0022-538X/91/105410-07$02.00/0
Copyright C) 1991, American Society for Microbiology
Recombination of the Internal
Direct
Repeat
Element DR2
Responsible for the
Fluidity
of the
a
Sequence
of
Herpes
Simplex Virus Type 1
KENICHI UMENE
Departmentof Virology, Faculty of Medicine, Kyushu University 60, Fukuoka812, Japan Received 13 March 1991/Accepted 10 July 1991
A seriesof herpes simplexvirustype1derivatives,havingasequencescomposedofDR1, Ub, (DR2)3-7,DR4t (a truncated formofDR4), and Uc were isolated and examined. The derivative havinga sequences with six copies ofDR2 generated progeny viruses having a sequences with the same number (six copies) of DR2. Anotherderivative, havingasequenceswith threeand sevencopiesofDR2, generatedprogenyviruseshaving asequenceswith varied numbers (4,5, 8, and 10copies)ofDR2, besides theoriginal DR2arrays (threeand seven copies). Therefore, the variation in copy number of DR2 was assumed to be caused mainly by recombination between DR2arraysrather thanbyslippagewithin a DR2arrayduringDNAreplication.The presence of DR2-like sequencesin internal directrepeatelementsofDR4andDR3.5supportedthehypothesis oftherecombinogenic propertyofDR2. Theequal distribution ofdivergence ofasequencestoboth ends of the virus genome favors the double-strand break and gap repair model to explain gene conversion and amplification ofthe asequence.
The 155-kb genome of herpes simplex virus type 1 (HSV-1) consists of two covalently linked components, L and S (Fig. 1A), which invert relative to each other (L-S inversion)(12, 19). A short sequence, a, isrepeated directly atboth ends of the genome and is also present in the inverse orientation attheL-Sjunction. One toseveralcopiesof the asequence is present atthe endofthe L component andat the L-S junction, but only one copy is presentattheend of the S component (6, 13, 14, 16, 19, 34). The a sequence encodes several cis-acting sites involved in (i) the circular-ization of viral DNA after infection (13-15, 33), (ii) the cleavage of unit-length DNA from concatemeric forms gen-erated by rolling-circle replication and encapsidation of the excised molecules (4, 7, 14, 15, 17, 19, 33), (iii) the expres-sion of an mRNA extending from the a sequence (1, 3, 5), and (iv) theL-S inversion caused by homologous recombi-nation between a sequences arranged in an inverse orienta-tion (2, 16, 20, 32).
Thea sequencecontains unique (U) and directly repeated (DR) sequence elements and is flanked by direct repeats of a 20-bp sequence termed direct repeat 1 (DR1) (Fig. 1B). Tandemly reiterated a sequences share the intervening DR1, and processing of the concatemeric form into unit-length viral DNA involves asymmetric cleavage of DR1 shared by thetwoadjacenta sequences (6, 13-15, 17, 32). Theinternal repeated arrays of DR2 and DR4 were formerly assumed to be cis-acting sites for the L-S inversion mediated by the a sequence (2, 32); however, the recombination between other inverted repeats (besides the a sequences) can also cause inversion (9, 18, 26, 31, 35, 36). Smiley et al. (20) recently showed that recombination events leading to L-S inversion didnotoccur at asingle site within the a sequence, and they proposed that recombination between a sequences occurs by astandard homology-dependent generalized recombination. Since the sequence of DR2 and DR4 might enhance the recombinational activity of the a sequence, the function of theinternal repeated arrays remained to be addressed.
The a sequences of various HSV-1 strains range in size from 250 to 500 bp and mainly depend on the number of
reiterations of internal DR elements (6, 13, 15, 32). In the presentstudy,a series of HSV-1 derivativesdifferingin the copy numberof DR2wereisolated and examined.Analyses of progeny viruses from these derivatives revealed that recombinational events between DR2 arrays are the main causeof variation in DR2 copy number. Theresults, and the presence of DR2-like sequences in other internal DR ele-ments, strengthened the case for the recombinogenicity of DR2 (2, 13, 15). Equal distribution of a sequences with different copy numbers of DR2 to both ends of the virus genomefavors the double-strand break and gaprepairmodel to explain gene conversion and amplification of the a se-quence (7, 8).
MATERIALS ANDMETHODS
Viruses and cells. HSV-1 strains SP23 (28), Bi (25), K41, K57, andTy154 (30)were asdescribedpreviously. Working stocks of HSV-1 were madeon Vero cells inEagle minimal essential medium and 2% fetal bovineserum.
Single-cloneisolation ofvirusesandextraction of viralDNA. Toeach well of the 96-wellmicrotest platewasadded0.2 ml containing 5 x 104 cells and 0.1 PFU of virus (24). The clonedprogeny viruseswererecovered after the cytopathic effect become apparent.AVero cell monolayer (5 x
106
cells per culture) infected with a cloned HSV-1 stock was col-lectedby low-speed centrifugation, and the viral DNA was extracted by the method of Hirt, as previously described (24).Restriction endonuclease digestion, gel electrophoresis, and Southernhybridization. Restriction endonucleases were pur-chased fromToyobo Co. (Osaka, Japan), and conditions for digestion were those recommended by the manufacturer. The DNAs digested with restriction endonucleases were separated in a 5% acrylamide gel or a 1% agarose gel. Southern hybridization was carried out on a Biodyne A transfer membrane (Pall Ultrafine Corp.) as previously de-scribed (23).
DNA sequencing. A restriction fragment was subcloned
5410
on November 10, 2019 by guest
http://jvi.asm.org/
a L S U,
A
Kpni ,
IRL aiRs IL TRsa
R I K
I IL
B DRI Ub
EJJDR2
t UcDRI UB=
Smal
Dral
FIG. 1. Maps of HSV-1 DNA. (A) Structure of the HSV-1 genomearranged in the prototype orientation (19). L and S compo-nents constitute 82 and 18% of the genome, respectively. Each componentconsists of unique sequences (UL andUs)bracketed by inverted repeat sequences (TRL, IRL,IRS,andTRs).One to several copies of the a sequence is present at the end of the L component and at the L-S junction, but only one copy is present at the end of theS component (6, 13, 14, 16, 19, 34).KpnIfragments at the ends of L andS components are shown (11). (B) Schematic representa-tionof the asequence, based on the nucleotide sequences of that of TW14determined in this work (see Fig. 4a). The cleavage sites of SmaI and DraI (anisoschizomer ofAhaII)areshown. The region corresponding to the 0.205-kb SmaI fragment (23, 30), used as a probein Southernhybridizationfor the detection of a sequences, is indicatedas a double line. TheDral fragment, corresponding to a unit length of the a sequence, is also indicated as a double line.
into both M13mplO and M13mpll and then sequenced by using the dideoxynucleotide chain termination procedure
(27).
RESULTS
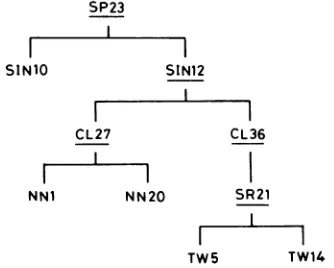
Segregation of HSV-1 derivatives having a sequences of different lengths. The length of the a sequence differs with the strain ofHSV-1 (6, 13). The difference was evident in Southern hybridization analyses of SmaI-cleaved HSV-1 DNAs, with a0.205-kb SmaI fragment (containing most of the a sequence) of XDD-1 (carrying the 9.5-kb EcoRI frag-ment of classIdefective DNAof HSV-1 strain Patton) asa probe (Fig. 1B) (23). The length of the a sequence of single-plaque clones derived from SP23 was examined by Southern hybridization analysis (30), and three classifica-tions were made: those generating two SmaI fragments of 0.21-and0.22-kb,such asthe parent SP23;thosegenerating
SP23
SINIO SIN12
CL27 CL36
I
NN1 NN20 SR21
[image:2.612.64.298.79.195.2]TW5 TW14
[image:2.612.320.562.101.478.2]FIG. 2. Genealogyof clones derived fromanHSV-1 strain SP23 (28). The clonesunderlinedarethosegeneratingtwoor more kinds ofa sequences (Table 1).
TABLE 1. Summary of lengths ofSmalfragments of SP23 derivatives shown in Fig. 3
Parentclone No. of
Panel in (total no.of L Length ofSmaI clones
Fig. 3 examined ane fragment (kb) (nameof
clones)" clone)
A SIN10 (32) P 0.21
1-10 0.21 32
B SIN12 (39) P 0.20,0.21, 0.22, 0.24
1 0.21, 0.24 7
2 0.21, 0.22 5
3 0.21 5
4 0.22 3
5 0.22,0.24 6
6 0.21,0.22, 0.24 5
7 0.20, 0.21 3(CL27)
8 0.20, 0.21, 0.24 3 9 0.22,0.24, 0.26 1 (CL36) 10 0.20, 0.21, 0.22 1 C CL36 (25) P 0.22, 0.24, 0.26
1 0.22, 0.23 1
2 0.22,0.26 5
3 0.22, 0.24, 0.26 5
4 0.22 11
5 0.22, 0.26, 0.285 1 6 0.175, 0.22 1 (SR21) 7 0.24, 0.26, 0.275 1 D CL27(24) P 0.20, 0.21
1 0.20 2(NN20)
2 0.20, 0.21 11
3 0.21 11 (NN1)
E SR21 (25) P 0.175, 0.22
1 0.175, 0.22 11
2 0.175, 0.22, 0.25 1
3 0.22 9(TW5)
4 0.22, 0.23 1
5 0.175 1(TW14)
6 0.185, 0.22 1
7 0.175, 0.20, 0.22 1 "P,parentalclone.
oneSmaIfragmentof0.21kb(one clonewasnamedSIN10); and an isolate (named SIN12) generating four SmaI
frag-mentsof0.20, 0.21, 0.22, and 0.24 kb (Fig. 2) (Table 1). Single-plaque clones derived fromSIN10 and SIN12were
further examined to analyze the mode offluctuation of the lengthof thea sequence and alsotoisolate the clone with a
shortera sequence. All 32clones fromSIN10hada0.21-kb SmaIfragment, asdid theparental
SIN10,
while those from SIN12 wereheterogeneousfor the lengthof thea sequence (Fig. 3A and B).Thirty-nine
clones from SIN12 contained one to three SmaIfragments
of0.20, 0.21, 0.22, 0.24, and 0.26kb(notdetected in theparental
SIN12clone)
andwereclassified into 10 groups (Table 1). CL27 (with the shortest SmaIfragment, of 0.20kb)andCL36
(with
thelongest
one, of 0.26kb) were usedfor furtheranalysis (Fig. 2).
Inadditionto theSmaI fragmentspresentin the
parental
clone CL36 (0.22, 0.24, and 0.26
kb),
four otherfragments
(of0.175, 0.23, 0.275, and 0.285 kb) were found insingle-plaque clones derived from CL36
(Fig.
3C). The 0.175-kb fragment of SR21 was the shortest(Table
1). Clones from CL27 had one or two SmaIfragments
of 0.20and 0.21 kb (Fig. 3D). NN20 (with one SmaI fragment of 0.20kb)
and.3 -L -1 --". tis I
on November 10, 2019 by guest
http://jvi.asm.org/
[image:2.612.101.268.569.703.2]A)
S P 1 2 3 4 5 6 7 8 9 10 P M -0.2 71
* * __ -0.234
-0.194
S P 1 2 3 4 5 6 7 8 9 10 P M
B)
S P 1 2 3 4 5 6 7 P M
C)
E)
-0.27i
-0.234
-0.194
D)
P1 2 3 M
VW :-"-. -0.271
&&***_^
*a
olg4
-0.234-.l194
S P 1 2 3 4 5 6 7 P M
-0.271
,,W
40 .,0.234z-0.194
FIG. 3. Southernhybridization profiles showing variation in the lengths of a sequences among HSV-1 derivatives. HSV-1 DNAs
were cleaved with SmaI, electrophoresed in 5% acrylamide gels,
transferred toanylon membrane, and hybridized with 32P-labeled
0.205-kbSmaIfragment (containingmostofa sequence)of XDD-1
as aprobe (Fig. 1B) (23, 30). Electrophoretic patterns of single-plaque clones from SIN10 (A), SIN12 (B), CL36 (C), CL27 (D), and SR21 (E)areshown in lanes 1to3,7, and 10, respectively,ineach panel. Lengths of the SmaI fragments are summarizedinTable 1. Parentalclone ineach panel is shown in lane P. Lane S wasSP23, as a reference (28). Lane M was the marker mixture of HaeIIl
digests of phage OX174 DNA (23). Sizes of fragmentsaregiven in
kilobasepairs.
NN1 (withoneof 0.21 kb)wereused for further study (Fig.
2).
SR21 generated twoSmaIfragments of0.175 and 0.22 kb (Fig. 3C, lane 6). In additiontothetwoSmaI fragments, four otherSmaIfragments (of 0.185,0.20, 0.23,and 0.25kb)were
found in single-plaque clones derived fromSR21 (Fig. 3E). TW5 (with oneSmaI fragment of 0.22 kb) and TW14 (with oneSmaIfragment of 0.175 kb [the shortest]) wereused for
furtherstudy (Fig. 2).
Characterization of four syngenic HSV-1 derivatives dif-feringinthelength oftheasequence.The fourHSV-1 isolates
(TW14,NN20, NN1, and TW5) derived from SP23 hadone
kind of a sequence and differed in the length of the a
sequence. Restriction endonuclease Dral (an isoschizomer
ofAhaIII) cleaved theasequence atasiteonUb (Fig. 1B).
DraIfragments ofthe asequences of these isolates,
repre-senting a unit length of the a sequence, were cloned into
pUC18andsequenced. TheasequenceofTW14 (the
short-est) consisted of 217 nucleotides (nt) (from DR1 to Uc), containing three copies ofDR2(Fig.4a). The composition of
thea sequences of NN20, NN1, andTW5 wasthe same as
that ofTW14, exceptfor the copy number of DR2. The a
sequences of NN20, NN1, and TW5 consisted of 241 nt (5 copies of DR2), 253nt(6 copies of DR2), and 265nt(7 copies of DR2), respectively.
The four isolates TW14, NN20, NN1, and TW5 could be supposed to be the same, except for the copy number of DR2. When one-step growth curvesof the four isolates on Vero cells were constructed (27), no differences were evi-dent. The difference in copy number of DR2 (from three to seven)probably hadnosignificant effectonthegrowth of the isolates in cell culture.
Distribution ofdivergence of theasequencestoboth ends of the HSV-1 genome. SR21 generated two SmaIfragments of 0.175 and 0.22 kb (divergenceofa sequences). The original virus particle of SR21 had packageda DNA molecule with three or morecopies ofa sequences presumedto be oftwo kinds (Fig. 1A). The presence of multiple copies of the a sequenceat the end of the L componentmight accountfor thedivergence ofasequencesatthe end of this component. Since only one copy of thea sequence is presentatthe end ofthe S component(14),thea sequence atthe end of the S componentof the original SR21 should be ofone kind, not two.Thequestionarose astowhether theasequencesatthe endofthe S componentofthereplicatedgenomemolecules remained homogeneous (onekind)orbecameheterogeneous (twokinds).
DNAs of SR21 were cleaved with KpnI (Fig. 1A) and electrophoresed in a low-melting-temperature agarose gel (11). KpnI-I and -Kfragments (containing the endofthe S component) andKpnI-R1 and -R2fragments (containingthe endof the L component and having one and twocopies of the a sequence, respectively) were purified (Fig. 5A). The DNAs ofKpnI-I, -K, -R1, and -R2fragments were cleaved with SmaI and hybridized with the 32P-labeled 0.205-kb fragment(correspondingtothea sequence) usedas aprobe (23). All KpnI fragments generatedboth 0.175- and 0.22-kb SmaIfragments (Fig. SB), therebyindicatingthe
divergence
of the a sequence at both ends of the replicated genomes, while the a sequence at the end of the S component of the originalvirus genome should have beenofonekind. Similar studies were carried out with SP23 (generating 0.21- and 0.22-kb SmaI fragments) and CL27 (generating 0.20- and 0.21-kbSmaIfragments),anddivergenceof the asequences at both ends of the replicated genomes was likewise ob-served.
Nucleotide sequencesof asequences ofHSV-1strainshaving short a sequences. The nucleotide sequences ofthe a se-quencesof strainsJustin(244 nt), KOS (297 nt), 17 (399 nt), USA-8 (415 nt), and F(482 nt) were determined (6, 13, 15, 32), and structural requirement in the a sequence was inferred (7). Shorta sequences should contain less dispens-able sequences than do the long a sequences. Therefore, a comparison of nucleotide sequences of the shorter a se-quences is worthwhile when attempting to elucidate the basicstructure of thea sequence.Thenucleotide sequences of the a sequences of 4 HSV-1 strains having shorter a sequences of K57(234 nt), K41 (255 nt),
Bi
(255 nt), and TylS4(256 nt) (25, 30)were determined (Fig. 4b to 4e).DISCUSSION
Variation in length of the a sequence among clones derived from SP23 was seento be due todifferences in the copy number of DR2. SIN10, which bears one SmaI frag-mentof0.21 kb, generated only progeny viruses bearing the SmaIfragmentof 0.21kb,identicaltothatof the parent virus (Table 1). All25 clones derived from TW14 bore one SmaI
A.IP. 'no 40 a
.10 4M... .AM&k
,O oiq'Jillillippilliliv
on November 10, 2019 by guest
http://jvi.asm.org/
[image:3.612.71.284.78.376.2](a) TW14
30 60 90
CCGCGGGGGGCCCGGGCTGCCCGCCGCCGCGCTTTAAAGGGCCGCGCGCGACCCCCGGGGGGTGTGTTTGGGGGGGCCC
CGGGG
DR1 Ub
120 150 180
TCCGCCGCTCCTCCCCCCGCTCCTCCCCCCGCTCCTTCCCGCGGCCCCGCCCGCCGC
CCGCGCGCGCGCACGCCGC
DR2 DR2 DR2 DR4t Uc
210 217
cCGGACCGCG0C S TTCCoCoCoCoC
(b)
K5730 60 90
CCGCCGGCGGCCCGGGCTCCCCGCCGCCGCGCTTTAAAGGGCCGCGCGCGACCCCCGGGGGGTGTGTTTCGGGGGGGGGCCGTTTTTGGG
DR1 Ub
120 150 180
GTCasGGcGCTcCTc UC*CGCTCCTCCCCCGCTCCTCCCCCGCTCCTCCCCCGCCGCCCGCCTTTTTCGGCCCCGCCCCCCACGCCCGCC
DR2 DR2 DR2 DR2 DR4n Uc
210 234
GcGcGcGcGcAcGLccGcccGAccGccGcccGccnnTaTTGCGCGCGCGCACGC
(C) K41
30 60 90
CCGCX*GGGGGcccGGGCTGccCGCCGCGCGCTTTAAAGGGCCGCGCGCGACCCTCGGGGGGTGLTSTGOC0GGGGGGCCCGTTTTCGG
DR1 Ub
120 150 180
GGTCTGGcCGTrCTCCCCCCGCTCTCCCCCCGCTCCTCCCCCCGCTCCTCCCCCCGCTCCTCCCCCCGCTCCTCCCCCCGCTCCCGCG
DR2 DR2 DR2 DR2 DR2 DR2 DR4t
210 240 255
GCCccGccccccAcGcc GJAucGccGCGCAcGcCGCCCGGACCGCCGCCCGCC1TSSAZCOZOCGCGCGCGC
Uc
(d) Bi
30 60 90
cts LZGGGGGGGGCCGCCGCTTTAAAGGGCCGCGCGCGACC L CGTTTTTGGGG
DR1 Ub
120 150 180
TCTGGGCCTCCTCCCGCTCCTCCCCCGCTCCTCCCCCGCTCCTCCCCCGCTCCTCCCCCGCTCCTCCCCCGCCGCCCGCCTTTTTCG
DR2 DR2 DR2 DR2 DR2 DR2 DR4n
210 240 255
GCC X*c c cc cCGCCX CoCoCoCGCCoCCCooACCoCCGoCCcoc-trr CoCC 0'CGCACGC
Uc
(e) Ty154
30 60 90
CCGCGG*GGGCCC GGGCGCCCGCCGCCGCGCTTTAAAGGGCCGCGCGCGACCCCCGGGGGGTGTGTTTCGGGGGGGGCCCGTTTTTGGG
DR1 Ub
120 150 180
GTCTGGGCGCTCCTCCCCCGCTCCTCCCCCGCTCCTCCCCCGCTTCTCCCCCGCTCCTCCCCCGCTCCTCCCCCGCCGCCCGCCTTTTC
DR2 DR2 DR2 DR2 DR2 DR2 DR4n
210 240 256
[image:4.612.74.544.87.599.2]GGCCCCGCCCCCCACGCCCGCCGCGCGCGCGCACGCCGCCCGGACCGCCGCCCGCCTTTTTTGCGCGCGCACGC Uc
FIG. 4. NucleotidesequencesofHSV-1 strainsTW14(aderivativeofSP23) (a)(28),K57(b),K41(c)(30), B1 (d)(25),andTy154(e) (30). Nucleotide numbersstart attheleft end oftheleftDR1andterminateattherightend of Uc. Theleftendof eachcomponent of theasequence
is indicated. DR1and DR2 ofTW14(a)areunderlined. Definitions ofDR4tand DR4nare
given
in thetext.fragment of0.175 kb,asdid the parent. Uniformity of thea
sequence in progeny viruses derived from a virus with homogeneousa sequences indicated that variation in
length
of the a sequences was
rarely
causedby slippage
during
DNA replication (29, 30).
CL27,
which bears two SmaI fragments of 0.20 and 0.21kb,
generated
only
progeny viruses bearing SmaI fragments of 0.20 and/or 0.21 kb. However, three isolates, SIN12,CL36,
andSR21,
which bear more heterogeneous a sequences,generated
progenyvirusesbearingasequencesabsent in eachparentvirus,
i.e.,
a0.26-kb SmaI
fragment
fromSIN12; 0.175-,
0.23-, 0.275-, and0.285-kb SmaIfragments
fromCL36;and0.185-, 0.20-, 0.23-, and 0.25-kb SmaIfragments
from SR21(Table
1). The modeoffluctuationoflength
ofthe a sequence(i.e.,
varia-tion in copy number ofDR2)
waselucidatedby considering
recombinational events between two DR2 arrays(Fig.
6).Two a sequences
bearing
the same copy number of DR2 (e.g., SIN10 andTW14)
could form apair
homologously;
on November 10, 2019 by guest
http://jvi.asm.org/
(A)
H 1 2 34 M -23.1
(a)
I I IDR2 I I 1I4 I I I I I lF
(B)
H 1 2 3 4 M
a
to'I.
a,
*
-0.271
-9.4 e -0.234
-6.6 -0.194
0 d* 1f.i,
-4A
-0.118
-2.3
FIG. 5. Southern hybridization analyses of terminal regions of HSV-1 DNAs. Kpnl-l(lane 1), -K(lane2), -R1(containingonecopy of the a sequence) (lane 3), -R2 (containing two copies of the a sequence) (lane 4) fragments from SR21 were purified from a
low-melting-temperature agarose gel. Kpnl-l and -K fragments contain the terminalregionsof the S component, andKpnI-R1and -R2fragments contain those of the L component(Fig. IA) (11). (A) SR21 genome DNAs digested with Kpnl (lane H) and the Kpnl fragments from SR21 (lanes 1 through 4) wereelectrophoresed in a 1%agarose gel, transferred to a nylon membrane, and hybridized with a 32P-labeled plasmid DNA carrying the BamHl-(T+R) frag-mentof HSV-1 (containing the L-Sjunction) (11,23). Lane M isa
marker mixtureofHindlIl fragmentsofphageXDNA(23). Sizesof fragments are shown in kilobase pairs. (B) SR21 genome DNAs (laneH)andtheKpnI fragments fromSR21(lanes1through4) were digestedwithSmaI, electrophoresedina5% acrylamide gel,
trans-ferred to a nylon membrane, and hybridized with the 32P-labeled
0.205-kb SmcaI fragment (containing most of the a sequence) of XDD-1 (23, 30). Lane M is a marker mixture of Haellldigests of phage4X174DNA (23).
hence, progeny virusesbearingan aberrant copy number of DR2 wererarelygenerated(Fig. 6a). When copy numbers of DR2 of two DR2 arrays were similar (e.g., CL27), the unpaired stretch (not forming a homologous pair between two a sequences) should have a limited length; hence, progeny viruses differing from the parent virus in copy number of DR2 were rarely generated by recombination between the two DR2 arrays (Fig. 6b). However, a longer stretch was assumed to remain unpaired between two a sequenceslargely differing in the copy number ofDR2(e.g., SIN12, CL36, and SR21), and the recombinations between the unpaired DR2 arrays were likely to generate progeny viruses having a sequences with aberrant copy numbers of DR2(Fig. 6c).
The divergence of a sequences at the end of the S component ofreplicated virus genome suggested the pres-ence of a mechanism mediating the exchange of genetic information between a sequences located at different sites (Fig. 5). The genetic information contained in a sequences neighboring inverted repeats of the L component (RL) would be madenextto theinverted repeats of the S component
(Rs)
by either mechanism (i.e., gene conversion between a se-quences or the presence of a single a sequence at the L-S junction) (Fig. 7). Consequently, the a sequences at the end of the S component could become divergent, though the a sequenceatthe endof the S component is a single copy and would be originally of one kind. Deiss et al. (7) proposed two alternative models for the cleavage-packaging of HSV-1 DNA; one is the directional cleavage model. The other model involves the interaction of two directly repeated junctions, resulting in amplification of the a sequence by a
(b) --- ~ D
1 1 1
Al__ I _%
(C) _IIDR
IT-I
A1---' C'XE
A/,I
''
,B 'C '0
FIG. 6. Recombination between two DR2 arrays. (a) TwoDR2 arrays with thesamecopynumber of DR2canmakeahomologous pair;hence,aDR2 arraywithanaberrant numberofDR2 sequences is rarely generated by recombination. (b) Two DR2 arrays with a similar copy numberofDR2formanunpaired stretch, which isnot
longenoughfor thefrequent generation ofa DR2 array bearingan
aberrant copy number of DR2. Recombinations shown by dotted lines B and C generate DR2 arraysconsisting ofsix and fivecopies
of DR2(thesame asthose of theparental DR2array). Recombina-tionsby the dotted lines A and D generate DR2 arraysconsistingof sevenandfourcopiesof DR2(differingfrom thoseofparental DR2 array), and occurrence of these recombinations is assumed to be
rare. (c) Two DR2 arrayslargely differingin copy number of DR2 should make a long unpaired stretch, leading tothe generationof DR2arrayswithanaberrant copy numberofDR2. Recombinations shown bydotted linesB,C,and D generate DR2 arraysconsisting ofsix, five,andfourcopiesof DR2(differingfrom those ofparental DR2array). Those shownbythe dotted lines A and E generate DR2 arraysconsistingofsevenandthreecopies ofDR2(sameasthoseof parental DR2 array). The model for the gaprepair recombination formingthe loopedstructuremaybe fittedtothegeneration ofthe DR2 array withanaberrant copy number of DR2(10).
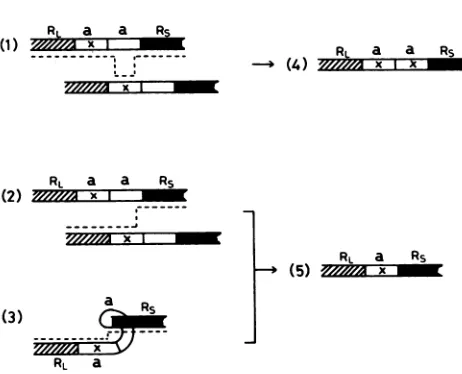
gene conversion-like mechanism, based on the double-strandbreakrepairmechanism first proposed bySzostak et al. (21, 22). The double-strand break and gap repair model foraamplification wasequally favorable toboth of thetwo presumed mechanisms explaining the divergence ofa se-quence attheend of the S component(i.e.,geneconversion orthepresenceofasinglea sequenceatL-Sjunction) (Fig. 7). The results described in the present report support the double-strand break and gap repair model as a mechanism for recombinational events ina sequences (7).
Theshortestasequence(that of TW14)consisted of DR1, Ub, three copies of DR2, DR4t, and Uc (Fig. 4a). The components of the a sequence of TW14 were detected commonly in a sequences from nine other strains (Justin, KOS,17,USA-8,F,K57, K41, B1, andTy154) (6, 13, 15, 32) (Fig. 4), and this presumably represents the basic structure of the a sequence. Of the three kinds of internal repeated arrays (DR2, DR3.5, and DR4), DR2 wasalways present in the a sequences previously examined. The DR2-like se-quences which are found in DR3.5 and DR4 are perhaps involved in the generation and amplification of DR3.5 and
on November 10, 2019 by guest
http://jvi.asm.org/
[image:5.612.347.522.72.305.2] [image:5.612.87.272.76.216.2](1)
-4(4 tXRL a a RS
--44) rzzzzAx Ix~
RL a a RS
a R
(3)
RL a
RL
_5)iIMMA(5 )
[image:6.612.61.292.70.257.2]-FIG. 7. Models forgeneration of
Pathwaysby whichthea sequence
component(Rs) becomedivergent
first, a sequence (marked x) next
component(RL) isdifferentfromthe
Rs(panel 1). Thegene conversion a-sequence
nexttoRsand theasequence (marked
dotted line inpanel 1, generates molecule
sequence next to Rs, as shown in
quencesnext toRsbecomedivergent kindsofasequences, i.e.,theones
2,3,and 5)Aftercrossoverbetween
toRsandthe asequence(marked
line in panel 2) or an intramolecular
neighboring a sequences (as shown
marked x a sequence is made to Rs
yielding a single a sequence at
sequences next toRsbecome divergent.
panel
5 can be amplified (8) by the double-strand
repair mechanismproposed by
DR4, suggesting the recombinogenic
DR2-like sequences. DR4-related
into three types: DR4 (originally
DR4t (a truncated form of DR4, DR2-like
quences), corresponding to nt 133
DR4n, corresponding to nt 142
probably formed from DR4t after
stretch(nt 145 to152 inFig.4b)and
154 to 157 and 171 in Fig. 4b),
strains isolatedinJapanandmay
classifyingHSV-1 strains
ACKNOWLEDGMENTS
IthankM. Oharaforhelpful
Part of this work was supported
Education, Scienceand Culture
REFERENCES
1. Ackermann,M., J. Chou,M.
Roizman.1986.Identificationby
ofaprotein specified byadiploid
repeatsof the Lcomponentofherpes
Virol. 58:843-850.
2. Chou, J., and B. Roizman. 1985.
plexvirus 1genome: identification cis-acting
recoin-binationsites withinthedomainof
811.
3. Chou, J., and B. Roizman.1986.
herpes simplex virus genome contains the promoter ofa gene
located in the repeat sequences ofthe L component. Virol.
57:629-637.
4. Chou, J., and B. Roizman. 1989. Characterization of DNA sequence-common and sequence-specific proteins binding to cis-acting sites for cleavage of the terminal a sequence ofthe
herpes simplex virus 1genome.J. Virol. 63:1059-1068.
5. Chou, J., and B. Roizman. 1990. The herpes simplex virus 1
gene forICP34.5,whichmaps in invertedrepeats,isconserved inseveral limited-passage isolates but not in strain 17syn+. J.
Virol. 64:1014-1020.
6. Davison, A. J.,andN. M.Wilkie.1981.Nucleotidesequencesof
thejointbetween the LandSsegmentsofherpes simplexvirus
types 1 and2.J. Gen. Virol. 55:315-331.
7. Deiss, L. P.,J.Chou, and N. Frenkel. 1986. Functionaldomains
within the a sequence involved in the cleavage-packaging of herpes simplex virus DNA. J. Virol. 59:605-618.
8. Deiss, L. P., and N. Frenkel. 1986. Herpes simplex virus amplicon: cleavageof concatemeric DNA islinkedtopackaging and involves amplification of the terminally reiterated a
se-quence. J.Virol. 57:933-941.
9. Harland, J., and S. M. Brown. 1989. A herpes simplex virus type 2 variant in which a deletion across the L-Sjunction is
replaced by single ormultiple reiterations ofextraneous DNA.
J. Gen. Virol. 70:2121-2137.
10. Jessberger, R., and P. Berg. 1991. Repair of deletions and double-strandgapsbyhomologousrecombination ina
mamma-lian in vitrosystem. Mol. Cell. Biol. 11:445-457.
11. Locker, H., and N. Frenkel. 1979. BamI, KpnI, and Sall restriction enzyme mapsofthe DNAs of herpes simplex virus strains Justin and F.: occurrence ofheterogeneities in defined regions oftheviral DNA. J.Virol. 32:429-441.
12. McGeoch, D. J., M. A. Dalrymple, A. J. Davison, A. Dolan,
M. C. Frame,D.McNab, L. J. Perry, J. E. Scott, and P. Taylor.
1988. The complete DNAsequenceof the long uniqueregionin
the genome of herpes simplex virus type 1. J. Gen. Virol. 69:1531-1574.
13. Mocarski, E. S., and B. Roizman. 1981. Site-specific inversion sequence of the herpes simplex virus genome: domain and
structural features. Proc. Natl. Acad. Sci. USA 78:7047-7051. 14. Mocarski,E.S.,and B.Roizman. 1982. Structureand roleofthe
herpes simplexvirus DNA termini in inversion, circularization and generation of virion DNA. Cell 31:89-97.
15. Mocarski, E. S., L. P. Deiss, and N. Frenkel. 1985. Nucleotide sequence and structural features of a novel Us-a junction present in a defective herpes simplex virus genome. J. Virol.
55:140-146.
16. Mocarski, E.S., L.E. Post, and B.Roizman. 1980. Molecular
engineeringofthe herpes simplex virus genome: insertion ofa
second L-S junction into thegenome causesadditional genome
inversions. Cell 22:243-255.
17. Nasseri,M.,andE.S. Mocarski. 1988.The cleavagerecognition
signal is containedwithinsequencessurroundingana-ajunction
in herpes simplex virus DNA. Virology 167:25-30.
18. Pogue-Geile, K.L.,G. T.-Y. Lee,and P. G.Spear. 1985. Novel rearrangementsof herpes simplex virus DNAsequences
result-ing from duplication ofa sequence within theuniqueregion of
the L component. J. Virol. 53:456-461.
19. Roizman, B. 1979. The structure and isomerization of herpes
simplexvirusgenomes. Cell 16:481-494.
20. Smiley, J. R., J. Duncan, and M. Howes. 1990. Sequence
requirementsforDNArearrangementsinducedby theterminal repeat of herpes simplex virus type 1 KOS DNA. J. Virol. 64:5036-5050.
21. Szostak, J. W.,T. L. Orr-Weaver, R. J. Rothstein, and F. W.
Stahl. 1983. Thedouble-strand-break repairmodel for
recombi-nation. Cell 33:25-35.
22. Takahashi,
I.
N.,and Kobayashi. 1990. Evidencefor thedouble-strand break repair model ofbacteriophage X recombination.
Proc.
Natl.
Acad. Sci. USA 87:2790-2794.23. Umene,K. 1985. Variability of theregionof theherpes simplex
virus type 1 genome yielding defective DNA: Smai fragment polymorphism. Intervirology 23:131-139.
on November 10, 2019 by guest
http://jvi.asm.org/
24. Umene, K. 1985. Intermolecular recombination of the herpes simplex virustype1genomeanalysed usingtwostrainsdiffering in restriction enzyme cleavage sites. J. Gen. Virol.
66:2659-2670.
25. Umene, K. 1987. Restriction endonucleases recognizing DNA sequencesof four base pairs facilitatedifferentiation of herpes
simplex virustype 1 strains. Arch. Virol. 97:197-214. 26. Umene,K.1987. Transition fromaheterozygous toa
homozy-gousstateofapair ofloci in the invertedrepeatsequencesof the
L component of the herpes simplex virus type 1 genome. J.
Virol. 61:1187-1192.
27. Umene, K. 1989. Short, duplicated sequence indicative of the
recombinogenicity of the junction between a unique and an
inverted repeat sequence in the S component of the herpes simplex virustype 1genome. J. Virol. 63:1877-1883.
28. Umene, K., and L. W. Enquist. 1985. Isolation ofnovel herpes
simplex virus type 1 derivatives with tandem duplications of DNA sequences encoding immediate-early mRNA-5 and an
origin ofreplication. J. Virol. 53:607-615.
29. Umene, K., R. J. Watson, and L. W. Enquist. 1984. Tandem
repeated DNA in anintergenic region of herpes simplex virus type 1(Patton).Gene30:33-39.
30. Umene, K., and M. Yoshida. 1989. Reiterated sequences of
herpes simplex virus type 1 (HSV-1) genome can serve as
physical markers for the differentiation of HSV-1 strains. Arch. Virol. 106:281-299.
31. Varmuza,S.L., andJ.R.Smiley. 1984. Unstableheterozygosity
in a diploid region of herpes simplex virus DNA. J. Virol. 49:356-362.
32. Varmuza, S. L., andJ.R.Smiley. 1985. Signalsforsite-specific cleavage of HSV DNA: maturation involves two separate cleavageevents atsitesdistaltotherecognitionsequences.Cell
41:793-802.
33. Vlazny, D. A., A. Kwong, and N. Frenkel. 1982. Site-specific cleavage/packaging of herpes simplex virus DNA andthe
selec-tive maturation of nucleocapsids containing full-length viral DNA. Proc. Natl. Acad.Sci. USA79:1423-1427.
34. Wagner,M.J., and W.C. Summers. 1978. Structure ofthejoint region and the termini of the DNA of herpes simplexvirus type
1.J. Virol. 27:374-387.
35. Weber, P. C., M. D. Challberg, N. J. Nelson, M. Levine, and J.C.Glorioso. 1988. Inversioneventsin theHSV-1genomeare
directly mediated by the viral DNA replication machinery and lacksequence specificity. Cell 54:369-381.
36. Weber, P.C.,M.Levine,andJ. C. Glorioso. 1990. Recombino-genic properties of herpes simplex virustype1DNAsequences
resident in simian virus 40 minichromosomes. J. Virol. 64:300-306.