0022-538X/92/127533-05$02.00/0
Copyright © 1992, American
Society
forMicrobiology
Mutational Analysis of the Fingers Domain of Human
Immunodeficiency Virus
Type 1
Reverse
Transcriptase
PAUL L. BOYER, ANDREA L. FERRIS, AND STEPHEN H. HUGHES* ABL-BasicResearch Program, NCI-Frederick Cancer Research and
Development
Center,
P.O. BoxB,
Frederick,
Maryland
21702-1201Received 6 April 1992/Accepted 18 August 1992
UsingBspMI cassette vectors, we have constructed a series of mutations in human immunodeficiency virus type 1 (HIV-1) reverse transcriptase (RT) that cause specific amino acid substitutions within the polymerase domain.The RNA-dependent DNA polymerase, DNA-dependent DNA polymerase, and RNase H activities of themutant RTs were assayed. Theelucidation of the structure of HIV-1 RT makes it possible to determine the locationsof specific mutations in the three-dimensional structure of HIV-1 RT[E.Arnold, A.Jacobo-Molina, R.G. Nanni, R. L. Williams, X. Lu, J. Ding, A. D. Clark, Jr., A. Zhang, A. L. Ferris, P. Clark, A. Hizi, and S. H. Hughes, Nature
(London)
357:85-89, 1992; L. A. Kohlstaedt, J. Wang, J. M. Friedman, P. A. Rice, and T.A. Steitz, Science256:1783-1790,1992].Themutations described in this report are between amino acids 25 and 81, within the "fingers" domain of RT (Kohlstaedt et al., Science 256:1783-1790, 1992). It has been suggested that this domain may play a role in positioning the template. Although the fingers domain does not contain the active site for polymerization, several of the mutations within this domain disrupt polymeraseactivitywithout
significantly
affecting RNase Hactivity.Human immunodeficiency virus type-1 (HIV-1) is the
etiological agent of AIDS. HIV-1 virions contain reverse transcriptase (RT), the enzyme that converts the single-stranded viral RNA genomeinto double-stranded
DNA
(31,32). HIV-1 RT is the target of drugs such as AZT
(zidovu-dine) and dideoxyinosine. Despite extensive study, further characterization of HIV-1 RT is necessary for the design of
moreeffective drugs.
In virions, HIV-1 RT exists as a heterodimer with two subunits of 66 and 51 kDa. The 66-kDa subunit contains both the RNA-dependent DNA polymerase domain and the RNaseH domain. The polymerase domain is composed of
approximately the first 430 amino acids in HIV-1 RT; the RNase H domain consists of the 130 amino acids at the
carboxy terminus. The smaller subunit is a proteolytic
cleavage product of the 66-kDa protein or of a larger
precursor and contains approximately the sequences that
comprisethe DNApolymerase domain in thelargersubunit
(4, 6, 9, 17, 20).
Thepolymerase domainsof HIV-1 RT and HIV-2 RTwere
initiallyanalyzedbystudyinginsertion anddeletion mutants
(7-10, 25).Mostof the insertion and deletion mutations that
disruptthepolymerase activityofeitherHIV-1 RTorHIV-2 RT also disrupt the RNase H activity. Similarly, insertion and deletionmutantsinthe RNase H domains of HIV-1 RT and HIV-2 RT often affectpolymerase activity. These data led to the proposal that elements in both domains are required for the proper foldingof each domain (8-10, 25).
There is biochemical evidence which provides support for
this
conjecture.
Hostomsky
et al.(12) expressed
thepoly-merase and RNase Hdomainsseparately. Neitherofthese
separatelyexpressedproteinshad ahighlevelofenzymatic activity.SignificantRNase Hactivitywasrestored when the
two domains were mixed; however, the
activity
of thepolymerasedomainwasnotenhanced(12).Thepolymerase
domain of HIV-1 RT has been further characterized by
* Correspondingauthor.
analyzingpoint mutants.Pointmutantshave beengenerated
bysite-directed mutagenesis(3,15,16, 21,22)orbyisolation from drug-resistant strains of HIV-1 (5, 18, 19, 24, 26, 28,
30). Most of the point mutants created by site-directed
mutagenesis have targeted evolutionarily conserved amino acidresidues(2, 23) and haveidentified anumber of amino acid residues critical for theactivityof HIV-1 RT.Analyses of the RTs from drug-resistant strains of HIV-1 revealed
changes at amino acid positions that arenot evolutionarily conserved.
Wehaveprepareda series ofBspMIcassettes thatallow mutations to be introduced into the polymerase domain of the 66-kDa form of HIV-1 RT(3).Thecassettes are deriva-tives oftheplasmidHIV-1RT, which induces theexpression
of the 66-kDa RT in Escherichia coli (9, 13). We have
previouslydescribeduseof theBspMIcassettes to create20
amino acid substitutions within the polymerase domain
between aminoacids 55 and 186. The 20 mutant RTswere
assayed forRNA-dependentDNApolymeraseand RNaseH
activities. Seventeen of the mutants havevarious levels of RNA-dependent DNA polymerase activityand retain
mod-erate orhighlevels of RNase Hactivity.Twoof the mutants,
Pro-55--+Glyand Ser-156--3Ala, eliminate RNaseHactivity
but retain a significant level of polymerase activity, while
mutant Pro-150--)Glycompletelyabolishes both activities. Wehave made 33 additional amino acidsubstitutions and
rearrangements
within the HIV-1 RT polymerase domain between amino acids 25 and 81. TheRNA-dependent DNApolymerase, DNA-dependentDNApolymerase,and RNase H activity levelswere assayed for all of the mutants. The
recentlypublished structure of HIV-1 RT (14) shows that these mutations lie
within
the"fingers"
domain. It has beensuggestedthat thisdomain hasarole in
binding
thetemplate.
Results and discussion. Thirty-three amino acid substitu-tions and rearrangements were made in thepolymerase
domain of HIV-1 RT between aminoacids 25 and 81(Table
1).Most ofthechangesare atamino acids thatareconserved inawide range ofretroviral RTs
(2,
23).
Themutations areprimarily singleamino acid
substitutions,
but someinvolve 7533on November 9, 2019 by guest
http://jvi.asm.org/
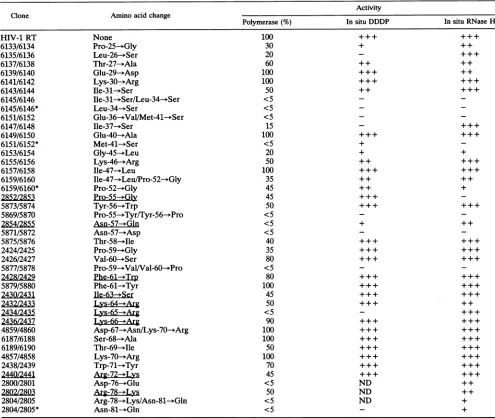
TABLE 1. Summaryofresults ofRNA-dependentDNApolymerase, DNA-dependentDNApolymerase,and RNase Hassaysa
Activity
Clone Aminoacidchange
Polymerase (%) In situ DDDP In situ RNase H
None Pro-25-*Gly Leu-26-->Ser Thr-27-->Ala Glu-29-*Asp Lys-30--+Arg Ile-31--Ser
Ile-31-*Ser/Leu-34--Ser Leu-34--Ser
Glu-36--ValIMet-41--+Ser
Ile-37--3Ser Glu-40--*Ala Met-41--*Ser Gly-45-*Leu Lys-46-*Arg Ile-47- Leu
Ile-47--+Leu/Pro-52-->Gly Pro-52-*Gly
Pro-55--Gly Tyr-56--Trp
Pro-55--Tyr/Tyr-56---Pro Asn-57-*Gln
Asn-57-->Asp Thr-58-*Ile Pro-59--)Gly Val-60-*Ser
Pro-59--Val/Val-60--*Pro
Phe-61-+Trp
Phe-61-->Tyr Ile-63Ser Lys-64--+Arg Lys-65--+Arg Lys-66-*Arg
Asp-67---Asn/Lys-70--3Arg Ser-68--->Ala
Thr-69- Ile Lys-70--*Arg Trp-71--+Tyr
Arg-722±Lys
Asp-76-*Glu Arg-78-ALys
Arg-78-- Lys/Asn-81--+Gln Asn-81--*Gln
100 30 20 60 100 100 50 <5 <5 <5 15 100 <5 20 50 100 35 45 45 50 <5 <5 <5 40 35 80 <5 80 100 45 50 <5 90 100
100
50 100 70 45 <5 50 <5 <5
++
++ ++
++ ++
++
+++
++ +
ND+
+++
++
+++
++
++
++
+++
+++
+++
+++
++ ++
aLocation(s) and the amino acid substitution(s) areshown for eachmutant. Previously describedmutants(3)areunderlined.TheRNA-dependentDNA
polymerase activity of themutantswasassayed inextractsasdescribed in thetext.Theamountof HIV-1 RTpolymerase activityseenwith thewild-type RT
wasset at100%, and the results for themutantRTswerenormalizedtothis value. The in situDNA-dependent DNA polymerase (DDDP)assayandthe in situ RNase Hassay weredone aspreviously described (3, 29). For bothassays,activity is expressedasapproximately equivalenttowild-type activity (+++), moderate(+ +), weak (+),orundetectable(-). ND,notdetermined.
multiple substitutions orchanges in the order of the amino
acids. We have previously described 20 mutations intro-duced into theHIV-1 RT polymerase domain between amino
acids55 and 186 (3). These mutationswere located in four areas designated group I through group IV, based on a
schemeoriginally proposed by Larder etal. (16). Since the structureof HIV-1 RTisnowavailable,wewill discuss the mutations with reference to the subdomains in which they
reside. Thethree-dimensional structureofHIV-1 RT shows that the polymerase domain of the 66-kDa subunit is com-posed of four subdomains that, taken together, resemble a
right hand (1, 14). The active site for polymerase lies in the palm, and the thumb holds the double-stranded primer/
template. The role of the fingers subdomain is less certain; however, it has been suggested that the fingersmayserveto
hold and appropriately position the template strand. The
connection domaintowhich RNaseHis attachedappearsto haveprimarilyastructural role. These four subdomainsare
also present in the 51-kDa subunit; however, they are arranged in a much different configuration. As a
conse-quence, the 51-kDa subunit can play no direct role in the polymerization reaction. Reconstitution studies have shown
that mutations in the 51-kDa subunit do not significantly
affect thepolymerase activity of the 66-kDa subunit (11, 21).
Ahomodimer oftwo 66-kDa subunits alsoappearsto be enzymatically active (27). Although the structure of this homodimeric enzyme has not been solved, it has been suggested that this formshouldresemble the66-kDa/51-kDa heterodimer, with the RNase H domain of the 66-kDa
subunitthat takes the placeof the 51-kDa subunit partially HIV-1 RT
6133/6134
6135/6136
6137/6138
6139/6140 6141/6142
6143/6144 6145/6146
6145/6146* 6151/6152 6147/6148 6149/6150 6151/6152* 6153/6154
6155/6156
6157/6158 6159/6160 6159/6160*
2852/2853
5873/5874 5869/5870
2854/285
5871/5872 5875/5876 2424/2425 2426/2427 5877/5878 2428/2429
5879/5880
2430/2431 2432/2433 2434/2435
2436/2432
4859/4860 6187/6188 6189/6190 4857/4858
2438/2439 244012441 2800/2801 2802/2803
2804/2805 2804/2805*
on November 9, 2019 by guest
http://jvi.asm.org/
cc.
97.4
66.2
kd-42.7 kd
31.0kd _
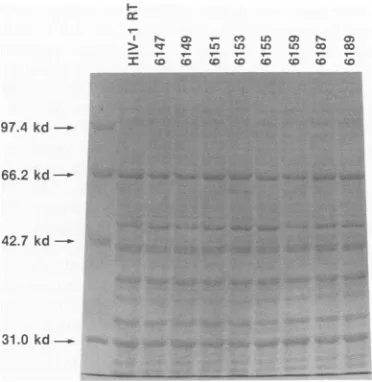
FIG. 1. Level of expression of mutant HIV-1 RTs inE. coli. Bacteriacontaining the parental HIV-1 RT cloneoramutantHIV-1 RTweregrownfor 12to16 hat 37°C with agitation. The bacteria
werecollectedby sedimentation inamicrocentrifuge and thenlysed
in1x Spear's buffer. The proteinextractswerefractionatedon an
SDS-9.0% polyacrylamide gel and visualized by staining with
Coomassie brilliant blue. The arrow on the right indicates the predicted size (66 kDa) of the parental HIV-1 RT protein. The level
of RTexpression is approximately equivalentamongthe different E.
coli strains. Theposition of migration ofsomeof themutantRTs (e.g., 6159/6160) varies slightly from that of wild-type HIV-1 RT.
unfolded and exposed to proteolytic cleavage. We have assumed in our discussions that the primary effect of the
mutations that we have made is exerted from within the
66-kDa subunit that is active inpolymerizationand that the
presenceof the mutation in the other subunit should havea
less significant effect. Although the available data support
thissuggestion,it hasnotbeendirectlytested for each of the
mutantsthatwewilldescribe,and it remainspossiblethatat
least in some cases, the mutation exerts part or all of its effect throughthealteration in the second subunit.
The mutations were constructed by using HIV-1 RT BspMI cassettes (3). Each mutation was validated by dideoxynucleotide sequencing, andthe mutantHIV-1 RTs
werevisualizedonsodiumdodecylsulfate
(SDS)-polyacryl-amidegels.As shown inFig. 1,theamountof RTproduced bythemutantsis similartothe level ofwild-typeHIV-1RT, althoughthepositionofmigrationofsomeof themutantRTs variesslightly.The levels ofRNA-dependentDNA
polymer-ase,DNA-dependentDNApolymerase,and RNase H activ-itiesweredetermined for each of themutantRTs(Table 1).
The resultswereusedtogroupthe amino acid substitutions into four categories: substitutions that are functionally si-lent, substitutions thatprimarilyaffect polymerase activity,
substitutions that primarily affect RNase H activity, and
substitutions that affect both polymerase and RNase H
activities.
Fourteensingleamino acid substitutions and threedouble
amino acid substitutions have been introducedin theregion
between amino acids 25 and 52. The sequence PLTXEKI (aminoacids 25to31)is conservedin lentiviralRTs;there is apartialconservation inawiderrangeof retroviral RTs(2, 23).The amino acids in this motifwerealteredindividually.
The motif containstwohydrophobic aminoacids, aleucine
atposition26 andanisoleucineatposition31.Each of these
was replaced with the uncharged hydrophilic amino acid serine. Mutant HIV-1 RTs with either Leu-26-*Ser or Ile-31-+Ser had diminished polymerase activitybut nearly full RNase H activity. The mutant Leu-26--Ser had
approxi-mately
20%of thewild-typelevel ofRNA-dependent DNApolymerase
activity (Table 1)
butwas inactive in theDNA-dependentDNApolymeraseassay.If thefingersdomainhas
a role in binding of template (14), it is possible that this mutation could selectively affect the enzymes interactions with RNA and DNAtemplates.
Two othermutantsin the PLTXEKImotif, Pro-25--*Gly
andThr-27--+Ala,have diminishedpolymerase activityanda
slight decrease in RNase H activity. We considered the
possibility
that these mutations affect theability
of the enzyme to bind orappropriately position
theprimer/tem-plate. It is believed that the
polymerase
domain and not RNase H hastheprimaryrole inbindingtheprimer/template (1, 14). Since these mutants are reasonably active in the RNase H assay,primer/template
should be bound to thepolymerase domain. The amino acid substitutions
Glu-29--Asp andLys-30--*Arg do not
significantly
affect eitherpolymerase
orRNase Hactivity.
Itisnotclear whether thereplaced
aminoacid isnotimportant
forpolymerase
activity
or whether the
replacement
amino acid is an effective substitute for theoriginal
amino acid. Further mutationalanalysis
shoulddistinguish
between thesepossibilities.
Other
changes
made in conserved amino acids includeGlu-40--+Ala and Ile-47--Leu. These substitutions did not
significantly
affect either RNase Horpolymerase
activity.
The Ile-47--*Leu mutation doesnotsignificantly
change thenature of the
original
side group, and leucine appears to substituteeffectively
for isoleucine at thisposition.
The Glu-40--*Alamutation, however, replaces
thecharged,
acidic side group ofglutamic
acid with thehydrophobic
sidegroupof alanine. The RTsofmousemammarytumor
virus,
Mason-Pfizermonkey virus,
and HIV-1 all have aglutamic
acidatequivalent positions.
Mostof the otherretroviral RTs haveeitherarginine
orlysine.
HIV-2RT also hasalysine
at thisposition. Although
theevolutionary
data suggest thatahydrophilic
amino acid is necessaryatthisposition, putting
alanineatthisposition
in HIV-1 RT hasnomeasurable effect on ourinvitropolymerase
or RNase H assays.The Leu-34--*Ser and Met-41--*Ser mutations
replace
ahydrophobic
amino acid with thehydrophilic
amino acid serine. In both cases, thesubstitutionsignificantly
decreases oreliminates bothpolymerase
and RNaseHactivities. The mutantGly-45--*Leu
hasasimilar effect. When thehydrogen
ofglycine
isreplaced
with thebulky
side group ofleucine,
activities are
significantly
reduced. The Ile-37--+Sermutant has an effect similar to that of Leu-26---Ser. Ile-37--+Ser retains 15%of thewild-type
levelofRNA-dependent
DNApolymerase
activity
(Table 1)
but has no detectableDNA-dependent
DNApolymerase activity.
The mutations
Lys-46--*Arg,
Ile-47--+Leu/Pro-52--
Gly,
and
Pro-52---*Gly
decreasepolymerase
activity
but have different effectsonRNase Hactivity. Lys-46--*Arg
has full RNase Hactivity.
Pro-52--+Gly
retains 45% ofwild-type
polymerase
activity
buthasonly
minimalRNase Hactivity.
We
previously
described anearby
mutant,Pro-55--*Gly,
which also retains45%ofwild-type
polymerase
activity
but has no detectable RNase Hactivity
(3) (Table
1).
Wesuggested previously
that thisregion
of thepolymerase
domain would be in contactwith the RNase Hdomain(3).
However, the structure of HIV-1 RT suggests a different
possibility:
thattheexactpositioning
oftheprimer/template
isaffectedby
thefingers.
Ifthis view is correct, amutationon November 9, 2019 by guest
http://jvi.asm.org/
[image:3.612.88.274.75.266.2]in the fingers domain which still allows properinteractions with the polymerase active site could affectthe position the primer/template and prevent proper interactions with the RNase H active site. Interestingly, the RNase H activity is partially rescued in the double mutant Ile-47--+Leu/Pro-52-*Gly (Table 1).
Twelve single amino acid substitutions and four double amino acid substitutions or rearrangements that have been generated between amino acids 55 and 81 arelisted in Table 1.The prolineatposition 55 and the asparagineatposition 57 arerequiredfor full polymerase and RNase H activities (3) (Table 1). We made a number of amino acid substitutions and rearrangements between amino acids 55 and 60 to characterize this regionmorecompletely. Several retroviral RTs have thearomatic amino acid tyrosine ortryptophan at positions equivalent to position 56 in HIV-1 RT, while the other RTs have either cysteine or glycine at this position. Theclosely related RT of HIV-2 has tyrosineatposition 56. When the tyrosine in HIV-1 RT is replaced with a tryp-tophan(Tyr-56--3Trp), approximatelyhalfof thepolymerase activityremains and RNase H activity isunaffected.
Appar-ently,thelargersidegroupoftryptophancauses adistortion
that decreases the polymerase activity. We tested whether
the order of the amino acids at positions 55 and 56 was important. The mutant Pro-55--+Tyr/Tyr-56- Pro is a rear-rangement of the normal HIV-1 RT amino acid sequence. This mutation eliminates both polymerase and RNase H activities. When the asparagine at position 57 is replaced
with the closely related amino acid glutamine, almost all of
thepolymerase activityis lost and the RNase Hactivity is diminished(3) (Table 1). Both activities areeliminated when asparagine is replacedwith aspartic acid.
Threeothermutationsinthe region from amino acids 55to 60, Thr-58-dle, Pro-59--3-Gly, and Val-60--Ser, alter
con-served amino acids. Most RTs have threonine or serine at
the equivalent position, and HIV-2 RT hasthreonine. Sub-stitutingahydrophobic isoleucine for the hydrophilic
threo-nine doesnotaffect the RNase H activity ofHIV-1 RTbut does decrease the polymerase activity to40% of the wild-typelevel. Theprolineataminoacid59 is highlyconserved. Replacing thisproline withaglycine affects polymerase but notRNaseHactivity. Atpositions equivalenttoposition 60,
mostretroviral RTs have ahydrophobic amino acid, either valine,isoleucine,orleucine. HIV-2 RT has threonineatthis
position. The mutationVal-60--+Ser has onlyaminor effect on the HIV-1 RT polymerase activity. Apparently this positioncan accommodate different amino acidsand is not
crucial to the structure or function of the enzyme. A
rear-rangement mutant was constructed at positions 59 and 60.
Themutation Pro-59--*Val/Val-60--+Proreversestheposition
of the proline at position 59 and the valine at position 60. Substitution of individual amino acids showed that the prolineatposition60isrequiredfor full polymerase activity,
butthevalineatposition 59 isnotrequired (Table1). Amino acidsubstitutionsateitherposition didnotaffect theRNase
Hactivity. Both thepolymerase and RNase H activitiesare
eliminated when the order of the two amino acids is
re-versed.
We have previously described mutations in a motif
(IKKK) atamino acids 63to66(3). We have constructeda
number of additional amino acid substitutions near this
motif. Two of these substitutions are of particular interest
because they have been identified in virus strains that are
resistant tothe drug AZT (5, 15, 18, 30). The single amino acid substitution Lys-70--*Arg and the double substitution
Asp-67--+Asn/Lys-70--+Arg have been linked to AZT
resis-tance. Although the lysine residue at amino acid 70 is conserved in various RTs, including HIV-2 RT, neither the single mutation Lys-70--+Arg nor the double mutation Asp-67--Asn/Lys-70---*Arg reduced the levels of either polymer-ase or RNase H activity in our assays (Table 1). We have made amino acid substitutions at positions 68 and 69, be-tween twoof the positions involved in AZT resistance. Most RTs, including HIV-1 RT, have serine at position 68,
al-though threonine or asparagine is found in some RTs and
HIV-2 RT has a lysine at this position. The mutation Ser-68- Ala haslittleor noeffectonpolymeraseorRNase H activity. Position 68 canapparently accommodate a number ofamino acids besides serine and isprobably notcrucial for the structure orfunction of HIV-1 RT. Most retroviral RTs have aglycine at the position equivalent to amino acid 69 in HIV-1 RT. HIV-1 RT has a threonine at this location, while the RTs from Moloney murine leukemia virus and HIV-2 have an asparagine residue at the equivalent position. The Thr-69-*Ile mutation replaces the hydrophilic threonine with thehydrophobic isoleucine and results in a loss of half of the polymerase activity.
The remaining six substitutions are between amino acids 71 and 81. These amino acids lie near the junction of the fingers and palm domains. At the position, equivalent to amino acid 71 of HIV-1 RT, most RTs have a tryptophan residue. The RTs of Rous sarcomna virus and Moloney murine leukemia virus both have a tyrosine at theequivalent position. The HIV-1 RT mutant Trp-71 -Tyr has a slight decreasein polymerase activity, indicating that a tryptophan residue is preferred to tyrosine at this position. The amino acid residues at positions 72, 76, 78, and 81 are all well conserved in retroviral RTs. The individual mutations Arg-72-*Lys, Asp-76-+Glu, and Arg-78--*Lys each result in significant decreases in polymerase activity, with littleorno measurable effect on the RNase H activity. The Asn-81- Gln mutant shows a significant reduction in both poly-merase and RNase H activities.
This research was sponsored in part by the National Cancer Institute under contract NO1-CO-74101 with ABL and by the National Institute of GeneralMedical Sciences.
We aregrateful to our colleagues, in particular Edward Arnold and Amnon Hizi forhelpful discussions, Marilyn Powers for the oligonucleotides, and HildaMarusiodisforpreparation of the manu-script.
REFERENCES
1. Arnold,E., A.Jacobo-Molina, R. G. Nanni, R. L.Williams, X. Lu, J. Ding, A. D. Clark, Jr., A. Zhang, A. L. Ferris, P. Clark, A. Hizi, and S. H. Hughes. 1992. Structure of HIV-1 reverse transcriptase/DNA complex at 7 A resolution showing active site locations. Nature (London)357:85-89.
2. Barber, A., A. Hizi, J. V. Maizel, Jr., and S. H. Hughes. 1990. HIV-1 reverse transcriptase: structurepredictionsfor the poly-merasedoniain. AIDS Res. Human Retroviruses 6:1061-1072. 3. Boyer, P. L., A. L.Ferris, and S. H. Hughes. 1992. Cassette
mutagenesis of the reverse transcriptase of human
immunode-ficiency virus type 1. J. Virol. 66:1031-1039.
4. DiMarzo Veronese, F., T. D. Copeland, A. L. DeVico, R. Rahman, S. Oroszlan, R. C. Gallo, and M. G. Sarngadharan. 1986. Characterization of highly immunogenic p66/pSi as the reverse transcriptase of
HTLV-III/LAV.
Science231:1289-1291.
5. Gao, Q., Z. Gu, M. A. Parniak, X. Li, and M. A. Wainberg. 1992. In vitro selection of variants of humanimmunodeficiency virus type 1 resistant to3'-azido-3'-deoxythymidine and 2',3'-dideoxyinosine. J. Virol. 66:12-19.
6. Hansen, J., T. Schulze, W. Mellert, and K. Moelling. 1988. Identification and characterization of theHIV-specificRNase H
on November 9, 2019 by guest
http://jvi.asm.org/
by monoclonal antibody. EMBO J. 7:239-243.
7. Hizi, A., A. Barber, and S. H. Hughes. 1989. Effects of small insertions on the RNA-dependent DNA polymerase activity of HIV-1 reverse transcriptase.Virology 170:326-329.
8. Hizi, A., S. H. Hughes, and M. Shaharabany. 1990. Mutational
analysis of the ribonuclease H activity of human
immunodefi-ciencyvirus 1 reverse transcriptase. Virology 175:575-580.
9. Hizi, A., C. McGill, and S. H. Hughes. 1988. Expression of
soluble, enzymatically active, human immunodeficiency virus reverse transcriptase inEscherichia coli and analysis of mu-tants.Proc.Natl. Acad. Sci. USA 85:1218-1222.
10. Hizi, A., R. Tal, and S. H. Hughes. 1991. Mutational analysis of
the DNApolymerase andribonuclease H activities of human
immunodeficiencyvirus type 2 reverse transcriptase expressed inEscherichia coli.Virology180:339-346.
11. Hostomsky, Z., Z. Hostomska, T. B. Fu, and J. Taylor. 1992. Reversetranscriptase of human immunodeficiency virus type 1:
functionality of subunitsof the heterodimer in DNA synthesis. J. Virol. 66:3179-3182.
12. Hostomsky, Z., Z. Hostomska, G.0. Hudson, E. W. Moomaw, and B. R. Nodes. 1991. Reconstitution in vitro of RNase H
activity by using purified N-terminaland C-terminal domains of
human immunodeficiency virus type 1 reverse transcriptase. Proc. Natl. Acad. Sci. USA 88:1148-1152.
13. Hughes, S. H., A. Ferris, and A. Hizi. 1990. Analysis of the reverse transcriptase of human immunodeficiency virus
ex-pressed in Escherichia coli, p. 297-307. In W. G. Laver and
G. M. Air(ed.), Use ofX-raycrystallographyinthedesign of
antiviral agents. Academic Press, Inc., SanDiego, Calif. 14. Kohlstaedt, L. A., J. Wang, J. M. Friedman, P. A. Rice, and
T. A. Steitz.1992. Crystal structure at 3.5Aresolution of HIV-1 reverse transcriptase complexed with an inhibitor. Science 256:1783-1790.
15. Larder, B., S. Kemp, and D. J. M. Purifoy. 1989. Infectious
potential of human immunodeficiency virus type 1 reverse
transcriptase mutants with altered inhibitor sensitivity. Proc. Natl.Acad. Sci. USA86:4803-4807.
16. Larder, B., D. J. M.Purifoy, K. Powell, and G. Darby. 1987.
Site-specific mutagenesis of AIDS virus reverse transcriptase.
Nature(London)327:716-717.
17. Larder, B., D. J. M.Purifoy,K. Powell, andG. Darby. 1987.
AIDS virusreversetranscriptase defined by highlevel
expres-sionin E. coli. EMBOJ. 6:3133-3137.
18. Larder, B.A., K. E. Coates, and S. D. Kemp. 1991.
Zidovudine-resistant humanimmunodeficiencyvirusselectedbypassagein
cell culture.J. Virol. 65:5232-5236.
19. Larder, B. A., and S. D. Kemp. 1989. Multiple mutations in HIV-1 reverse transcriptase confer high-level resistance to
zidovudine(AZT). Science 246:1155-1158.
20. LeGrice,S.,R.Zehnle,andJ.Mous. 1988. Asingle
66-kilodal-tonpolypeptide processed from the human immunodeficiency
virustype2polpolyprotein inEscherichia colidisplays reverse
transcriptase activity.J. Virol.62:2525-2529.
21. LeGrice, S. F. J., T. Naas, B.Wohlgensinger, and 0. Schatz. 1991. Subunit-selective mutagenesis indicates minimal polymer-aseactivity inheterodimer-associated pSl HIV-1 reverse
tran-scriptase.EMBO J. 10:3905-3912.
22. Lowe, D. M., V. Parmor, S. D. Kemp, and B. A. Larder. 1991.
Mutational analysis oftwoconservedsequencemotifsin HIV-1 reversetranscriptase. FEBS Lett.282:231-234.
23. McClure, M. A., M. S. Johnson, D. F. Feng, and R. F. Doolittle.
1988. Sequence comparisons of retroviral proteins: relative ratesofchange and general phylogeny. Proc. Natl. Acad. Sci. USA 85:2469-2474.
24. Nunberg, J. H., W. A. Schleif, E. J. Boots, J. A. O'Brien, J. C.
Quintero, J. M. Hoffman, E. A. Emini, and M. E. Goldman. 1991. Viral resistance to human immunodeficiencyvirus type
1-specific pyridinone reversetranscriptase inhibitors.J. Virol. 65:4887-4892.
25. Prasad, V., and S. P. Goff. 1989. Linkerinsertionmutagenesisof
the human immunodeficiency virus reverse transcriptase
ex-pressed in bacteria: definition of the minimal polymerase
do-main.Proc. Natl.Acad. Sci. USA 86:3104-3108.
26. Prasad,V., I.Lowy,T. D. Santos,L. Chiang, and S. P. Goff. 1991. Isolation and characterization of a dideoxyguanosine triphosphate-resistantmutantofhumanimmunodeficiencyvirus
reverse transcriptase. Proc. Natl. Acad. Sci. USA
88:11363-11367.
27. Restle, T., B.Muller, and R. S.Goody. 1990. Dimerizationof
humanimmunodeficiencyvirus type 1reversetranscriptase.A targetfor chemotherapeutic intervention. J. Biol. Chem. 265:
8986-8988.
28. Shih, C.-K., J. M. Rose,G.L.Hansen,J.C.Wu,A.Bacolla,and J.A. Griffin. 1991. Chimeric human immunodeficiency virus
type1/type 2 reversetranscriptasesdisplayreversedsensitivity tononnucleosideanalog inhibitors.Proc. Natl. Acad.Sci. USA 88:9878-9882.
29. Spanos, A.,S.G.Sedgwick, G.T.Yarranton,U.Hubscher,and G. R. Banks.1981. Detectionof thecatalyticactivities ofDNA
polymerasesandtheir associated exonucleasesfollowing SDS-polyacrylamidegel electrophoresis. Nucleic AcidsRes.
9:1825-1839.
30. St.Clair,M.H.,J.L.Martin,G.Tudor-Williams,M.C.Bach, C. L.Vavro, D. M. King,P. Kellam, S. D. Kemp, and B. A. Larder.1991. ResistancetoddIandsensitivitytoAZT induced byamutationin HIV-1reversetranscriptase.Science 253:1557-1559.
31. Varmus, H.1988. Retroviruses. Science 240:1427-1428.
32. Weiss, R., N.Teich, H. E.Varmus,and J.Coffin (ed.). 1985.
RNA tumor viruses. Cold Spring Harbor Laboratory, Cold Spring Harbor,N.Y.