadily with protein amino groups,
resulting in theformation of cross-links with a
maximum span of 0.5 nm. Since this span is
considerablyshorterthan that of DSP
(approx-imately1.1nm),DFDNBis useful as aprobe for
protein associations of closer proximity than
those detectedby DSP. For thisparticular
ex-periment,aviruspreparationat aconcentration
of 1 mg/mlwas used, and the proteinpattern
wasdeterminedby staining thepolyacrylamide
gelswithCoomassieblue. The patterns observed
upon cross-linking were independent of virus
concentration,andsimilar results were obtained
for more dilute, radioactively labeled
prepara-tions.
Treatment of intact virions with 100 ,ug of
DFDNBperml resulted in the loss of thegp7O
band and the appearance of
higher-molecular-weight material which possesses a similar mo-bility to that of the envelope complex formed
with DSP (Fig. 5C).There does not appear to
be adetectable decrease in theintensityof the
p30 band, and bands corresponding tothe p30
oligomersarenotpresent.DFDNBtreatmentof
virus which had beenpretreatedwith NEM also
resulted in the cross-linking of gp7O and the
resolution of a large complex (Fig. 5D). This
band appears slightly larger than the
corre-spondingbandobserved in laneC;this may be
an indication that thecomplexformedwithout
NEM treatment contains less than the
maxi-mum number of gp7O or p15(E) constituents.
For the NEM-treated virions, DFDNB
treat-mentdoesresult inadecrease in theintensityof
the p30 band, indicatingthat cross-linkinghas
occurred; however, the p30 intermediates
de-tected after DSP trea,tment are not observed.
Thismay indicatethat NEM treatmentinduces
achangeinconformation of thep30 molecules,
whichallowstheoccurrenceoflarge-scale
cross-linking by DFDNB. Figure 5E shows the gel
patterns
obtainedafter DSPcross-linking.The p30 dimer,tetramer, and hexameraredetected, and a broadband,encompassingthemobilitiesof thecross-linkedenvelopecomplexesresolved
inlanesCandD,isobserved.
VW4..
4WAIP
4..
VOL. 30, 1979
on November 10, 2019 by guest
http://jvi.asm.org/
162 PINTER AND FLEISSNER
A E
i-E
TS
--v
dirner
!gp9O} C
qp7O- *_
1 e
e
31-IC
FIG. 5. AKR Coomassie blue
alyzedbySDS-1 afterthefollowi
(B)virions treat linked with IOC
treated with 0.19 100pg ofDFDJ with
100,pgofD
dimer" is an ingrates with pur
nonreducingcoi not observed a monomericLE2
3 C D E Cross-linking of FeLV. The studies
de-scribed above have been performed with the
_
, -t-~ mAKR strain of
MuLV;
similar results have been,,
-~ obtained with other MuLV strains, including
-
-w-x
~~Rauscher
andMoloney
virions(data
notshown).
To determine whether these structures arepres-ent in retroviruses of otherspeciesaswell, the
DSP
cross-linking
experiments were repeatedwitharepresentative strain of FeLV. Upon
SDS-PAGEanalysis of '4C-amino acid-labeled FeLV,
the overallpattern obtained was similar to that
observed with AKRMuLV,althoughthe
abso-lute mobilities of the variousproteinswere
dif-ferent. As was the case forMuLV, analysisunder
nonreducing
conditions demonstrated thepres-enceof free gp7O(Fig. 6B andC),whereas NEM
treatment resulted in the formation of a
disul-fide-linked gp7O-pl5(E) complex (Fig. 6D).
Upon treatment with increasing amounts of
DSP, the intensity of the [gp9O] band
dimin-ished, as did that of the core components. The
onlynewcomponentresolvedwas alarge band
withsimilarmobilitytothat of the cross-linked
virions cross-linked with DFDNB. MuLV envelope complex (Fig. 6G).
Cross-link-staining pattern of AKR virions an- ing of
[3H]glucosamine-labeled
FeLV resultedinPAGE undernonreducing conditions, the appearance of a band of identical
mobility,
[ng treatments:(A) untreatedvirions; * * *
tdwith 0.1%NEM; (C) virions cross-
proving
thatthls
does factrepresent thecross-)pg of DFDNB per ml; (D) virions linkedFeLV envelope
complex
(Fig. 6J and K).%0
NEMfollowedby cross-linking with Thus, it appears that a similar envelope complexNBper ml; (E) virions cross-linked exists for FeLV as for
MuLV,
suggesting
that'SP
per mL (The bandlabeled "LETS thismay be auniversal feature ofmammalian%purity found in virions which comi- leukemia virus structure. Cross-linking of the
ifiedLETSprotein analyzed under FeLV p27 apparently occurred, although
inter-rtditions.Uponreduction, this band is mediate-sized oligomers were not detected.
This
nd a band which
comigrates
with may be a reflection of the differences in therS proteinappears)
14C-aa
A B C D E
x
-
[gP9o]-gp7O- fl
p
27-_e a a
se
m
e
pl5(E)-FIG. 6. Cross-linking of FeLV proteins withDSP. "C-amino acid-labeled (A-G) and [3H]glucosamine-labeled(H-K)FeLVanalyzed bySDS-PAGE under reducing (A)and nonreducing(B-K)conditions afterthe
followingtreatments:(A and B) untreated virus; (C)+1%iodoacetamide; (D) +0.1% NEM;(E) +0.1%NEM, +DSP(4 pg/ml); (F) +0.1% NEM, +DSP(20 pg/ml); (G) +0.1% NEM, +DSP(100pg/ml); (H)+0.1%NEM; (I)
+0.1%NEM, +DSP (10 pg/ml); (J) +0.1%NEM, +DSP (100pg/ml);(K) +0.1% NEM, +DSP(1,000pg/ml).
F G
a
3H-gln
H J K
SW
dik-J. VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:6.501.63.249.56.304.2] [image:6.501.128.408.442.625.2]CROSS-LINKED COMPLEXES OF MuLV AND FeLV 163
primary structures ofp27and p30, or may
ac-tuallyindicate a difference between the
struc-turalarrangementof the majorcoreproteins of
the twoviruses.
DISCUSSION
The cross-linking experiments described in
this paper demonstrate thepresence of distinct
associations between leukemia virus envelope
components and between viral core proteins.
Treatment ofMuLVwith thereversible
cross-linking reagent, DSP, results in the formation of alarge complex (component "x") which is
gly-cosylated (Fig. 2) and can be
immunoprecipi-tated by both anti-gp70 and anti-pl5(E) sera.
SDS-PAGE oftheimmunoprecipitatedcomplex after reduction demonstrates that the only
com-ponents present inappreciableamountsaregp7O
andp15(E) (Fig.3).Thiscomplexisformed both
withnativevirionsand with virionswhich have
been treated with NEM, and which therefore
contain the gp70-p15(E) disulfide-linked
com-plex. The acylationrate ofproteins byDSP is
veryrapid, with ahalf-lifeof less than 1 min at
neutral pH (19); thus it doesnot appearlikely
that the formationof"x" requireslateral
diffu-sionofcomponents in theviralmembrane. The
envelope complex is observed over a greater than 100-fold concentration range of DSP; in
addition, an apparently identical complex is
formed aftertreatment of virions with a second
cross-linking reagent, DFDNB, which has a
shorterspan (0.5nmcompared with1.1nmfor
DSP)anddifferent functionalgroups thanDSP (Fig. 5). These results indicate that gp7O and
p15(E) exist in theviralmembrane in theform
of a specific structural complex, which
corre-sponds to the component "x" observed in gels after stabilizationwith thecross-linkingagents.
The fact that theenvelope complexsurvives in
the presence of NP-40 (Fig. 4)is a further
dem-onstrationof the
stability
of thecomplex.Theapparent sizeof theenvelope complexas
determined by SDS-PAGE, usingtheLETS
di-mer asa molecular-weight marker, is
approxi-mately450,000to
500,000
(Fig. 5). However,thecross-linked structure ofthe complex could
re-sult in an anomalousmigrationrate. It has
re-centlybeenreported thatcross-linked
proteins
with molecular weights larger than 50,000
mi-grate in SDS-Laemmli gels asifthey were 17 to
25%largerthan themolecular weights calculated
fromtheirpolypeptide composition (21). By
us-ing the larger value as the limit of error, a
minimumi
molecular weight of 360,000 can becalculated for thecross-linkedcomplex,
suggest-ing that it is composed of at least four
gp7O-p15(E) components; by calculating a molecular
weight directly from the observed mobility of
the complex, a maximum value of six subunit
componentsisobtained. The demonstrationthat
asimilar complex ispresent on FeLVindicates
that this structure is a conserved feature of
retroviruses. Since the size of the complex is
large enough to account for the 8- to 10-nm
projectionsobservedonthe surface ofMuLV in
the electron microscope, it is attractive to
hy-pothesize that this complexactuallycorresponds
totheindividualknobsofthe viralenvelope.
The majorcross-linked core components
de-tected after DSP treatment of intact MuLV
consistedofp30 oligomers. Thep30dimer,
tet-ramer, andhexamer wereresolvedand
charac-terizedbyimmunoprecipitation (Fig. 3) andby
two-dimensional gel electrophoresis, in which
thefirstdimension wasrununder nonreducing
conditions and the second dimension was run
after reduction (data not shown). Larger
com-plexes were also formed, but these were not
resolved in the gels and formed abroad band
nearthe top of the gel.
Tne
formationofonlytheeven-numberedoligomersindicates the
sta-bility ofthe p30 dimerandsuggests that it is a
basic subunit of the viral core. Ourobservation
that thep30 complexes are notcross-linked in
thepresence of NP-40 suggests that
hydropho-bic forces playa rolein the self-association of
p30 molecules; thisis consistent with the
dem-onstratedpresence ofhydrophobicsitesonp30
(20,27).It isinterestingtocompare theseresults
with those ofBurnette et al. (4), who
demon-strated thatpurified p30ofMoloney MuLV
self-associates into multimeric forms at increasing concentrations. At the highest concentration
tested,the observed molecularweightfor
p30
as determined by sedimentation velocity analysis wasapproximately120,000,andacomputer-sim-ulated analysis of the data suggested a
mono-mer-octomerequilibrium.Thesep30 complexes
mayberelatedtothe6-nmcorestructureswhich
have beenvisualized in theelectronmicroscope
(22).
Cross-linked complexes of the other major componentsofthe viralcore,
p15,
p12,andplO, were not detected under conditions whichre-sulted inextensivecross-linkingof theenvelope
components and ofp30.This was notdueto a
lack of accessible amino groups on these
pro-teins,since wehaveobserved thataradioactive
analog of DSP, N-succinimidyl
[2,3-3H]pro-prionate(Amersham Corp.),caneffectivelylabel
all of the core proteins (data not shown). In
addition, after treatmentofvirions withDSPat
100
ltg/ml,
p15
can nolongerbeimmunoprecip-itated with anti-15 sera, indicating that it is
heavilysubstituted with DSP molecules.Alack
ofcross-linking isnotconclusive evidencethat
molecularassociations donotexist,since it may
VOL. 30, 1979
on November 10, 2019 by guest
http://jvi.asm.org/
merely indicate that the necessary functional groups are not arranged in the proper spatial
orientation to be cross-linked by a particular
reagent. Nonetheless, the fact that the smaller
core components are not readily cross-linked
may bean indication thatthey donotexist as
specific multiprotein complexes in virions.
Structural studies of otherenveloped viruses,
performed with the sameorrelated cross-linking
reagents, have demonstrated the existence of
heterocomplexes between virion core and
enve-lope components. Treatment ofSemliki Forest
viruswithdimethyl suberimidate resulted in the
cross-linking of viral envelope components to
cores (11). Cross-linking of vesicular stomatitis
virus withDSP and related reagents resulted in
thedetection of complexesformed between the
viral glycoprotein, G, and the membrane protein,
M(6), and also betweenthe Gprotein and the
nucleocapsid protein (21). Such
heterocom-plexes between envelope and core components
of retroviruses were not detected in ourstudy.
In particular it is of interest that cross-linked
complexes of p15 and theenvelopeproteins were
notobtained(Fig.3B), since p15 has been shown
topossesspropertiesofamembrane-associated
protein (2).Although failure todetect such
in-teractionsbythis type ofastudyis not
conclu-sive, it does indicate an increased probability
that such interactions are not present. It has
beendemonstrated that the envelope and core
components of MuLV aresynthesizedvia
sepa-rateclasses of mRNA's (29); thus they must find
their waytotheviralassembly site,theplasma
membrane, byindependent routes. Since
retro-viralproteins are relatively minorcomponents
ofinfectedcells,there mustbe anefficient and
specific interaction between the viral envelope
andcoreproteinswhich results in their
associa-tion in the presence ofalargeexcessofcellular
material. Recently it has been shown that the
assembly of budding virus particles can occur
with uncleaved core precursor Pr7O (30). It is
interestingto speculate that the association of
viral core and envelope components at the
plasma membrane requiresthe uncleaved core
precursor and that, in extracellular particles
which contain the cleaved core proteins, these
interactions are not retained. A possible
ap-proach toward resolving this question would be
tocross-linkbudding virus particles and to look
for thepresence ofenvelope-core complexes in
either solubilized membranes or subsequently
released virusparticles.
ACKNOWLEDGMEENTS
This workwas supported by grants CA-02214 from the National Cancer Institute and VC-222 from the American CancerSociety.
We thankJudyLieman-Hurwitz and Anne Swartout for providing excellent technical assistance.
LITERATURE CIMD
1. August,J.T., D. P. Bolognesi,E.Fleissner, R. V. Gilden, and R. C. Nowinski. 1974. A proposed no-menclature for the virionproteinsofoncogenicRNA viruses.Virology 60:595-601.
2. Barbacid, M., and S. A. Aaronson.1978. Me&brane propertiesof the gaggene-coded p15 proteinofmouse
type-C RNA tumor viruses. J. Biol. Chem. 253:1408-1414.
3. Bonner,W.M.,and R. A.Laskey.1974.A filmdetection methodfortritium-labeledproteinsandnucleic acids in polyacrylamide gels. Eur.J. Biochem.46:83-88. 4. Burnette, W.N., L.A.Holladay,and W. M. Mitchell.
1976.PhysicalandchemicalpropertiesofMoloney mu-rine leukemiavirusp30protein:amajorcorestructural componentexhibiting highhelicityand self-association. J. Mol Biol. 107:131-143.
5. Demsey,A., D.Kawka,and C. W.Stackpole. 1977. Applicationoffreeze-dryingintact cellstostudies of murine oncornavirusmorphogenesis. J.Virol.
21:358-365.
6. Dubovi,E.J., andR.R.Wagner.1977.Spatial relation-ships oftheproteinsofvesicularstomatitis virus: in-ductionof reversible oligomers by cleavable protein cross-linkers andoxidation. J.Virol.22:500-509. 7. Eisenman,R.N., and V. M.Vogt. 1978. The
biosyn-thesis ofoncovirusproteins. Biochim. Biophys. Acta 473:187-239.
8. Famulari, N.G.,D.L.Buchhagen, H.D.Klenk, and E.Fleissner.1976.Presenceof murine leukemia virus envelope proteinsgp7O and p15(E)in a common poly-protein ofinfected cells.J.Virol.20:501-508. 9. Frank,H.,H.Schwarz,T.Graf,and W.Schafer. 1978.
Properties ofmouseleukemia viruses. XV. Electron microscopicstudies on theorganizationofFriend leu-kemiavirusand othermammalian C-type viruses.Z. Naturforsch.Teil C33:124-138.
10. Gahmberg, C. G., and S. Hakomori. 1973.External labellingofcellsurfacegalactoseandgalactosaminein glycolipidandglycoprotein of human erythrocytes. J. Biol. Chem. 243:4311 4317.
11. Garoff, H.,and K.Simons.1974.Locationofthespike glycoproteins in theSemliki Forestvirusmembrane. Proc.Natl. Acad.Sci.U.S.A.71:3988-3992.
12.Helenius, A., and K. Simons. 1972. The binding of detergents to lipophilic and hydrophilic proteins. J. Biol.Chem.247:3656-3661.
13. Ikeda, H.,W.Hardy,E.Tress,and E. Fleissner. 1975. Chromatographicseparationandantigenic analysisof proteins of theoncornaviruses.V. Identification ofa
newmurineviral protein,p15(E).J.Virol.16:53-61. 14.Jamjoom,G.A.,R. B.Naso,and R. B.Arlinghaus.
1977.Further characterization of intracellular
polypro-teinprecursorsofRauscher leukemiavirus. Virology
78:11-34.
15. Karshin,W.L,L J.Arcement,R. B.Naso,and R. B. Arlinghaus. 1977. Common precursor for Rauscher leukemia virus gp69/71,p15(E),andpl2E.J.Virol.23: 787-798.
16.Keski-Oja, J.,D. F. Mosher, andA. Vaheri. 1977.
Dimeric character offibronectin,amajorcell surface-associatedglycoprotein. Biochem.Biophys.Res. Com-mun. 74:699-706.
17. Kessler, S. W.1975.Rapid isolation of antigens fromcells witha staphylococcal protein A-antibodyadsorbent: parameters of the interaction ofantibody-antigen com-plexeswithproteinA.J. Immunol.115:1617-1624. 18. Laemmli, U. K. 1970. Cleavage ofstructural proteins
duringtheassembly ofthehead ofbacteriophageT4.
on November 10, 2019 by guest
http://jvi.asm.org/
Nature(London) 227:680-685.
19.Lomant, A. J., and G. Fairbanks. 1976. Chemical probes of extended biologicalstructures:synthesis and properties of the cleavable protein cross-linkingreagent
[nS]-dithiobis(succinimidyl proprionate). J. Mol. Biol. 104:243-261.
20. Marcus,S. L, S. W. Smith, J.Racevskis, and N. H. Sarkar. 1978. The relativehydrophobicity of
oncorna-viral structural proteins.Virology 86:398-412. 21. Mudd, J. A., and R. E. Swanson. 1978. In situ
cross-linking of vesicular stomatitis virus proteins with
re-versibleagents.Virology 88:263-280.
22.Nermut, M. V., H. Frank, and W. Schafer. 1972. Prop-erties ofmouseleukemia viruses.m.Electron
micro-scopicappearance asrevealed afterconventional
prep-arationtechniquesaswellasfrfeez-drying and
freeze-etching. Virology 49:345-358.
23. Pinter, A., and E. Fleissner. 1977. The presence of
disulfide-linked gp7O-p15(E) complexes in AKR MuLV. Virology 83:417422.
24. Pinter, A., J. Lieman-Hurwitz, and E. Fleissner. 1978. Thenatureofthe association between themurine leu-kemia virusenvelope proteins. Virology 91:149-158. 25. Ruoslahti, E., and A. Vaheri. 1975. Interaction of
solu-ble fibroblast surfaceantigen with fibrinogen and fibrin. Identity with cold insoluble globulin of human plasma. J. Exp. Med. 141:497-501.
26.Shapiro, S. A., M. Strand, and J. T. August. 1976. High molecular weightprecursorpolypeptidesto
struc-turalproteins of Rauschermurine leukemia virus. J. Mol.Biol. 107:459-477.
27.Swanson, S. K., E. Sulkowski, and K. F. Manly.1978.
Hydrophobic binding site(s) onMoloney-murine
leu-kemia virus p30.Virology 85:211-221.
28.VanZaane, D., M. J. A. Dekker-Michielson, and H. P. J. Bloemers.1976.Virus-specificprecursor
polypep-tides incells infectedwith Rauscher leukemia virus: synthesis, identification, and processing. Virology 75: 113-129.
29.Van Zaane, D., A.L. J. Gielkins, W. G. Hesselink,
and H. J. Bloemers. 1977. Identification of Rauscher murine leukemiavirus-specific mRNAs for the synthe-sisofgag-andenv-geneproducts. Proc. Natl. Acad. Sci. U.S.A. 74:1855-1859.
30.Witte,0.N., and D. Baltimore. 1978. Relationship of
retrovirus polyprotein cleavagestovirion maturation studied withtemperature-sensitive murine leukemia
vi-rusmutants.J. Virol. 26:750-761.
31.Yoshinaka, Y., and R. B. Luftig.1977.Murine leukemia virus morphogenesis: cleavage of P70 in vitrocanbe accompanied byashift fromconcentrically coiled inter-nal strand("immature")toacollapsed ("mature") form of the viruscore.Proc.Natl. Acad. Sci. U.S.A.
74:3446-3450.
on November 10, 2019 by guest
http://jvi.asm.org/


![FIG. 6.following+0.1%labeled+DSP Cross-linking of FeLV proteins with DSP. "C-amino acid-labeled (A-G) and [3H]glucosamine- (H-K) FeLV analyzed by SDS-PAGE under reducing (A) and nonreducing (B-K) conditions after the treatments: (A and B) untreated virus; (C) +1% iodoacetamide; (D) +0.1% NEM; (E) +0.1% NEM, (4 pg/ml); (F) +0.1% NEM, +DSP (20 pg/ml); (G) +0.1% NEM, +DSP (100 pg/ml); (H) +0.1% NEM; (I) NEM, +DSP (10 pg/ml); (J) +0.1% NEM, +DSP (100 pg/ml); (K) +0.1% NEM, +DSP (1,000 pg/ml).](https://thumb-us.123doks.com/thumbv2/123dok_us/1514851.104100/6.501.63.249.56.304/following-proteins-glucosamine-nonreducing-conditions-treatments-untreated-iodoacetamide.webp)