JOURNAL OF VIROLOGY, June 1992,p. 3833-3845 0022-538X/92/063833-13$02.00/0
Copyright©) 1992, American SocietyforMicrobiology
Functional Diversity
of ElA Gene Autoregulation
among
Human
Adenoviruses
JOY D. COGAN,t
STEPHEN
N.JONES,:
ROBERT K. HALL,§ AND CLARKTIBBETTS*Departmentof
Microbiology
andImmunology, Vanderbilt University Schoolof Medicine, Nashville, Tennessee 37232Received 16 July 1991/Accepted 21 February 1992
Autoregulationof the adenovirus EIAgeneinvolves its constitutive expression and positively and negatively
regulated transcription. Dissection of this process will identify basal-level cis elements and autoregulatory
targetsoftheElApromoterand functional domains within the trans-acting ElAgeneproducts. In thisreport,
the DNAsequenceofthe human subgroupB adenovirustype3 (Ad3)ElAgeneis presented and compared with
that of theEIAgenesof similar and distantly related human adenoviruses. The cDNAforms ofthe Ad3 ElA gene, corresponding to two majorearlymRNA species, are cloned, sequenced, and subcloned into plasmid
expression vectors. Cotransfections ofcell cultures are performed with Ad5 or Ad3 ElA gene expression
plasmidsandareportergeneunder controlof theAd5orAd3ElApromoter.TheAd5 and Ad3 ElApromoters
aresimilarly repressed by either serotype's 12S cDNAgeneproducts.The Ad3 ElApromoterresponds much
morestrongly thantheAdSEIApromoter totransactivationby13ScDNAgeneproducts. Incontrast,the 13S cDNAgeneofAd5 hasgreatertransactivationactivitythanthat ofAd3.Experimentswith missense mutations of the Ad5 ElAgeneindicatethattransactivationof theAd5EIA promoterisweak, just reversingorbalancing
negativeautorepression. Single amino acidsubstitutions in theconserved, repressive functionaldomain 2 of the ElAgenemodulate transactivating activity thatis usuallyassociatedwith the separate and distal conserved
functional domain 3. These results suggestastrongstructure-function relationshipinfluenced bythevariable sequencesseparating theseconserved domains.
Natural variation among similar viruses or strains of virusesprovidesmanyexamplesofevolutionary constraints,
based upon the structure or function of essential gene
products. In a complementary fashion, the immune re-sponses of infected host organisms impose divergent pres-sures oncertain viralgenes.Lateviralgenesareoften cited as examples of these counterpoised forces in viral gene
evolution.Essentialearly viralgenes mayalsobesubjectto complementaryconstraints and divergentpressures.
Theadenovirus EIA earlygene is thefirstcomponent of viralgenomeexpression in the nucleus early after infection.
The intimate associations of ElA gene products with the
cellularmechanics oftranscription regulation (3, 4, 8, 36, 41, 55) create a set of functional constraints on this essential early gene. A role for ElA antigens as targets for cell-mediatedimmunosurveillance(1, 2, 32)maybeanalogousto
the immunological basis for antigenic drift of late-gene capsid antigens.
The levels and the kinetics of ElAgeneexpressionin cells
infected by adenovirus type 5 (AdS) or adenovirus type 3 (Ad3) are remarkably different (51). We attribute this to
evolved differences ofpositive and negative autoregulation of thisessential viralgene's expression (18, 51).Adenovirus ElAgeneexpressioninvolvesautoregulatoryrepressionand transactivation oftranscription (13, 18, 23, 25, 28, 37, 46,
* Correspondingauthor.
t Present address: Department ofPediatrics, Division of
Genet-ics, Vanderbilt University School of Medicine, Nashville, TN
37232.
tPresent address: InstituteforMolecular Genetics, Baylor
Col-legeofMedicine, Houston,TX 77030.
§Present address: Department of Molecular Physiology and
Biophysics,Vanderbilt University School ofMedicine, Nashville, TN 37232.
51). These opposed levels of control are mediated by two
early ElA gene products, distinguished by their 5' donor
sites for the RNAsplicingofacommonElAgenetranscript.
Two functional domains (14, 30, 33) are associated with
enhancer-targeted repression, and these are present in the translation products of both ElA mRNA species. A third functional domain (14, 30, 33) is associated with transacti-vation of transcription, and this domain is found in the translation products of the longer 13S mRNA. The EIA promoter contains elements that represent targets for the regulatoryactivities of the ElAgeneproducts (18), although suchtargets maybeaccessed indirectly, through intermedi-ate cellular factors associated with developing or active
transcription complexes.
The ElAgeneis alsonoted for thevarietyof cellulargenes
whichrespondtoElA-mediatedpositive and negative
regu-lation oftranscription (14, 16, 17, 34, 35,44, 45, 47,48,53). Evolutionary balances of functional constraints and anti-genic pressureson the ElAgene mayhave determined the variation ofmorecomplex propertiesofadenoviruses,such
as tissue tropism and oncogenicity. The variety of such
virus-host cell-host animal interactions and the involvements of the ElAgene are seminal featuresofthelarge family of (human) adenovirusserotypes (42, 57).
The different intensity and kineticsofAdS andAd3 ElA
geneexpression earlyafterproductive infectionare
recapit-ulated in transfections ofthe same host cells with plasmid expression vectors. Our earlier studies (18, 51) were done
withvectors expressingcDNA forms of the AdS ElAgene
or genomic forms of the AdS and Ad3 EIA genes. These
ElA gene constructs are cotransfected into cells with a
reporter gene (chloramphenicol acetyltransferase [CAT]) expressed under control of either the AdS or Ad3 ElA promoter. The ElA promoter of Ad3 responds more to
transactivation by the 13S cDNA of AdS than does the 3833
Vol.66, No. 6
on November 9, 2019 by guest
http://jvi.asm.org/
3834 COGAN ET AL.
homologous AdS ElA promoter. The Ad5 ElA promoter responds more strongly than the Ad3 ElA promoter to
autorepression mediated by the 12S cDNA form of theAdS ElA gene.
In this report, we present the DNA sequence of the Ad3 ElA gene and describe the successful cloning and
expres-sion of its 12S and 13S cDNAs. New plasmid expression
vectors enable cotransfectionswith the ElA cDNAgenesof
both Ad3 and AdS and the CAT reporter gene, expressed under control of the homologous or heterologous Ad3 and
AdS ElA promoters. The repressive activities of the 12S
cDNAgeneproductsof Ad3 andAdSareverysimilar.There
isa significant difference between the activities ofAd3 and
Ad5ElA 13S cDNAgene productsin transactivation of the Ad3 ElApromoter.Experimentswith missense mutationsin
functional domains2 and 3 of theAd5ElAgene address the
remarkably different responses of the AdS and Ad3 ElA promoterstotransactivationbyElA products.The variable region separating conserved functional domains 2 and 3
appearsto be importantin the role of both domains in ElA
trans autoactivation.
MATERIALSAND METHODS
Cells and viruses. HeLa cell cultures were maintained in
110-mm dishes in Dulbecco's modified Eagle's medium (DMEM) supplemented with 10% calf serum, antibiotics (100 U ofpenicillin per ml and 100 ,ug ofstreptomycin per
ml), andanantimycoticagent(1 ,uMp-hydroxybenzoicacid butyl ester). Adenovirus type 3 (strain G.B.)wasoriginally
obtained from the American Type Culture Collection and waspropagatedandpurified by proceduresusedroutinelyin
this laboratory (50, 51).
Enzymes.Restriction endonucleases,T4 DNAligase, and the Klenowand T4 DNApolymeraseswerepurchasedfrom
New England Biolabs. Chemically modified T7 DNA
poly-merase, Sequenase, was purchased from United States Biochemical Co. Enzymes were used with the buffers and
incubation conditionsspecified bythe vendors.
Plasmid expression vectors. The construction and use of
plasmids expressing the CAT gene under the control of adenovirus ElA promoterswere described in earlier publi-cations (18, 51). In the present study, we usedan Ad3 ElA
promoter-CAT plasmid (51)and theAd5/3 chimeric promot-er-CAT plasmid described byJones and Tibbetts (18). The latter CAT expression vector has upstream ElA promoter
sequences derived from Ad5, joined at a common PvuII restriction site (-50 bp) tothe identical transcription initia-tion region of the Ad3 ElA CAT plasmid. Construction of the plasmids expressing the genomic form and the 12S and 13S cDNA forms of theAd5ElAgenewasdescribedearlier (18).
We modifiedouroriginal plasmidvector,whichexpresses
the genomic form of the Ad3 ElAgene (51). This plasmid DNA was first restricted at the unique sites for ClaI and SphlI, followedby blunting andpolishing ofthe noncomple-mentarystickyends with T4 DNApolymeraseand blunt-end ligationwith T4 DNAligase.The modifiedplasmidwasthen cloned by transformation of Eschenichia coli competent cells, screening for ampicillin-resistant colonies with plas-mids about 540bp smaller than theoriginal Ad3 ElAgene
expressionvector. Thesequences deletedin this procedure
remove a remaining portion of the tetracycline resistance
gene of the original pBR322 vector without affecting the expressionof theclonedgenomicform of the Ad3ElAgene
in transfected human cells. Deletion of these plasmid
se-quences removes the
pBR322-derived
BamHI
andHindIII
restriction sites from the Ad3 ElA
expression
vector,pro-moting
theremaining,
Ad3-specific
BamHI
andHindIII
sites tounique
status. This facilitatesourprocedure,
which sub-stitutes cDNA sequences forgenomic
DNA sequences be-tweenthesetworestrictionsites,
asdescribed inafollowing
section.
Genomic
expression
vectors,corresponding
to missensemutations hr3 and hr4 of theAd5 ElAgene, as described in
an earlier
publication
(9),
were obtained fromGary
Glennand Robert Ricciardi of the Wistar Institute. The missense mutations of these mutants debilitate the transactivation
activity
of theAd5ElA(13S) products,
consistent with the loci of the mutations in functional domain 3 of the ElA gene.Expression
vectorscorresponding
to the13S
cDNA ofAd5 ElA mutants
pm936
andpm953,
as described in anearlier
publication
(29),
were obtained from Jim Lillie andMichael Green of Harvard
University.
Themissense muta-tions of these mutants debilitateenhancer-targeted
repres-sion of theAdSElA(12S
and13S) products,
consistentwith the loci of the mutations in functional domain 2 of theElAgene.
DNA
sequencing
of the Ad3ElAgene.The DNA sequence of theleft-end,
1,565-bp
BglII
Kfragment
of the Ad3 genomewas determined from viral DNA
(21, 23, 24,
52)
and from viral DNAfragments
derived from afunctional,
plasmid-cloned Ad3ElAgene
(51).
Avariety
ofpartial
andcomplete
MspI
and MboI restrictionfragments
of viral DNA werecloned into the AccIorBamHI site of
phage
M13mp18
DNA and thensubjected
to routinedideoxynucleotide
sequenceanalysis (43). Sequences
from the left-endregion
ofAd3,
corresponding
to the inverted terminalrepetition
and ElApromoter of
Ad3,
wereobtainedby
Maxam-GilbertchemicalDNA
sequencing
protocols
(31),
asreported
earlier(21,
24,
52).
Theexon-intronjunctions,
corresponding
tothe 12S and 13S ElA mRNAspecies
ofAd3,
have been confirmedby
fluorescenceautomated
sequencing
(38)
of Ad3ElA-specific
cDNAclones,
described in afollowing
section.Infection of cells and
early
viral RNApreparation.
HeLa cell cultures were infected with Ad3 at 1.5 x1010
virionparticles
per 5 x 106cells perplate
(cells
were about 80%confluent).
Mediumwasremovedfrom thecells before viruswasadded in avolume of 0.5 ml per
plate,
using
phosphate-buffered saline with2%calfserum.After 30 minat
37°C,
the virusinoculum fluidwas removed andreplaced
with 10 ml perplate
of DMEM culture mediumcontaining
5% calf serum,antibiotics,
andantimycotic.
Six hours after
infection,
the cultures were washed withphosphate-buffered
salineprior
toextraction of RNAby
the method ofChirgwin
et al.(5).
Theinfected cellswerelysed
by
addition of 0.8 ml of4Mguanidine isothiocyanate-25
mM sodiumacetate(pH 6)-120
mM3-mercaptoethanol
perplate.
Lysates
were forcedthrough 18-gauge
syringe
needles to reduceviscosity,
thenlayered
over a1.2-mlcushion of 5.7MCsCl,
andcentrifuged
at37,000
rpm for 16 h at20°C
withaDuPontAH50rotorand
OTD75
ultracentrifuge
topellet
the RNA. The supernatant wasremoved,
and the total RNApellet
wasresuspended
in 300 ,ul of 10 mM Tris-HCl(pH
7.5)-i
mM EDTA.Polyadenylated
RNAwas isolated from total RNAby
oligo(dT)-cellulose
(Collaborative
Research)
column
chromatography.
Samples
wereapplied
tocolumns in 0.5 MNaCl-10mMTris-HCl(pH
7.5)-i
mMEDTA-0.5% sodiumdodecyl
sulfate. Columns were washed with theapplication
buffer and then with 0.1 M NaCl-10 mMTris-HCI
(pH
7.5)-i
mM EDTA. Thepolyadenylated
RNA fractionwasrecoveredby
elution with 10 mMTris-HCI
(pH
J. VIROL.
on November 9, 2019 by guest
http://jvi.asm.org/
DIVERSE ADENOVIRUS ElA GENE AUTOREGULATION 3835 7.5)-i mM EDTA, and the concentration was estimated
from the A260 measured with a microcell with a Beckman DU70 spectrophotometer. The eluted polyadenylated RNA fractions were ethanol precipitated and resuspended in di-ethylpyrocarbonate-treated water at a concentration of 0.5 ,ug/,ul.
Synthetic DNA oligomers. The DNA sequencing primer (-40, M13 universal forward primer) was obtained from New England Biolabs or United StatesBiochemical Co. We are now preparing oligonucleotide primers for DNA se-quencing in this laboratory with a Du Pont Generator single-column DNA synthesizer with standard phosphoramidite reagents (NEN-Du Pont). The two oligomers used in our Ad3 ElA cDNA cloning protocol were synthesized with a SAM-1 instrument, phosphoramidite reagents, and solvents from BioSearch: oligomer 365,5'-TGATTTAGAGATAGAC GGGCCGGAG-3'; and oligomer 366, 5'-TACTAAGGTCCA AAGGTCCATCCCC-3'. The DNA sequences of these two oligomers flank, on opposite strands, the unique BamHI and HindlIl restriction sites in the 5' and 3' exons of the Ad3 ElA gene.
Ad3 ElA-specificcDNA. First-strand cDNA synthesis was performed with the polyadenylated fraction of RNA isolated early after infection of HeLa cells by Ad3 and the compo-nentsof a cDNA synthesis kit from Promega. Priming of the first-strand cDNA was done with 1 ,ug of oligomer 366 (above) and 2 ,ug of polyadenylated RNA; it was annealed by heating and then cooled in a total volume of 10 ,ul. DNA synthesis proceeded for 60 min at 42°C in a 25-,ul reaction mix containing 50 mM Tris-HCl (pH 8.3), 75 mM KCI, 10 mMMgCl2, 0.5 mM spermidine, 10 mM dithiothreitol, 4 mM sodium
PPi,
1 mM each dATP, dCTP, dGTP, and dTTP, 25 U ofRNasin, and 30 U of avian myeloblastosis virus reverse transcriptase. The reaction mixtures (and products) were frozen and stored at -20°C until the next step of cDNAamplification by polymerase chain reaction (PCR). These
first-strand cDNA products represent the noncoding strand ofthe Ad3 ElA gene from the 3' exon position flanking the
HindIll site (oligomer 366 sequence) towards the 5' exon and
siteofElA transcription initiation.
Amplification and cloning of Ad3 EIA cDNA sequences. Thecomponents of a PerkinElmer-Cetus GeneAmp PCR kit and the vendor's procedures were used, with some
modifi-cations, to synthesize a second-strand cDNA from the
first-strand cDNA of the Ad3 ElA gene (above) and then to amplify the cDNA segments prior to cloning.
First-strand cDNA products (2
RI
of the reaction mix described above) were combined with 10RI
of lOx PCR buffer(0.1 MTris-HCl [pH8.3], 0.5 M KCl, 15 mM MgCl2, 0.1% gelatin), 18 nmol each of dATP, dCTP, dGTP, and dTTP, 100 pmol of oligomer 365, 90 pmol of oligomer 366, 64 ,u of H20, and 2.5 U of Taq DNA polymerase in a final volumeof 100[lI.
Samples were covered with a layer of light mineral oil and amplified in a Perkin Elmer-Cetus thermal cycler. The initial denaturation step was 94°C for 150 s, followed by 45 amplification cycles of three steps: 60 s at 94°C, 120s at 47°C, and 180 s at 72°C. The finalextension step was 10 min at 72°C. The sample was removed from beneath the oil overlay and ethanol precipitated. DNA amplification products were analyzed by agarose gel electro-phoresis.The PCR amplification products, generated as described above with Ad3 early mRNA and cDNA primers 366 and 365,weredigested with BamHI and HindIll attherestriction sitesjust inside theprimer DNAsequences. TheDNAwas ligated to BamHI- and HindIll-restricted pEMBL18 (6)
vector DNA and transformed intoE. coli UT481 (F' bearing) competent cells, using the ampicillin resistance marker for plasmid selection and white colonies to indicate an inter-rupted lacZ gene.
Recombinant phagemid DNA was isolated for DNA se-quencing to identify the intron junctions associated with the 12S and 13S cDNA species and to screen for possible adventitious mutations resulting from the cDNA synthesis or PCR amplification steps prior to cloning. Thefl phage strain IR1 was used as the helper for propagation and packaging of the recombinant phagemid DNA. The universal M13 se-quencing oligomer was used for sese-quencing of the single-stranded phagemid DNA, with a fluorescence automated Du Pont Genesis 2000 DNA sequencer (38). The sequences of the cDNA clones were compared with established sequences ofthe wild-type (wt) Ad3 ElA gene. Eight candidate clones of the 12Sand the 13S cDNAs were analyzed, with only one apparent (missense) mutation found in one 13S cDNA clone. Later subcloning of this mutant's cDNA in an expression vector revealed modestly diminished transactivation of the Ad3ElA-CAT reporter plasmid. The four 12S and four 13S clones each have intron junctions which agree precisely with the junctions reported for the closely related ElA gene of adenovirus type 7 (7).
The cDNA sequences of the eight cDNA candidates were excised from the pEMBL18 vectors by restriction with BamHI and HindIll for substitution into the corresponding BamHI-HindIII region of the modified genomic Ad3 ElA expression plasmid (described above). The restricted plas-midand cDNAfragments were mixed, ligated with T4 DNA ligase, and used to transformE. coli, with selection based on the plasmid ampicillin resistance gene. The desired plasmids wereselected by agarose gel electrophoresis on the basis of reduced sizes compared with the genomic Ad3 EIA expres-sion vector. The expresexpres-sion and activities of the cloned 12S and 13S Ad3ElA genes were evaluated in cotransfections of HeLa cellcultures withElA promoter-CAT gene expression plasmids. The four 12S candidate clones were indistinguish-able incotransfection functional assays (18, 51). Three of the four 13S candidate clones were also indistinguishable from one another in such assays.
Transfections of HeLa cells and analysis of ElA gene expression. HeLa cells were prepared for DNA transfections from cultures at approximately 50 to 70% confluence on plates. Transfections generally followed the procedures of Graham and Van der Eb (11), Gorman et al. (10), and Tibbetts et al. (18, 51) with modifications. Cells were re-moved fromplates bytrypsinization, pooled, and dispensed asequal aliquots into separate tubes for individual transfec-tions insuspension. Each tube of cells (4 ml per transfection in DMEM-5% calf serum) received 200 ,u of a transfection mixture containing 20 ,ug of plasmid DNA (carrier plus expression vector) and 5 ,ug of sheared, denatured salmon sperm DNA inHEPES-buffered phosphate (125 mM NaCl, 5 mMKCl, 0.6 mM Na2HPO4, 5 mM dextrose, 20 mM HEPES
[N-2-hydroxyethylpiperazine-N'-2-ethanesulfonic acid, pH
7.05]). CaCl2 was added to the transfection mixtures at 125 mMforprecipitation with DNA, 30 minprior to transfection. Following 3 h of gentle rotation at 37°Cin the CO2 tissue culture incubator, the transfected cells were mixed with equalvolumes of cold 20% glycerol in DMEM and immedi-atelycentrifuged for 5min at 2,500 rpm. The cell pellets were resuspended in 10 ml of DMEM-10% serum-antibiotics-antimycotic and returned to new culture plates for posttrans-fection incubation at 37°C.
Transfected cells were harvested at various times after VOL. 66,1992
on November 9, 2019 by guest
http://jvi.asm.org/
AATATACCTT CTGTGTGGTG b GACGTGACTT c GAGGTGTGGT TAGTTTTGGG TGAATTTCTG ACTTTGACCG GGTGTCAAAG ACGAGTTCCG GCAAGTCAGT AAGATGGAAT ATTGGCTGCG ATGTGGGAGG TTGAACACGG CGGATGCAAG g AGTCATTTCG TTTACGTGGA TCCTGTGTTT TCAAGAGGCC TCTGCGCTTT TCCAGTGAGA CCGGGASCGA CAGTGCAGCC 1 CTGCTAGCTG GGGGTGGTTG ATGGACTTGC GAGCAGTCCA TTGGACTGTC CA.GGAATAAC ACTGGAATGA n CCACTTTATT TACA< GTAAGT TAAACTGTTG AATGGTAGAT CTGLTGALGG GTTGTTATGA TCCATACCGC CGG&GCTGCC GTATTTAAGT TTATGTTTTT GGTGCCAACA a GGGTTAACGG AGTTATGTTG AAGTAGACAG TGAAAATTCT CGGTTATGCC GGTTTCGATT TTACGTAGGT ACTCTTGAGT GAAA ATGAGA AATACTGGAG TTTCGATCCA CALTGAGGAA ATTGGACATA CATAGGTGGG AGAGGGTTTT AGTAAATGAG TGGA.CATGGC TGTAAATGAG CTAAAAGGGG CAAGTTATTA TTTTCCCACG d CCATTTTCGC AGGGTGGAGT ACCGTGTTTT GTCAGCTGAT GCCAGCGAGA CACCTGCGCT TTTGTGGTAA CCTACGCTGC GCTGTGAATG AACCCCTC AAAAAATTGC CCTCCCAGTG GGAGTAAAAG m T< GTAA,GTCTT TGCTATATGA GAAATTTAAA GGAATAGTGT 0 TTCTTGCGAT TTTTTGTAG
CAMCTTCTCC TGATTCA&CT ACCTCACCTC
TATGC&LGCC CLTTCCTGTG ALGCCThLGC q
AGGACTTGTT GGAGGG 2t;GQ T r
ALTGa s
GTGCC CTGCAGCTGT GTTTATTTAA
v
TGCTGAGTGT TTTATTACTT CTTGGGTGGG 1569 CTGAAATTCA CTGGGAAACG _[:I&CZAL GTAATTTAAA 60 CGGCGCGACC 120 CGGTAAATGT 180 CTTACTGACA 240 e GCGAAAACTA 300 ATTTGCCGAG 360 TCACCTAAAT 420 h CGTCAGGGTA 480 AGAGTTTTCT 540 TCCTGCCACL 600 ATACCCTAAT 660 ACGATCTGTA 720 GGTTTTTTAC 780 CTGLGCLCT 840 CTGATCTGGG 900 ATGLTGaaGa 960 CTGCCAGCGL 1020 GTGAATTTCA 1080 GAATGCA.CTG 1140 AGCTGTTTAA 1200 TCCTGTGTCT 1260 GGCGCCCGCA 1320 CCCTGCTGTG 1380 _.Z_CCCGGLAA 1440
TGTGACGTCA TGTAATAAAA 1500 GACTTGGATA TATAAGTAGG 1560
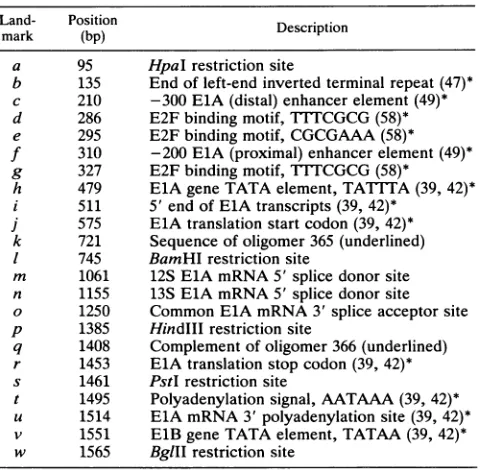
[image:4.612.172.455.55.424.2]FIG. 1. DNA sequence of the Ad3BglIIKfragment,spanningtheElAgene.This sequence hasbeenreportedinU.S.patent4,920,211. Coding regionsareindicatedinboldface. Numbersindicate basepairs alongtheElAgene-encodingrstrand,proceeding from the leftend of the viral genome. Lowercase italic letters above the DNA sequence highlight landmarks, whichare identified in Table 1. Sequences correspondingtooligomers 365 and366,usedin cDNAcloningprocedures(seeMaterials andMethods), areunderlined.
TABLE 1. Landmarksin the DNA sequenceof the Ad3ElA gene'
Description
HpaI restriction site
End ofleft-end invertedterminalrepeat (47)*
-300 ElA(distal) enhancerelement (49)*
E2Fbinding motif,T'llCGCG (58)* E2Fbinding motif, CGCGAAA (58)* -200ElA(proximal) enhancer element (49)* E2Fbinding motif, T'TlCGCG (58)* ElAgeneTATAelement,TAT'TTA (39, 42)*
5'end ofElAtranscripts(39, 42)*
ElAtranslationstartcodon(39, 42)* Sequenceofoligomer365 (underlined) BamHIrestriction site
12SElAmRNA5'splice donor site 13SElAmRNA5'splice donor site
Common ElAmRNA3'splice acceptorsite
HindIII restrictionsite
Complementofoligomer 366(underlined) ElAtranslationstopcodon (39, 42)* PstIrestriction site
Polyadenylation signal, AATAAA (39, 42)* ElAmRNA3'polyadenylation site (39, 42)* E1BgeneTATAelement, TATAA (39, 42)* BglII restriction site
transfectionbyscrapingthecells from eachplateinto 1.2 ml
of Versene (0.5 mM EDTA in phosphate-buffered saline).
Thesuspensionswerecentrifugedin 1.5-mlmicrocentrifuge
tubes for approximately 2min at 12,000rpm, and the cells were then resuspended in 150
[lI
of 0.25 M Tris-HCl (pH7.5).Thecellsuspensions weresubjected tothreecycles of
freeze-thawing between -80 and 37°C and then centrifuged in the microcentrifuge for 10 min at about 12,000 rpm. Cellular extractswere assayedforCAT enzyme activity in 100-,u reaction mixes with 80 nmol of acetyl coenzyme A and 2 to 2.5 nmol of
['4C]chloramphenicol,
as describedpreviously (51). One unit of CAT activity corresponds to
acetylation of 1% of substrate (20 nmol) in 15 minat 37°C
underourstandardassayconditions. Sampleextracts(50
pul
of the
100-pul
reactionmix)areroutinely diluted in assaysto provide results in the range of1to50%substrateacetylation. Higher levels ofacetylation are not linear with the concen-tration of CAT in the extract. Activities of CAT in theaFeaturesofAd3ElAgenetopographyareidentifiedaslandmarks which correspond tolowercaseitaliclettersabovetheDNAsequenceinFig. 1. A schematicmapof the Ad3 ElAgeneispresentedinFig.2. Features inferred
by comparisons withpublished results for human Ad7 orotherwork are indicatedby asterisksand referencecitations,respectively.
3836 1 CTATCTATAT AAAGTGCGCG GTGGGAAAAT GACGTAAAAC GGATATGAGG f AATGAGGAAG GGCCGAGTAT TTCCGCGTAC TTTAAACCTG CCTCCGCGCC GGGGTTATC k
TGAT TL TTCC
TGATTCTATG TGTTACCCCA AGCAGCTGAA TGGGGCAACT TGTTTTTAAG AAGAACTATT GTGCTCGCTT GLTGATGAGT CCTGCAAACG p GATAAGCTTG CTGCCAAGGC u TTATGTCAGC w AGCAGATCT Land-mark a b c d e
f
g h i k I m n 0 p q r s t U V w Position (bp) 95 135 210 286 295 310 327 479 511 575 721 745 1061 1155 1250 1385 1408 1453 1461 1495 1514 1551 1565on November 9, 2019 by guest
http://jvi.asm.org/
[image:4.612.67.309.496.731.2]DIVERSE ADENOVIRUS ElA GENE AUTOREGULATION 3837 extract are calculated to takc into account the extent of
extract dilution (if any). Average levels of CAT activity recovcred from transfected cell cultures are presented plus orminus the standard crror of the mean.
RESULTS
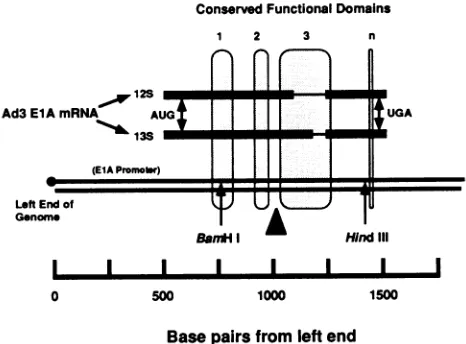
DNA sequence of the Ad3 EIA gene. The DNA sequence of the 1,569-bpBglIl K restriction fragment of human Ad3 is presented in Fig. 1. This figure represents new data and previously published data for the entire length of both complementary strands. Landmarks within the DNA se-quence include the genomic inverted terminal repetition of Ad3 (52), some of the cis clemcnts of the Ad3 EIA promoter (12, 18, 21), and sites associated with transcription and mRNAprocessingofthe EIA gene (7, 54). These landmarks and others are described in Table 1. Figure 2 provides a schematic mapof the Ad3 EIAgenc and its two major early mRNAspccies.
The DNAsequenccof the Ad3 EIA gene is verysimilarto that determined for the closelyrelated subgroup B serotype Ad7 (7). The EIAgcnc-coding regionsof Ad3 and Ad7 differ by only10of 786 bp.Their13S mRNA translation products differby only3 of 262 amino acids, as I for V, V forI, and I for R (single-letter amino acid code). The 12S- and 13S-specific5' splicingdonor sitesand common 3' splice accep-tor sitesofAd3, detcrmined asdescribed later,are identical to thosc specificd byAd7. The 13S intron and the 5' and 3' noncoding DNA sequences of these two viruses differ by only 19 of 779 bp. The enhancer elements (12) and other idcntificd cis elcmcnts oftheirElA promoters are identical DNA sequenccs.
The close relationship of the subgroup B Ad3 and Ad7 EIA genes is reminiscent of that of the EIA genes of subgroup C serotypes Ad2 and Ad5 (54). Aligned DNA sequences ofAd3 and Ad7 or ofAd2 and Ad5 reveal 98 to 99%5 sequcnce identity. Aligned DNA sequences represent-ingthe twodifferent subgroups, suchasAd5and Ad7, show only
52%,c
sequence identity through the EIA gene region. Conscrved features in the EIA regions of subgroup B and subgroup Cadenoviruses includeaninverted terminalrepeat regionand left-end origin of DNAreplication,transcriptional enhancer elements at -200 and -300bp,aTATA element of the EIA promoter as TATTTA, 40 bp of nearly identical DNA sequcncc about the EIA5' RNA cap site, conserved functional domains 1,2, and 3 of the EIAcodingsequences, and anuclcarhoming signal atthecarboxyterminus of EIAproteins.
We and others (14, 18, 25, 54) have observed differences amongfunctional components of subgroup B and subgroup C EIA genes. Thcsc distinguishing features include the number and arrangement of E2F binding sites in ElA promoters, diffcrent 12S EIA splicc donor sites, and DNA sequence and separationof conscrved functional domains 2 and 3. A schematic comparison of the Ad5 and Ad3 EIA gene structures, similar to the format of Fig. 2, was pre-sented in ourprevious report (Fig. 1 ofrefercnce 18).
Cloning andexpression of Ad3ElA-specific cDNA. Diffcr-cnt levcls ofAd5 and Ad3 EIA genc expression early after infcctionwith wtand mutantviruses,have beenattributcdto diffcrent responses of their EIA promotcrs and also to different activitics of EIA gene products (18, 25, 51). Our rccent workwith cDNA exprcssion cloncs of the Ad5 EIA gcnelocalizcd dcterminantsofthesedifferentautoregulatory
rcsponses in the upstrcam DNA scquences of the Ad5 and Ad3EIA promotcrs. Inordertoanalyzepossible differences
ConservedFunctional Domains
..12S
12SAd3 ElA mRNA AUGS
(EIA 13S
_ (EIPrombr
Left End of Genome
i
3 n
ESHIE I PE.-1P!I tUGA .iI.ISISS
BfmHI HIndII1
I I
I
I
I I 0-I
I
0 500 1000 1500
Basepairs fromleftend
FIG. 2. Map of the left end of the Ad3 viral genome, including the EIA gene and its 12S and 13S mRNA species. The thick horizontal lines above the double-stranded DNA genome (left-end segment) represent thetwomajor early mRNA species of the ElA gene. The thin lines in the middle of the mRNAscorrespondtothe introns removed by RNA splicing of the primary EIA transcript. The shaded areas surrounding the DNA and mRNA sequences correspond to the three highly conserved functional domains of adenovirusElAgenes(30-32). The very thinarea nearthe 3' end of the gene, labeled n, corresponds to the conserved pentapeptide region which has been associated with nuclear targeting of ElA proteins from the cytoplasmic site of mRNA translation (44). The unique sites for restriction endonucleasesBamHI and Hindlllare shown in the 5' and3' exons of the gene, respectively. The AUG
startcodon andUGA stop codon of the mRNAspeciesareindicated
by the vertical doublearrows.Thelarge black wedge highlights the nonconservedregionof variable length and DNA sequence between conserved functional domains 2 and 3. This region may be of importance in modulating the negative and positive transcriptional regulation activities associated primarily with the conserved do-mains(25). An alignment of these landmarks of the Ad3ElAgene withcorresponding features of the more familiarAdSEIA gene was presentedasFig. 1 inour recent report(18).
in Ad3 and Ad5 EIAgene productactivities, itwas neces-sary to constructcDNAexpression clones correspondingto the 12Sand 13S mRNAspecies oftheAd3ElA gene.
Several conventional protocols forsynthesis and cloning ofElAcDNAexpression clones wereunsuccessful, in our laboratory andothers. The successful procedure described in Materials and Methods involvedsynthesis ofa segmentof first-strandcDNA withmRNApurified from cells early after Ad3infection, reverse transcriptase, andanoligonucleotide primer complementarytothe 3' exon sequence of Ad3 ElA mRNA. Asecondprimer, representingthe 5'exonsequence of the Ad3 ElA mRNA, Taq DNA polymerase, and an additional quantity of the first primer were used in a PCR procedure to synthesize the second-strand cDNA andthen amplifythe corresponding cDNAsequence.
Ananalysis of thecDNAsynthesis-amplification products
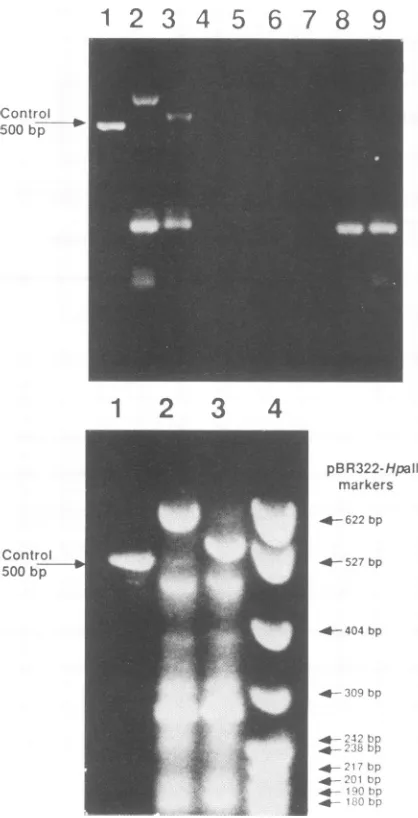
by agarose gel electrophoresis ispresented in Fig. 3. More than90%of the totalproductsrepresentspeciessmallerthan 500bp, and these do not appear torepresent the Ad3 EIA gene. Inspiteofthespecificoligonucleotide primersusedfor
cDNA synthesis and amplification, an identical pattern of
these abundant, smaller products is obtained with mRNA isolated from uninfected HeLa cells. These
spurious,
if perhaps significant, products appearwitha widevariety
of reaction conditions, includingdifferentdurations,
tempera-la!
!
m
I allI I I I I I t
1N
T
of
Vot..66, 1992
on November 9, 2019 by guest
http://jvi.asm.org/
[image:5.612.322.555.78.250.2]3838 COGAN ET AL.
6
7
8 9
Control
500bp
-1
2
3
4
pB R322- Hpa ll markers
-622bp
-4-52 bp
404bp
[image:6.612.78.287.77.486.2]* 3/ hp
FIG. 3.
Analysis
of theproducts
ofcDNAsynthesis
and PCR amplification in 1.8% agarose gels (3 V/cm for 16 h in 0.1 MTris-borate [pH 8.3]-1 mM Na,H2EDTA-0.5 ,ugofethidium
bro-mide per ,ul). Theproducts ofthe cDNA PCR amplification
reac-tions are visualized by ethidium fluorescence on longwave UV
transillumination. (Top) Lane 1 correspondsto the 500-bp control
productDNAprovided with the PCR GeneAmp kit. Lanes2, 3, 8,
and9show PCRproducts amplified fromHeLa cellmRNA
prepa-rationswith oligomers 365 and366, as described inMaterials and
Methods.Lanes 8 and9showproducts frommRNAisolated from
uninfected HeLa cells. Theseextraneous products also appear in
lanes 2 and3,whichwere derived from Ad3-infected HeLa RNA.
Lanes 3and 9 showthesameproductsasinlanes 2 and 8following
digestionwithBamHI andHindIIIrestriction enzymes.Ad3
E1A-specific products, susceptible tocleavage by BamHI andHindIII,
appearasthelargest speciesinlanes 2 and3 and correspondtothe 13S cDNAproduct. A slightly smaller and less abundant species,
the 12ScDNAproduct,is alsopresentbut difficulttoseeunless the
gels are overloaded with products. The empty lanes 4 and 5
representnegativecontrols for mock reactions lacking onlyreverse
transcriptase,andlanes 6 and 7representnegative controlsfor mock
reactionslacking only theoligonucleotide primers.(Bottom) In this
overloaded gel, lanes 1, 2, and3 correspondto lanes 1, 2, and 3 in
the toppanel, respectively. Lane4 shows length markers derived fromapBR322DNAHpaIl restriction digest. ThetwoAd3 E1A-specificcDNAproductsappear as630- and535-bp speciesin lane 2.
The smallerspecies corresponds to 12S Ad3 ElA cDNA product
turesand repetitions of the thermal cycling, and concentra-tionsof monovalent and divalent saltsorTaqDNA polymer-ase.
The products of reactions initiated with mRNA from Ad3-infected cells containtwoadditional species, about630 and 535bp. The ratio ofyields of the larger and the smaller products, estimated by ethidium fluorescence in the gel, is typically about 10:1. The lengthsarethoseexpectedforthe Ad3 ElA cDNA products, predicted from the Ad3 DNA sequence, the specific oligomer primers, and the intron
junctionsdescribed for Ad7 ElA mRNA (7).Treatment with
BamHI and HindlIl reduces the lengths of the two Ad3-specific products by about 75 bp each. The sizes of the nonspecific products do not appear to change following treatmentwiththese restriction enzymes. Apparently, only the two Ad3mRNA-specific products have theBamHIand HindlIl restriction sites associated with the amplified re-gions of the 5' and 3' exons of the Ad3 ElA gene.
The BamHI- and HindlIl-treated cDNA products from Ad3-infected mRNApreparationswereligatedwith BamHI-andHindIII-restricted pEMBL18(6)vectorDNAforcloning of the tentatively identified Ad3 cDNA sequences. Several transformants with inserts of 550 or457 bp were selected. DNA sequence analysis confirmed these to be cDNA frag-ments corresponding to the Ad3 ElA gene 13S and 12S mRNAspecies, respectively. Ad3 ElA 12S and 13S cDNA insertswereexcised from the pEMBL18 vectors and substi-tuted for corresponding genomic DNA sequence between theBamHI and HindIII sites of the modified Ad3 ElA gene expression plasmid (51) (see also Materials and Methods). These newAd3ElAcDNA constructs were thenevaluated for transcriptional regulation activities by cotransfection analysis of transient-gene expression with CAT reporter gene expression plasmids. Ad3 ElA-CAT reporter gene
expressionwasanalyzed incells cotransfected with the 12S
or 13S Ad3ElA cDNAexpression clone (Fig. 4).
Significantlylowerlevels of CAT geneproduct,29% of the
controlvalues,wererecovered from cells cotransfected with the 12S cDNA clone than from cells transfected with the Ad3 ElA-CATplasmid alone. This is consistent with
enhancer-targeted repression of transcription, expected for the 12S
cDNAspecies and theAd3 ElA promoter used for expres-sion of the CAT gene. The activities of four different constructs of the Ad3 ElA 12S cDNA expression plasmid
wereindistinguishable in similar cotransfection assays with the repressible Ad3 ElA-CAT, Ad5 ElA-CAT, and pSV2-CAT(10)reporterplasmids(resultsof thesecomparisonsnot shown).
Transactivation of the Ad3 ElA-CAT plasmid is evident in cotransfections with the Ad3 ElA 13S cDNA expression clone(Fig. 4). Three different constructs of the Ad3ElA 13S
cDNA expression plasmid showed similar transactivation
activities, 20-fold stimulation over controls, in cotransfec-tion assays under these conditions. A fourth 13S cDNA clone hadtwo- tothreefold-lowertransactivation activity in such assays, but itwaslaterdeterminedtohave sustained a single base change
(C1142-
T), a missense mutation (H--Y) near the right end of conserved functional domain 3 (results not shown). This mutation is likely to have been introduced early during the rounds of DNA synthesis andand istypically recovered in amounts lessthan 10%of the larger, 13S-specific cDNA product. Each is reduced by about 75 bp following cleavage byBamHIand HindIII (lane 3).
1
2 3 4 5
J. VIROL.
on November 9, 2019 by guest
http://jvi.asm.org/
DIVERSE ADENOVIRUS ElA GENE AUTOREGULATION 3839
a
-
I-4c
Uc
1000
100
10
Alone +12S cDNA +13S cDNA
FIG. 4. CATgeneexpression in cell cultures cotransfectedwith Ad3 ElA-CAT (10 ,ugperplate) and the 12Sor13ScDNA clone of
the Ad3 ElAgene (3 ,ugperplate). Repressive activity ofthe12S cDNAclone isevident from the lower level of CATgene product
harvested 42 h after cotransfection. The average amount ofCAT product recovered from these cultureswas only 25to30% ofthat recovered from cells transfected by Ad3 ElA-CAT alone. Transac-tivationactivity of the 13S cDNA clone appears aslevels of CAT
product 20 timesgreaterthan in cells transfected by Ad3 ElA-CAT alone. Error bars indicate intervals 3 standard errors above the
averagelevels of CAT (6to15 samplesperaverage).
amplification with avian myeloblastosis virus reverse tran-scriptase andTaq DNApolymerase.
ElAgeneautoregulationwith 12S and13S cDNAclones of Ad5 and Ad3.The availability of 12S and 13S cDNA clones of both theAd5 and Ad3 ElAgenesenablesanalysis ofthe
positive and negative autoregulation ofthetransient
expres-sion ofthesegenesintransfectedcells. Parallel experiments
permit comparisonsof ElApromoterresponsesand effects
of ElA gene products in homologous and heterologous
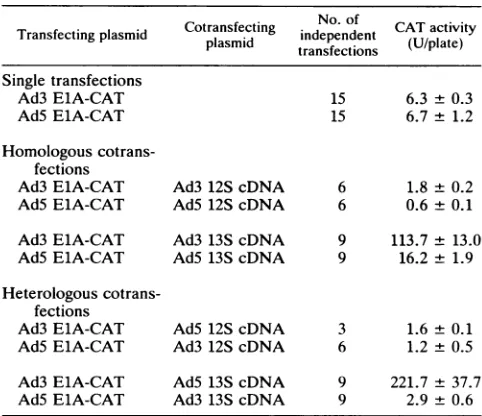
cotransfections. Thedata summarized in Table 2 represent
multiple transfection experiments performed with several different batches of HeLa cells. Each set of transfections
wasperformed with either the Ad5orAd3 ElApromoterfor CAT gene expression, at 10 ,ug of ElA-CAT plasmid per
plate, alone or cotransfected with the 12S or 13S cDNA
clone of the Ad5 orAd3 ElAgene at 3 ,ugperplate. CAT
gene product activity was assayed in extracts from the cultures 40to44 hafter transfection.
Theaverage levelsofCAT fromtransfected cell cultures
presentedinTable 2 havelarger variances than would have been the case if the experiments were performed as
repli-cates with a single batch preparation of HeLa cells for
transfection. Our otherdose-response and time course
ex-periments (Fig. 5 and 6) (reference 18 and unpublished results) indicate that thisparticularsetof conditions-batch ofcells, inputsofreporterplasmid (10 ,ugperdish)andEIA
geneplasmid (3 ,ugperdish), glycerolshocktreatment,and time aftertransfection-typicallyunderestimates the poten-tial levels for ElA autotransactivation. With 10to100 times less ElA (13S cDNAor genomic) plasmid input, transacti-vation often approaches 500-fold-higher expression of the Ad3 ElA-CATgene. Nevertheless,the resultspresentedin Table 2 indicate that the differentresponsesof Ad3 andAd5 ElApromoters are statistically significant (P < 0.05).
Sta-tistically significant differences are also observed for the transactivatingactivities of the Ad3 andAd513SElA cDNA
genes.
Homologous cotransfection experiments, in which the ElA-CAT and the E1A-cDNA clones have the same
pro-moters, generate CAT product which should reflect the levels oftranscription from that ElApromoterin the
trans-fected cells. With this consideration, the data in Table 2 supportthe followingconclusions.
TABLE 2. ElA-CAT gene expression in cells transfected alone orwith homologous and heterologousElA12S
and 13S cDNAclones"
Cotransfecting No. of CATactivity
Transfecting plasmid plasmid Independent (U/plate)
Single transfections
Ad3 ElA-CAT 15 6.3± 0.3
Ad5ElA-CAT 15 6.7± 1.2
Homologous cotrans-fections
Ad3 ElA-CAT Ad3 12S cDNA 6 1.8 ±0.2 AdS ElA-CAT Ad5 12ScDNA 6 0.6 ±0.1 Ad3ElA-CAT Ad3 13S cDNA 9 113.7± 13.0 AdSElA-CAT AdS 13ScDNA 9 16.2 ± 1.9 Heterologous
cotrans-fections
Ad3ElA-CAT AdS 12S cDNA 3 1.6± 0.1 AdS ElA-CAT Ad3 12S cDNA 6 1.2± 0.5 Ad3 ElA-CAT AdS13S cDNA 9 221.7± 37.7 AdSElA-CAT Ad3 13S cDNA 9 2.9 ± 0.6
a HeLa cell cultures were transfected by the procedure described in Materials and Methods. Each culture dish of HeLa cells received 10Vgofthe
indicated ElA-CAT reporter plasmid and, if cotransfected, 3 FLg of the
indicatedElAcDNAexpression plasmid.The number ofindependent trans-fections isspecifiedwith the mean and standard error for CATactivityper
plate,recovered 40 to 44 hafter transfection.
(i) Similar levels of ElA-CAT transcription are supported by the Ad3 and Ad5 ElA promoters in the absence of ElA
cDNAexpression clones.
(ii) Lower levels oftranscription from the Ad3 and Ad5 ElA promoters occur in cells homologously cotransfected with 12S ElA cDNA expression vectors. This autorepres-sion assay led tolevels ofCATproduct whichwere 9 and 29%, respectively, of those expressed by the Ad5 and Ad3 ElA-CAT vectors alone. These data suggest, as reported earlier(18),that theAd5 ElA promoter ismore sensitiveto autorepression than the Ad3EIApromoter.
(iii) The autoactivated expression of CAT from the Ad3 ElA promoter with the homologous 13S cDNA of Ad3 ElA isatleastseventimes greater than the homologous transac-tivation of the AdS ElA promoter with the 13ScDNAofAd5 ElA.
Comparisons of the results ofhomologous and
heterolo-gouscotransfectionexperiments leadtoadditional
observa-tions, includingthe following.
(iv) Transcription of Ad3 and AdS ElA promoters is
repressed by expression of either the Ad3orAd5 12ScDNA gene. The levels ofrepressioneffectedbytheAd3 and Ad5 12S cDNA clonesare notstatistically distinguishable.
(v) The Ad3 ElA promoterresponds muchmorestrongly
to transactivation bythe Ad3 or AdS 13S ElAcDNAthan does the Ad5 ElA promoter, as reported earlier
(18).
The Ad5 13ScDNAgene isatleast two times more effective than the Ad3 13S cDNA cloneas atransactivator of the Ad3 ElA promoter.The responses of the Ad3 and Ad5 ElA promoters to
homologous or heterologous autorepression are
similar,
al-though the Ad5 ElA promoter appears to be
slightly
more sensitiveto repression. Theautorepressive
activities of the Ad3 and Ad5 ElA 12S cDNA genes appear to be verysimilar. VOL.66, 1992
on November 9, 2019 by guest
http://jvi.asm.org/
[image:7.612.92.257.78.181.2] [image:7.612.313.554.106.314.2]3840 COGAN ET AL.
0.4 0.8 1.2 1.6 2.0 2.4 2.8 3.2 0.4 0.8 1.2 1.6 2.0 2.4 2.8 3.2 pg
Co-transfecting
EIAPlasmidyg
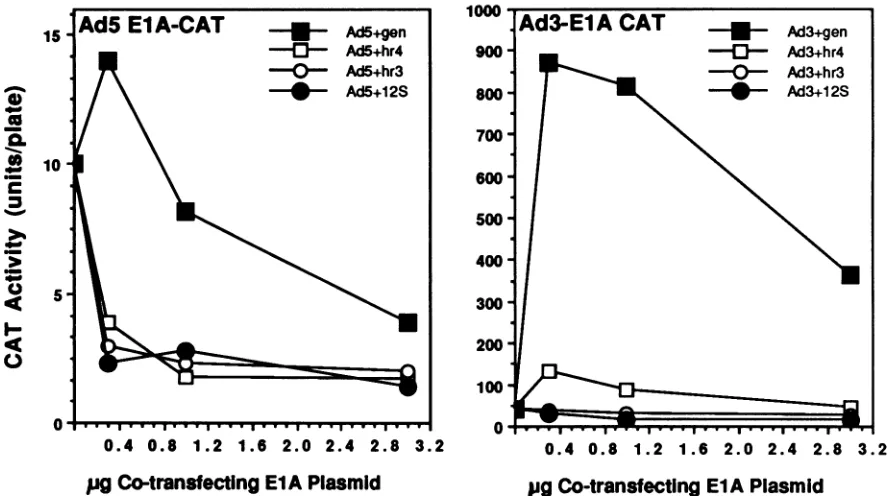
Co-transfecting ElA PlasmidFIG. 5. Effects of Ad5 hr4 and hr3 mutants (domain 3, transactivation)on the cotransfection CATgene expressionfrom Ad3 andAd5 ElA-CATvectors.(Left) Dose-response analysisofcotransfectingAd5 ElAplasmidsforCATgeneexpressionunder control of theAdSElA promoter. Thegenomic (gen) form of thewtAdS ElAgeneisweaklystimulatingtoCATgeneexpressionatverylowinputandrepressive
athigherinputscomparedwithexpression in cells transfected with theAdS ElA-CATplasmidalone. Therepressive12S cDNA ofwtAdS strongly decreases the level ofexpression from the CAT reporter vector, as is also the case in cells transfected with genomic clones representing the domain 3mutantshr4 andhr3.Themutantsappeartoabrogatewhat littletransactivating activityis revealedbythisassay
with the poorly activated AdS ElA promoter. (Right) Corresponding experiments performed with the strongly activated Ad3 ElA-CAT
vector.Thechangein ordinate scale reflects theveryhighlevels of transactivation obtained in cells cotransfected with thegenomicAdS ElA
gene. Again, the hr4 and hr3mutants are severelydebilitated in their capacitytotransactivatethe Ad3 EIApromoter,although the data indicate that the hr4 lesion is somewhat less deleterious.
Indistinctcontrast, the AdS 13SElAcDNAgeneproduct
appears to have about 15-fold-higher transactivation activ-ity, with the transactivation-responsive Ad3 ElA promoter
target, than does the homologous Ad3 13S cDNA gene
product. We draw this conclusionfrom acomparisonof the results from different cotransfections. Lower levels of AdS than Ad3 EIA 13S cDNA gene expression take place in cotransfected cells, as judged from the relative levels of
homologous ElA-CAT expression. Nevertheless,
two-times-higher levels of Ad3 ElA-CAT transactivation are
observed in cells cotransfected with the Ad5 than with the Ad3 13S cDNA gene. The results from different
heterolo-gous and homologous cotransfection experiments, using single batches of HeLa cells and lower inputs of the 13S cDNA expression plasmids, suggest that this difference in Ad5 and Ad3 13S ElA transactivation activities may be
significantly greater than maybe inferred from the data in
Table 2alone.
We acknowledge that alternative explanations of these results canbe framed on the basis of variations in the ElA
protein levels (stability, turnover, translatability,
compart-mentalization) in cells transfected with the various ElA expressionmutants. Lackingreagentsand data pertainingto
the differential stability of variant EIA proteins, we prefer
the plausible assumption that levels of transcription,
re-flected by the relative accumulations of the CAT reporter
gene product, are the primary determinant of variation in
EIA protein levels. A second concern arises in our use of
ElA genes under the control of diffcrent ElA-responsive
ElApromoters. Todate, however, noeukaryoticpromoter which is quantitatively refractile to the positive and/or negative transcriptional regulation effected by ElA gene
products has been described.
PositiveElAgeneautoregulation and conserved functional domain 3. It is plausible that the conserved functional domains of the adenovirus ElAgene, associatedwith posi-tive and negative regulation of the transcription of other early viral and cellular genes (14, 30, 33), act in similar fashion in the transcriptional autoregulation of the ElA
gene. Well-characterized plasmids bearing missense
muta-tions in functional domains 2 and 3 ofthe AdS ElA gene
were generously made available to usto test this
assump-tion. In addition, these new cotransfection experiments provided further insight into the different balances of posi-tive andnegative autoregulation which characterize the ElA
genesofAd5 and Ad3.
Figure 5 presents the results ofcotransfections of HeLa cells with the Ad3 and Ad5 ElA-CAT expression vectors
and EIA genes of the Ad5 mutants hr3 and hr4. These missensemutations, characterized by the work of Glenn and Ricciardi (9), represent single-base changes in functional domain 3 of theAd5ElAgene. They compromise
transac-tivation ofElA-dependent earlypromoters, suchasthose of
the adenovirus E2, E3, and E4genes.The leftpanel of Fig. 5 showsthedose-responsecurvesofAd5 ElA-CATreporter
gene expression with increasing amounts ofcotransfecting ElA expression plasmids. Genomic wt Ad5 ElA provides limited transactivation of the Ad5 ElA promoter, but CAT 15
a.
U
I-U
*0
c 10
51
0
J.VIROL.
on November 9, 2019 by guest
http://jvi.asm.org/
[image:8.612.90.535.74.322.2]DIVERSE ADENOVIRUS ElA GENE AUTOREGULATION 3841
#-M
0.0CD 200-0
:'E
o 100
I-LA0
S 0
0._
U
co
I-U
-000 0.0
1-. 0
25000
*-3
* 1500.
-1000'
Uc
0.4 0.8 1.2 1.6 2.0 2.4 2.8
0.4 0.8 1.2 1.6 2.0 2.4 2.8
yjg Co-transfecting
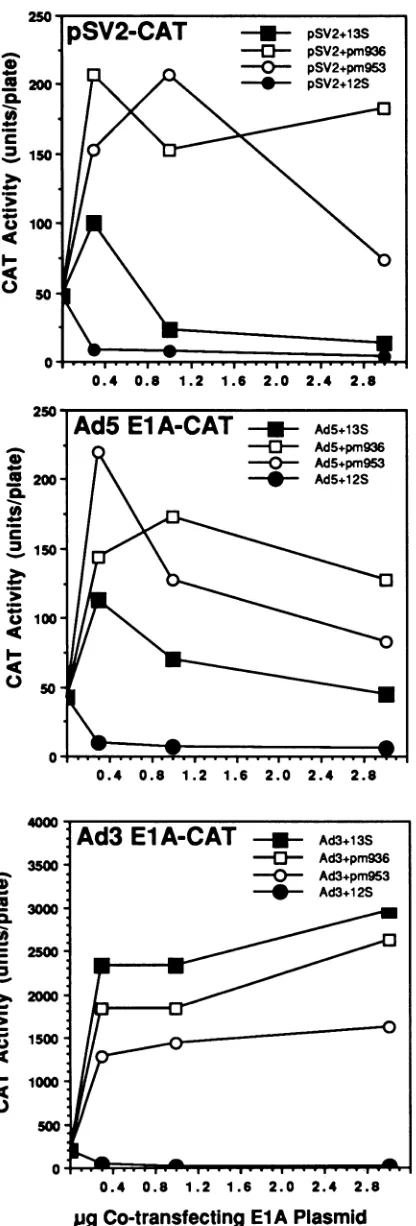
El APlasmidFIG. 6. Effects of theAd5pm936 andpm953mutations(domain2,
repression), ascotransfecting 13S cDNA,on CATgeneexpression
from thepSV2-CATand ElA-CAT vectors. (Top)The 12S cDNA,
and to a lesser extent the 13S cDNA, of the wt Ad5 EIA gene
products do appear at higher levels than in the cotransfec-tions with the repressing wt Ad5 12S cDNA clone. The genomic clones expressing the mutant hr4 and hr3 ElA genes lead torepression levels similar to that of the wt 12S cDNAclone. This is consistentwith the loss of autotrans-activation activity duetothemutationsinfunctionaldomain 3 of the Ad5 ElAgene.
The right panel of Fig.5shows the corresponding analysis performed as cotransfections with the Ad3 ElA-CAT re-porterplasmid.The very different ordinate scalesofthe two panels reflect the much greater response of the Ad3 ElA promoter to transactivation by the wtAd5 EIA gene. The level of Ad3 ElA-CAT expression is much lower in cells cotransfected withthe mutanthr4and hr3ElAgenes. In the caseof the hr4mutant, thisexperiment suggestsdiminished but not entirely abrogated transactivation activity.
Negative EIA gene autoregulation and conserved functional domain 2.Point mutations infunctional domain2oftheAd5 EIA gene shouldcompromise the enhancer-targeted repres-sion activity ofboth 12S and 13S cDNAclones of the Ad5 ElA gene. Such mutations are represented by the Ad5 missensemutantspm936 and pm953, characterized by Lillie andGreen (29). Figure6 presents an analysis ofCATgene expressionfrom three different promoters incotransfections withwt and mutant formsofthe Ad5 EIAgene.
The toppanel of Fig.6shows the resultsobtained with the SV2-CAT (10) reporter gene. The early simian virus 40 (SV40) promoter has an enhancer element (72-bp repeat) which is a target for repression mediated by Ad5 EIA gene products. Thispromoterisnotgenerally recognizedasbeing responsive totransactivation by adenovirus ElA products. Except at the lowest inputs of cotransfecting 13S wt AdS cDNA, both cDNA forms of the wt Ad5 ElAgene repress transcription from the SV2 promoter. In cotransfections with 13S cDNA clones representing mutants pm936 and pm953, there appears to be no repression of the SV2 promoter but rather a modestly elevated level of CAT expression, only twice thatrecovered fromcells transfected with SV2-CAT alone.
Expression of the Ad5 ElA-CATreportergene was eval-uated with the same set of cotransfecting Ad5 ElA genes (Fig. 6, middle panel). The results areremarkably similarto
repressestheenhancer-dependent transcription from the earlySV4t)
promoterin theplasmid pSV2-CAT (41). The dose-responsecurves
revealno repression by the 13ScDNAclonescorrespondingtothe twomissense mutations of the Ad5EIA gene.(Center)Remarkably similar resultsobtained whenAd5ElA-CAT is usedasthereporter.
Again, the 12S cDNA of the wt Ad5 ElA gene represses the
expressioncontrolledby the Ad5EIA promoter. Thisrepressionis compromised by the missense mutations. The transactivation
ex-pected incotransfection with thewtAd5 13S cDNAgene isweak. The overallpatternsofresults with theSV40earlypromoterandthe
Ad5 ElA promoter are remarkably similar, includingthe ordinate
scales of CATactivity. (Bottom)Effects of the AdS EIAdomain 2 lesions on transactivation oftranscription from the Ad3 ElA
pro-moter. In this case, the 13S cDNA clonesof the wt Ad5 and the
mutants are all transactivating the reporter gene through its Ad3
ElApromoter.TheAd5 12ScDNAcontrolrepresses theAd3ElA
promoter. Theplasmidswith eithermutationofdomain2,however,
have lesstransactivation activitythan thewtAd5 13ScDNAgene.
This suggests modulating effects of mutations in one domain on
complementary activities associated with another domain in the sameprotein. Noticeagainthechangeinordinate scalein thispanel
toreflect thesignificantly higherlevelsofCATgeneexpressionfrom the transactivation-responsive Ad3 EIA promoter.
VOL.66, 1992
on November 9, 2019 by guest
http://jvi.asm.org/
[image:9.612.74.282.73.686.2]3842 COGAN ET AL.
thoseobtainedwith SV2-CAT described above. Cotransfec-tion ofAd5 ElA CAT with the wt Ad5 13S cDNA shows onlyslightly greateractivation than cotransfections with the AdS 12S cDNA. The 13S cDNA clones of point mutants pm936 andpm953 donotappeartobecapable of repressing the AdS ElA promoter, but the levels of transactivation effectedby these mutants' ElA genes(as13ScDNA)arenot significantly greater than observed in the SV2-CAT
experi-ment.Theseresultswerealso confirmed inreplicate cotrans-fectionsperformed under the conditions described in Table 2, footnote a (results not shown). The Ad5 ElA promoter thus appears tobe intrinsically weak in its response to 13S ElA-mediated transactivation. Autorepression doesnot ap-pear to be a functional block to an otherwise stronger response to transactivation by the 13ScDNAgenesofAd5 wt, pm936, or pm953.
The lastexperiment of this serieswasdone with theAd3 ElA-CAT reporter gene (Fig.6,bottom).The ordinate scale inthis panel again reflects theintrinsically high response of the Ad3 ElA promoter to activation by the 13S cDNA products of the Ad5 ElA gene. wt Ad5 and the mutants pm936 and pm953 all lead to strong transactivation of the Ad3 ElA promoter when cotransfected as 13S cDNAs. A surprising result in this experiment is the lower level of transactivationactivityassociated with bothmutantforms of the Ad5 13S cDNA. The locus of each mutation is within functional domain 2, associated with enhancer-targeted re-pressionactivity. Apparentlyeven asubtlechange, suchas a missense mutation in one functional domain of the ElA protein,canmodulate other functions associated with
adja-cent ordistal domains.
Theapparently lower transactivation of Ad3 ElA CATby the Ad5pm936 andpm953ElA genes, shownin thebottom panel ofFig. 6, may reflect differences associated with the heterologous Ad3ElA promoteras atarget for transactiva-tion. This isnot particularly surprising, since the Ad5ElA promoter, usedto express the Ad5wtand mutant 13S ElA cDNAgenes inthese experiments, doesnotrespondwellto transactivation.
DISCUSSION
DNA sequencing of similar genes from closely related
organismsis often the firstpointof entry intoproblemareas
associated with prior and continuing evolution of gene structures and functions. Alignments of similar DNA or RNA sequences readily reveal a molecular topography of conserved and nonconserved domains in the gene of inter-est,reflectinga crosssection of variation of that gene within a contemporary population. Establishing consensus se-quences may prompt entertaining speculation about the derivation of contemporary variants from a putative com-mon precursor. The conclusions drawn from such analysis mayevenbe correct, ifthe contemporarypool of variants is derived from a relatively homogenous founder population. In general, however, this type of analysis is suspect, to the extent that one discounts the likely heterogeneity of the gene's precursors and temporal recombining intermediates. A more immediate andproductive application of compar-isons of DNA sequences is correlation of conserved or nonconserved regions of the gene with specific functions of the gene products. Such mapping of functional domains
develops by assay of activities associated with thegene as
derived fromwild-type strains, prevalent allelic variants, or spontaneous and induced mutants. There are risks associ-ated in studiespreoccupied with singularmodels of a
partic-ular gene's structure-function
relationships.
There are alsorisks to be managed with the notion that the most
highly
conserved of variant DNA sequences must be associated with domains of essential function.Structuraland functional dissection of the adenovirusElA gene. Consideration of the adenovirus ElA gene and its
autoregulation,asrevealed from studies of the Ad3 and Ad5
models, leads to several ironies. The ElA promoters of
subgroup B and subgroup C adenoviruses have identical
duplicate EIA enhancer elements
(12, 21, 40, 54).
Sponta-neous or induced deletions of the enhancer
region
cripple
Ad5 EIA gene
expression
and Ad5infections,
but similar deletions in the Ad3 genomeimpair
neither ElA geneexpression nor the course of
productive
infection. Whatdistinguishes these
highly
conserved elements as eitheressential or dispensable for the
expression
of the Ad5 andAd3 ElAgenes?
Dissection of the adenovirus ElA promoter has revealed
multiple arrays of othercis
elements,
inparticular,
knownmotifs for interactions with cellular
transcription
factorssuch as E2F, CREB/ATF, and
enhancer-binding
factors.Brute-force deletions of selected
regions
of the ElA pro-moter canpromoteordinarily
subordinate elementsto moredominant roles in the basal-levelor
autoregulated
expression
of the ElA gene. Thedesign
of assays, the selection ofaparticular model
adenovirus,
and the nature ofspecific
mutageniclesions may confound
interpretation
ofstructure-function relationships. Forexample, separate reports have asserted both DNA sequence
independence
andspecific
DNAsequence determinants involved in
positive
and nega-tiveautoregulation
of theElA promoter(13, 18). Similarly,
conflictingreportshaveappearedwith respecttothe role of
promoter targets of other
ElA-dependent
transcription
units(20,
22, 26,27,
44).
Ad3 EIAgeneDNA sequence. TheDNA sequence of the Ad3 EIA gene,
presented
inFig.
1 and Table1,
is very similar to that of theclosely
related Ad7 ElA gene. Thisanticipated
resultprovides
a limited set of sequence differ-enceswhich may bepredicted
tohave minimalimpact
ontheautoregulated
expression
of the ElAgenes of these similarviruses.Alignmentofthe Ad3ElA gene sequence with that ofAd5 follows the
precedents
established in the literature(14, 30, 33,
54)
and promptsgeneralizations
withregard
tothe coincidence of functional domains
(generally mapped
in Ad2 or AdS) with thehighly
conserved domains of DNA sequences. The DNAsequence of the Ad3 ElA gene sup-portsourselection of this serotypeasarepresentative
oftheB subgroupwith ourpresent
emphasis
on functionaldiver-sityofElA gene
autoregulation
insubgroupBandsubgroup
C adenoviruses.
Diverged patterns of promoter and product roles in EIA
geneautoregulation. The results of
homologous
andheterol-ogous cotransfections presented in this report suggest that extensivefunctionaldiversityhasdeveloped inthe autoreg-ulation of the Ad3 andAd5ElAgenes.Atthesimplest level,
this diversity isreflected bylower expression levels of the
Ad5 ElA gene than of the Ad3 ElA gene (51). Ourrecent report (18) suggested that a primary determinant of this difference intheautoregulated ElAgeneexpressionof Ad3 andAd5 liesin theupstreamDNAsequences,whichspecify
different responses ofElA promotersto 13S ElA-mediated transactivation. The resultspresented in Table 2and
Fig.
6 further support this conclusion. The Ad5 ElA promoter behavesmoreliketheSV40earlypromoter, which isgener-ally
thoughttoberepressed
andnot transactivatedatallby
the action of adenovirusElA gene
products.
The Ad3ElA J. VIROL.on November 9, 2019 by guest
http://jvi.asm.org/
DIVERSE ADENOVIRUS ElA GENE AUTOREGULATION 3843 promoter can be repressed by 12ScDNA products, but it is
remarkable for its strong response to transactivation by 13S EIA gene products.
These results suggest a complementary coevolution of the ElA promoters and their autoregulatingEIAgene products. The poorly transactivated Ad5 ElA promoter supports expression of a 13S cDNA product with greater transacti-vating activity. The opposite situation is found in the Ad3 ElA gene. The strongly transactivated Ad3 ElA promoter supports expression of a 13S cDNA product with lower transactivating activity.
Theperspective on ElA gene autoregulation and comple-mentarypatternsofpromoter-product coevolution provokes interest in the regulated expression of other early genes of Ad3 andAd5.Adenoviral early-gene expression is generally recognized to be ElA dependent. In the case of Ad5, early-gene expression takes place in a milieu of limited quantities of the apparently strong Ad5 13S EIA transacti-vator. Basal-level expression ofAd5 early genes appears to besufficienttoevadeabortive infection (at high multiplicity)
with Ad5 d1312, a mutant which lacks the AdS ElA gene
altogether.
Inthe case of Ad3, the early genes are expressed in the presence of very large quantities of what appears to be a weaker 13S EIA transactivator product. The ElA autoreg-ulation mutant Ad3 hrlS (23, 25) invites comparison with
Ad5d1312. Although Ad3 hrl5 encodes wt Ad3 EIA
prod-ucts, the result is apparent shutdown of further EIA tran-scription. Limited expression of the ElA gene supported by the mutant promoter is lethal; infection is aborted, with minimal cytocidal effects even at very high multiplicity. This mayimply that theexpression of other Ad3 early genes has coevolved to a state of greater dependence on the ElA transactivation function.
Another provocative problem is presented by the differ-ences in ElA product activities and responses of the co-evolved EIAgene promoters. Many host genes of adenovi-rus-infected or -transformed cells respond to positive or negative regulation by adenovirus ElA gene products. The promoters of these cellular genes donot typically resemble the particular arrays of enhancer elements or E2F motifs which characterize ElA promoters. We should anticipate that functional divergence of adenovirus ElA genes, re-vealed in our studies of autoregulation, is likely to be manifested in highly variable regulation of cellular genes, which are not constrained to coevolvewith the adenovirus ElA gene. As suggested in the introduction, such variation may play determinant roles in the diverse tissue tropisms
andpathoepidemiology of humanadenovirus serotypes and
subgroups.
Interdependenceoffunctional domainsand activitiesofEIA proteins. A structural relationship of conserved domains 2 and 3 appears to mutually influence their complementary functions intranscriptional regulation.The resultspresented
in Fig. 6 illuminate this point. The missense mutations in
Ad5pm936
and hr953compromise enhancer-targetedrepres-sionactivity, associated with their loci in conserved domain
2. However, the assayswiththe transactivation-responsive
Ad3 ElA-CAT reporter gene showsignificantlylower trans-activation with these mutant Ad5 ElA genes. The single
amino acidchanges corresponding to their missense muta-tions appear to change the overall structure of the mutant 13S cDNAproducts and thereby compromise (in
quantita-tiveterms) the function of the distal transactivation domain. ThisleadsustoconsiderthattheDNAsequencebetween conserved domains2and3(black wedgeinFig. 2)isaregion
of the EIAgene which is balanced in coevolution with the upstream, transactivation-responsive elements of the
EIA
promoter. This is reminiscent of the structure-function rela-tionshipassociated with mutant Ad3 hrlS and its second-site revertants during passage of viral stocks in this laboratory (23, 25). The level of EIA gene expression in Ad3 hrlS represents effective transcriptional shutdown of the gene, while the same promoter overexpresses the ElA genes which bear reverting deletions in the variable region between conserved functional domains 2 and 3. Perhaps this mutant and its revertants functionally recapitulate aspects of the evolution of ElA gene autoregulation among adenovirus serotypes in the field. These aspects are the apparent reiter-ation and divergence of cis elements in the ElA promoter and deletions (or insertions) in variable regions~separating conserved, functional domains in the ElA gene-coding re-gion.
ACKNOWLEDGMENTS
This investigation was supported by Public Health Service grant CA-34126 from the National Cancer Institute. R.K.H. and S.N.J. were also supported in part by Training Program grant CA 09385 from the National Cancer Institute. Oligomers 365 and 366 were synthesized in the laboratory of R. Stephen Lloyd, Center for MolecularToxicology, Vanderbilt University School of Medicine, supported in part by U.S. Public Health Service grant ES 00267. Fluorescent dideoxyterminators, other sequencing reagents, the Genesis 2000 automated DNA sequencer, and Generator oligonu-cleotidesynthesizer were made available to our laboratory as part of the Collaborative Research Agreement between E. I. du Pont de Nemours & Co., Inc., andVanderbilt University School of Medi-cine. We thank Ed Chait, Richard Baxter, and Paul Levy for the important roles they have played in the initiation and support of that program.
We gratefully acknowledge the Ad5 ElA gene expression plas-mids(missense mutants) provided by Robert Ricciardi of the Wistar Institute and Michael Green ofHarvard University (currently Uni-versity ofMassachusetts School of Medicine, Worcester). We also thank Ricciardi, Green, and Lloyd for thoughtful comments on reading early drafts of themanuscript. During these investigations, expert technical assistance was provided by Nancy Love, Patrick Adams, and SusanGlass.
REFERENCES
1. Ackrill, A. M., and G. E. Blair. 1988. Regulation of major histocompatibility class I gene expression at the level of tran-scription in highly oncogenicadenovirus transformed rat cells. Oncogene3:483-487.
2. Bellgrau,D., T. A.Walker, and J. L. Cook. 1988. Recognition of adenovirus EIA gene products on immortalized cell surfaces by cytotoxic Tlymphocytes. J. Virol. 62:1513-1519.
3. Berk, A. J. 1986. Adenovirus promoters and ElA transactiva-tion. Annu. Rev. Genet. 2:45-79.
4. Borelli, E., R. Hen, and P.Chambon. 1984. Adenovirus-2 ElA products repressenhancer-induced stimulation of transcription. Nature (London) 312:608-612.
5. Chirgwin, J. M., A. E. Przybyla, R. J. MacDonald, and W. J. Rutter. 1979. Isolation of biologically active ribonucleic acid from sources enriched in ribonuclease. Biochemistry 18:5294-5299.
6. Dente, L., G. Cesareni, and R. Cortese. 1983. pEMBL: a new family of singlestranded plasmids. Nucleic Acids Res. 11:1645-1655.
7. Dijkema, R., B. M. M. Dekker, and H. Van Ormondt. 1980. The nucleotide sequence of the transforming BglII-H fragment of adenovirus type 7 DNA. Gene 9:141-156.
8. Flint, J., and T. Shenk. 1989.AdenovirusElAprotein paradigm viraltransactivator. Annu. Rev. Genet. 23:141-161.
9. Glenn, G. M., and R. P. Ricciardi. 1985. Adenovirus 5 early region 1A host range mutantshr3, hr4, andhr5 contain point VOL.66, 1992