0022-538X/86/050493-07$02.00/0
CopyrightC) 1986,American Society forMicrobiology
Persistent Infection of
a
Temperature-Sensitive G31 Vesicular
Stomatitis Virus
Mutant
in
Neural and Nonneural
Cells: Biological
and
Virological Characteristics
JAYASHREE HUPRIKAR,l* STANLEY G. RABINOWITZ,"2MAURO C. DALCANTO,3 AND
MARY KATHLEEN RUNDELL4
Departments of Medicine,' Pathology,3 and Microimmunology,4 Northwestern University Medical School, andVeterans
Administration Lakeside Medical
Center,2 Chicago,
Illinois 60611Received 23 September 1985/Accepted 29January 1986
MouseL-929 cells(L cells), humanoligodendroglioma cells, and ratgliomacellswere persistentlyinfected withvesicular stomatitis virus(VSV)mutanttsG31 and maintained foratleast 4yearsat37°C. Thestriking
observation in thisstudywasthat there is'amarked difference inneurovirulenceamongthepersistentinfections
(PIs) derived from the three cell lines. tsG31VSV derived from persistentlyinfected L cells and oligodendro-glioma cells remained highly virulentasassayed byintracerebral(i.c.)inoculation into 3-week-old Swiss mice. In contrast, tsG31 VSV isolated from glioma cells lost neurovirulence by passage 20. Persistently infected
glioma cells were carried through more than 180 passages without reemergence of neurovirulent virus.
Importantly, gliomaPI virus neurovirulence wasrestoredquickly byi.c. passageinmice andmoreslowly by passagethroughnormal L cells. Incontrast,the neurovirulence of L-cellPI viruswasenhancedbyi.c.passage in mice and slowly reduced by passage through normal glioma cells. Furthermore, no alteration in neurovirulencewasobserved in thecaseofoligodendroglioma PIvirus. Althoughthemechanism(s) underlying the lossof virulence ingliomacells isunclear,ourstudies suggest that either strict temperature sensitivityor
thepresence ofaheat-labiletranscriptaseor bothplaya majorrole in this phenomenon.
Many enveloped RNA viruses can establish long-term
persistent infection (PI) in vivo or in vitro (5, 10, 22, 25).
Persistent infection(PI) withrhabdoviruseshas been studied
ingreatdetailand overalongperiod oftime (6), andmultiple
potential mechanisms have been reported. For example, it
has been reported that alteration of viral proteins and
multiple substitutionsin the RNA appearcontinually during
PI(2, 17, 19). InPIwith wild-type vesicular stomatitisvirus (WT VSV) and rabies virus, the presence of defective
interfering particleshas beenobserved(3, 7, 23).There also
has been phenotypic alteration in RNA synthesis (2) and
base mutations in the ternimal noncoding region (8) with
persistent VSV infection. Production of interferon as a
mechanism ofestablishingPIs has also been reported(26).
We concentrated on establishing PIs in neurally derived
cell lines (glioma and oligodendroglioma) to dissect more
carefully potential pathogenic events underlying our
previ-ous observations about temperature-sensitive (ts) mutants
and host-virus central nervous system interactions (1, 14,
15). PIswere established in neurally derived cells and one
nonneural cell line (L-929 cells) with tsG31 VSV over a
4-year period, and important differences in capacity to
produce neurovirulence were found. It is clearfrom these
observations that every mutant shows different replicative
patternsindifferent cell lines and thatalterationin thevirus
duringthe courseofpersistence leads tounique changes in
its biological characteristics.
In our PIs, with increasing passage, all persistently
in-fected cells continued to produce virus particles, although
gradual decreases were noted in plaque size and titer. In
contrasttovirus derivedfromLcells andoligodendroglioma
cells, the virus fromglioma cells lost its neurovirulence in
* Correspondingauthor.
mice afterpassage 20(about 120days after the initiation of
PI). We now reportvirological characteristics and
biochem-ical behavior of the neurovirulent and avirulent viruses
derivedfrom PIsof these threecelltypes.
MATERIALSANDMETHODS
Cells. BHK-21 cells (Microbiological Associates,
Be-thesda, Md.), mouse L cells (clone 929; Microbiological
Associates),and humanoligodendroglioma cells (generously
supplied by Nelson Levy,DukeUniversity, Durham, N.C.),
ratglioma cells (C-6 clone; a gift from Mary Christensen,
NorthwesternUniversity, Chicago, Ill.), andneuroblastoma
cells(N-18 andN-27clones;agift fromT.Johnson, Kansas
State University, Manhattan, Kans.) were used. Dulbecco
modified Eagle medium supplemented with 7% fetal calf
serum,
gentamicin,
andanti-pleuropneumonia-like
organism(GIBCOLaboratories, Grand Island, N.Y.)wasusedtogrow
and maintain all cell cultures.
Viruses. WT VSV and the tsG31 mutant were originally
obtained from M. E. Reichmann (Urbana, Ill.), plaque
purified twice,and then grownin BHK-21 cells. The titers of
WTVSVpools andtsG31 VSV stockswere109and 5 x 108
PFU/ml, respectively, in a standard BHK-21 cell assay.
BHK-21 cellswere usedfor all plaque assays except when otherwise indicated. Venezuelan equine encephalomyelitis
(VEE) virus was obtained and grown as described
previ-ously(16).
Initiation and maintenance ofPI. PI in C-6glioma, L-929,
oligodendroglioma, and neuroblastoma cells was initiated
and maintained with tsG31 VSV at a low multiplicity of
infection (0.01 to 0.1) at the semipermissive temperature
(37°C) (24). We were unsuccessful in establishing PI with
neuroblastoma cellsaswellas othercell typesnotreported
here(unpublished data),but PIwasreadily establishedin the
493
on November 10, 2019 by guest
http://jvi.asm.org/
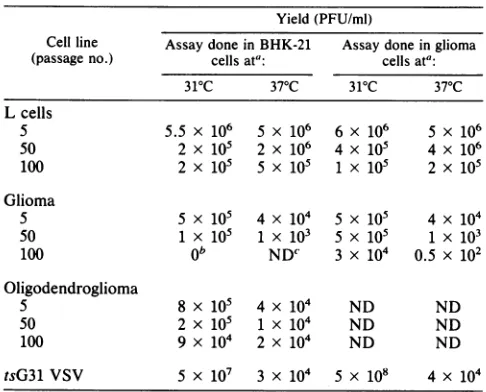
TABLE 1. Plaquing efficiency of virus derived from persistently infected cells
Yield (PFU/ml)
Cell line Assaydone in BHK-21 Assay done in glioma
(passage no.) cells ata: cells ata: 310C 370C 310C 37°C
Lcells
5 5.5x106 5x106 6x106 5x106
50 2 x 105 2 x 106 4 x 105 4x 106
100 2 x 105 5x 105 1x 105 2 x 105
Glioma
5 5x 105 4 x 104 5x 105 4x 104
50 1 X 105 1 X 103 5x 105 1X 103
100 ob NDC 3 x 104 0.5 x 102
Oligodendroglioma
5 8 x105 4x104 ND ND
50 2 x 105 1 X 104 ND ND
100 9 X 104 2 x 104 ND ND
tsG31 VSV 5 x 107 3 x 104 5x 108 4 x 104
aSupernatant media harvested 3 to 4 days after each passage were assayed
at31 and37°C. Plaques were counted after 48 h.
bNo plaques were seen after 48 h of incubation, so incubation was
extended for 24 h. At 72 h, 104plaques were counted.
cND, Not done.
above two neurally derived cells as well as in L-929 cells.
Persistently infected cultures were maintained at37°C
rou-tinelyexceptwhen otherwise indicated.
Plaque assays and growth curvesfor virus isolates fromPIs.
Plaqueassaysandgrowthcurveswith cell-freeculturemedia
collected from each of the PIs were done as described
previously(15).
Virulence studies. Outbred Swiss mice (3 weeks old;
Charles River Breeding Laboratories, Inc.,
Wilmington,
Mass.) wereusedfortheseexperiments.PIisolates(0.03ml)
were serially diluted and inoculated intracerebrally.
Deter-mination of the 50% lethal dose (LD50) for various viral
isolateswas performed asdescribed before (13).
Detection ofinterferon andsuperinfection. Thepresenceof
interferonwas detected asdescribedby others (24).
Super-infection ofPIs withhomologous andheterologous viruswas
performed accordingtopublished work(11). The details of
both proceduresare discussed inthe Results.
RNA labeling and hybridization. Extraction of RNA and
hybridization procedures were carried out as described
previously (2, 18).Inbrief, confluent monolayers ofBHK-21
cells were infected with virus isolated fromPI in the
pres-enceof1
,uag
ofactinomycinD(AmershamCorp.,Arlington
Heights, Ill.)perml and1%fetalcalfseruminmedium. Cells
wereincubatedattheappropriatetemperatures.Atspecified
intervals mRNAwasextracted withaphenol-cresol mixture
(containing 0.1% 8-hydroxyquinoline). Subsequently,
pre-cipitationof the aqueousphasewithethanolwas
performed,
and the precipitates were collected. tsG31 viral RNA was
extracted from purified virus obtained from
[3H]uridine-labeled cultures (25,uCi/ml). Forannealing,0.2 mlofmRNA
wasmixedwith 0.1 ml of labeled viral RNA(approximately
10,000 cpm) in sealedtubes, and the mixturewas incubated
for 18 h in a 70°C water bath. Each sample was then
precipitated with 10% trichloroacetic acid before and after
digestionwith 20 ,ugofpancreatic RNaseA(SigmaChemical
Co., St. Louis, Mo.) per ml at 37°C for 30 min.
RNase-resistanthybridized viral RNAwasdeterminedbycounting
the radioactivity of trichloroacetic acid precipitates in a
scintillationcounter(Beckman LS-7000).
RESULTS
Characterization of virus production. L-929, glioma, and
oligodendroglioma cells which were persistently infected
with tsG31 VSV released virus continually, although with
timethere was a gradual drop in titer from 106 to 104 PFU/ml (Table 1), and plaque size was reduced topinpoint with time. On the otherhand, if the incubation period of the plaque
assay was extended to another 24 h, plaque size increased
dramatically(data not shown). This has been reportedbefore
andexplainedasresulting from attenuation oftheinhibition
of host-cell protein synthesis and, hence, limitation of the
cytocidaleffects of the virus(20). Interestingly,in the case of
glioma PIs, after passage 90 (approximately 650 days) we were unable to plaque virus in BHK-21 cells unless the
incubation period was increased by 24 h, but could easily
plaque the same virus in glioma cells (Table 1).
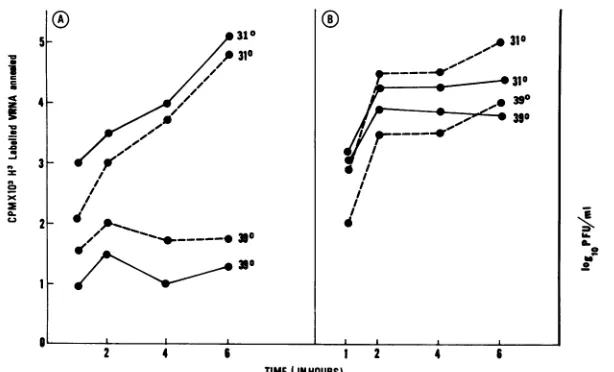
Since all PIs were initiated with the ts mutant, the ts natureof theisolates was assessed (Fig. 1A, B, and C). All PI isolates were ts, although, as compared with L cells,
glioma PIs seem to be more restricted with regard to their temperature sensitivity. For glioma PIs, rates of replication at 37and 39°C were much slower than those of L-cell PIs
(Fig. 1B through G). After passage 90, glioma PIs did not
growin BHK-21 cells at 37 and39°C (Fig. 1D and E),butno change in growth was observed in glioma cells (Fig. 1F and G). This characteristic of glioma PIs suggests that this tsG31 VSVisolate becomesincreasinglyhost restricted as well as ts. Table 1 depicts the plaquing of virus derived from all three cells at the permissive and semipermissive (37°C) temperature. It is notable that virus from L cells plaqued
similarly or even better at37°Cthanat31°C,suggestingthat partial reversion of the tsG31 mutation may be occurring (Table 1). In contrast, virusfromboth neurally derivedcell
linesplaqued at least alog lessat37°Cthanat31°C(Table1).
Thesecharacteristics are alsosupported by growth curves. To assess thetemperature sensitivity further, we shifted the established PIs to thenonpermissive temperature(39°C)
aswell as to thepermissivetemperature (31°C). At39°C we could maintainpersistently infectedLcells for aslongas our 37°C PIs, but we failed to do so with glioma or oligodendro-glioma cells. This newly established PI at 39°C shows characteristics similar topersistentlyinfected L cells at 37°C (unpublished data). At 31°C, owing to the viral replication, cytocidal effect was seen within 48 to 72 h. Interestingly,
gliomaPIs at later stages (afterpassage 90) when shiftedto
31°C could be maintained at 31°C for at least 15 passages
(approximately3months).This was notobserved in the case
of L cells or oligodendroglioma cells at any stage. This
indicates the striking changes that occurred ingliomaPI.
Finally, all PIs were studied at different stages of
persis-tence by means of an indirectfluorescent-antibody technique
with hyperimmune anti-VSV serum. All PIs showed 60 to 70% cells infected asearlyaspassage 4. The number of cells infected did not change significantly with time (data not
shown).
Interferon activity. As reported earlier (24), interferon
production can exert amajor influence in the establishment
of PI. In ourstudies, althoughthe PI cells with serialpassage became increasingly resistant to challenge with a homolo-gous WT VSV and aheterologous virus, VEEvirus (Table
2), we could not detect interferon after acid treatment of supernatant fluids harvested from PIs. When all three PIs were superinfected with homologous virus, it was notable
on November 10, 2019 by guest
http://jvi.asm.org/
Ts G31
390
1 , ,I I
4 6 8 10 12
L- 10
390
4 6 8 10 12
12 4
TIME (hours)
GL-10
310
370
* * 390
4 6 8 10 12
[image:3.612.95.516.71.381.2]FIG. 1. (A through E) Growth curves done in BHK-21 cells. (F and G)Growthcurvesdone in glioma cells. Virus yieldassayswere performedin BHK-21 cells. L, Lcells; GL, glioma cells; 10,passage10; 100, passage 100.
TABLE 2. Superinfection of persistentlyinfected cells with homologous and heterologous virusa
Cell line Uninfected PItiterb Superinfection titer(PFU/ml)
(passageno.) (PFU/ml) WTVSV VEE
Normal Lcells 0 5.3 x 108 5.9 x 108
Lcells
31 2 x 104 2 x 104' 2.5 x 107
54 7 x 104 3.5 x 104c 1 X 107
115 6.5 x 104 8 x 104, 2.5 x 105
Glioma
16 1 x 104 2 x 106 2 x 108
116 od 3 x 104 1x 104
Oligodendroglioma
56 3 x 104 1 X 106 4 x 107
109 9x 103 3 x 104 1 X 103
a Semiconfluent cells were infectedwith 106PFU of either WT VSV or
VEEperflask and incubatedat37'Cfor24 h.Supernatants harvested from
infected and uninfected cultures were assayed in BHK-21 cells at 37TC.
Valuestabulatedherearemean values of fiveindependentexperiments.
bSupernatantsfromuninfectedcontrol orpersistently infectedcells were
assayed at 31°C in BHK-21 cells. Plaques were counted after 48 h of
incubation.
' WT plaques wereidentifiedbyplaquesize.
dNoplaquesweredetectedat 48 h, but 3x 103plaqueswerepresentafter anadditional24-hincubation.
that L cells showed more dramatic inhibition ofgrowth of WTVSVatanearlierperiodthan eithergliomaor
oligoden-droglioma cells (Table 2). When all three PIs were
superinfected with heterologous virus, although there was someinhibition ofgrowthof VEE virusexpressed byLcells at earlierstages, significantinhibition wasobservedatlater stages in all three PIs. This suggests early autointerference and laterproductionof interferon. Resistanceto VEE
repli-cationbypersistently infectedgliomacells does suggest the
presenceof interferonatlaterstages. These studies suggest
a possible role for interferon in PI, but the early loss of
virulence in glioma PI without the demonstration of inter-feron activity suggests that interferon is not an important mechanism indetermining the alteration of virulence inour
PIcultures.
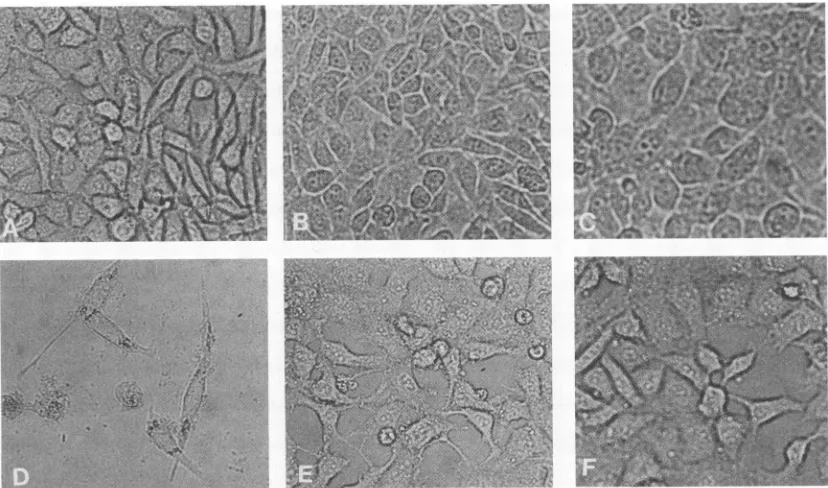
Formoredirectdetectionof thepresenceofinterferon, L cells andoligodendroglioma cells persistently infected with VSV tsG31 weretreated with rabbit anti-mouse interferon
immunoglobulin G and goat anti-human interferon serum,
respectively (both generously supplied by the Antiviral Substances Program, National Institute ofAllergy and In-fectiousDiseases). Becausegliomacellsareratderived, we were unable to detect interferon by directly utilizing anti-interferon serum.The results obtainedwith both L-cell and
oligodendrogliomaPIs(Table 3)showed that thepresenceof anti-interferonantibodyinthe mediumincreased virus titers 20 to 30 times in supernatants harvested 4 days after the treatmentwasbegun.Therewasaclosecorrelation between the increased virus yields observed with anti-interferon 6
5
4
3
2
1
0
on November 10, 2019 by guest
http://jvi.asm.org/
[image:3.612.57.298.490.646.2]TABLE 3. Influenceofanti-interferonantibodyon virus yieldin PIs
Yield 4 days
(passageno.) Cells treatedwitha: aftertreatment Lcells
100 Rabbit anti-mouse interferon
immunoglobulin 4.5 x 107
100 Control serum 1.5 x 106
100 Untreated control 1.2 x 106
Oligodendroglioma
127 Anti-interferon serum 5.5 x 105
127 Control serum 3 X 104
127 Untreated control 1.8 x 104
a Anti-interferon serum (90 units) was added to each culture (T-25 flask), andincubation continued at 37°C. At 1 to 5 days after treatment, supernatant from eachculture was harvested and assayed at 31°C in BHK-21 cells. Peak virus titers were obtained 4 days after the cultures were treated with anti-interferon serum and are reported here.
treatment and significantly increased cell destruction, as
shown in phase-contrast photomicrographs of normal and
persistentlyinfected Lcells(Fig. 2).
RNAsynthesis andhybridization. The evolution of
pheno-typically alteredRNAmutantsinPIisolateswasobservedin
oneofourmodels as otherinvestigators have noted(4, 24,
26). Theselection ofaphenotype(RNA-)was notobserved
in our persistently infected L cells but was consistently observed inglioma cells. Hybridization experimentssuggest
thattherewaseithernoordecreasedRNAsynthesisat39°C
for glioma PIs (Fig. 3A). However, L-cell PIs
synthesized
almostasmuchmRNA at39°Casthey didat31°C(Fig. 3B).
The selection ofthe RNA- phenotype was observedat an
early stage of glioma PIs and seems to be a contributing
factor in lossof neurovirulence sincebothchanges occurred
about the same time. No difference in structural protein
synthesis was observed when assessed by polyacrylamide
gel electrophoresis with [35S]methionine-labeled samples
preparedfromall threePIsasdescribedpreviously (9). The
data for mRNA synthesis confirmed previously described
findings concerning viral growth characteristics of glioma
PIs. In contrast to our observations with L-cell PI, other
investigators (24, 26) did observe RNA- phenotypes in L
cells persistently infected with VSVmutant tsO23.
Virulence in mice. Supernatant media from PIs were
examined for neurovirulence by intracerebral inoculation
into3-week-old Swiss mice. Inthe caseofLcells,virulence
seemed to be increased three to eight times as compared
withthat of parental virus tsG31 VSV, although the time of
deathwassimilartothatwithparentaltsG31 VSV (Table4).
Inoligodendroglioma cells, neurovirulencewascomparable
to that ofthe initial tsG31 mutant. Of major interest is the
fact thatthere was aloss of virulence byglioma cells after
passage 20.Of furthernoteis thefactthat theseresultswere
confirmed with other tsG complementation group mutants
(i.e., tsG22 [group II], tsG41 [group IV] [datanot shown]).
Infectionwith virus fromgliomacells wasinvariably benign
even when doses as large as 2 x
105
PFU were used.Furthermore, after amplification by growth at 31°C in cul-ture, preparations containing 2 x
107
PFU were inoculatedintomice, and no deaths orclinical illnesswere apparent.
Virusreplicationin mouse brains wasobserved in all three
PIs. Maximum yield of virus production wasfoundtobe at
4 days after infection in all three PIs, and the titers were
comparable
(Table
5). When 4-day brain poolsprepared
from glioma PI-infected mice were inoculated into normal
FIG. 2. Anti-interferon treatmentofpersistently infectedLcells with tsG31 VSV. (A, B, C)Normal L-cell control; (D, E, F)Lcells persistently infectedwith tsG31 at passage 100; AandD, effect of anti-interferon treatment onnormal and persistently infectedLcells, respectively;BandE,effectof controlserumtreatment onnormal andpersistently infectedLcells,respectively; CandF, untreatedcell controls.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:4.612.108.522.445.689.2]0310
310
..,/ ---O---* no
2 4 6
310
390
390
1 2 4 6
TIME(INHOURS)
FIG. 3. H3-labeled viral RNA annealedtomRNA fromPIs is shownbysolid lines (cpmx 103), anddotted lines representreplicationof virus(PFU/ml).(A)Hybridization and growthpatternofgliomaPIsfrompassage8to90.(B)HybridizationandgrowthpatternsofL-cell PIs
atevery stageofpersistence.
mice, dramaticregeneration of neurovirulencewasapparent
(Table 5). Regenerated neurovirulence in glioma PI brain
pools wascomparable tothatofthe original tsG31 VSV in
that the LD50 and time ofdeath were similar. In contrast,
L-cell PI showed enhanced neurovirulence and accelerated
death ratecomparable to thatofWT VSV.
Theregeneration of neurovirulence inthe4-day brainpool
prepared from mice infected with virus from glioma cells
suggested additional in vitroexperiments. We nextinfected
normal L-929 cells with established gliomaPI isolates and
normalglioma cellswithestablished L-cellPIisolates. Since
alterations in neurovirulence did not occur in a single
[image:5.612.157.464.71.257.2]pas-sage, wemaintainedthesecross-infected culturesat37°C for
TABLE 4. Lethality of virus harvested frompersistently infected cells
Cellline (passageno.) (PFU/mouse)a
Lcells
21. 6 x 101
55.1.7x 102
118. 8x 102
Glioma
10. 4x 103
15.. >5x 104b
20. >2 x 105b
40.>2 x 105c
100.>2 x 105c
Oligodendroglioma
50.. 5X102
80. 5x102
tsG31. 5 x 102
aSerial dilutions
(10-1
to 10-5) (0.03 ml) ofsupernatant fluid harvestedfromcells were inoculated intracerebrally into Swissoutbred mice (3 to 4
weeks old),and themicewere observed for 21 daysafterinoculation forsigns
of illnessand death. LD5O,NumberofPFU thatgive50%lethality in6 to 8
days.
bNo deaths observed,but20%hind-limbparalysiswasnoted. GliomaPIs
atpassages15, 20, 40,and 100 wereamplified firstat31°C,and isolates were
then inoculated.
cNoclinical symptomswereobserved with this doseofvirus at any time.
aperiod of time. Interestingly, the virulence of virus derived
frompersistently infected L cellswasreducedatleastby 2 logs within a few weeks ofpassage in glioma cells. Con-versely, infectivity of glioma PIs showed sharp enhancement of virulence(Table 6) afterpassagein Lcells. However,the
emergence of neurovirulent virus in these cultures was
extremely slow compared with that of viruspassed through
mousebrain.
DISCUSSION
Establishment ofPI with VSV tsG31mutantsinnonneural (L cells) and neural (human oligodendroglioma and rat
glioma) cellswasachieved withoutgreatdifficultyor
manip-ulation, and all three persistently infected cultures were
maintained forover4 years.
All three PIsshowedagradual decrease in titer and plaque
size. However, onlyin glioma PI did we observe a unique
model of PI in which PI was associated with loss of
neurovirulence.
The presence of endogenous interferon may play an
important role in persistence, but it was less apparent in earlypassages. Thisisparticularly trueforglioma PI. Based
onsuperinfection withaheterologous virus,VEE, glioma PI
showed nearly full sensitivityto VEEinfectionatastagein which the virus had become nearly avirulent. Thus, lack of virulence does notappeartocorrelate with the presenceof
interferon. The probability of glioma PI being a better interferoninducer thanL-cellPIwasexplored byus,but the
datawere notconvincing.
The earliestchangeobservedwasselection ofaphenotype
(RNA-) by virus from glioma PI. At a similar passage
number virus from glioma PI became avirulent and much
moretsforgrowth.
From the results ofourannealing experiments it is clear
that thereisno orverylittle mRNA synthesizedat39°C by glioma PIs. This lack of synthesis of mRNA at 39°C is characteristic ofa complementationgroup I mutant known tobedefectiveintranscriptase activity. It has been reported previously that L cells persistently infected with the ts023 VSVmutant(complementationgroupIII) show consistently rapid selection of RNA-, groupI ts mutants (4, 24, 26). On the other hand, in our system L cells persistently infected with VSV tsG31, another group III mutant, failed
consis-0
4
a
IL
a.
._i
c0
z
at
9
2
.0a
i
e x
2 1,
L3 3
11
on November 10, 2019 by guest
http://jvi.asm.org/
[image:5.612.60.301.474.656.2]TABLE 5. Virus replication in brains of mice infected with virus isolatedfrom persistently infected cells
Titerof LD50of Cellline LDSo of
vPIs
4-daybrain 4-daybrain (passageno.)(PFU/mouse)m
pool pool(PFU/mouseY~ (PFU/ml) (PFU/ml)c
Lcells
15 60 5 x 104 5 x10
30 80 5 x 104 5 x10
67 90 4 x 104
4x10'
Glioma
10 4 x 103 5x 104 5x102
30 >1 x 105 5x 104 5x102
80 >1 x 105 9X 103 9X102
Oligodendroglioma
25 2 x102 3 x104 2x 2
80 2 x 102 3 x 104 2x 102
a These valuesarefor the passage number itself.
bSwissmice (3 weeksold)wereinoculatedintracerebrally witha50%LD_O
of virus isolated from PIs. At reported times the brains were removed asceptically, and brain homogenateswereassayedin BHK-21cellsat31°C. Titers givenaremeansof threeexperiments.
C ForLD50determination serial dilutions of 4-day brainpoolswere
inoculat-edintracerebrally.
tently toselect the RNA- phenotypeandremained virulent
tomiceand leaky in temperature sensitivityaswell as RNA
synthesis.No one has correlated the RNA- phenotype with
the virulent or avirulent nature of the ts mutant, but the
tendencytogenerate revertant mutantsandleakinessat37°C
has been shown to contribute to virulence (21). We are
currently studyingwhetherourPIs develop analterationin
neurotropismin vivo, as reported byothers (12).
When normalcellswere infected with virusisolatedfrom
these three PIs, replication ofvirusat 37°C was at a much
slowerrateforgliomaPIthanfor L-cell and
oligodendrogli-omaPIs(Table 1).The slowerrateofgrowthexperiencedin
thecaseofgliomaPI may in partallow immuneintervention
in vivo to aid in the recovery ofinoculated mice from the
virulent effects.
The emergenceofavirulent virus after several passagesof
avirulent virus ofgliomaPI in L cells(Table 6)suggeststhat virus-host interactions can have pronounced effects in the alteration ofpathogenesis. Toemphasizethecontribution of the host to the pathogenesis of central nervous system
infections by ts mutants, we also established persistently
TABLE 6. Cross-infection of normal cells with virus from PIs
Cell line Titer inglioma h
(passageno.) cells(PFU/ml)' LD5 (PFUIml)
Lcell(100) 2 x 104 8 x 10'
Glioma (100) 3 x 104 >105'
Y-16ld1.4x 105 1.4 x 102
L-lge 1 x 105 1 X 104
"Supernatants harvestedfrompersistentlyinfected cells wereassayedin
glioma cells at 31°C for 48 h. Titers given here are the means of three
experiments.
bLD50,Numberof PFU thatgive 50%lethality.
'Noclinicalsignswereobservedin micefor21daysafterinoculation.
dY-16 wasthe 16th passage of normalgliomacells infected with L-cell PI at
passage 82 and maintained at37°C.
"L-19 wasthe 19th passage ofnormal L cellsinfectedwithgliomaPI at
passage 62 and maintained at37°C.
infected L cells andglioma cells with other ts mutants from complementation groups II (tsG22 VSV) and IV (tsG41 VSV). L cells persistently infected with tsG22 VSV and tsG41 VSV showed evolution ofenhanced virulence when
comparedwiththeirrespective parentalmutants. However,
inglioma PIs with tsG22 VSV ortsG41 VSV virulence was alsodecreased dramatically withinafew weeks of initiation (data not shown). This again supports the observation that virus from L-cell PIs consistently showed enhanced neurovirulence, while virus from glioma PIs tended to lose virulence during persistence.
Wealso documented in this paperthe recoveryof virulent virus in the 4-day brain pools of mice which were infected with avirulent glioma PIs. This reemergence of neuro-virulence was vastly more rapid than recovery in tissue culture. This may be due to the replication of inoculated virus occurring in brain cells other than glial cells which enhances the virulence over a 4-day period. As we have reported earlier (14, 15), virus isolated from the brains of miceinoculated with tsG31wasnolonger ts but still behaved the same in vivo. Our observations of the reemergence of
neurovirulence and the infectivity of brain pools prepared
from L-cell andgliomaPIssuggest that the loci for virulence and the ts phenotype of tsG31 VSV aredistinct.
Fromall these observations weconclude that in the case
ofpersistenttsVSVinfection establishedin ourlaboratory,
in addition to the critical role played by host cells, the
avirulent nature ofglioma PIs in vivo is afunctionof strict
temperature restriction as well as selection of the
RNA-phenotype.
ACKNOWLEDGMENTS
Wewish tothank MaryGrover forhertechnical assistance and Roberta Meyer formanuscript preparation.
This study was supported by Veterans Administration research service project545andby Public HealthServicegrants 2 R01-NS-13011and2R01-NS-13045from the National Institutesof Health.
LITERATURECITED
1. DalCanto, M. C., S. G. Rabinowitz, and T. C. Johnson. 1976. Status spongious resulting fromintracerebralinfection of mice withtemperaturesensitivemutantsof VSV.Br.J. Exp. Pathol. 57:321-330.
2. Frey, T. K.,andJ. S. Youngner. 1982.Novelphenotype ofRNA synthesis expressed by vesicular stomatitisvirusisolated from persistent infections. J. Virol. 44:167-174.
3. Grabau, E.A., andJ. J. Holland. 1982. Analysis of viral and defective interfering nucleocapsids in acute and persistent in-fectionsby rhabdoviruses. J. Gen.Virol. 60:87-97.
4. Holland, J. J., E. A. Grabau, C. L. Jones, and B. L. Semler. 1979.Evaluation ofmultiplegenomemutationsduringlongterm
persistent infection by vesicular stomatitis virus. Cell 16: 495-504.
5. Holland, J. J.,S. I. T.Kennedy, B. L. Semler,C. L.Jones,L.
Roux, and E. A. Grabau. 1980. Defective interfering RNA viruses and thehost-cellresponse. Compr. Virol. 16:137-192. 6. Holland, J. J.,L. P.Villarreal,R. M.Welsh,M.B.A.Oldstone,
D. Kohne, R. Lazzarini, and E. Scolnick. 1976. Long-term persistentvesicularstomatitisvirus andrabiesvirus infectioa of cells in vitro. J.Gen. Virol. 33:193-211.
7. Kawai, A., S. Matsumoto, and K.Tanabe. 1975. Characteriza-tionof rabiesvirus recovered frompersistently infected BHK cells. Virology 67:520-533.
8. Keene, J. D., J. S. Youngner, and J. Wilusz. 1985. Base mutations in theterminal, noncoding regions ofthegenomeof VSV isolated from persistent infections of L-cells. Virology
140:249-256.
9. Laemmli,U.K.1970.Cleavageof structuralproteinsduringthe assembly ofthe head ofbacteriophage T4. Nature (London)
on November 10, 2019 by guest
http://jvi.asm.org/
[image:6.612.61.298.599.659.2]227:680-685.
10. Lehmann-Grube, F., L. Martinez-Peralta, M. Bruns, and J. Lohler. 1983. Persistent infection of mice with lymphocytic choriomeningitis virus. Compr. Virol. 18:43-103.
11. Nishiyama, Y. 1976. Growth of L cells persistentlyinfected with VSV: factors involved in the regulation of persistent infection. Virology 35:265-279.
12. Preble,0. T., L. E. Costello, D. D. Huang, and M.A.Barmada. 1980. Neurovirulence mutantof vesicular stomatitis virus with an altered target cell tropism in vivo. Infect. Immun. 29:744-757.
13. Rabinowitz, S. G., M. C. DalCanto, andT. C.Johnson. 1976. Comparison of central nervous system disease produced by wild-type and temperature-sensitive mutants of vesicular stomatitis virus.Infect. Immun. 13:1242-1249.
14. Rabinowitz, S. G., J. Huprikar, and M. C. DalCanto. 1981.
Comparative neurovirulence of selected vesicular stomatitis virus temperature-sensitive mutantsof complementation groups IIandIII. Infect. Immun. 33:120-125.
15. Rabinowitz, S. G.,T. C. Johnson,and M. C. DatCanto. 1977. Theuncoupled relationship between the temperature-sensitivity andneurovirulence in mice ofmutantsof VSV. J. Gen. Virol. 35:237-249.
16. Rabinowitz, S. G., andR. A. Proctor. 1974. Invitro studies of antiviralactivityof immune spleen cells inexperimental Vene-zuelanequineencephalomyelitis infection in mice. J. Immunol. 112:1070-1077.
17. Rowland, D., E. A. Grabau, K. Spindler, C. Jones, B.Semler, andJ. J. Holland.1980. Virusprotein changesand RNAtermini
alterations evolving during persistent infection. Cell 19:871-880. 18. Saghi, N., and A. Flammand. 1979. Biochemical characteriza-tions oftemperature-sensitive rabies virus mutants. J. Virol. 31:220-230.
19. Semler, B. L., and J. J. Holland. 1979. Persistent vesicular stomatitis virus infection mediates base substitutions in viral RNAtermini. J.Virol. 32:420-428.
20. Stanner, C. P., A. M. Francoeur, and T. Cam. 1977.Analysis of VSV mutants with attenuated cytopathogenicity. Mutation in viral function P or inhibition of protein synthesis. Cell 11:273-281.
21. Stanner, C. P., and V. J. Goldberg. 1975.On the mechanismof neurotropism of vesicular stomatitis virus in newborn hamsters: studies with temperature sensitive mutants. J. Gen. Virol. 29:286-296.
22. Ter Meulen, V., J. R. Stephensson, and H. W. Kreth. 1983. Subacutesclerosing panencephalitis. Compr. Virol. 18:105-159. 23. Villarreal, L. P., andJ. J. Holland. 1976. RNA synthesis in BHK-21 cells persistently infected with vesicular stomatitis virus and rabies virus. J. Gen. Virol. 33:213-224.
24. Youngner, J.S.,E.J. Dubois,D.0. Quagliana,M. Kelly, and 0.T. Preble. 1976. Role oftemperature-sensitive mutants in persistent infection initiated with vesicular stomatitis virus. J. Virol. 19:90-101.
25. Youngner, J. J., and 0. T. Preble. 1980. Viral persistence: evolution of viralpopulations. Compr.Virol. 16:73-125. 26. Youngner, J. S.,0.T.Preble,and E. V.Jones. 1978. Persistent
infections ofLcells with vesicular stomatitis virus: evaluation of viruspopulations.J. Virol. 28:6-13.