CopyrightC1976 AmericanSociety forMicrobiology Printed in U.SA.
Fractionation of DNA
Nucleotide Transcripts from Moloney
Sarcoma Virus and Isolation of Sarcoma Virus-Specific
Complementary DNA
ARTHUR E. FRANKEL,* RICHARD L. NEUBAUER, AND PETERJ. FISCHINGER Laboratory ofViral Carcinogenesis, National Cancer Institute, Bethesda, Maryland 20014
Received forpublication10December1975
Radioactive
DNAcomplementary to nucleotide sequences inMoloney
murinesarcoma virus
(MSV) and
Moloneyleukemia
virus(M-MuLV) complex
wasmade
by the endogenous reverse transcriptase reaction. These virus stocks contained a threefold excess ofMSV
over M-MuLV as measured by biological assay. The complementary DNA was an accurate copyof the
viral RNA
inthat
86%
of35S
viral RNAhybridized
withcomplementary
(cDNA) DNA
at a 1.5 to1cDNA-RNA molar
ratio.The
complementary DNA, of
a4-6Ssize,
wasfraction-ated by sequential absorptions
with MuLVand the feline leukemia
viruspseu-dotype of MSV,
[MSV(FeLV)]
RNA.In
this
mannerthree
setsof nucleotide
sequences
which
representdifferent portions of the MSV viral
complex
wereobtained:
a sarcomavirus-specific fraction
(cDNAa,)
with
sequencesthat had
no
homology
toM-MuLV
RNA but which
hybridized
toMSV(FeLV)
RNA,
asarcoma-leukemia
fraction
(cDNAcommon)
with
sequences common toMSV
aswell
as M-MuLVviral RNA, and
acDNAIeuk
representingthose
nucleotide
sequences
found
only
inM-MuLV.
Hybridization of
MSV-MuLV viral 35S
RNAwith
athreefold
molar excessof
cDNA's revealed that
-20% washybridized
with
cDNAsarc,
whereas -75% was hybridized with cDNAommon. M-MuLV35S RNAalone did
nothybridize with
cDNA,
but did
hybridize
40and
50%with
cDNAleuk
and
cDNAcOmmon,
respectively. The
cDNAsac
representsabout
25%of
the total MSV
sequences,whereas the
cDNAcommon
representsthe remainder of
the
MSV virus genome.Some
cDNA ommon
sequences wereshared
by
twoother
sarcomaviruses
and several
distinctly different isolates of MuLV. In
contrast,the MSV "sarc"
sequenceshad
little
or nohomology with
twoother murine
sarcoma virus
isolates.
Sarcomagenic
viruses isolated from variousmammalian
speciesgenerally transform
fibro-blasts
inculture and
coexist withleukemia
helper
viruses,which furnish the
virusenve-lope
and possibily supply the
reversetranscrip-taseenzyme (10, 11, 15, 24, 27; P.
T.
Peebles,
B.I. Gerwin, and
E. M. Scolnick, Virology, inpress).
The
morethoroughly studied
murine sarcoma viruses(MSV)
weregenerally
shownto
be defective for
replication
but not defectivefor
transformation.
Accordingly,
cellstrans-formed
only by
MSV
inthe absence ofreplicat-ing
leukemia
virus wereisolated
(1, 2).Varioustypes of
nonproducing
cells of several specieswere
characterized,
someof
whichexpressed
some murine
luekemia
virus(MuLV) antigenic
determinants
and others which
didnotexpress these functions (1, 2, 13).MSV-transformed,
nonproducing
cells of variousspecies
stably
transcribe
someMuLV-specific
RNAwhether
the cells contain MuLV
antigens
or not(20).
Thus
MuLV-specific
information seems tobeagenetically stable
partof
the MSV
genome.The
nucleotide
sequencesof
MSV associated with
transformation have
notbeen clearly
identi-fied. Some
strainsof MSV derived after
passagethrough
rats contain as apartof their
genomerat and/or rat leukemia virus sequences (22,
23).
Recently,
inthe avian oncornavirus systemthe availability
ofnondefective transforming
Rous
sarcoma virusand
itscorresponding
transformation-defective deletion
variantsal-lowed the identification
of Rous sarcoma virussequences
associated with transformation
(D.Stehelin
etal.,
J.Mol. Biol.,
in press).Although
thenature of mammaliansarcoma
viruses
does
not allow this approach, analter-nate
method
wasderived. The Moloney isolateof MSV
(M-MSV),
notknown
to have beenpassed through
rats, could give rise to murinesarcoma-positive,
leukemia-negative (S+L-)
cells,
which when infected with MuLV gaverise to virus stocks in which infectious MSV was in excess over MuLV (2, 18). This MSV
481
on November 10, 2019 by guest
http://jvi.asm.org/
482 FRANKEL, NEUBAUER, AND FISCHINGER
excesspersisted when MSV was carried overby transspecies rescue into the cat system (16). M-MSV was then
propagated
forseveral years in catcells as apseudotype
virus withonly felineleukemia virus
(FeLV)
as helper[MSV(FeLV)1.
It wouldseemreasonable toas-sumethatonly
M-MSV-specific
sequenceswereconserved in cat cells that do not contain M-MuLV sequences in uninfected cat cell DNA (A. Frankel et al., inpress).
Accordingly,
sin-gle-stranded DNA could be madefrom the M-MSV-MuLV complex by the endogenousre-verse transcriptase reaction, and the MuLV-specific sequences could be removedby
exhaus-tive
hybridization
with MuLV RNA. Theresid-ual
complementary
DNA(cDNA)
could behy-bridized to
MSV(FeLV) pseudotype
RNA toverify the presence of
sarcoma-specific
se-quences. After several
sequential cycles
ofhy-bridization,
we isolatedsequences,
termed"sarc," only associatedwith M-MSV. Weshow that the remainder of
M-MSV-specific
DNA shares itssequences with M-MuLV. Hybridiza-tion experiments of MSV viral35S
RNA withisolated
sarcoma-specific
cDNA showed thatapproximately
25%of the M-MSVgenomecon-sists of
sarcoma-specific sequences,
whereas theremainder
is shared with MuLV. TheseM-MSV
sarcoma-specific
sequences were notfound
inKirsten
and
Harvey isolates
of MSV.MATERIALS AND METHODS
Viruses. The murine sarcoma-leukemia virus mixtureused to make probe wasobtained from the 319 S+L- clone ofMSV-transformed 3T3FL cells infected with the IC isolate of M-MuLV (2, 7). The virushad athree- to fourfold excess of focus-forming sarcoma virus toleukemia focus-inducing virus by
biological assays. The viral supernatant was con-centrated 2,000-foldbypolyethyeleneglycol precipi-tation (8%, wt/vol), banded through a sucrose gra-dient (20 to 60% sucrose in phosphate-buffered sa-line), pelleted through phosphate-buffered saline, andsuspendedinphosphate-buffered saline prior to preparingcDNA.
MuLV was obtained from the supernatant of 3T3FLcells infected with the IC isolate of M-MuLV. The supernatantwasconcentratedby polyethylene glycol precipitation 2,000-fold andsuspendedin 0.15 M sodiumchloride, 0.015 MTris, 0.0015 M EDTA, pH7.4(NTE) for RNA extractionorbandedthrough sucroseandsuspendedinphosphate-buffered saline ifusedtomake cDNA.
TheMSV(FeLV) virus complexwasgeneratedby rescue from the FG10 clone of S+L- cells into cat embryo fibroblasts with FeLV and grown in the P521 producer clone of feline CCC cells (8, 16, 17, 19). The P521 viruscomplex hadan MSVtoFeLV ratio of3:1 bybiological titrations. Thevirus com-plex was purified from supernatant fluid by banding in a continuous-flow ultracentrifuge
(Electro-Nu-cleonicsLaboratories, Inc.). Thevirus waspelleted
and dissolved in NTE prior to RNA extraction. FeLVvirus,Rickard strain(422),wasobtained from
Electro-Nucleonics, Inc. and dissolved in NTE for RNA extraction. Kirsten MSV-MuLV
complex
grown in NRKcells,
Rauscher MuLV, Kirsten MuLV virus, and BALB-2 xenotropic MuLV were also obtained from Electro-Nucleonics, Inc. The FeLVpseudotypeofHarveyMSVwasobtainedfrom P. Sarmaandwasgrownincatembryofibroblasts,
and the virus was purified identically toMSV(FeLV).
cDNA's. Radioactively labeled DNA sequences
complementary to MSV-MuLV viral RNA were madebytheendogenous reverse transcriptase reac-tion (26). The reaction mixture contained 1 ml of banded MSV-MuLV virusinphosphate-buffered sa-line, 1mlof0.5 MTris-hydrochloride (pH 7.8), 1ml of5mMMnCl2, 1ml of0.1Mdithiothreitol, 1mlof
actinomycinD (1,000
jig/ml;
Calbiochem), 0.1mlof 2 x 10-2MdATP, dGTP, and dCTP, 0.9 ml of0.1 M NaF,1mlof 0.5MNaCl,1mlof0.15%TritonX-100, 1ml of[3H]TTP(5x10-5M;1mCi/ml;NewEngland
Nuclear Corp.), and 1 mlof0.1M CMP. The final concentrationswere0.1mgof viral protein perml,0.05 M Tris, 0.5 mM MnCl2, 0.01 Mdithiothreitol,
100
jig
ofactinomycin D perml,2 x 10-4 Mbases,9 mM NaF, 0.05 M NaCl, 0.01%Triton, 5 x 10-6M [3H]TTP, and 0.01 M CMP. The reaction mixture was vortexed and incubated for 45 min at 37C. It wasthentreated with0.7%sodium dodecylsulfate, 0.01 M EDTA and shaken. The mixture was ex-tracted with an equal volume of NTE-saturated phenol-chloroform-isoamyl alcohol (1:1:0.04), and thewaterphasewasmade0.5NNaOH and boiled for10min. Thissolutionwasdiluted with6volumes ofdistilled water, neutralized with 2 N HCl, and concentratedona1-cmhydroxylapatite column. The cDNA was eluted from the column with 0.5 ml of 0.3 Mphosphate buffer (PB) washes. Testing the enzy-matic product prior to boiling in alkali revealed <10% double-stranded DNA by salt elutions from hydroxylapatite (6). The DNA had a specific activity of 2 x 107 counts/min per,ug.RNA. 65S RNA was prepared from fresh MSV-MuLV virus suspended in sterile NTE. The suspen-sion was treated with 1% sodium dodecyl sulfate, extracted twice with equal volumes of NTE-satu-rated phenol, and precipitated with 2 volumes of -20C dehydrated ethanol. Diethyloxydiformate (Eastman) (0.1%) was added prior to precipitation to inhibit nucleases (24). The RNA was left over-night at -20 C and thencentrifuged at 10,000 x g for 30 min. Theviral RNA pellet wasresuspendedin 0.5ml ofNTEand layered on a 10 to 30% sucrose in NTEgradient andcentrifuged at 40,000 rpm for 3 h in anSW41 rotor.Opticaldensities at 260 nm on 0.5-mlfractions were read, and the 60-70S absorbance peakwaspooled and precipitated with 2 volumes of ethanol in apolyallomer tube. After 6 h the high-molecular-weight RNA was pelleted and resus-pendedin 0.5 mlof NTE or 0.12MPB.30-40S RNA wasprepared from 65S RNA by heating for 1 min in NTEat 100C andcentrifuging at 40,000 rpmina10 to 30%sucrosegradientinNTEfor 5h inan SW41 J. VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
rotor. Fractions were collected, and the30-40S ab-sorbance peak was pooled and precipitated with 2 volumes of ethanol. After 6 h the 30-40S RNA was resuspended in 20Mulof distilled water for iodination or 400 MAl of NTE for hybridization. Approximately 30
jig
of high-molecular-weight RNA could be ob-tained from 2 liters of MSV-MuLV supernatant or 15 liters of MuLV supernatant.ForMSV(FeLV)-FeLV RNA the viral concentrate was extracted once with 1%sodium dodecyl sulfate and twice with phenol and precipitated with ethanol at -20 C. The RNA was pelleted at 10,000xg for 30 min and resuspended in 0.5 ml of NTE or 0.12 M PB. High-molecular-weight viral RNA from MSV-(FeLV)-FeLV RNA was not isolated because the virus was grown in cat cells. Contaminating cat cellular RNAdid not hybridize with the mouse viral sequences (Frankel et al., in press).
Total viral RNA of Kirsten sarcoma-leukemia vi-rus, Kirsten-MuLV, Rauscher leukemia-MuLV, and Harvey sarcoma-FeLV was extracted in the same way.
High-molecular-weight RNA was iodinated by a modification of the technique of Colcher et al. (5). A
2.5-Mlamount of New EnglandNuclear Corp. 125I (1 mCi) was mixed with 3Mlof 0.3 mM NaSO3, 0.3 N H2SO4, and 10 ul of distilled water and incubated for 5 min at room temperature. Then 2Mulof viral RNA in distilled water (1 ug) was added along with 1
M1A
of 1.8 MNaOAc, pH 4.5, and 1 Mlof 50 mM thallium trichloride. This mixture (19 Mig total) was incu-bated for 30 min at 65 C. Then 1.2Mulof 0.1 M NaSO3 wasadded, and the mixture was reincubated for 10 min at 65C. The material was then chilled and passed through a 10-cm Sephadex A-50 column to removefree iodine and, when necessary, separatedfrom iodinated protein by a CF-1l column. '25I-la-beled MSV-MuLV 30-40S RNAwaspreparedfrom unlabeled 30-40S viral RNA. The prepared cDNA
hybridizedwith85%oftheiodinated RNA testedby
RNaseT.digestionataDNA-RNA ratioof1:1and 96%at aDNA-RNAmolar ratioof12:1(see Table1). Fractionationof cDNA's. cDNA (0.25 to 0.75Mug
or 5 to 15 million counts/min) was prepared from MSV-MuLVvirus and suspendedin 1 mlof 0.3 M PB. This cDNA wasdiluted to 0.22MPB,and6 to 8
M(gof70S MuLV RNAwasadded. Thishybridization
mixture wasincubated at 63 C for 48 h. It wasthen diluted1:1withdistilled water and loadedon a 1-cm
hydroxylapatitecolumn at 50 C. Washes of0.14and 0.3M PBweredone.Aliquotswereprecipitatedwith trichloroaceticacid, filtered onto0.45-,umcellulose acetatefilters,and counted. The unhybridized 0.14 M PBwash material wasdiluted fivefold with dis-tilled water and concentratedon a 1-cm
hydroxylap-atite column.The cDNAfractionwaselutedwith 0.3 M PB 0.5-ml washes. Again aliquots were taken,
and acidwasprecipitated, filtered,and counted. The hybridizing fractionwastreated with 0.5 MNaOH and boiled for 10 min to remove the RNA. It was then diluted with distilled waterfivefold, neutral-ized, and concentratedastheunhybridizedfraction. With thisproceduretwocDNAfractions,onehaving
MuLV-related sequences and one without MuLV sequences, weremade.
Then 1 ml of each cDNA fraction in 0.3 M PB was mixed with 40 ,Mg of MSV(FeLV)-FeLV RNA and diluted to 0.22 M PB. These were again incubated for 48 h at 63C. The hybridization mixtures were diluted 1:1 with distilled water and loaded on 1-cm hydroxylapatite columns at 50 C. This fractionation step, carried out as before, further segregated the MuLVsequences into those common with MSV and those that were MuLV specific. Further, this step selected only those sequences not hybridizing to leu-kemia that can still hybridize to MSV, the "sar-coma-specific" sequences. Nonhybridizing se-quences and those representing contaminating mouse cellular RNA are selected against at this fractionation and are discarded. It was found neces-sary to repeatthe fractionation of the sarcoma-spe-cific cDNA with MuLV RNA because the probe con-tinued to display residual hybridization to MuLV RNA. After this second fractionation with MuLV the sarcoma-specific or
cDNAsa.
material had very little hybridization with MuLV RNA. Further frac-tionations with MuLV did not yield any further significant hybridization of thecDNAsar,.
We were able to obtain from 15 million counts/min of MSV-MuLVcDNA approximately 1 million counts/min of cDNAcommon-materialhybridizing to leukemia and MSV(FeLV) RNA; 500,000 counts/min of cDNAIeuk -material hybridizing to leukemia, but not sarcoma, MSV(FeLV); and 200,000 counts/min of cDNAsare-material hybridizing to sarcoma but not leukemia. There was about 60% recovery of DNA at each frac-tionation.The losses occurred at boiling and concen-trating stepsand not during incubations for hybridi-zation.Characterization of cDNA fractions. At each stageof fractionation 300to1,000 counts/min of each cDNA fractionwas mixed with 2 Mgof65S MSV-MuLV RNA, 0.7 Mig of70S MuLV RNA, 4 Mig of MSV(FeLV)RNA,or noRNA in0.1mlof0.22M PB andincubated for 20 hat63C. The mixtureswere then diluted into3mlof 0.12M PBand loadedontoa 1-cm hydroxylapatite column at 50C. PB washes (0.14 Mand 0.3M)werepassed throughthecolumn,
andacid-precipitablecounts inthe washeswere de-termined. The ratio of0.3M PBcounts/mintototal counts/min represents the fractionof cDNA
hybrid-ized to RNA.
The sizeof the cDNA fractionwasdeterminedby
centrifuging200 to 1,000 counts/minof each cDNA fraction on a 10 to 30% (wt/vol)sucrosegradientat 34,000 rpm in an SW56 rotor for 18 h at 4C and
determining acid-precipitable counts per minute and optical densities on fractions. An Escherichia coli tRNA markerwascentrifuged
concurrently.
The purity of the DNA transcripts was ascer-tained by determining
Crtlu2
for hybridization of 1,000- to 10,000-counts/min probewith 2 ug of 658 MSV-MuLVRNA in 0.1 mlof0.22 M PBandtakingaliquotsat various times. Theexperiment was
re-peated with 0.8
Mug
of 30-40S MSV-MuLV RNA. Percentage ofhybridization of thealiquotswas de-termined byhydroxylapatitechromatographyas be-fore. Crt curvesweremadeas suggested by Leongand Britten(3, 4, 14).Hybridsof eachprobefraction withMSV-MuLV RNA were dilutedin 0.12 M PB
on November 10, 2019 by guest
http://jvi.asm.org/
484 FRANKEL, NEUBAUER, AND FISCHINGER and loaded on hydroxylapatite columns at 50C. Theseelutedat5-Cintervals with0.12 MPB washes
toobtainthermal denaturation profiles.
Hybridization of viralRNA with cDNAfractions. The specificity of the cDNA fractionswasstudiedby hybridizing cDNA's withhigh-molecular-weight, io-dinatedviral RNAatlowand highDNA-RNA molar ratios.cDNAwasdialyzed against NTE, co-precipi-tated in 70% ethanol with 0.1 M sodium acetate buffer andtRNA, and resuspended in 5 to 40 pLI of 0.3 M PB.cDNA (10,000to 30,000counts/min) in 1
,ul
wasmixed with1 .ulof 1.2 M PB and 1,1
of iodi-natedviral RNAcontaining 1,000to 10,000counts/ min of RNA. The hybridization mixture (0.75 M Na+)wasincubatedinsmallEppendorfpolyallomer tubesat68CuntilobtainingaCotof 0.3. The solu-tionwasthen dissolved in 2 ml of 2x SSC andsplit into 2equal volumes.One fractionwastreated with RNaseT, (200U/ml),and both fractionswere incu-batedfor 30minat37C. The nucleicacidswerethen precipitated with trichloroacetic acid, filtered, and counted inbothagammacounter andaliquid scin-tillationcounterwithEconoflour. The acid-precipi-tablegammacounts perminute afterRNase treat-ment divided by the untreated acid-precipitable gammacountsperminuterepresentsthepercentage ofhybridization. Thetritium countsperminute di-vided bytwotimes the gamma counts perminuterepresentsthe cDNA-RNA molarratio(basedon2x 107counts/minper,tgfor cDNA and107counts/min per,ugforiodinatedRNA). Hybridizationswere
per-formed atboth 10to12:1 and 1 to 3:1 molar ratios of cDNA-RNA.
Distribution of Moloney sarcomaand leukemia viral sequences. Each probe fraction (500 to 1,000 counts/min) was hybridized with total viral RNA
extract from Kirsten MSV-MuLV, Harvey MSV(FeLV)-FeLV, Kirsten MuLV and Rauscher MuLV.Thehybridizationsweredonein 0.22 MPB
at63C toafinal Crt ofatleast10mol-s/liter. The extentofhybridizationwasanalyzed by hydroxylap-atitechromatography asbefore.
RESULTS
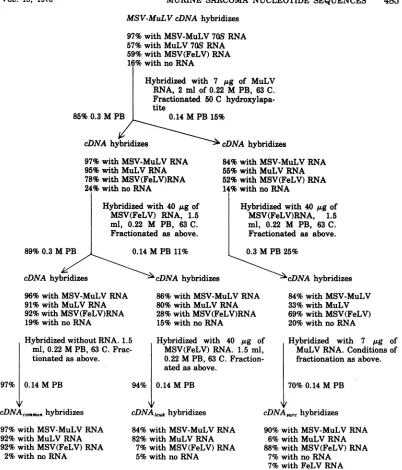
Fractionation scheme of MSV-MuLV
cDNA. Ten cDNA preparations from MSV-MuLVwereexamined inaseriesofsequential
absorptions with
high-molecular-weight
viral RNAstofractionate DNAsequences. The totalcDNA was a reasonably homogeneous
tran-scriptinthat70 to86%of homologous iodinated MSV-MuLV65S RNAwashybridizedtocDNA
at a 1.5:1 molar excess of probe DNAto viral 70S RNA at a
Cot
of 0.24. A fractionationschema is shown in Fig. 1. The diagramed MSV-MuLV cDNAhybridizedmost(97%)with
MSV-MuLV 65S RNA and less and
approxi-mately equally (57 to 59%)to MuLV70S RNA
andMSV(FeLV) RNA and hada16%
hybridi-zation withno RNA present.This background
was showntobe double-stranded DNAby salt
elutionsonhydroxylapatite (6).The cDNAwas
absorbed with MuLV 70S RNA to eliminate
MuLV-related
sequences. After fractionationwith MuLV
70S RNA the fractions show achange
inhybridization
pattern (Fig. 1). Thematerial hybridizing
to MuLV RNA had anenriched hybridization (95%) to MuLV RNA and a somewhat lower hybridization to
MSV(FeLV) RNA
(78%)and
24%with no RNA.The
material
nothybridizing
to MuLV RNAshows
areduction
in both MuLV andMSV(FeLV)
RNAhybridizations
(55 and 52%,respectively).
Furtherfractionation of
thecDNA fractions
withMSV(FeLV)
RNAyields
aset
of
sequences,cDNAcommon,
with
ability
tohybridize equally well
to MuLV orMSV(FeLV)RNA-92%
for each. This
fraction constitutes0.73 of the
original MSV-MuLV
cDNA.An-other
setof
sequences,cDNAleuk,
isable
tohy-bridize with
MuLV sequences (82%) but notwith
MSV(FeLV)
sequences (7%). ThiscDNAeuk represents 0.09 of the starting
mate-rial. The final
fraction,cDNAsarc,
represents0.06
of the
startingmaterial. The other
0.15of
the
original cDNA
isnon-hybridizable
mate-rial. With
different
preparationsof
cDNAsar.
the
ratioof
MSV(FeLV)hybridization
to MuLVhybridization
ranges from 4:1 to 15:1 (as in Fig.1). A
third hybridization
with MuLV RNA ulti-mately reduces thehybridization with MuLV tonear
the value of hybridization with
no RNApresent
for all "sarc" probes. The preparation
ofcDNAsar.
minimally
requiresboth
twoleuke-mia
fractionations and
oneMSV(FeLV)
frac-tionation.
The order of fractionation does
not make anydifference.
The requirementfor
twoleukemia fractionations
cannotbe reduced
by
increasing
the leukemia
RNA inthe
hybridiza-tionmixture, increasing
the
salt
concentrationof
the
hybridization
to 0.3 MPB,
lowering the
incubation temperature to 50 C, or
extending
the
timeof incubation
up to 72 h.The
cDNAeuk
preparation at times
required repeated
frac-tionations with
MSV(FeLV)
RNA toreduce
hy-bridization
toMSV(FeLV) RNA
toless than
10%. A
single fractionation enriches but does
not
lead
toabsolute exclusion of
sequences.It should be stressed that the terms cDNA
"sarc,"
"leuk,"
and "common" are operationalterms describing different sets of viral se-quences. These cDNA fractions do not necessar-ily representsingle specific genes; in the case of "common" and "leuk" these are almost cer-tainly composed of many genes.
Properties of isolated cDNA fractions. All cDNAfractions
yielded
broadpeaks
invelocity
density
gradients. The unfractionatedMSV-MuLV cDNA was
-4S.
ThecDNAcommon
was4S;
thecDNAleuk
was3S; and thecDNAsarc
wasattimes more heterogeneous but averaged
4S
to6S.
J. VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
MSV-MuLV cDNA hybridizes 97%withMSV-MuLV70S RNA 57%with MuLV 70S RNA 59%withMSV(FeLV) RNA 16% with no RNA
Hybridized with 7 ,ug of MuLV
RNA, 2 ml of 0.22 M PB, 63 C.
Fractionated 50C
hydroxylapa-tite
85% 0.3M PB 0.14 M PB15%
cDNA hybridizes cDNA hybridizes
97%withMSV-MuLV RNA 84% with MSV-MuLVRNA 95% withMuLV RNA 55%with MuLV RNA 78%withMSV(FeLV)RNA 52%with MSV(FeLV) RNA 24% withnoRNA 14%withnoRNA
Hybridized with 40,ug of Hybridized with 40 ,ug of MSV(FeLV) RNA, 1.5 MSV(FeLV)RNA, 1.5 ml, 0.22 M PB, 63 C. ml, 0.22 M PB, 63 C. Fractionated as above. Fractionated as above.
89% 0.3M PB 0.14 M PB 11% 0.3 M PB 25%
cDNAhybridizes cDNAhybridizes cDNAhybridizes
96% withMSV-MuLV RNA 91% with MuLV RNA 92%withMSV(FeLV)RNA 19%withnoRNA
Hybridized without RNA.1.5
ml,
0.22 MPB,
63C. Frac-tionatedasabove.97% 0.14 M PB 9
CDNAcommon hybridizes cj
86% with MSV-MuLV RNA 80% with MuLV RNA 28% withMSV(FeLV)RNA 15%withnoRNA
Hybridized with 40 ug of MSV(FeLV) RNA. 1.5 ml, 0.22 M
PB,
63C. Fraction-atedasabove.)4% 0.14M PB
wDNAl,.k
hybridizesc
84% withMSV-MuLV 33% withMuLV 69% withMSV(FeLV) 20% withnoRNA
Hybridized with 7 /ug of MuLV RNA. Conditions of fractionationasabove. 70% 0.14 MPB
,DNAiarc hybridizes 97%withMSV-MuLV RNA 84%with MSV-MuLV RNA 90%with MSV-MuLV RNA 92%with MuLVRNA 82%with MuLV RNA 6%with MuLV RNA 92%withMSV(FeLV) RNA 7%withMSV(FeLV) RNA 88%withMSV(FeLV) RNA
2%withnoRNA 5%withnoRNA 7%withnoRNA
7%withFeLV RNA FIG. 1. FractionationschemeofMSV-MuLV cDNA.
cDNA of MSV-MuLV hybridized to 65S
MSV-MuLV RNA witha
CrtI/2
of0.05 (Fig. 2).ThecDNAcommo,, fraction hybridized'ata simi-larrate, witha
CAtM2
of 0.07.ThecDNA,.,,
had.a
CAtM2
of 0.08. The cDNAlOUk hybridized moreslowly, with a
Crt112
of 0.26. The data show there are3.25 times as manyMSV-related se-quences in MSV-MuLV as MuLV-related se-quences. Furthermore, the complex cDNAprobably does not have a major contaminant
because the MuLV sequences and MSV
se-quencesaccountfor atleast 85% of the
hybridiz-able sequences. The observations concur with
biologicaldata whichdescribeathree- to four-fold excess ofinfectious MSV to MuLV. With
35S MSV-MuLV RNA
cDNA.,,
cDNAcommon,andMSV-MuLV cDNAhybridizedwitha
CrtAi2
of0.02; cDNAlOUk hybridizedwitha
CrtI/2
of0.07.Accordingly,noneofthe cDNA fractions
repre-sents a minor contaminant in the 65S or 35S
viralRNA.
Thermal denaturation ofhybrids. The hy-485
on November 10, 2019 by guest
http://jvi.asm.org/
[image:5.503.55.457.62.532.2]486 FRANKEL, NEUBAUER, AND FISCHINGER
0 A~~~~~~~~~~2
~20
D30
30
.-100
140
<50
<~~~~~~~~~~~~~50-Z06
160-
10-70
70-80
R80-if
0-~~~~~~~~~~~~~~~~~~~~~~~~~~~
lOC
10
10-3
102
01
100
110-3
10-2
1010
i0
Crt (mole
xsec/liter)
Crt (mole
xsec/liter)
FIG. 2. (a)Hybridization of cDNA fractions with 65S MSV-MuLVRNA.Hybridizations were performed in 0.1 mlof 0.22 MPB with2 pgof 65SMSV-MuLV RNA and 1,000 to 10,000counts/minof each cDNA fraction.Aliquots (0.01 ml) were taken at times varying from 2 min to 13 h.Incubation temperature was 63 C. Extent ofhybridization was measured byhydroxylapatite chromatography.Symbols: *,MSV-MuLV cDNA;
0,
cDNAcommo;
A,
cDNAteuk;
0,cDNAsarc
(b)Hybridization of
cDNAfractions
with33SMSV-MuLVRNA.Hybridizations were performed in 0.1 ml of 0.22 M PB with 0.8
pg
of30-40SMSV-MuLV RNAand1,000to 10,000counts/min of each cDNA fraction. Aliquots (0.01 ml) were taken at times varying from before the addition of RNA to 23 h after the addition of viral RNA. Incubation temperature was 63 C. Extent ofhybridization wasmeasured byhydroxylapatite chromatography.Symbols are the same as in (a).
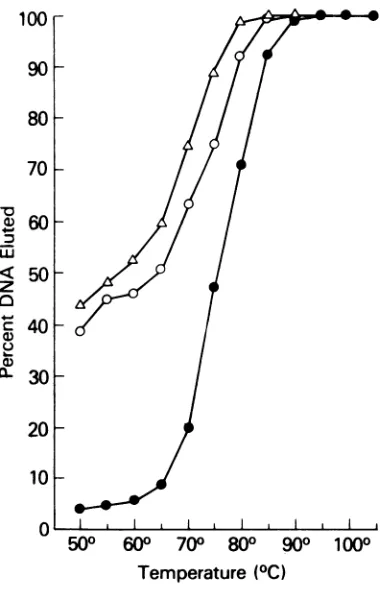
brids between the
cDNAfractions
andMSV-MuLV65S
RNA had
consistently high
Tm's
(78to83 C)
suggesting that the selected
sequenceswere
closely
matched
withsequences
in thehigh-molecular-weight
viral RNA
of
MSV-MuLV
(Fig.
3).The
unfractionated
cDNAhy-brid
with
homologous
RNAhad
aTm of
82.5C
as did the
cDNAsarc
hybrid.
cDNAcommon
andcDNAleuk had Tm's of
77.5 Cwith the
homolo-gous
MSV-MuLV
65S RNA.Composition
ofviral RNAs
based onprotec-tion of
hybrids from RNase.
Itwasof interest
to
determine what portions of MSV
or MuLV35S
RNAs
werecomprised
of"common,"
"leuk,"
or
"sarc" sequences.
Initially cDNA
wasalso
made
toM-MuLV
inthe
sameway
asfor the
MSV-MuLV
complex.
This MuLV cDNA
hy-bridized
83%with 70S
M-MuLV
RNA,
50%with MSV
(FeLV)-FeLV
RNA,
24% withFeLV
RNA, and
5%without
RNApresent.
Thermal
denaturation
of theM-MuLV cDNA
hybrid
withMSV
(FeLV)-FeLV was
13 Chigher
than
withthe FeLV
RNA(75 and 62
C,
respectively).
The
MSV-MuLV and MuLV cDNA's were
hybridized
to theirrespective viral 35S RNAs,
and
the
degree
ofhybridization
determined
byRNase
T,
digestion
atrelatively
lowmolar
ra-tios of
cDNA-RNA
ispresented
onTable
1.Large
proportions
ofeach RNA
hybridized with
either MSV-MuLV
or MuLV cDNA.The
ho-mologous
hybridization was larger,
but thecDNA of
MSV-M-MuLV
alsointrinsically
hy-bridized
more with theiodinated RNAs than
the
M-MuLV cDNA.
The
"common"
and"leuk" cDNA
hybridized
with
portions
ofMSV-MuLV
andMuLV RNA.
The "sarc" cDNA
only hybridized
withMSV-MuLV RNA
and notMuLV RNA. Hence
thefractions have the
expected
sequencespecific-ity.
The MuLV
genome appears to consist ofapproximately equal portions
of"common"
and"leuk"
sequences,whereas the MSV
genomehas
about
70to80%"common"
sequencesand 20to30%
"sarc"
sequences. Thesumofthe values
of the
degree
ofhybridization by
individual
probes
exceeds 100%.The "leuk"
sequences donot
have
the provenspecificity
of"sarc"
se-quences as no
iodinated RNA specific
forMSV
was
used.
This is
discussed below.
Distribution
ofMoloney
sequences amongmurine
RNA
tumorviruses.
cDNAsar,
hadnohomology
to KirstenMSV-MuLV
sequencesand
little
(4%
above
background) homology
tothe
Harvey
MSV-MuLV
viruscomplex
(Table
2). There
was nohybridization
of "sarc"se-quences with RNA
from the
murineecotropic
leukemia viruses
orwith
producer
cellRNA of
the
BALB-2
xenotropic
virus.cDNAcommon
sequenceshybridize
to Kirstensarcomaand
Harvey
sarcomaRNAs
(50
to61%)
but less wellto the
helper
virus RNA
present
with the
Harvey
sarcomasequences-15%with
FeLV
RNA.
Further,
cDNAcommon
hybridizes
only slightly
more with amixture
ofKirsten
and
Harvey
sarcomaMSV RNA than
toHar-J. VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:6.503.67.458.74.232.2]MURINE SARCOMA NUCLEOTIDE
100r
90h
80h-70
z
a)
C,
60
50
40
F
30
_20_
10 _
vI
50
60
70
80
90
100 [image:7.503.256.450.369.547.2]Temperature
(0C)
FIG. 3. Thermal elutions of cDNA hybrids with MSV-MuLV 70S RNA. Thermal elutions were from 1-cm hydroxylapatite columns with 0.12 M PB washes at 5 C intervals. Symbols: *, MSV-MuLV
cDNA;
O.cDNAcommon;
Ay,
cDNAleuk E,
cDNAsarc
vey
MSV RNA
alone (Table
3).This
suggeststhat
the
commonsequences present inKirsten
and Harvey MSVs
arefrom the
samesubgroup
of total
Moloney
commonsequences.Addition-ally,
aKirsten MSV-MuLV
cDNA,
prepared
similarly
tothe other
probes,
hybridized
more(44%)
with MSV-FeLV RNA than with FeLV
RNA
alone (20%). This Kirsten MSV cDNA
hybridized
90%with
homologous RNA with
a5%
background.
The
hybrid
of
Moloney
com-mon sequences
with
homologous RNA had
aTm
of
80C
(Fig.
4),the
hybrid of Moloney
commonsequences with Kirsten MsV-KiMuLV RNA had a
Tm
of 69C, and
that withHarvey MSV
RNA was 73 C.
Thus,
there is at least a 10% mismatch of common sequences.DISCUSSION
Complementary
DNA was transcribed fromthe
viruscomplex
consisting of MSV and M-MuLV. The cDNA was fractionated withse-A I .
quential absorption into
three subsets
repre-senting
various portionsof the
viral
genomes.The total cDNA
was ahomogeneous transcript
in that essentially complete
hybridization
ofhomologous
RNA wasachieved
at aslight
mo-lar
cDNA excess.The purity of cDNA
and its fractions
issup-TABLz
1. Percentageof hybridization ofiodinatedviral RNA to cDNA
firactionsa
%Hybridization of viral35S cDNArepresentative of: RNA
MSV-MuLV MuLV
MuLV 67 68
MSV-MuLV 89 66
"tcommon"p 74.5 52
"leuk" 28 42
"sarc" 20.5 7
0 1 5
aHybridization was performed as described in
Materials and Methods, and thehybridswere ana-lyzed at a
Cot
of 0.3 with acDNA-RNA molar excess of 2:4.1. In each case various cDNA concentrations were used, but a threefold excessof cDNA was suffi-cientto attain a maximalplateauvalue. RNaseT,
digestions of the hybrids were done as described in Materials and Methods.TABLz 2. Distribution of Moloneysarcoma and leukemia virusnucleotidesequences among murine
RNA tumorviruses Hybridization(%)to0: Viral RNA
MSV-MuLVcDNA,.,cDNAc0 cDNAIeuk cDNA
0 11 6 2 3
M-MSV-MuLV 84 67 97 90
M-MSV-FeLV 74 69 92 7
Kirsten MSV- 62 7 50 42
MuLV
HarveyMSV-FeLV 40 11 61 7
M-MuLV 86 7 92 90
Rauscher-MuLV 75 6 79 65
Kirsten-MuLV 33 3 37 40
BALB-2 35 6 51 44
NZBlive' NJI 0 26 j 19
a500-1,000counts/min ofeach cDNA fractionwas hy-bridizedin 0.22M PB at 63 C to afinal C,tof atleast10
mol-s/literwithcrude viral RNAs from each of the indicated viruses. The extent of hybridization wasanalyzed by hy-drozylapatitechromatography.
ICell RNA from A673 human cellschronically infected
withBALB-2 virusfromC3H/Hin-inducedmousecells used instead of viral RNA.Hybridization was taken toafinalCrt of2,500mol-s/liter. The RNAs from all eight viruseswere tested withhomologous complementary cDNApreparedin the presenceof100 jigof actinomycin D permlandwere foundtohybridizeto atleast70%.
eCell RNAfromNZBliverwasused. FinalCrtof19,000 mol-s/liter used.
dNT, Not tested.
on November 10, 2019 by guest
http://jvi.asm.org/
488 FRANKEL, NEUBAUER, AND FISCHINGER TABLE 3. RelationofMoloney common sequences
withnucleotide sequences in Kirsten MSV and Harvey MSV RNAs
Hybridiza-ViralViralRNARNA
~cDNk OA00a
tionto(%)
O
... .. . .. .. . .. . . 2KirstenMSV-KiMuLV .51
HarveyMSV-FeLV ... .. 61
Kirsten MSV-Kirsten MuLV + Harvey 71 MSV-FeLV ...
Kirsten MuLV .37
FeLV 15
aHybridization conditions andevaluations were the same as inTable 1.
bKirstenMSV-Kirsten MuLV and Kirsten MuLV
werebothprepared in the same NIH 3T3 cell sys-tem.
This
washopefully
avoidedby
the fact thatcDNAsarc
washybridized
toMSV(FeLV)
RNA.Because six consecutive viral
passages
ofMSV(FeLV)
wereperformed
in cat cells andthenthe infected
virus-producing
cat cell cul-ture waspropagated
for >3years,
it is assumed thatonly
thosesequences
that were the MSV genome itself were conserved.Second,
based on lack ofhybridization
ofcDNAsarc
withRNAs
of two types of murinexenotropic
viruses,
cDNAsa.,
consisted of
se-quencesother than those absent from
MuLVbut
present
inmurinexenotropic
oncornavirus.Third, the cDNAsarc partly hybridized with
ratcell
DNA, indicating
that thisM-MSV isolate isquite
dissimilar from several otherMSVs,
which also
containinformation other than
thatof
MuLV (22, 23).
This
M-MSV genomegives rise in
variousported by the
Crt curveswith unfractionated
and
fractionated cDNA's. A
CrtAi2
of
0.02 to0.05
isnear
the predicted
Crtl
for
amixture
of
twoRNA
tumorviruses.
The similar
CrtA2
of the
"common"
and
"sarc" sequences attests totheir
not being transcripts
of
minor contaminants,and the lower
Crt,/,
of
the "leuk"
sequencessupports
the
-3:1 sarcoma excess inthe
mix-ture.
The FeLV RNA
hybridizations with the
probe
fractions
were at nearbackground level
for
the sarcoma-related
sequences.Thus, there
is no
enrichment of FeLV-related
sequences inthe MSV(FeLV)-FeLV
fractionations.
Finally,the
high homologous
Tm's of cDNA fractions
with
65S MSV-MuLV RNA favors
accuratetranscripts
of viral
sequences inthe
cDNA's.An
understanding
ofthe compositionof
MSVRNA could
be obtained from the hybridization
experiments
of iodinated viral
RNAwith cDNAtested
by RNase
digestion. Sarcoma-specific
cDNA
represents 20% of the totalMSV-MuLV
viral
sequences orabout
25%of the MSV
sequences
based
onthe
3:1sarcoma excessby
biological titrations and
Crtcalculations
per-formed
onthe
sameviral harvest
used to makeiodinated subunit RNA. The other 75% of the MSV RNA hybridizes with "common" cDNA. TheMoloney leukemia viral genome consists of aboutequal portions of common sequences and
leukemia-specific
sequences. The "sarc" portionis atleast as large or up to twice the size of the
"sarc"
portion of the Rous sarcoma genome, asreported by Stehelin et al. (in press).
The nature
ofsequences
of thecDNAr,
isof
interest
because
it may contain the sequencesresponsible for
celltransformation. It isimpor-tant to rule out that
these
are not spurious normal mouse DNAsequences transcribed andrecognized
asapart ofthe
trueMSV genome.100
r
90h
80
H
70F
-0 a)
LU
z 0
40)
C.)
a-60H
50
V
40
H
30
H
20 K
10o
II
v500
600
700
800
90Temperature
(0C)
FIG. 4. Thermal elutions ofcDNAommon hybrids
with different MSVRNAs. Thermal elutions were from 1-cm hydroxylapatite columns with 0.12 M PB washesat5Cintervalswerecarriedoutonhybrids betweencDNAcommonand viralRNA of MSV-MuLV,
Kirsten MSV-Kirsten MuLV, and Harvey MSV-FeLV.Thehybridswerestudied after reachingatCrt of 10 mol-slliter.Symbols: 0, M-MSV-MuLV RNA;
A, Kirsten MSV-Kirsten MuLV RNA; 0, Harvey
MSV-FeLV RNA.
1000
[I
J. VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:8.503.264.454.281.576.2]species
toS+L-
typecells,
which express theantigenic gs-1 determinant
normally found onthe
p30
polypeptide (9, 19). Actually the pre-dominent gs-1 containing polypeptide inheter-ologous
S+L- cells is a molecule of ap60
sizewhich does contain, with some other determi-nants, the gs-1 site (17). No heterologous
S+L-cell
tested expresses the MuLVgp71,
andre-cent
data indicate that the
detectable reversetranscriptase enzyme isonly that of the helper virus (Peebles et al., in press; Fischinger and Bolognesi, unpublished data). Thus, some
MuLV-specific
products are a part of theS+L-typeMSV genome and some other MuLV-coded
information
may be missing or not expressed.This
is compatible with several of theother
MSV
genomes,which do
not express MuLV proteinsbut do contain MuLV-specific informa-tionexpressed in the RNA of transformed cells (20).Apparently the
S+L- MSV genome isanalogous
toother
hybrid
oncornavirusioslates
in
that
it arose by the accretion of new se-quences,presumably by
arecombinational
event (12, 20).
The
lackof homology
betweenthe
cDNAsar
of the S+L-
genomeand the
Kirstenand
Harvey MSV RNAs poses a special question.
Apparently the
transcriptionof dissimilar
mo-lecular
sequencesfound
in variousMSVs
canresult
in mousefibroblast transformation.
Even
the
different
aviansarcoma virus genomecan
give
rise tothe
sameresult,
although avian
cDNAsarc and mammalian
transforming
sar-coma viruses
have
no sequences in common(Stehelin
etal.,
inpress).
Itis
important
todetermine
whether cDNAsarc
sequencesfrom
S+L-
typeMSV
arerepresented
innormal
mouse DNA.
Recently
these
sequences wereindeed
found
innormal
mouse DNA (ourun-published data). Also,
one mustask whether
transformation of cells by different
virusesleads
toexpression
of
common sarcomavirus-associated
sequences incell RNA.
Since the submission
of this
manuscript,
analogous data have been
published
which
arein
general
agreementwith the above
conclu-sions
(21).
ACKNOWLEDGMENTS
We thankDave Colcher and Dave Swan for iodination of high-molecular-weight RNAs, Cy Cabradilla and Stuart Aaronson for thegiftof Kirsten sarcoma-Kirsten leukemia viralRNA, and DanielHaapalaforvaluable discussions.
LITERATURE CITED
1. Aaronson, S.A., andW.P. Rowe. 1970. Nonproducer clones of marinesarcoma virus transformed BALB 3T3 cells. Virology42:9-19.
2. Bassin, R. H., N. Tuttle, andP. J. Fischinger. 1970. Isolationof murinesarcomavirustransformedmouse
cells which are negative for leukemia virus from agar suspensionculture. Int. J. Cancer6:95-107. 3. Britten, R. J., and D. E. Kohne. 1968. Repeated
se-quences in DNA. Science 161:529-540.
4. Britten, R. J., and J. Smith. 1970. Abovine genome. Carnegie Inst. Washington Yearb. 68:378-386. 5. Colcher, D., W. Drohan, and J. Schlom. 1976.
Mason-Pfizer virus RNA genome: relationship to the RNA of morphologically similar isolates andothers oncorna-viruses. J.Virol. 17:705-712.
6. Fanshier, L., A. C. Garapin, J. McDonnell, A. Faras, W.Levinson, and J. Bishop. 1971.Deoxyribonucleic acid polymerase associated with aviantumorviruses: secondary structure of the deoxyribonucleic acid product. J. Virol. 7:77-86.
7. Fischinger, P.J., C. 0. Moore, and T. E. O'Connor. 1969. Isolation and identification of a helper virus found in the Moloney sarcoma-leukemia virus com-plex. J.Natl. Cancer Inst. 42:605-622.
8. Fischinger, P. J., and T. E. O'Connor. 1969. Viral infection across species barriers: reversible alteration of murine sarcoma virus for growth in cat cells. Sci-ence 165:714-716.
9. Fischinger, P. J., W. Schafer, and E. Seifert. 1972. Detection of some murine leukemia virus in virus particles derived from 3T3 cells transformed only by murine sarcoma virus. Virology47:229-235. 10. Hartley, J. W., and W. P. Rowe. 1966. Production of
altered cellfoci in tissue culture by defective Moloney sarcoma virusparticles. Proc. Natl. Acad. Sci. U.S.A. 55:780-786.
11. Harvey, J. J. 1964. An unidentified virus which causes the rapid production of tumorsinmice.Nature (Lon-don) 204:1104-1105.
12. Hayward, W. S., and H. Hanafusa.1975. Recombina-tionbetweenendogenousandexogenousRNA tumor virus genes asanalyzed bynucleicacidhybridization.
J. Virol. 15:1367-1377.
13. Huebner, R. J., J. W. Hartley, W. P. Rowe, W. T. Lane, and W. J. Capps. 1966.Rescue of the defective genome ofMoloney sarcomavirus from a
noninfec-tioushamster tumor and the productionof
pseudo-type sarcoma viruseswith various murine leukemia viruses.Proc. Natl. Acad. Sci. U.S.A. 56:1164-1169. 14. Leong, J., A. C.Garapin,N.Jackson, L.Fanchier, W. Levinson, and J. Bishop. 1972.Virus-specific ribonu-cleic acid incellsproducing Rous sarcoma virus: de-tectionandcharacterization. J. Virol. 9:894-902. 15. Moloney, J. B. 1966. Avirus-induced
rhabdomyosar-comaof mice. Natl. Cancer Inst. Monogr.22:139-142. 16. Nomura, S., P. J.Fischinger, C. F. T.Mattern, P. T. Peebles, R. H. Bassin, and G. P. Friedman. 1972. Revertants of mouse cells transformed by murine sarcomavirus. I.Characterization of flat and trans-formedsublines withoutarescuablemurine sarcoma virus.Virology 50:51-64.
17. Oskarsson, M.K., W.G. Robey, C. L.Harris, P.J. Fischinger,D.K.Haapala,and G. F. VandeWoude. 1975. Ap60polypeptideinthe feline leukemia virus
pseudotype ofMoloney sarcomavirus with murine leukemia virus p30 antigenic determinants. Proc. Natl. Acad. Sci. U.S.A.72:2380-2384.
18. Peebles, P. T., R. H. Bassin, D. K. Haapala, L. A. Phillips, S. Nomura, andP.J.Fischinger.1971. Res-cueof mouse sarcomavirusfromasarcomapositive
leukemianegative cellline:requirementfor
replicat-ing leukemiavirus. J.Virol.8:690-694.
19. Peebles,P.T.,P.J.Fischinger,R. H.Bassin, andA. G.Papageorge.1973.Isolationofhumanamnioncells transformedbyrescuablemurine sarcoma virus. Na-ture(London)New Biol.242:98-101.
20. Scolnick, E. M., R. T. Goldberg, and W. P. Parks. 1975.Abiochemical andgeneticanalysisof
on November 10, 2019 by guest
http://jvi.asm.org/
490 FRANKEL, NEUBAUER, AND FISCHINGER lian RNA-containing sarcomaviruses. ColdSpring
Harbor Symp. Quant.Biol. 39:885-895.
21. Scolnick, E. M., R. S. Howk, A. Anisowicz, P. T. Peebles, C. D.Sher, and W. P. Parks. 1975. Separa-tion ofsarcoma specific and leukemia
virus-specific geneticsequencesofMoloneysarcomavirus. Proc. Natl. Acad. Sci. U.S.A. 72:4650-4654. 22. Scolnick, E. M., and W. P. Parks. 1974. Harvey
sar-comavirus: asecond murinetypeCsarcomavirus withratgeneticinformation. J. Virol. 13:1211-1219. 23. Scolnick, E. M., E. Rands, D. Williams, and W. P. Parks. 1973. Studiesonthe nucleic acidsequencesof
Kirstensarcomavirus: a model forformation ofa
mammalian RNA-containingsarcomavirus. J. Virol.
12:458-463.
24. Snyder, S. P., and G. H.Theilen. 1969. Transmissable feline fibrosarcoma. Nature (London) 221:1074-1075. 25. Solymosy, F., I. Fedorcsak, A. Gulyas, G. L. Forkas, and L.Ehrenberg. 1968. Anewmethod basedonthe
useofdiethyl pyrocarbonateasanuclease inhibitor for the extractionof undegraded nucleic acid from plant tissues. Eur. J.Biochem. 5:520-527.
26. Temin, H. M., and D. Baltimore. 1972. RNA-directed DNAsynthesis and RNAtumorviruses. Adv. Virus Res. 17:129-136.
27. Wolfe, L. G., F. Deinhardt, G. H. Theilen, H. Rabin, T. Kawakami, and L. K. Bustad. 1971. Induction of tumorsinmarmosetmonkeys by simiansarcoma
vi-rustypeI(Logothrix):apreliminary report. J. Natl.
Cancer Inst. 47:1115-1120.
J. VIROL.