0022-538X/95/$04.0010
Copyrightq1995, American Society for Microbiology
Structure and Role of the Terminal Repeats of Epstein-Barr
Virus in Processing and Packaging of Virion DNA
JULIE ZIMMERMANNANDWOLFGANG HAMMERSCHMIDT*
Institut fu¨r Klinische Molekularbiologie und Tumorgenetik, GSF-Forschungszentrum fu¨r Umwelt und Gesundheit GmbH, D-81377 Munich, Germany
Received 28 November 1994/Accepted 14 February 1995
The linear virion Epstein-Barr virus (EBV) DNA is terminated at both ends by a variable number of direct, tandemly arranged terminal repeats (TRs) which are approximately 500 bp in size. The number of TRs at each terminus can vary. After infection of host cells, the EBV DNA circularizes via the TRs by an unknown mechanism, and replication of the viral DNA during the lytic phase of the EBV life cycle leads to large DNA concatemers which need to be cleaved into virion DNA units, eventually. This cleavage event occurs at an
unknown locus within the TRs of EBV, which are the cis-acting elements essential for cleavage of the
concatemers and encapsidation of the virion DNA. To investigate the mechanism of DNA processing during genome circularization and cleavage of concatemeric DNA, the genomic termini of EBV were cloned, se-quenced, and analyzed by direct labeling of the virion DNA. Both termini ended with identical 11-bp elements; the right end has acquired an additional 9-bp stretch that seemed to originate from the leftmost unique
sequences. The left terminus is blunt, whereas the right terminus appears to have a 3*single-base extension.
In a transient packaging assay, a single terminal repeat was found to be sufficient for encapsidation of plasmid DNA, and mutagenesis of the TR element defined a region of 159 bp, including the 11-bp element, which is essential for packaging. These results indicate that the genomic termini of EBV are not generated by a simple cut of a hypothetical terminase. The mechanism for cleavage of concatemers seems to involve recombination events.
The termini of Epstein-Barr virus (EBV) virion DNA are involved in various processes of the viral life cycle, which con-sists of a latent and a lytic phase. Shortly after infection of B lymphocytes, the linear virion molecules are circularized. Dur-ing the latent phase, the EBV genome is maintained as a circular molecule and its DNA is replicated via oriP, the plas-mid origin of DNA replication (26). During the lytic phase, which is induced spontaneously or by expression of the BZLF-1 gene (3, 22), DNA replication is mediated by a dif-ferent origin of DNA replication, termed oriLyt (9). Lytic rep-lication leads to long head-to-tail concatemers which are cleaved to generate virion DNA units which are packaged into viral capsids. The protein or complex of proteins responsible for processing of the EBV virion DNA is called the terminase and has not been identified so far.
The termini of EBV DNA consist of a variable number of directly oriented 538-bp terminal repeats (TRs). The TRs of EBV are involved in the circularization event of the ge-nome, and they are the essential elements for cleavage and packaging of the EBV virion DNA (10). The mechanisms by which these processes are accomplished are not clear. The hypothetical terminase cleaves EBV concatemers into unit-length monomers somewhere within the TRs, but the cleavage and recognition sites of the terminase have not been identified yet. The sequence of the circular EBV ge-nome was determined, and the sequence of the fused ter-mini is known (1). The sequences at the terter-mini of the linear
virion DNA of EBV, however, have not been studied in detail so far.
We determined the terminal sequences of EBV by clon-ing and sequencclon-ing the terminal fragments of linear EBV DNA. Surprisingly, a sequence of 11 bp is repeated at both ends in direct orientation. An element of 9 bp derived from the most distal unique region near the left terminus appears at the right terminus. These findings could be confirmed by mea-suring the lengths of small single-stranded end fragments of linear EBV virion DNA. In addition, the fine structure of the EBV genomic termini was analyzed. The linear form of the EBV genome has two different end structures: the left
terminus is blunt, and the right terminus has a 39single-base
extension. We developed a functional assay that allowed us to test recombinant plasmid DNA for packaging by EBV. Recombinant plasmid DNA containing the cis elements for replication was transfected into P3HR1 helper cells and tested for packaging by the products of the endogenous EBV. Functional analysis of the TR sequence led to the observation that a single TR element is sufficient for cleav-age and packaging of the plasmid DNA by EBV, and dele-tion analysis of a single TR element showed that a sequence of 159 bp contains the signals essential for packaging. This finding indicates that the sequence requirement for cleavage and packaging extends beyond the vicinity of the site of cleav-age itself.
MATERIALS AND METHODS
Cells and virus.HH514 is a cell line derived from a single cell clone of the Burkitt’s lymphoma cell line P3HR1 (16). Cells were grown in RPMI 1640 cell culture medium supplemented with 10% fetal calf serum, 2 mML-glutamine, 100 U of penicillin per ml, and 100mg of streptomycin per ml. The virion DNA of the EBV strain, B95-8, that was used for cloning the terminal fragments of EBV and labeling experiments was a gift from Bill Sugden (Madison, Wisconsin). * Corresponding author. Mailing address: Institut fu¨r Klinische
Molekularbiologie und Tumorgenetik, GSF Forschungszentrum fu¨r Umwelt und Gesundheit GmbH, Marchioninistr. 25, D-81377 Munich, Germany. Phone: 49/89/7099-506. Fax: 49/89/7099-500. Electronic mail address: [email protected].
3147
on November 9, 2019 by guest
http://jvi.asm.org/
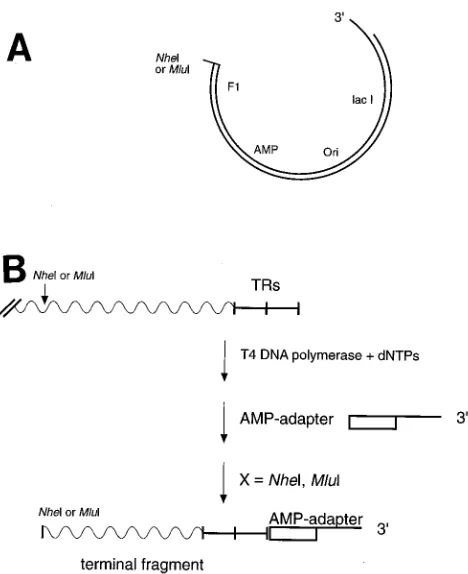
Cloning of the genomic termini.The strategy of cloning the terminal fragments of EBV is as follows (see Fig. 2). EBV virion DNA was made blunt by treatment with T4 DNA polymerase in the presence of the four deoxynucleotide triphos-phates. The AMP adapter with the sequence
59-TCGTCCCATGGGAATTCTGGACTTTCTACGCATGATGATGATG-39
39-AGCAGGGTACCCTTAAGACCTGAAAGATGCG-59
was ligated to the blunt ends. After cleavage of the modified virion DNA with
BamHI, the fragments with sizes of 4 to 6 kbp were isolated on preparative
agarose gels. Fragments of sizes appropriate for cloning were generated by cleavage of the BamHI fragments with NheI (left end) or with MluI (right end). Only the terminal fragments contained the AMP adapter on one side, and they were cloned in defined orientation into the modified pAMP1 vector (Gibco BRL). Clones carrying the left or right terminal fragments were identified by colony hybridization with radioactively labeled probes specific either for the left or for the right end of EBV DNA. An EcoRI-SalI fragment with the coordinates 1 to 644 of the EBV(B95-8) DNA cloned into pUC18 (p950) was used for detection of the left terminus; an MluI-BstNI fragment with the coordinates 169566 to 170117 was cloned into pUC18 (p951) for detection of the right terminus.
5*and 3*labeling of the EBV genomic termini.For 59labeling, 20mg of linear native untreated EBV DNA of the B95-8 strain was first dephosphorylated with calf intestine phosphatase and subsequently cut with BamHI after inactivation of the phosphatase. Fragments in the range of 4 to 6.5 kbp, including the end fragments, were recovered from a preparative agarose gel and labeled at the 59
ends with T4 polynucleotide kinase and 50mCi of [g-32
P]ATP. These labeled
BamHI fragments were cut with enzymes that have unique cleavage sites within
the terminal repeat. One half of the prepared fragments was cleaved with
Sau3AI to generate a left-end fragment of appropriate size, and the other half
was cleaved with BstEII to produce the right-end fragment. After cleavage, the DNA was extracted with phenol and butanol and the probes were loaded on an 8% denaturing sequencing gel.
EBV DNA was labeled at the 39ends with terminal deoxynucleotidyltrans-ferase (TdT) (Promega), which adds nucleotides to the free 39hydroxyl group of the DNA strands. Twenty micrograms of linear, native EBV DNA was melted at 708C in the presence of spermidin (1 mM) to make the 39strands more accessible for the enzyme. After being cooled to room temperature, the DNAs were labeled at the 39ends in a volume of 100ml containing 250mCi of [a-32
P]ddATP, 20ml of TdT buffer (500 mM cacodylate buffer [pH 6.8]–1 mM CoCl2-0.5 mM dithio-threitol–500mg of bovine serum albumin per ml), and 2ml (20 to 40 U) of TdT for 30 min at 378C. The reaction was stopped by heating of the mixture for 10 min at 708C. The 39-labeled DNA was cleaved with BamHI, and the BamHI frag-ments of between 4 and 6.5 kbp were prepared and cut with Sau3AI or BstEII. After extraction with phenol and butanol, the fragments were loaded on an 8% denaturing sequencing gel.
Plasmids.Plasmids p554 and p588 contain the origins of DNA replication,
oriP and oriLyt, and the TRs of the B95-8 strain. The two plasmids were used as
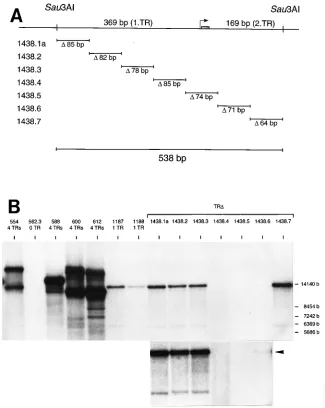
positive controls for packaging (10). The negative control p562.3 contains oriP and oriLyt but lacks the TRs (9). A single copy of the terminal repeats of EBV was isolated with Sau3AI or EcoNI and cloned into the BamHI site of plasmid p562.3 (pHEBo [26]), resulting in the plasmids p1187 and p1188 (see Fig. 7A). Plasmid p1187 contains B95-8 sequences from nucleotide position 170263 to position 170800; p1188 contains B95-8 sequences from position 170591 to posi-tion 171129. Oligonucleotide-directed mutaposi-tions were introduced into plasmid p1187 as described elsewhere (17) and were confirmed by DNA sequencing. The different mutants which are based on the p1187 plasmid (see Fig. 8) encompass the following deletions: p1438.1a, from 170263 to 170347; p1438.2, from 170348 to 170429; p1438.3, from 170430 to 170507; p1438.4, from 170508 to 170592; p1438.5, from 170593 to 170666; p1438.6, from 170667 to 170737; p1438.7, from 170738 to 170800.
Transient packaging assay. (i) DNA transfections.The plasmids were elec-troporated into HH514 cells by using the Bio-Rad gene pulser at 960mF and 220 V. HH514 cells (107cells in a volume of 250ml of RPMI 1640) were transfected with 10mg of pCMV BZLF-1 and 10mg of test plasmid. Four transfections were combined and resuspended in 15 ml of RPMI 1640 with 10% fetal calf serum.
(ii) Preparation of virion DNA from the viral capsids.The supernatant of the transfected cells was harvested after 4 days, and cell debris was removed by centrifugation at 3003g for 10 min and at 4,0003g for 20 min. The viral
particles were pelleted through a 20% sucrose (in Tris-EDTA-NaCl [TEN]) cushion by centrifugation at 25,000 rpm for 1.5 h at 158C in an SW28 rotor and resuspended in 900ml of 0.05% Tween for 1 h at 378C. To remove cellular DNA contamination, 100 ml of DNase buffer (0.5 M Tris-HCl [pH 7.5]–0.1 M MgSO4–1 mM dithiothreitol) and 30 U of DNase were added and the mixture was incubated for 1 h at 378C. The viral particles were lysed with 100ml of proteinase K (10 mg/ml) and 50ml of 10% sodium dodecyl sulfate overnight at 378C, and the viral DNA was isolated and purified.
(iii) Analysis of the packaged plasmid DNA.The virion DNA was cleaved either with BamHI or HindIII in the presence of XbaI, which was added to remove contaminating cellular DNA. XbaI does not cleave the plasmids but has many restriction sites in cellular DNA. Treatment with DpnI led to the removal of the nonreplicated bacterial input DNA. The restriction fragments were
sep-arated by electrophoresis on agarose gels and transferred to nylon filters. The filters were hybridized with DNA of vector pUC18, which detected the prokary-otic plasmid DNAs only.
RESULTS
The terminal fragments of linear EBV DNA.To select
ap-propriate restriction enzymes to clone the terminal frag-ments of EBV DNA, linear EBV(B95-8) virion DNA was digested with a set of various enzymes that do not cleave with-in the TR sequence. The EBV fragments were then separated on an agarose gel, and the terminal fragments were made visible by hybridization of the Southern blot with probes specific for the left and for the right ends (Fig. 1). A ladder of bands represents the terminal fragments, which vary by an increment of ca. 500 bp according to the number of TRs at each end. The distribution of the band intensities is the same as that reported earlier (12). Four equal populations of molecules have one, two, three, or four TRs at the left end. About 70% of the molecules have four TRs at the right end. The restriction enzymes NheI (for the left end) and
MluI (for the right end) were chosen for cloning because they
produce terminal fragments of convenient sizes (1.5 to 3.5 kbp).
Cloning and sequencing of the genomic termini. Several
technically different attempts were made to clone the virion DNA termini molecularly. The highest efficiency for cloning was achieved by using the strategy outlined in Fig. 2. The
FIG. 1. Restriction analysis of the EBV termini. Linear EBV DNA was cleaved with BspHI, MluI, XhoI, and NheI and additionally with HindIII for the right end. The cleaved EBV fragments were separated by electrophoresis, trans-ferred to nylon filters, and hybridized with probes specific for the left and right ends. (A) Autoradiography of a blot hybridized with radioactively labeled probe containing the unique sequence of the left terminus (strain B95-8, nucleotides 1 to 644). (B) Autoradiography of a blot hybridized with a radioactively labeled probe containing the unique sequence of the right terminus (strain B95-8, nucle-otides 169566 to 170117). Four major bands with similar intensities and a size difference of ca. 500 bp represent the left-terminus fragments with one to four TRs. The right-terminus fragment with four TRs has the strongest intensity. After longer exposure, fragments with up to 10 TRs can be detected. The restriction enzymes chosen for cloning the EBV ends were NheI for the left terminus and MluI for the right terminus, which produce small fragments of between 1.5 and 3.5 kbp.
on November 9, 2019 by guest
http://jvi.asm.org/
pAMP1 vector is a linearized DNA molecule with two 39 over-hangs of 12 bases that are not complementary to each other. The vector recircularizes and becomes transformable into
Escherichia coli only after a fragment with compatible ends is
inserted into it.
To prepare the appropriate inserts, the genomic termini of linear EBV DNA were made blunt and an oligonucleotide
adapter for one of the 39 12-base overhangs of the pAMP 1
vector was attached. The BamHI EBV fragments of between 4 and 6.5 kbp were cleaved with NheI (left terminus) or MluI (right terminus). The AMP adapter was attached to the termi-nal fragments of EBV only, which could be cloned in defined orientations into the pAMP 1 vector (Fig. 2B).
Clones containing the terminal fragments were identified by colony hybridization with a radioactive probe specific for either the left end or the right end. The junction between vector and insert was sequenced, and the boundaries of left and right termini are summarized in Table 1 and Fig. 3.
Fifteen clones containing the left terminus were analyzed. They all showed the same terminal base pair at position 172241 of the EBV prototype strain B95-8 with the exception of a single clone. Clones with the right fragments were found to be
heterogeneous, since the EBV inserts terminated at nucleotide positions 172251 to 172249.
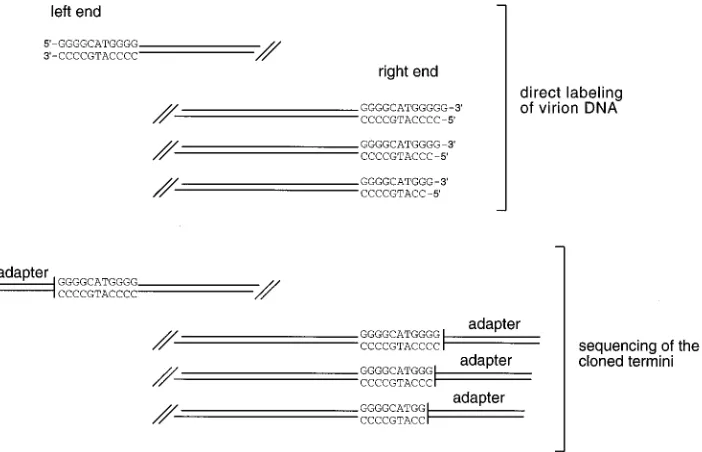
The terminal sequences of linear EBV DNA.The sequence
analysis of the left and right termini revealed that an 11-bp sequence is duplicated at each end (Fig. 3). This finding was surprising, because other herpesviruses do not show such a terminal sequence redundancy (2, 4, 8, 15, 23). Moreover, the right termini of EBV virion DNA seemed to have acquired a 9-bp unique sequence that appears to originate from the ulti-mate left boundary of the unique EBV sequence. Figure 4 depicts the arrangements of the 9- and 11-bp elements in the circular EBV genome (1) in comparison with the sequences of the cloned linear termini. The 11-bp element is repeated once in every TR and in addition appears next to the last TR in combination with the 9-bp element. The right termini of EBV end with a sequence of 20 bp (the 9-bp element plus the 11-bp element). All of the left termini end with the 11-bp element. Comparison of the cloned terminal sequences with the pub-lished sequence of the fused termini revealed that the right termini could have acquired these 20 bp from the ultimate left end.
Determination of the lengths of terminal fragments in EBV
virion DNA.This analysis led to the question of whether these
sequence structures do exist at the ends of EBV DNA or whether they result from a cloning artifact. Therefore, the precise lengths of the termini of native, unmodified linear EBV DNA were determined.
B95-8 virion DNA was labeled at the 59ends with T4
polynu-cleotide kinase and [g-32P]ATP and at the 39ends with TdT
[image:3.612.62.296.68.355.2]and [a-32P]ddATP. After digestion with BamHI, the labeled
FIG. 2. Cloning strategy for the terminal fragments of EBV DNA. (A) The linearized pAMP 1 vector (Gibco BRL) was chosen for cloning of the terminal DNA fragments. This vector contains 39extensions of 12 bases that are not complementary and cannot recircularize unless a fragment with appropriate end structures is inserted. To clone the left and right EBV DNA terminal fragments, the pAMP 1 vector was modified by restriction with NheI and MluI, respectively. (B) EBV(B95-8) linear virion DNA was treated with T4 DNA polymerase in the presence of nucleoside triphosphates to provide blunt ends. An AMP adapter complementary to the 39extension of the pAMP 1 vector was attached to the termini. Isolated BamHI fragments in a range between 4 and 6.5 kbp were cleaved with either NheI (left end) or MluI (right end). The resulting EBV DNA fragments were then cloned into the appropriately modified pAMP 1 vector. Clones containing the terminal fragments were identified by colony hybridization with probes specific for the left and right ends (for details, see Materials and Methods).
[image:3.612.317.555.90.191.2]FIG. 3. The terminal sequences at the ends of EBV DNA. Summary of the sequence analyses of the cloned termini revealed that both termini of EBV DNA end with the same 11-bp element. The appearance of the 9-bp element at the right end was unexpected, since this sequence belongs to the unique sequence of the left terminus.
TABLE 1. Determination of the terminal nucleotides of the EBV DNA end fragmentsa
Coordinate No. of clones
sequenced
Left terminus
172241... 14 172243... 1
Right terminus
172251... 13 172250... 8 172249... 1
a
Fifteen colonies containing the left terminus and 22 clones containing the right terminus of EBV DNA were sequenced at the junction between vector and insert. With the exception of one clone, all left-end inserts ended at position 172241 of the EBV(B95-8) DNA (1). The terminal base in the right-end clones varied between position 172251 and position 172249. The left cloned termini contained one to three TRs, and all contained the 11-bp element. Two clones derived from the right terminus contained two copies of the TR element; the remaining clones contained only one copy.
on November 9, 2019 by guest
http://jvi.asm.org/
[image:3.612.318.553.606.683.2]DNA was electrophoresed on an agarose gel and the DNA fragments of 4 to 6.5 kbp, including the terminal fragments, were recovered. The BamHI fragments were then cleaved sep-arately with additional restriction enzymes (the left end with
Sau3AI and the right end with BstEII) to yield small terminal
fragments. These small fragments, end labeled on either the 59
or the 39 end, were electrophoresed on denaturing
polyacryl-amide gels next to sequencing ladders of the corresponding cloned termini as length markers.
As shown in Fig. 5A, the lengths of the expected single-stranded DNA fragments were calculated on the assumption of blunt end structures of the termini. Sau3AI digestion of a blunt left terminus would yield two single-stranded fragments
la-beled at either the 39or the 59position with a size difference
of 5 nucleotides. Sau3AI produces a 59 overhang of 4
nucle-otides, and an additional nucleotide is added by labeling of the
39 end with [a-32P]ddATP. The labeled single-stranded
frag-ments of the right terminus would differ in their lengths by 6 nucleotides because of the additional ddATP nucleotide and
the 59 overhang of five nucleotides derived from the BstEII
cleavage site.
The results shown in Fig. 5B showed that the 59- and 39
-labeled fragments of the left terminus have the predicted
lengths (59left, 168 bases; 39left, 173 bases). The size
differ-ence of 5 nucleotides indicated that the left terminus of EBV must be blunt. The right terminus showed a ladder of
three bands for both the 39- and 59-labeled strands. This
minor variability of 3 bases at the right end is in agreement with our cloning and sequencing results for the right ter-minus. Bands of corresponding relative intensities have a size difference of 7 instead of the expected 6 nucleotides.
The 39-labeled strand is longer by one base (39 right, 117
bases). Therefore, the right end seems to have a single-base extension.
The results of sequencing the cloned terminal fragments and of labeling the native genomic termini are compared in Fig. 6. The termini of EBV DNA have different end structures: the
left end is blunt, whereas the right end probably has a 39
overhang of a single G. In the case of the right cloned terminal fragments, the treatment of virion DNA with T4 DNA
poly-merase has removed the single-base extension at the 39end by
the exonuclease activity of the enzyme. Thereby, the cloned right termini appeared to be truncated by a single residue compared with the right end of the native virion DNA. The heterogeneity of 3 bases found in the right-end clones is not the result of a cloning artifact and was confirmed by the label-ing experiment. It seems that the right end of EBV is gener-ated with less specificity than the left end. The results obtained with two completely independent approaches are identical. This coincidence indicates that the complicated terminal struc-tures of EBV virion DNA, including the duplication of the 11-bp elements and the apparent translocation of the 9-bp element, exist in virion DNA.
One TR element is sufficient and essential for DNA
pack-aging by EBV. The determination of the terminal sequences
of EBV DNA supplied us with enough information about the actual cleavage site to study the TRs functionally. Such an analysis should lead to the identification of the cis-acting signals essential for cleavage and packaging of EBV DNA.
A transient packaging assay that allows recombinant plasmid DNA to be tested for packaging by the endogenous EBV DNA was developed. The recombinant plasmid DNA to be tested and a BZLF-1 gene expression vector were cotransfected into
FIG. 4. Comparison of the circular genome and the cloned linear termini of EBV DNA. The upper part of the figure shows the fusion of the EBV DNA termini with four TR elements. Position 1 of the EBV genome is defined by the first EcoRI site following the TRs. The 11-bp element is repeated at the beginning of every terminal repeat and once outside the repetitive sequence together with the 9-bp element. In the lower part, the structures of the cloned left and right termini are depicted. The left termini contained one to three TRs and all ended with the 11-bp element. Most right termini had only one TR element, and only 2 of 22 clones contained two TRs. The right-terminus fragments end with a sequence of 20 bp consisting of the 9-bp element and the 11-bp element that seems to be derived from the left end. We assume that the structures of the terminal fragments with three or four TRs are consistent with these findings.
on November 9, 2019 by guest
http://jvi.asm.org/
a B-cell line latently infected with EBV (16). The lytic cycle of the virus was induced by expression of the BZLF-1 gene, and the viral capsid DNA was prepared and analyzed after 4 days. Plasmids p554 and p588 contain the fused termini with four TRs and oriLyt, the origin for lytic replication. Since their packaging has been shown previously, they were used as pos-itive controls (10).
To determine whether one TR element is sufficient for pack-aging, we cloned a single copy of the TRs into a vector con-taining oriLyt. The cloning strategy is shown in Fig. 7A. To investigate the significance of the position of the 11-bp ele-ment in a single TR copy, two fragele-ments were subcloned ei-ther with Sau3AI, so that the 11-bp element was positioned in the center of the repetitive sequence, or with EcoNI, which positions the 11-bp element more to the end of the TR se-quence.
Restriction analysis of the encapsidated DNA should lead to the detectable fragments shown in Fig. 7B. After lytic replica-tion, the concatemeric plasmid DNA is processed and pack-aged as multimers of the size of the EBV genome (data not shown and reference 22). Cleavage of these linear molecules of approximately 170 kbp with restriction enzymes will lead to internal fragments containing the fused TRs. In addition, ter-minal fragments are produced by the terminase activity and cleavage with restriction enzymes.
The positive control p554 showed the expected fusion frag-ment of 15 kbp and the terminal fragfrag-ments (9 to 11 kbp) that are identified in Fig. 8. No fragments were seen with the negative control plasmid p562.3, since this plasmid lacks the TRs and therefore was not encapsidated. The positive control p588 also showed the fusion fragment (16 kbp) and the termi-nal fragments (3.3 to 4.8 kbp), of which only the top fragment with four TRs is clearly visible in Fig. 7C. Both p1187 and p1188 were packaged, although less efficiently than the plas-mids with four TRs. The position of the 11-bp element within the TR sequence had only a slight effect on the efficiency of packaging.
The endogenous EBV of the cells was always packaged along with the test plasmids but was not detected with the vector probe (pUC18). As an internal control for the efficiency of DNA transfections, packaging, and equivalent preparations of the DNA samples, the Southern blots were stripped and rehybridized with EBV(B95-8) DNA. On the basis of these experiments, we consistently found that the plasmids p1187 and p1188 were less efficiently packaged than the positive con-trols p554 and p562.3, which is more evident in Fig. 8B (see below). Although the number of TR copies determines pack-aging efficiency to a certain extent, a single TR copy is suffi-cient and essential for processing and packaging of concate-meric plasmid DNA by EBV.
A region of 159 bp contains thecis-acting signals essential
for cleavage and packaging.To identify signals important for
packaging within the TR sequence, we constructed seven de-letion mutants based on plasmid p1187 (Fig. 8A). These mu-tants were tested for packaging, and the result of the experi-ment is shown in Fig. 8B. Four mutants, p1438.1 to p1438.3 and p1438.7, were packaged with the same efficiency as p1187. The sequences that were deleted in these constructs do not contain signals required for encapsidation. Packaging of plas-mid p1438.6 was greatly reduced, but a weak band representing the fusion fragment of 13 kbp was visible after longer exposure of the Southern blot. Plasmids p1438.4 and p1438.5 were not packaged, since their fusion fragments could not be detected at all. The signals required for processing and packaging of plas-mid DNA by endogenous EBV must lie within the 159-bp sequence which had been deleted in these plasmids. The
FIG. 5. Determination of the terminal structures of EBV DNA. (A) The predicted sizes of the 59- and 39-labeled single-stranded terminal fragments in the case of blunt termini. (B) Autoradiography of the sequencing gels. For analysis of the left (L) terminus, the 59- and 39-labeled Sau3AI EBV DNA fragments were loaded separately next to the sequencing ladder of the corresponding sequence of the cloned left terminus. Various exposures of the sequencing gel allowed the exact determination of the fragment lengths. The short exposure of the gel confirmed that the 39-labeled left fragment consists of a single band. The 59- and 39-labeled left terminal fragments (59L and 39L) showed the lengths expected for blunt left termini. Similarly, the 59- and 39-labeled BstEII fragments of the right terminus were loaded next to the sequencing ladder of the cloned right terminus. A ladder of three bands represents the heterogeneity of 1 to 3 bp at the right terminus. The 59-labeled top fragment (59R) had the expected length of 110 bases, but the 39-labeled fragment (39R) was longer by one nucleotide (117 bases), indicating that the right terminus has a 39single-base overhang. The stronger intensities of the 39-labeled fragments show that 39labeling had oc-curred with higher efficiency than 59labeling. Plasmid p1179 contains one TR element and was sequenced with two primers (1179/L and 1179/R) at the region of the 11-bp element to generate size markers.
on November 9, 2019 by guest
http://jvi.asm.org/
159-bp sequence includes the 11-bp element and the EBV equivalents of the putative packaging signals, pac1 and pac2, the only conserved regions found at the termini of herpesvi-ruses so far (Fig. 9). Plasmids p600 and p612 contain the TRs and differ only in the orientations of oriLyt to the TRs. Both were packaged with equal efficiency, showing that the orienta-tions of the TRs relative to oriLyt have no effect on the pack-aging event (Fig. 8B).
DISCUSSION
This study was performed to learn about the genomic ter-mini of EBV virion DNA. Several structural elements which constitute the EBV termini could be identified. An 11-bp ele-ment with direct orientation was found at both termini in all sequenced clones. In addition, the right end has acquired a 9-bp element that seems to originate from the ultimate left boundary of the unique segment of EBV DNA. In a compar-ison of terminal sequences found in the clones with the se-quence of the circular genome (1), it appears as if the ends display a 20-bp overlap. These sequence structures really exist at the native genomic termini, since they could be confirmed by measurement of the sizes of small labeled terminal fragments. Our findings indicate that the hypothetical terminase does not generate the termini during lytic replication by a simple cut as has been proposed for other herpesviruses (2, 8, 15, 23). The mechanism is more complex and may involve recombinational events.
Comparison of the TR elements of the B95-8 and the Na-malwa strains of EBV revealed that the 9-bp element is an integral part of each TR element in Namalwa EBV but is present only once within the unique sequence adjacent to the TRs (Fig. 4) (13). This observation could be confirmed by sequencing of the cloned TR elements derived from B95-8 DNA which were used in this study (data not shown). Accord-ing to our results, the 9-bp element was found attached to every right terminus. It appears, however, that the 9-bp
ele-ment does not have to be present in cis for the plasmids to be packaged. Plasmids p1187 and p1188 carry only one TR ele-ment and consequently lack the 9-bp eleele-ment (Fig. 7). Either the 9-bp element is not required for cleavage and packaging, or it may have been rescued by a recombination event with the endogenous EBV in the B cell during lytic replication. Alter-natively, it is possible that the 9-bp element is needed for a different process, like the circularization of the EBV genome, but not for cleavage and packaging, which alone were mea-sured in our transient packaging assay.
The left terminus of EBV is blunt, whereas the right ter-minus probably carries a single unpaired G. Most
herpesvi-ruses studied so far contain complementary single-base 39
extensions, indicating that the linear virion DNA could circu-larize by base pairing after infection of the host cells (2, 4, 8, 15, 23). Like pseudorabies virus with its blunt L terminus and 2-base extension at the S terminus (11), EBV has two different end structures that make circularization by base pair-ing and ligation improbable. In addition, the duplicated 11-bp elements at the termini of EBV must be removed upon cir-cularization. A recombination event could expel the dupli-cated 11-bp element during recircularization to prevent it from becoming amplified with each virus generation. Other-wise, viral propagation would lead to an unlimited gain of redundant sequence elements at the viral termini. A highly conserved cellular protein has been found to bind to the ter-mini of EBV in vitro, and its binding site spans the 11-bp element. It has been suggested that the protein is involved in such recombination processes, which remains to be demon-strated (21).
One TR element is sufficient for packaging, and deletion analysis identified a 159-bp sequence that includes all the re-quirements for processing and packaging of plasmid DNA. In Fig. 9, the terminal sequences of EBV and herpes simplex virus type 1 (HSV-1) are compared. The only two homologous se-quences found at the termini of herpesviruses are the packag-ing signals pac1 and pac2 (5). The packagpackag-ing signals are
lo-FIG. 6. The terminal structures of the linear EBV genome. The results of the sequence analysis of the cloned termini and analysis of the single-stranded terminal virion DNA fragments are compared. The left terminus is blunt and ends with the 11-bp element. The heterogeneity of 3 bases observed in the clones containing the right end could be confirmed by the labeling experiments with virion DNA. The size difference of the right single-stranded fragments indicates that the right terminus has a 39overhang of 1 base, which probably is a guanosine.
on November 9, 2019 by guest
http://jvi.asm.org/
[image:6.612.131.482.73.299.2]cated at conserved distances from the actual ends of the genome (41 to 47 bp for pac1 and 20 to 35 bp for pac2) among different herpesviruses. It has been proposed that the cleavage-packaging machinery is directed to defined distances from the packaging signals. Alignment of the terminal sequences of EBV and HSV-1 revealed that the spacing between the pack-aging signal homologous regions and the genomic ends of EBV is similar to those of HSV-1 and other herpesviruses (Fig. 9). The packaging signals as well as the 11-bp element are in-cluded in the 74-bp sequence region that was deleted in the nonfunctional TR mutant in p1438.5. The other mutant im-paired for cleavage or packaging, p1438.4, contained the
pack-aging signals and the 11-bp element. Thus, it is very likely that there are additional sequence elements adjacent to the pack-aging signal regions and the cleavage site itself which are re-quired for EBV processing.
The generation of the termini of EBV DNA can be ex-plained without the need for recombination by a model sug-gested for the resolution of concatemeric replication interme-diates of bacteriophage T7. In this case, the concatemers are
cleaved at two specific staggered points to yield 59overhangs,
which are then filled in by repair synthesis of the polymerase. Similarly, two single-stranded cleavages, one at the bottom strand on the right side of the 11-bp element located within the
FIG. 7. One terminal repeat is sufficient and essential for packaging. (A) The strategy for cloning a single copy of the TRs is shown. Plasmid p588 contains the fused termini of EBV DNA with four TRs and the essential signals for replication, oriLyt and oriP. Two different enzymes were used for removing the repetitive sequence of 538 bp containing different positions of the 11-bp element. Sau3AI produces a fragment with the 11-bp element positioned in the center, and the EcoNI fragment contains the 11-bp element at the end. Both fragments were cloned into plasmid p562.3. The resulting plasmids, p1187 and p1188, contain one TR element with different positions of the 11-bp element. (B) Restriction analysis of encapsidated plasmid DNA. Integral numbers of plasmid molecules are packaged as linear multimers of ca. 170 kbp. In addition to the fusion fragments, the restriction enzyme cleavage of these molecules should also generate terminal fragments resulting from the action of the hypothetical terminase that cleaves the DNA within the TRs. Presumably, the plasmid p554 is packaged as a hexamer. Cleavage with BamHI should lead to a ladder of terminal fragments (9 to 11 kbp) and to two fusion fragments, of which only the 15-kbp fragment with the TRs is detected with the pUC18 probe. HindIII restriction of p588, packaged as an 11-mer, generates a 16-kbp fusion fragment and terminal fragments of between 3.3 and 4.8 kbp. Plasmids p1187 and p1188 are probably packaged as 13-mers; their fusion fragments should have a size of 13 kbp, and the terminal fragments should have sizes of 6.9 kbp (p1187) and 7.2 kbp (p1188). (C) The transient packaging assay (see Materials and Methods) was used to analyze the packaging of plasmids p554, p562.3, p588, p1187, and p1188. The DNAs isolated from the virion preparations were cleaved with HindIII or BamHI, separated by gel electrophoresis, transferred to nylon filters, and hybridized with pUC18 DNA. The autoradiography of the blot is shown. The positive control plasmids p554 and p588, containing four TRs, are packaged and show the predicted sizes of the fusion fragments and the terminal fragments (Fig. 7B) which can be detected with this probe. The negative control p562.3 lacks the TR elements and was not packaged. Both plasmids with a single TR element were packaged, and the fusion fragments of p1187 and p1188 could be detected. The terminal fragments of the plasmids are indicated (arrowheads), but the terminal fragment of p1188 was clearly visible only after longer exposure (data not shown).
on November 9, 2019 by guest
http://jvi.asm.org/
unique leftmost sequence of EBV and one at the top strand on the left side of any 11-bp element, would provide staggered ends which could be filled in to yield the EBV termini (Fig. 4). This model is hampered by the observation that the copy num-ber of TRs at the right end is variable (Fig. 1), which could not be easily explained by such a staggered-cut repair model. Therefore, we favor a recombination model to explain the formation of the EBV termini.
The termini of many herpesvirus DNAs have been shown to be involved in recombination processes. The ability to in-duce recombination was most intensively studied with the
a sequence of HSV-1 (6, 14, 20, 25). A specific signal for
recombination could not be found at the termini of HSV-1 (19). However, the termini contain GC-rich sequences with a large amount of small oligo(dG) and oligo(dC) repetitive elements that are related to the immunoglobulin
heavy-chain sequences involved in class switch recombination (7, 18). It has been suggested that these GC-rich sequences me-diate the inherent recombinogenic activity of the a sequence. Although it is speculative, the class switch-like GC-rich se-quences, which are an integral part of the class switch-like GC-rich sequences present within the terminal repeats of EBV and the terminal structures of other herpesviruses, may be able to interact with the recombination machinery of the host cell to perform essential viral processes. In theory, such recombination processes during lytic replication of the EBV genome could lead to the duplication of the 11-bp element and the appearance of the 9-bp element at the right end. The fact, however, that all cloned termini showed the same terminal sequences indicates that such a mechanism must be highly specific and efficient. In addition, the two different end struc-tures of EBV indicate that the genome circularizes by a
mech-FIG. 8. Deletion analysis of the TR sequence. (A) Seven deletion mutants were constructed by in vitro oligonucleotide-directed mutagenesis of the 538-bp TR sequence of plasmid p1187. Mutants p1438.1a to p1438.7 and the sizes of the deleted sequences are depicted. A detailed description of the deletion endpoints in the individual mutants can be found in Materials and Methods. The shaded rectangle represents the position of the 11-bp element shown in Fig. 4. (B) Seven deletion mutants were tested for packaging. The plasmids analyzed in Fig. 7C were used as controls. Two plasmids, p600 and p612, containing the four TRs and oriLyt in different orientations, are packaged with similar efficiencies. Mutant plasmids p1438.1a, p1438.2, p1438.3, and p1438.7 were packaged. The packaging of p1438.6 was greatly reduced, but a weak band (13 kbp) could be detected after longer exposure (arrowhead) (shown below). No packaging was seen for the mutants p1438.4 and p1438.5. The 159-bp sequence deleted in these two plasmids (85 bp from p1438.4 and 74 bp from p1438.5) contains the essential signals for cleavage and packaging of plasmid DNA and corresponds to the nucleotide coordinates 170508 to 170666 of the EBV prototype strain B95-8.
on November 9, 2019 by guest
http://jvi.asm.org/
[image:8.612.147.472.70.478.2]anism that cannot easily be explained by a simple recombina-tion event. It is therefore likely that EBV has acquired a specific and yet to be defined mechanism to maintain the integrity of its ends.
ACKNOWLEDGMENTS
We thank Bill Sugden for B95-8 virion EBV DNA, suggestions, and valuable comments on the manuscript, and we thank our colleagues for helpful discussions.
This research was supported by grant AI-29988 from the National Institutes of Health and by grant Fa 138/3-7 from the Deutsche For-schungsgemeinschaft.
REFERENCES
1. Baer, R., A. T. Bankier, M. D. Biggin, P. L. Deininger, P. J. Farrell, T. J.
Gibson, G. Hatfull, G. S. Hudson, S. C. Satchwell, C. Seguin, P. S. Tufnell, and B. G. Barell.1984. DNA sequence and expression of the B95-8 Epstein-Barr virus genome. Nature (London) 310:207–211.
2. Chowdhury, S. I., H.-J. Buhk, H. Ludwig, and W. Hammerschmidt. 1990. Genomic termini of equine herpesvirus 1. J. Virol. 64:873–880.
3. Countryman, J., and G. Miller. 1985. Activation of expression of latent Epstein-Barr herpesvirus after gene transfer with a small cloned subfragment of heterogeneous viral DNA. Proc. Natl. Acad. Sci. USA 82:4085–4089. 4. Davison, A. J. 1984. Structure of genome termini of varicella-zoster virus. J.
Gen. Virol. 65:1969–1977.
5. Deiss, L. P., J. Chou, and N. Frenkel. 1986. Functional domains within the
a sequence involved in the cleavage-packaging of herpes simplex virus
DNA. J. Virol. 59:605–618.
6. Dutch, R. E., R. C. Bruckner, E. S. Mocarski, and I. R. Lehman. 1992. Herpes simplex virus type 1 recombination: role of DNA replication and viral a sequences. J. Virol. 66:277–285.
7. Gomez-Marquez, J., A. Puga, and A. L. Notkins. 1985. Regions of the terminal repetitions of the herpes simplex virus type 1 genome. Relationship to immunoglobulin switch-like DNA sequences. J. Biol. Chem. 260:3490– 3495.
8. Hammerschmidt, W., H. Ludwig, and H.-J. Buhk. 1988. Specificity of cleav-age in replicative-form DNA of bovine herpesvirus 1. J. Virol. 62:1355–1363. 9. Hammerschmidt, W., and B. Sugden. 1988. Identification and characteriza-tion of oriLyt, a lytic origin of DNA replicacharacteriza-tion of Epstein-Barr virus. Cell
55:427–433.
10. Hammerschmidt, W., and B. Sugden. 1989. Genetic analysis of immortaliz-ing functions of Epstein-Barr virus in human B lymphocytes. Nature (Lon-don) 340:393–397.
11. Harper, L., J. Demarchi, and T. Ben-Porat. 1986. Sequence of the genome
ends and of the junction between the ends in concatemeric DNA of pseu-dorabies virus. J. Virol. 60:1183–1185.
12. Kintner, C. R., and B. Sugden. 1979. The structure of the termini of the DNA of Epstein-Barr virus. Cell 17:661–671.
13. Matsuo, T., M. Heller, L. Petti, E. O’Shiro, and E. Kieff. 1984. Persistence of the entire Epstein-Barr virus genome integrated into human lymphocyte DNA. Science 226:1322–1325.
14. Mocarski, E. S., and B. Roizman. 1982. Herpesvirus-dependent amplifica-tion and inversion of cell-associated viral thymidine kinase gene flanked by viral a sequences and linked to an origin of viral DNA replication. Proc. Natl. Acad. Sci. USA 79:5626–5630.
15. Mocarski, E. S., and B. Roizman. 1982. Structure and role of the herpes simplex virus DNA termini in inversion, circularization and generation of virion DNA. Cell 31:89–97.
16. Rabson, M., L. Gradoville, L. Heston, and G. Miller. 1982. Non-immortal-izing P3J-HR-1 Epstein-Barr virus: a deletion mutant of its transforming parent, Jijoye. J. Virol. 44:834–844.
17. Schepers, A., D. Pich, J. Mankertz, and W. Hammerschmidt. 1993. cis-acting elements in the lytic origin of DNA replication of Epstein-Barr virus. J. Virol. 67:4237–4245.
18. Shimizu, A., and T. Honjo. 1984. Immunoglobulin class switching. Cell 36: 801–803.
19. Smiley, J. R., J. Duncan, and M. Howes. 1990. Sequence requirements for DNA rearrangements induced by the terminal repeat of herpes simplex virus type 1 KOS DNA. J. Virol. 64:5036–5050.
20. Smiley, J. R., C. Lavery, and M. Howes. 1992. The herpes simplex virus type 1 (HSV-1) a sequence serves as a cleavage/packaging signal but does not drive recombinational genome isomerization when it is inserted into the HSV-2 genome. J. Virol. 66:7505–7510.
21. Sun, R., T. A. Spain, S. F. Lin, and G. Miller. 1995. Autoantigenic proteins that bind recombinogenic sequences in Epstein-Barr virus and cellular DNA. Proc. Natl. Acad. Sci. USA 91:8646–8650.
22. Takada, K., N. Shimizu, S. Sakuma, and Y. Ono. 1986. trans activation of the latent Epstein-Barr virus (EBV) genome after transfection of the EBV DNA fragment. J. Virol. 57:1016–1022.
23. Tamashiro, J. C., and D. H. Spector. 1986. Terminal structure and hetero-geneity in human cytomegalovirus strain AD169. J. Virol. 59:591–604. 24. Vlazny, D. A., and N. Frenkel. 1981. Replication of herpes simplex virus
DNA: localization of replication recognition signals within defective virus genomes. Proc. Natl. Acad. Sci. USA 78:742–746.
25. Weber, P. C., M. D. Challberg, N. J. Nelson, M. Levine, and J. C. Glorioso. 1988. Inversion events in the HSV-1 genome are directly mediated by the viral DNA replication machinery and lack sequence specificity. Cell 54:369– 381.
26. Yates, J. L., N. Warren, and B. Sugden. 1985. Stable replication of plasmids derived from Epstein-Barr virus in various mammalian cells. Nature (Lon-don) 313:812–815.
FIG. 9. Comparison of the native genomic termini of EBV and HSV-1. The terminal sequences of EBV(B95-8) and HSV-1(KOS) were aligned (24). The packaging region homologies (An, Cn, Anmotif and GC motif), the 11-bp elements, and the 9-bp element of EBV DNA are indicated. Arrows mark the distances between the A motif and the native ends of EBV and HSV-1 DNA.