0022-538X/94/$04.00+0
Copyright C) 1994, American Society for Microbiology
Sequence of the Phosphoprotein Gene of Pneumonia
Virus of
Mice:
Expression of
Multiple Proteins
from Two
Overlapping
Reading
Frames
J.
BARR,'t
P.CHAMBERS,'t
P.HARRIOTI,2
C.R.PRINGLE,1
ANDA. J.EASTON"*
Department of Biological Sciences, UniversityofWarwick, Coventry CV47AL,
England,'
and MedicalBiologyCentre, The Queen's Universityof Belfast, Belfast BT9 7BL,Northem Ireland2Received 15 December1993/Accepted22April 1994
The gene encoding thephosphoprotein of the pneumoviruspneumonia virus of mice (PVM) has been cloned andsequenced.Thegeneis 903 nucleotides inlengthand contains alongopenreading frame (ORF) capable of encoding a polypeptide of 295 amino acid residues. A smaller, second, overlapping ORF encoding a polypeptide 137 aminoacids in lengthwas alsopresent. The large ORF directed thesynthesis of a39-kDa polypeptide andfouradditional polypeptideswithmasses of37kDa, 26 kDa,23kDa,and16 kDa in vitro. The smaller polypeptides were generated by internal initiation on in-frame AUG initiation codons to generate carboxyco-terminal products. Western immunoblot analysis indicated thatatleasttwooftheseproteins and several otherrelatedpolypeptides arepresentininfected cells,and thepossible originsof these are discussed. Western blot analysis using antiserumraisedagainst asynthetic peptideandspecificfor thepredictedsecond ORF product identifiedapolypeptideof 23kDainPVM-infectedcells.Thepatternof PVM Pgeneexpression isunlike that of the closely related respiratory syncytialvirus and is reminiscentofthatofparamyxoviruses such asSendai virus. This is thefirstexample of a pneumovirus encoding multiplepolypeptide productsfrom a singlemRNAin vivo.
Pneumonia virusof mice(PVM) is classifiedas amemberof the subfamily Pneumovirinae of the family Paramyxoviridae. Within this subfamily PVMis grouped in the genus Pneumo-virus along with human respiratory syncytial virus (HRSV), bovine respiratory syncytial virus (BRSV), and turkey rhino-tracheitisvirus (TRTV). HRSV is thebest-characterized mem-ber of this genus because it is the agent most frequently isolated in acute respiratory tract infection in children (29). PVMwasfirst identified as an agent responsible for a latent or inapparent infection of mice (12). The isolation of PVM, or an antigenically related virus, from a variety of rodents and other species has been reported. Serological studies have shown that the human population is frequently exposed to infection by PVMoranantigenically related virus(11, 25). No disease has been associated withseroconversion in humans, and the clin-icalsignificance of PVM infection is unclear.
Nucleotide sequence studies have demonstrated that while the fourpneumoviruses are similar to one another(4), they are quite distinct from members of the genera Morbillivirus, Paramyxovirus,andRubulavirus, which comprise the remainder of thefamily Paramyxoviridae. The 10pneumovirus genes show little homology to the 6 or 7 genes that make up the morbilli-virus orparamyxovirus genome (la, 5-7). One of many other distinguishing differences exhibited at the molecular level by parainfluenza viruses, morbilliviruses, and rubulavirus is the possession of a phosphoprotein (P) gene with a second, overlappingopen reading frame(ORF), whereas the P genes of HRSV and BRSV contain only one ORF(2,9, 13-15, 19,21, 23, 28, 30, 31).
cDNA clones representing PVM mRNAs have been
de-*Correspondingauthor. Phone: 203 523523.
Fax:
203 523701.tPresentaddress: DepartmentofMicrobiology, Universityof Ala-bama atBirmingham, Birmingham, AL 35294-0005.
tPresent address: MRC Collaborative Centre, Mill Hill, London NW7
lAD,
England.scribed previously (3). Many cDNA clones representing the PVMphosphoproteinwereobtained,noneof whichcontained thefull-length gene without cDNA rearrangements (la). The 5'-terminal sequence of thePgenewasobtainedbyPCR(26) between oligonucleotide PVMP1, representing the antisense of P-gene residues 115 to 129 (GGTGACATGAGTAGTG
GCAGTGTTCGG115),
and oligonucleotide Oli-C(la).
The underlined sequence indicates residues homologouswith the PVMPgene,and the number indicates the nucleotidetowhich the 3' nucleotide binds. Restriction enzyme-generated frag-mentsof the Pgenewere subcloned intobacteriophage M13 mpl8and M13mpl9 sequencingvectorsby standard molecu-lar techniques (22), and thenucleotide sequence was deter-minedbythe chaintermination method ofSangeret al. (27). Sequence ofthe PVM Pgene.The mRNA sequenceand the predicted polypeptide sequence of the PVM phosphoprotein geneareshown inFig.1.The gene is 903nucleotideslongfrom thefirst base transcribed into mRNAtothe last baseadjacenttothepoly(A)tractandcontainsalongORF whichstartsfrom amethionine residuecorrespondingtonucleotidesatpositions 10 to 12 and finishes at a termination codon at nucleotide positions 895 to 897. The determined length of the gene corresponds well with the estimated size of the mRNA ob-served in Northern(RNA)blots(3).ThePVM Pgenebegins and ends with the conserved PVM genestartand stop consen-sussequences,previously reported (3).ThislongORF has the potential to encode a polypeptide 295 amino acids inlength which hasapredictedmolecularweightof 32,916. Thisfigure is notin close agreement with the observed molecularweight of the PVM P protein as calculated by sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE), which is approximately 39,000 (18). We have shown that the 39-kDa protein is encoded by the PVM P gene by hybrid arrested translation in vitro (3). Aberrant migration in SDS-PAGE is, however, a common feature of paramyxovirus P
proteins. An immediately obvious difference between the P
5330
on November 9, 2019 by guest
http://jvi.asm.org/
AGGAUAAAUAUGGAGAAAUUCGCCCCCGAAUUUGUUGGCGAGGAUGCUAACAAGAAGGCAGAGGAGUUUCUCAAACAUAG 80
N E K F A P E F V G E D A N K K A E E F L K H R
AUCCUUCCCUUCGGAAAAACCACUAGCUG6UAUACCGAACACUGCCACUCAUGUCACCAAAUAUAACAUGCCCCCUAUAU 160
S F P S E K P L A G I P NT A T H V T K Y N M P P I
M S P N I T C P L Y
UGCGUAGCUCAUUCAAACUCCCUUCCCCGAGAGUUGCUGCAAAUCUUACU6GAACCCUCUGCUCCCCCUACCACUCCACCA 240
L R S S F K L P S P R V A A N L T E P S A P P T T P P
C V A H S M S L P R E L L a I L L N P L L P L P L H H
CCCACACCUCCCCAGAACAAGGAAGAGCA6CCCAAAGAG6UCUGAUGUUGACAUUGAGACUAUGCAUGUCUGUAAGGUUCC 320
P T P P Q N K E E Q P K E S D V D I E T N H V C K V P
P H L P R T R K S S P K S L N L T L R L C N S V R F
UGACAAUCCGGAACACAGCAAGAAGCCAUVCUGCUCAGAUGAUACCGAUACUAAGAAAACUA6GAAGCC6AUGGUCACCU 400
D N P E H S K K P C C S D D T D T K K T R K PN V T
L T I R N T A R S H A A H IDP I L R K L G S R W S P
UUGUGGAACCC6AGGAGAAAUUUGUCGGAUUGGGAGCUAGCUUGUACAG6GAGACCAUGCAGACCUUUGCUGCUGAUGGU 480
F V E P E E K F V G L G A S L Y R E T N Q T F A A D G L W N P R R N L S D W E L A C T G R P C R P L L L H V
UAUGAUGAA6AAAGCAACCUAUCGUUUGAGGAGACUAACCAAGAGCCGGGUUCUUCAUCUGUAGAACAAAGACUAGAUAG 560
Y D E E S N L S F E E T N a E P G S S S V E a R L D R
N N K K A T Y R L R R L T K S R V L H L
AAUAGAGGAGAAAUUGUCCUACAUAAUAG6CCUUUUAAACACCAUAAUGGUAGCGACUGCUGGACCUACCACUGCUAGAG 640
I E E K L S Y I I G L L N T I N V A T A G P T T A R
AUGAGAUUAGAGAUGCCCUUAUAGGCACUAGAGAAGAACUUAUUGAGAUGAUCAAGUCUGACAUCUUGACUGUCAAUGAC 720
D E I R D A L I G T R E E L I E N I K S D I L T V N D
AGAAUAGUGGCCAUGGAGAAGCUCAGAGAU6AGGAAUGCUCCAGAGCUGACACUGAUGAUGGAUCAGCCUGUUAUUUAAC 800
R I V A N E K L R D E E C S R A D T D D G S A C Y L T
D R A R I L D K I V S S N A E E A K E D L D V D D I
UGGGCAUUAiUUUUUAGUUAAUU 903 M I N F
FIG. 1. Nucleotide sequence of PVM P gene shown in mRNA
sense. Thesequencesofthepredicted polypeptides encoded by both the long and the short ORFs are shown underneath. The numbers
refertothe nucleotidepositions in thegene.
proteins ofPVM,HRSV, and BRSV is their length, deduced from the nucleotide sequences. The PVM Pproteinencoded
bythe long ORFis 295residues long, whereas theHRSV and BRSV Pproteins are both 241 residues in length (15, 19, 21,
28). The homology that is seen between the PVM P protein
and either the HRSV or BRSV P protein is not evenly distributed throughout the length of the proteins but is
con-centrated into two regions of high levels of homology
sepa-ratedbyaregion ofpoorhomology (Fig. 2).Duringsequence
analysis of the pneumovirus P proteins, all the gap
substitu-tions introduced to generate alignment were located in this
region, afeature thathighlights the lack ofsimilarities in this
PVM MEKFAPEFVGEDANKKAEEFLKHRSFPSEKPLAGIPNTATHVTKYNMPPILRSSFKLPSP 60 RSA2 MEKFAPEFHGEDANNRATKFLES IKGKF 28 18537 MEKFAPEFHGEDANNKATKFLES IKGKF 28 PVM RVAANLTEPSAPPTTPPPTPPQNKEEOPKESDVDIETNHVCKVPDNPEHSKKPCCSDDTD 120
RSA2 TSPKDPKKKDSIISVNSIDIEVTKESPITSNSTIINPTNETDDTA 73
18537 ASSKDPKKKDSIISVNSIDIEVTKESPITSGTNIINPISEADSTP 73
*..*.. ...***... . ...
PVM TKKT RKPMVTFVEPEEKFVGLGASLYRETMOTPAADGYDEESNLSFEETNOEPGSSS 177
RSA2 GNKPNYORKPLVSFKEDPTPSDNPFSKLYKETIET FDNNEEESSYSYEEINDOT NDN 130
18537 EAKANYPRKPLDSFKEDLTPSDNPFSKLYKETIET FDNNEEESSYSYEEINDOT NON 130
PVM VEORLDRIEEKLSYIIGLLNTIMVATAGPTTARDEIRDALIGTREELIEMIKSDILTVND 237 RSA2 ITARLDRIDEKLSEILGMLHTLVVASAGPTSARDGIRDAMIGLREEMIEKIRTEALMTND 190
18537 ITARLDRIDEKLSEILGMLHTLVVASAGPTSARDGIRDAMVGLREEMIEKIRAEALMTND 190
PVM RIVAMEKLRDEECSRADTDDGSACYLTDRARILDKIVSSNAEEAKEDLDVDDIMGINF 295
RSA2 RLEAMARLRNEESEKMAKDTSDEVSLNPTSEKLNNLL EGNDSDNDLSLED F 241
18537 RLEAMARLRNEESEKMAKDTSDEVSLNPTSKKLSNLL EDNDSDNDLSLDD F 241
[image:2.612.56.294.517.665.2]* * *.... ***...**. *...**
FIG. 2. Alignment ofthe amino acid sequence of the PVM and subgroup A and B HRSV (strains A2 and 18537, respectively) P proteins.Thealignmentwasperformedwith theCLUSTALprogram
(10). Amino acid identities (*)andsimilarities (-)areshown.
Num-berstotherightof thesequencesrefertothe residues ofeachprotein.
internal region of the pneumovirus P proteins. The first region of high-level similarity is located at the extreme amino termi-nusof these proteins and is only 17 amino acids in length. The secondregion of high-level homology between these P proteins is located between residues 181 and 249 of the PVM P protein sequence and 134 and 201 of theHRSV and BRSV P protein sequences. The percent identities for this region are64% for PVM and HRSV and 67% for PVM and BRSV. These figures increase to 84 and 86%, respectively, when homologous sub-stitutions are also considered. Comparison of the amino acid sequences of the PVM and TRTV P proteins reveals that in TRTV, this second region of homology is again conserved, although the first region of homology at the amino terminus is not(16). The P proteins of PVM and TRTV have 69% identity in thehomologous region, rising to 82% upon consideration of similarities. Thehigh level of homology that is seen between all fourpneumoviruses in this region suggests that this region of P mayhave an important function. This high level of amino acid identity is also consistent with the observation that the PVM P protein is serologically related to the HRSV and BRSV P proteins(18). In the amino-proximal region of poor homology, which spans residues 18 to 180, there are only 38 identities (23% identity) with the HRSV P protein. The P proteins of HRSV and BRSV also show relatively poor amino acid identity (37%) in this region (21). A single divergent region located in the amino-terminal half of the HRSV P protein has been reported (13, 19). This region lies within the portion of the HRSVPprotein that displays poor homology with the corre-sponding region of the PVM P protein. It seems that this region does not require strict conservation of amino acid residues and is consequently tolerant to nonconservative amino acid changes.
ThePVM Pgene also contains asecond, smaller ORF which is 411 nucleotides in length. The coding region of the second ORF extendsfromamethionine residue located at nucleotides 131 to133 ofthe PVM P gene to a termination codon located at nucleotides 542 to 544. This second ORF encodes a polypeptide 137 amino acids in length and with a predicted molecularweight of 15,932. The position of the PVM P-gene second ORF coincides with theregion of poor homology, with the termination codon for the second ORF lying just 8 nucleotides upstream of the codon that encodes the first residue of the large region of high conservation. The region of poor homology includes the entire length of the sequence correspondingto the second ORF. The apparent lack of need for strict conservation of amino acid residues in this region has permittedchanges at the nucleotide level and the creation of a substantial coding region in a second ORF. It may be that the need for more strict amino acid conservation in the immedi-ately adjacent conserved region of the PVM P protein has prevented extension of the second ORF coding region past the boundaryof the variable and homologous regions.
Expression of the PVM Pgene. The production of PVM-specificmonoclonal antibodieswas describedpreviously (18). Screeningof thehybridoma cell lines for the ability to produce anti-P-proteinspecific antibodieswas performed by immuno-precipitationof
[35S]methionine-labelled
Pprotein synthesized in a rabbit reticulocyte lysate system. Anantibody designated 26/3/B5 was positive by this procedure (data not shown). Western immunoblot analysis with antibody 26/3/B5 recog-nized 39-, 26-, and 23-kDa polypeptides and eight additional polypeptides, albeit in small amounts, in PVM-infected BSC1 celllysates.In anattempttoidentify thelikely originsof these polypeptides, a series oftruncated P-gene plasmidswas con-structed in atranscriptionvector.Afull-length P-genecoding region was generated by ligating together DNA fragmentson November 9, 2019 by guest
http://jvi.asm.org/
A
1 2 3 4B
39-
9'_-37 _,
1 2 3 45 kd
w 439
-37
'>.,'>>...
16_l~~~~l
:".
-OW 5_23
--_own -016
FIG. 3. (A) Identification of AUG codons usedtoinitiate transla-tion of P-related polypeptides in vitro. This autoradiograph of a
polyacrylamide gel shows radioactively labelled P-protein-related polypeptides translated in vitro with rabbit reticulocyte lysate
pro-grammedwith mRNA transcribed from plasmids pGEMP (lane 1), pGEMP37 (lane2), pGEMP26(lane3), and pGEMP23 (lane4). (B) ThePVM P-gene longORFencodesseveralproteins invivo. Proteins fromE.coliBL21transformed with plasmids pGEMP and pGEMP37
were separated by SDS-PAGE together with proteins from PVM-infectedandmock-infected BSC1 cells, Western blotted, and probed with PVM P-protein-specific monoclonal antibody 26/3/B5. Lane 1, PVM-infectedBSC1 cell material; lane 2,mock-infected cell material; lane3, E. coli BL21 transformed with pGEMP;lane 4, E. coli BL21 transformedwithpGEMP37; lane 5, E. coli BL21 transformed with pGEM1as acontrol. Polypeptide molecularmasses are indicated, in kilodaltons.
from PCRs and cDNA clones. The nucleotidesequenceof the
full-length constructwasverified. The insert wascloned into
themultiplecloningsite of the T7transcriptionvectorpGEM1
(Promega Corporation) to give the plasmid designated
pGEMP. Correct orientation of the cDNA fragment was
confirmed by restriction enzyme digestion. Next, various
amountsofthe 5'-terminal sequenceofthefull-lengthP-gene cDNA contained within pGEMPwere removed byrestriction
enzyme digestion between a site within the P-gene coding
region and the EcoRI site present in the pGEM1 multiple
cloning site at the 5' end of the P-gene cDNA. The cleaved
endswere blunted with the Klenow fragment ofDNA
poly-merase 1 and ligated to generate plasmids containing
trun-cated Pgenes.The sitesusedwerefor AccI, HincII, NsiI, and
NheI, producing pGEMP37, pGEMP26, pGEMP23, and
pGEMP16, respectively. The locations ofthe restriction
en-zymesusedtoremovethe5'-terminal sequences areindicated
inFig.1. TheT7promoter-proximalATGinitiation codons of these five transcription plasmids are the first, second, third,
fourth,and fifthATG codons, respectively, of the long ORFin thefull-length PVM P-gene cDNA. mRNAtranscribed from plasmidpGEMPwastranslatedinareticulocyte lysatesystem,
and the [35S]methionine-labelled polypeptide products were
subjected toSDS-PAGE. An autoradiograph of the resulting polyacrylamidegel isshown in Fig. 3A. A longexposureshows
that the full-lengthP-gene cDNA insertof pGEMPdirectsthe
invitro synthesisofa39-kDa (P39),full-length PVM Pprotein
and smalleramountsofatleast four additional P-gene-specific polypeptideswithmolecularweightsof 37,000, 26,000, 23,000, and16,000. All fivepolypeptideswereimmunoprecipitated by
the PVM P-protein-specific monoclonal antibody, 26/3/B5
(data not shown). pGEM37 directed the production of
polypeptides with molecular weights of 37,000, 26,000, 23,000, and 16,000 but not the full-length P39 protein. Thus, the removal of the AUG codon at position 10 of the mRNA prevents synthesis of the full-length P39 protein, indicating that this is initiated from the first AUG codon.
The first ATG initiation codon present on the PVM P-gene insert of pGEMP37 is at nucleotide positions 148 to 150 of the PVM P gene, and this is the second AUG codon ofthe large ORF. Removal of this second, in-frame AUG codon prevented the expression of the 37-kDa polypeptide when the mRNA transcribed from pGEMP26 was translated. Thus the 37-kDa protein (P37) is expressed from the second AUG of the PVM P-gene long ORF. The transcription plasmid pGEMP26 is capable of directing the synthesis of the 26-kDa (P26), the 23-kDa
(P23),
and the 16-kDa(P16)
polypeptides.Transcription plasmid pGEMP23, which directed the syn-thesis of P23 and
P16,
lost the ability to direct expression of the P26 polypeptide in conjunction with the loss of the third AUG in frame with the long ORF of the PVM P gene (nucleotide positions 301 to 303 of the PVM P gene), indicating that P26 expression is initiated from this AUG codon. Plasmid pGEMP23 only directed the synthesis of two detectable PVM P-related polypeptides, P23 and P16. The first AUG codon on the truncated PVM P-gene insert of pGEMP23 is that of nucleotide positions 391 to 393 of the PVM P gene. This is the fourth AUG codon in frame with the long ORF. There are no other AUG codons close enough to direct the synthesis of a polypeptide of such size, and so we conclude that P23 is initiated by this AUG.Similarly, the first available AUG codon of pGEMP16 is at nucleotides 457 to 459 and is also in frame with the long ORF. mRNA from pGEM16 directed the synthesis of a 16-kDa polypeptide when translated in a wheat germ translation system (data not shown), indicating that
AUG457
is responsible for initiating translation ofP16.
The T7 transcription plasmids pGEMP, pGEMP37, pGEMP26, pGEMP23, and pGEM16 were transformed into Escherichia coli BL21, which contains an
isopropyl-p-D-thioga-lactopyranoside (IPTG)-inducible chromosomal copy of the T7 RNA polymerase gene (32). The plasmids directed the synthesis of the range of polypeptides seen in vitro. The 39-, 26-, and 23-kDa polypeptides seen in vivo comigrated with the P39,P26, and P23 proteins expressed from constructs pGEMP and pGEMP37 in E. coli BL21 (Fig. 3B) and with proteins of corresponding size synthesized in vitro (data not shown). The antibody did not identify polypeptides with molecular weights corresponding to those of either theP16
or P37 polypeptide from the PVM-infected BSC1 cell lysate, though this may be because these polypeptides were present in small amounts. The identities of the other eight polypeptides recognized by monoclonal antibody 26/3/B5 are not presently known, al-though it is unlikely that they represent primary products of internal initiation since their molecular weights do not corre-spond to the P-protein-related polypeptides expressed in vitro from the first five P-gene AUG codons. The 35-kDa polypep-tide that was identified may be initiated from the same AUG that initiates translation of the 37-kDa protein in vitro(AUG148)
and subsequently is covalently modified, since both the 35- and 37-kDa proteins (and all other forms of P-related proteins) fail to react with the monoclonal antibody 26/4/C4 described by Ling and Pringle (18), which recognizes onlyP39,
this indicates that the 35- and 37-kDa proteins lack the amino-terminus of the full-length P39 protein. Alternatively, the 35-kDa polypeptide may arise through a cleavage event which removes the amino terminus of either the P39 or P37 protein.
I
on November 9, 2019 by guest
http://jvi.asm.org/
[image:3.612.79.276.67.221.2]Identification of the second ORF protein. The complete coding region present within the PVM P-gene second ORF wasamplified by PCR using oligonucleotide primers PVMP2A (positive sense) (CCGGGATCCTCATGTCACCAAATATA
ACGTGCCCC154)
and PVMP2B (negative sense) (CCTAAGCTTCTCCTCTATTCTATC556).
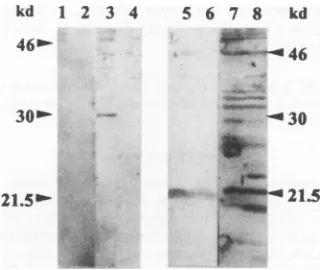
The underlined regionsindicate the sequences corresponding to the PVM P gene, and the number indicates the nucleotide position to which the terminal base anneals. The amplified fragment was end filled, cleaved with restriction enzyme BamHI, which cuts in PVMP2A, and then inserted by standard molecular techniques into theBamHI andSmaI sites present in the polylinker region of theprokaryotic expression vector pQE32 (Qiagen Inc.) (22). Theresulting plasmid, pQE32.P2, was transformed into E. coli M15 (Qiagen Inc.). Expression and purification of the PVM P-gene second ORF product was carried out according to directions supplied by the manufacturer. The expression vector pQE32 has a region of sequence preceding the multiple cloning site which encodes an initiation methionine followed closely by six histidine residues. These histidine residues form anaffinity tag which enables purification of the polypeptide to which the residues are attached, as insertion of a DNA fragment into the multiple cloning site in frame with the initiation codon leads to the expression of a polypeptide having sixhistidine residues at its amino terminus. This also allows the fusionprotein to bind to a nickel-chelated resin from which it can be subsequently eluted. In our experiments, we have observed that the hexahistidine leader region alters the migra-tion of thepolypeptide to which it is attached, corresponding to an apparent molecular weight increase of approximately 7,000 in SDS-PAGE (data not shown). A peptide, HHPHL PRTRKSSPKS, representing amino acids 36 to 50 of the second ORF of the PVM P gene, was coupled to keyhole limpet hemocyanin (Sigma Chemical Company Limited) by m-maleimidobenzoyl N-hydroxysuccinimide ester (20). The coupled peptide was purified on Sephadex G-25, and rats were injected subcutaneously with an equivalent of 0.5 mg of peptide emulsified in Freund's complete adjuvant and then givenboosters at 3 weekly intervals with a similar quantity of peptide in Freund's incomplete adjuvant. Blood was taken 3 weeks after the third booster and assayed for response of rats to the peptide by Western blot analysis against bacterially expressed PVM P-gene second ORF protein product. The Western blot analysis identified a 30-kDa polypeptide ex-pressed only in the induced cultures (Fig. 4). Preimmune serumfailed torecognize this protein. Taking into account the extrasequences from the expression vector, this suggested that the unmodified second ORF protein (P2) had an apparent molecular weight of 23,000 in contrast to the predicted value of 15,932. The serum from peptide-immunized rats detected a polypeptide with an apparent molecular weight of approxi-mately 23,000 in the PVM-infected cell material which was absent in mock-infected cells (Fig. 4). We believe that the 23-kDa polypeptide identified by the immune serum repre-sents P2, the product of the second ORF. The observed aberrant mobility in SDS-PAGE is consistent with that seen with thebacterially expressed second ORF protein and would explain the apparent absence of the protein in in vitro trans-lations, as itwouldcomigrate with P23.
These dataindicate that the PVM P gene, in common with thePgenes of all other morbilliviruses, paramyxoviruses, and rubulaviruses, is capable of expressing more than one polypep-tide in vivo and iscapable of utilizing more than one reading frame in order to do this. A second ORF has also been observed in the genesencoding the M2 proteins of respiratory syncytial virus, PVM, and TRTV, but the sequences of the
kd 1 2 3 4 5 6 7 8 kd
i-N4
..
"W-430
:0:
oft21.5
21.5b'-FIG. 4. The PVM P-gene second ORF encodes a polypeptide in vivo. Shown is a Western blot of a polyacrylamide gel probed with antiserum raised against a peptide based on the predicted sequence of the P-gene second ORF or with immune serum. Lanes 1 and 3, proteins from pQE32.P2-transformed E. coli probed with preimmune and antipeptide antisera, respectively; lanes 2 and 4, proteins from pQE32-transformed E. coli probed with preimmune and antipeptide antisera, respectively; lanes 5 and 7, proteins from mock-infected BSC1 cells probed with preimmune and antipeptide antisera, respec-tively: lanes 6 and 8, proteins from PVM-infected
BSC1
cells probed with preimmune and antipeptide antisera, respectively. The positions ofmolecular weight standards are shown.predicted proteins are not conserved and the presence of these predicted proteins has not been demonstrated in vivo (7, 8, 17). From the long ORF, the PVM P gene is able to express at least three primary polypeptide products in vivo, namely, the full-length PVM P39 protein and two internally initiated proteins having molecular weights of 26,000 and 23,000. These two internally initiated proteins are translated from the third and fourth AUG codons of the PVM P-gene long ORF, respec-tively, and the proteins are therefore carboxy coterminal. This is similar to the situation reported for Newcastle disease virus (24). Whether P26 and P23 play a role in the functioning of the nucleocapsid complex is unknown. These two internally initi-ated proteins are not capable of binding to the PVM N protein in in vitro binding studies (1). Thus, it seems that the func-tion(s) of these smaller, P-related proteins may not involve the same direct, strong association with the N protein. One of the major polypeptides, with an apparent molecular weight of 35,000, seen in vivo is possibly generated by amino-terminal proteolytic cleavage of either the full-length P39 protein or the internally initiated P37 protein, since it fails to react with an amino-terminus-specific monoclonal antibody (18). In addi-tion, several other P-related polypeptides were identified by Western blot analysis. The origin of these other polypeptides is not clear, but some may represent phosphorylation or other modification intermediates as has been suggested for the Newcastle disease virus P-related polypeptides detected in vivo (24).
The function of the second ORF product (P2) in vivo is unknown. The polypeptide that is translated from the second ORF of the PVM P gene has a predicted molecular weight of 15,932. However, in PVM-infected cells, the polypeptide which we believe to be the product of the second ORF migrates with an apparent molecular weight of 23,000. The reason for the discrepancy between the predicted and observed molecular weights of this protein is not known. The coincidence of the location of the second ORF with a region of amino acid sequence variability in the P39 protein encoded by the long ORF is very striking.
lt!k.: -i
llil. ..k.15.::1. , il
on November 9, 2019 by guest
http://jvi.asm.org/
[image:4.612.353.513.67.202.2]Nucleotide sequence accessionnumber. The sequences re-ported here have beenlodgedwith the EMBL data bank under accession number U09649.
WethankM.Hartley for theprovisionofwheat germ extract. J.B. wasfunded by a research studentship training grant from the Medical Research Council.
REFERENCES
1. Barr, J., and A. J. Easton.Submitted for publication.
la.Barr, J. N., P. Chambers, C. R. Pringle, and A. J. Easton. 1991. Sequence of the majornucleocapsid proteingeneofpneumonia virus of mice: sequence comparisons suggest structural homology between nucleocapsid proteins of pneumoviruses, paramyxovi-ruses, rhabdoviruses andfiloviruses. J. Gen. Virol. 72:677-685. 2. Bellini, W. J., G. Englund,S. Rozenblatt, H. Arnheiter, and C. D.
Richardson.1985. MeaslesvirusPgenecodes for two proteins. J. Virol.53:908-919.
3. Chambers, P., J. Barr, C. R. Pringle, and A. J. Easton. 1990. Molecularcloning of pneumonia virus of mice. J. Virol. 64:1869-1872.
4. Chambers, P., D. A. Matthews, C. R. Pringle, and A. J. Easton. 1990.Thenucleotidesequencesof intergenic regionsbetweennine genesofpneumonia virus ofmice establishthe physical order of these genes intheviral genome.VirusRes. 18:263-270. 5. Chambers,P., C. R. Pringle, and A.J. Easton. 1991.Genes1and
2ofpneumonia virus ofmice encodeproteins which havelittle homology with the 1C and 1B proteins of human respiratory syncytialvirus. J. Gen. Virol. 72:2542-2549.
6. Chambers, P., C. R. Pringle, and A. J. Easton. 1992. Sequence analysis of thefusion glycoprotein ofpneumonia virus ofmice suggests possible conserved secondary structure elements in paramyxovirus fusion glycoproteins.J.Gen. Virol. 73:1717-1724. 7. Chambers, P., C. R. Pringle, and A. J. Easton.Unpublished data. 8. Collins, P.L., and G. W. Wertz. 1985. The envelope-associated 22K protein of human respiratory syncytial virus: nucleotide sequence ofthe mRNA and a related polytranscript. J. Virol. 54:65-71.
9. Galinski,M.S., M. A. Mink, D. M. Lambert, S. L. Wechsler, and M.W. Pons.1986.Molecularcloning and sequence analysisofthe humanparainfluenza3virusRNAencoding the P and C proteins. Virology145:46-60.
10. Higgins, D. G., and P. M. Sharp. 1988. CLUSTAL: a package for performing multiple sequence alignment on a microcomputer. Gene73:237-244.
11. Horsfall, F. L., and M. D.Curnen. 1946. Studies on pneumonia virus ofmice(PVM).J. Exp. Med.83:43-64.
12. Horsfall, F. L., and R. G. Hahn. 1939. ApneumoniavirusofSwiss mice. Proc. Soc. Exp.Biol.Med. 40:684-686.
13. Johnson, P. R., and P. L. Collins. 1990. Sequence comparison of thephosphoprotein mRNAs ofantigenic subgroupsA and Bof human respiratory syncytial virus identifies a highly divergent domainin thepredicted protein. J. Gen. Virol. 71:481-485. 14. Kondo, K., B. Hisanori, M. Tsurudome, M. Kawano, M. Machiko,
andY. Ito. 1990. Sequence analysis ofthe phosphoprotein (P) genesofhumanparainfluenzatype 4Aand 4Bvirusesand RNA editingat transcript of the P genes: the number of Gresidues added isimprecise. Virology178:321-326.
15. Lambden, P. R. 1985. Nucleotide sequence of the respiratory syncytial virusphosphoprotein gene. J. Gen.Virol.66:1607-1612.
16. Ling, R., P.Davis, C. M. Wood, C. R. Pringle, D. Cavanagh, and A.J. Easton.Unpublisheddata.
17. Ling, R., A. J. Easton, and C. R Pringle. 1992. Sequenceanalysis of the 22k, SH and Ggenes ofturkey rhinotracheitis virus and theirintergenic regions reveals a gene order different from that of other pneumoviruses. J. Gen. Virol.73:1709-1715.
18. Ling, R., and C. R. Pringle. 1989. Polypeptides of pneumonia virus of mice. I.Immunological cross-reactions and post-translational modifications.J. Gen.Virol.70:1427-1440.
19. Lopez, J. A., N. Villanueva, J. Melero, and A. Portela. 1988. Nucleotide sequence of the fusion andphosphoproteingenes of humanrespiratory syncytial (RS) virus Long strain: evidence of subtypegeneticheterogeneity. VirusRes.70:249-262.
20. Lui, F. T., M.Zinneker, T. Hamaoka, and D. H. Katz. 1979. New proceduresforpreparationand isolationofconjugatesofproteins and asyntheticcopolymerof D-aminoacidsandimmunochemical characterization of suchconjugates. Biochemistry 18:690-697. 21. Mallipedi, S., and S. K. Samal. 1992. Sequence comparison
betweenthephosphoproteinmRNAsof human and bovine respi-ratorysyncytial virus identifiesadivergent domain in the predicted protein.J.Gen.Virol. 73:2441-2444.
22. Maniatis, T., E. F. Fritsch, and J. SambrookL 1982. Molecular cloning: a laboratory manual. Cold Spring Harbor Laboratory, ColdSpringHarbor, N.Y.
23. Matsuoka, Y., J. Curran, T. Pelet, D. Kolakofsky, R.Ray, and R. W. Compans. 1991.The P gene of humanparainfluenza virus type 1encodes P and Cproteinsbutnot acysteine-richVprotein. J. Virol. 65:3406-3410.
24. McGinnes, L. W., C. McQuain, and T. Morrison. 1988. The P protein and thenon-structural 38K and 29Kproteins ofNewcastle disease virus are derived from the same open reading frame. Virology164:256-264.
25. Pringle, C. R., and R P.Eglin. 1986. Murine pneumoniavirus: seroepidemiological evidence ofwidespread human infection.J. Gen.Virol.67:975-982.
26. Saiki, R K., S.Scharf,F.Faloona, K. B. Mullis, G. T.Horn, H. A. Erlich,and N. Arnheim.1988.Enzymatic amplification of ,B-globin genomicsequences and restriction site analysis for diagnosisof sickle cell anemia. Science 230:1350-1354.
27. Sanger,F., A. R.Coulson,B.G.Barrell, A. J. H.Smith, and B. A. Roe. 1980.Cloninginsingle strandedbacteriophageas anaidto rapidDNAsequencing.J.Mol.Biol.143:161-178.
28. Satake,M., N.Elango, and S. Venkatesan. 1984.Sequence analysis of the respiratory syncytialvirus phosphoprotein gene. J. Virol. 52:991-994.
29. Selwyn, B. J. 1990. Theepidemiology of acute respiratorytract infectionin youngchildren:comparison offindings from several developing countries.Rev.Infect. Dis. 2:5870-5888.
30. Shioda, T.,K.Hidaka, T. Kanda, H. Shibuta, A. Nomoto, and K. Iwasaka. 1983.Sequence of3687 nucleotides from the3' end of the Sendai virus genome RNA and the predicted amino acid sequences of viral NP, P and C proteins. Nucleic Acids Res. 11:7317-7330.
31. Spriggs, M.K.,and P.L.Collins. 1986.Sequenceanalysis ofthe P and Cproteingenesofhumanparainfluenza virustype3: patterns of amino acid sequencehomologyamongparamyxovirus proteins. J.Gen. Virol.67:2705-2719.
32. Studier, F. L., and B. A. Moffatt. 1986. Use ofbacteriophageT7 RNApolymerase to direct selectivehighlevelexpression of cloned genes. J. Mol. Biol. 189:113-130.