Mutational Analysis of Adeno-Associated Virus Type 2 Rep68
Protein Endonuclease Activity on Partially
Single-Stranded Substrates
MICHAEL D. DAVIS,† JIANWEN WU,‡ANDROLAND A. OWENS* Laboratory of Molecular and Cellular Biology, National Institute of Diabetes and
Digestive and Kidney Diseases, Bethesda, Maryland 20892
Received 12 October 1999/Accepted 15 December 1999
The endonuclease activity of the Rep68 and Rep78 proteins (Rep68/78) of adeno-associated virus type 2 (AAV) cuts at the terminal resolution site (trs) within the hairpin structure formed by the AAV inverted terminal repeats. Recent studies suggest that a DNA unwinding function of Rep68/78 may be required for endonuclease activity. We demonstrate that several mutant proteins which are endonuclease negative on a fully duplex hairpin substrate are endonuclease positive on a partially single-stranded hairpin substrate. Trunca-tion analysis revealed that the endonuclease funcTrunca-tion is contained within the first 200 amino acids of Rep68/78. This endonucleolytic cleavage is believed to involve the covalent attachment of Rep68/78 to the trs via a phosphate-tyrosine linkage. A previous report (S. L. Walker, R. S. Wonderling, and R. A. Owens, J. Virol. 71:2722–2730, 1997) suggested that tyrosine 152 was part of the active site. We individually mutated each tyrosine within the first 200 amino acids of the Rep68 moiety of a maltose binding protein-Rep68/78 fusion protein to phenylalanine. Only mutation of tyrosine 156 resulted in a protein incapable of covalent attachment to a partially single-stranded hairpin substrate, suggesting that tyrosine 156 is part of the endonuclease active site.
Adeno-associated virus type 2 (AAV) is a human parvovirus with a single-stranded, linear DNA genome containing in-verted terminal repeats (ITRs), which function as origins of replication (11, 32, 33, 41). AAV is nonpathogenic and usually requires an adenovirus or herpesvirus as a helper for efficient replication (3). The AAVrepgene encodes at least four over-lapping, multifunctional, nonstructural proteins encoded by RNA transcribed from two promoters. Rep68 and Rep78 are encoded by spliced and unspliced transcripts, respectively, from the promoter at map position 5, hence the first 529 amino acids of Rep78 and Rep68 are identical (6, 26, 40, 44). Rep40 and Rep52 are encoded by spliced and unspliced transcripts, respectively, from the promoter at map position 19 (5).
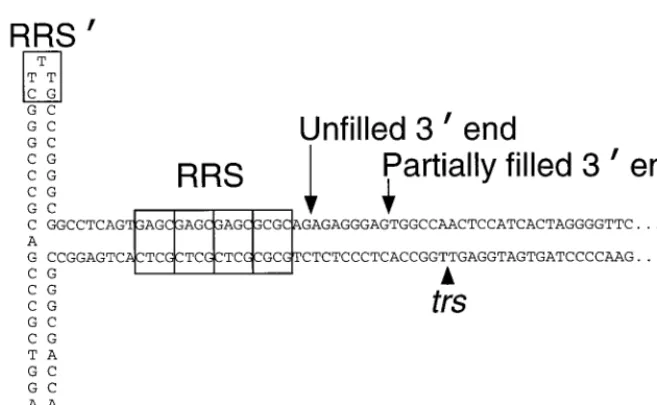
The AAV ITRs are palindromic and fold into hairpin struc-tures (see Fig. 1) which serve as primers for the synthesis of the complementary strand (4, 40). The resulting closed-end inter-mediates are resolved by a process called terminal resolution, which involves a site-specific, strand-specific endonuclease cut at the terminal resolution site (trs), followed by unwinding and replication of the hairpin (13, 37, 39). Rep78 and Rep68 (Rep68/78) display activities which are required for AAV DNA replication including the abilities to bind specifically to Rep recognition sequences (RRSs) within the AAV terminal hairpin DNA (13–15, 25, 28, 52) and to mediate nucleoside triphosphate-dependent, strand-specific nicking at the trs(13, 15, 37). Rep proteins also have nucleoside triphosphate-depen-dent DNA helicase (13, 15, 22) and DNA-RNA helicase (53) activities, as well as ATPase activity (53).
There is strong evidence that Rep68/78 are involved in the preferential integration of AAV genomes into a region on the q arm of human chromosome 19 (1, 10, 18–20, 23, 34, 46, 52). This is the only reported example of site-specific integration in a mammalian virus system. The chromosome 19 preferred in-tegration locus, designatedAAVS1, contains both an RRS and atrs-like sequence which can be cleaved by Rep proteins (18, 46, 52). Linden et al. (23) showed that a 33-bp segment of AAVS1, containing both the RRS and trs-like sequence, is sufficient to target AAV integration into an episome. If either the RRS ortrs-like sequence was mutated, then targeting was lost (23).
Three lines of evidence suggest that the trs endonuclease activity of Rep68/78 requires a Rep68/78 DNA unwinding ac-tivity. First, Snyder et al. (38) demonstrated that a nucleoside triphosphate cofactor is no longer required for Rep68trs en-donuclease activity if the region of thetrs is single stranded. Second, although we have been able to generate several heli-case-positive, endonuclease-negative Rep mutants, we have not been able to generate any helicase-negative, endonuclease-positive (on a fully double-stranded hairpin substrate) mutants (9, 22, 28, 48, 49). Recently, Zhou et al. (56) showed that Rep68 can unwind a blunt-ended, double-stranded DNA sub-strate if it contains an RRS. This apparent linkage between the helicase and endonuclease activities of Rep68/78 has compli-cated the interpretation of mutational analyses intended to identify specific amino acid residues involved in Rep68/78 en-donuclease activity (9, 24, 48, 51). By using AAV hairpin sub-strates in which thetrsis single stranded, we have uncoupled DNA cleavage from DNA unwinding.
We have used maltose binding protein (MBP)-Rep68/78 fusion proteins produced inEscherichia colifor this analysis. Our “wild-type” protein, MBP-Rep68⌬, contains Rep68/78 amino acids 3 through 522, nearly all of the region which is identical between Rep68 and Rep78 (7). MBP-Rep68⌬ has been shown to have all of the in vitro functions of Rep68/78 produced in human cells. It binds specifically to DNA contain-* Corresponding author. Mailing address: Laboratory of Molecular
and Cellular Biology, NIDDK, National Institutes of Health, Bldg. 8, Rm. 310, 8 Center Dr. MSC 0840, Bethesda, MD 20892-0840. Phone: (301) 496-3359. Fax: (301) 402-0053. E-mail: [email protected].
† Present address: Department of Health Administration, City of Detroit, Detroit, MI 48202.
‡ Present address: Department of Medicine, Louisiana State Uni-versity School of Medicine, New Orleans, LA 70112.
2936
on November 9, 2019 by guest
http://jvi.asm.org/
ing RRSs (7–9, 48, 49, 54), hastrsendonuclease (7, 9, 46, 48, 49), helicase (7, 9, 48, 49, 53), and ATPase (53) activities, and can complement Rep-deficient cell extracts in an in vitro AAV replication system (50). Given the ease with which mutant proteins can be generated and purified, we felt that this system was appropriate for identifying sequences important for the endonuclease activity of Rep proteins.
Protein expression. MBP-Rep68 fusion proteins were pro-duced in E. coli containing plasmids encoding these fusion proteins and purified as described previously (7, 53). Protein concentrations were determined by optical density at 225 nm using bovine serum albumin (BSA) standards. The production of MBP-Rep68⌬proteins of the predicted sizes was confirmed by sodium dodecyl sulfate (SDS)-polyacrylamide gel electro-phoresis (PAGE) and Coomassie blue staining (data not shown). All of our mutant MBP-Rep68⌬ proteins were iso-lated at concentrations and purity levels comparable to the wild-type protein.
trsendonuclease assays.The site-specific and strand-specific endonuclease assay was performed as described previously (13), with the modifications indicated below. Plasmid psub201 (31) was used for preparing AAV hairpin DNA in the flop configuration. psub201 contains a modified AAV genome in which each ITR is flanked by anXbaI and aPvuII site. The plasmid was digested with XbaI, followed by dephosphoryla-tion with calf intestine alkaline phosphatase and 32P 5⬘-end radiolabeling using T4 polynucleotide kinase. The resulting products were then digested withPvuII, denatured by heating to 100°C for 6 min, and immediately cooled on ice for 4 min to form radiolabeled AAV unfilled hairpin in which the trs is single stranded (Fig. 1). To make filled hairpin, the unfilled hairpin was treated with Klenow fragment in the presence of dNTPs to fill in the 5⬘overhang. The hairpin DNAs were then purified by nondenaturing PAGE (6% polyacrylamide). For the trs endonuclease assays, 32P 5⬘-end-labeled, filled or un-filled, AAV hairpin DNA (25,000 cpm) was incubated in the presence of MBP-Rep68⌬or mutant proteins in a 30-l reac-tion volume containing 25 mM HEPES/KOH (pH 7.5), 5 mM MgCl2, 1 mM dithiothreitol, 0.3g BSA, and 0.4 mM ATP.
The reaction mixtures were incubated for 1 h at 37°C and terminated by addition of 15l of gel loading buffer (0.5% SDS, 50 mM EDTA, 40% [vol/vol] glycerol, 0.1% [wt/vol] bromophenol blue, 0.1% [wt/vol] xylene cyanol) with subse-quent boiling to release the nicked fragment. The reaction products were resolved on a nondenaturing 6% polyacrylamide gel, which was then dried and autoradiographed.
[image:2.612.140.469.69.272.2]We first wished to test the hypothesis that many of our MBP-Rep68⌬ proteins, containing mutations previously re-ported to eliminatetrsendonuclease activity on a filled hairpin substrate, could nick an unfilled hairpin substrate in which the trs is single stranded. Plasmids encoding MBP-Rep68⌬, the D40A-D42A-D44A, E83A-K84A-E86A, Y121F, K146A-D149A-E150A, Y152F, G334A, G339A, K340H, T341A, E378A-E379A-K381A, D402A-K404A-K406A, V418S, D443A-K447A, D455A-D457A, and K463A-E465A-K467A-D468A mutant MBP-Rep proteins and MBP-LacZ have been reported previ-ously (7, 9, 48, 49, 53). The designations for proteins with amino acid substitutions are the single-letter code for the wild-type amino acid, followed by the Rep68/78 amino acid number, followed by the single-letter code for the new amino acid. The plasmid encoding the mutated MBP-Rep68⌬protein with a deletion of Rep68/78 methionine 225 (M225dl) was made by replacing thePstI-BamHI fragment of therepgene of pMBP-Rep68⌬ with the corresponding fragment from pHIV-Rep/ M225dl(28).
Figure 2 shows that the Y121F, M225dl, G334A, G339A, K340H, T341A, E378A-E379A-K381A, D402A-K404A-K406A, V418S, D443A-K447A, D455A-D457A, and K463A-E465A-K467A-D468A mutant proteins all had levels oftrs endonucle-ase activity similar to the wild-type fusion protein on the un-filled hairpin. The Y152F protein had low but detectable endonuclease activity. The D40A-D42A-D44A, E83A-K84A-E86A, and K146A-D149A-E150A proteins had no detectable trsendonuclease activity. An MBP-LacZ fusion protein (53) also showed no activity.
It is important to note that only proteins with alterations in the first 200 amino acid residues of the Rep68/78 moiety were devoid of endonuclease activity on the unfilled hairpin sub-FIG. 1. AAV ITR hairpin DNA (“flop” configuration). The positions of the primary Rep68/78 recognition sequence (RRS) (52) and the secondary binding site for Rep68/78 (RRS⬘) (30, 55) are within the labeled rectangles. The individual imperfect GAGC repeats of the RRS are indicated by subdivisions of its rectangle. The positions of the terminal resolution site (trs) (13) and the 3⬘ends of unfilled hairpin DNA and partially filled hairpin DNA are also indicated. Fifty-four base pairs at the right end of the hairpin sequence are indicated by dots.
on November 9, 2019 by guest
http://jvi.asm.org/
strate. Care was taken in the design of the D40A-D42A-D44A, E83A-K84A-E86A, and K146A-D149A-E150A mutations to minimize the possibility of inducing global misfolding of the proteins (2, 9). These proteins were previously demonstrated to have significant levels of DNA helicase activity (9), indicat-ing specific defects in functions required for endonuclease ac-tivity. The D40A-D42A-D44A mutant is also defective in hair-pin DNA binding and self-association (9). Previous mutational analyses suggest that the ability of Rep68/78 to form stable oligomers on hairpin DNA is required for both optimal DNA binding and trs endonuclease activity (9, 51). It is possible, however, that the endonuclease defects in the D40A-D42A-D44A, E83A-K84A-E86A, and K146A-D149A-E150A pro-teins could be the result of disruption of the secondary or tertiary structure of regions nearby in either the primary se-quence or the three-dimensional structure of the protein. The E83A-K84A-E86A mutation is near a sequence motif con-served in a superfamily of replication initiation proteins which includes Rep68/78 (17). This HUHUUU sequence (in which H represents histidine and U represents an amino acid with a bulky hydrophobic side chain) is believed to be involved in the coordination of metal ions (17). Thetrsendonuclease activity is known to be dependent on the presence of magnesium ions (13). The sequence in Rep68/78 is HMHVLV (amino acids 90 to 95). Mutation of H90 or H92 to alanine has been shown previously to knock out thetrsendonuclease activity of Rep78 on a filled hairpin substrate (45).
All of our point mutations that resulted in MBP-Rep fusion proteins which were endonuclease negative on a filled hairpin, but had endonuclease levels comparable to that of MBP-Rep68⌬ on an unfilled hairpin (Fig. 2), also resulted in pro-teins which were helicase negative (Table 1) (7, 9, 48, 49). We hypothesize that many of the mutant Rep68/78 proteins pre-viously reported to be endonuclease negative (7, 9, 24, 28, 48, 49, 51), especially those with mutations outside the first 200 amino acids of Rep68/78, are endonuclease negative on a filled hairpin because of an inability to unwind DNA and not be-cause of an inability to cleave at thetrs. Smith and Kotin (36) showed that amino acids 225 to 621 of Rep78 are sufficient for DNA helicase activity. Rep68/78 belong to a superfamily of
ATPase/helicase proteins (16). All of the sequence motifs con-served within this superfamily are found within Rep68/78 amino acids 330 through 422 (16). The G334A, G339A, K340H, T341A, E378A-E379A-K381A, D402A-K404A-K406A, and V418S mutations alter conserved amino acids within these helicase motifs.
[image:3.612.149.457.76.256.2]The D443A-K447A, D455A-D457A, and K463A-E465A-K467A-D468A mutations alter amino acids within a putative 3,4-heptad repeat motif which is believed to be involved in Rep68/78 oligomerization (9). The proteins containing these mutations are endonuclease negative on a filled hairpin sub-strate and helicase negative (Table 1) (9). The ability of these
FIG. 2.trsendonuclease assays of wild-type and mutant MBP-Rep68⌬fusion proteins on the unfilled hairpin substrate.trsendonuclease assays were performed (as detailed in the text, except that no ATP was added) with the mutant fusion proteins indicated above each lane. Each sample contained 25,000 cpm of 5⬘32P-end-labeled unfilled AAV terminal repeat hairpin DNA. Samples contained either no protein, 1.0g of MBP-LacZ (LacZ), or 1.0g of the indicated mutant protein. The wild-type (W-T) lanes contained 1.0 or 0.5 g of MBP-Rep68⌬. All reaction mixtures were incubated at 37°C for 60 min, boiled, and resolved on a nondenaturing 6% polyacrylamide gel. The position of the released cleavage product is indicated on the right.
TABLE 1. Activities of MBP-Rep68⌬substitution mutant proteins
MBP-Rep protein
Activitya
trsendonuclease
DNA helicase Unfilled
hairpin hairpinFilled
Wild type ⫹ ⫹ ⫹
D40A-D42A-D44A ⫺ ⫺ ⫾
E83A-K84A-E86A ⫺ ⫺ ⫹
Y121F ⫹ ⫺ ⫺
K146A-D149A-E150A ⫺ ⫺ ⫹
Y152F ⫾ ⫺ ⫹
Y156F ⫺ ⫺ ⫹
Y152F/Y156F ⫺ ⫺ ⫹
G334A ⫹ ⫺ ⫺
G339A ⫹ ⫺ ⫺
K340H ⫹ ⫺ ⫺
T341A ⫹ ⫺ ⫺
E378A-E379A-K381A ⫹ ⫺ ⫺
D402A-K404A-K406A ⫹ ⫺ ⫺
V418S ⫹ ⫺ ⫺
D443A-K447A ⫹ ⫺ ⫺
D455A-D457A ⫹ ⫺ ⫺
K463A-E465A-K467A-D468A ⫹ ⫺ ⫺
a⫹, activity comparable to wild type;⫾, low but detectable activity;⫺, no
detectable activity.
on November 9, 2019 by guest
http://jvi.asm.org/
[image:3.612.310.549.490.710.2]mutated proteins to cleave the unfilled hairpin substrate (Fig. 2) suggests that the proposed role of oligomerization, medi-ated by this motif, intrs endonuclease activity (9) is not re-quired for the actual DNA cleavage event.
We next wished to determine the minimal portion of the Rep68/78 protein sufficient fortrsendonuclease activity on an unfilled hairpin substrate. We made a series of C-terminal truncations of the Rep68/78 moiety of MBP-Rep68⌬. Plasmids encoding C-terminally truncated versions of MBP-Rep68⌬
were constructed by first amplifying a section of pMBP-Rep68⌬(7) using PCR. Part of each downstream primer was complementary to the region where the truncation was to be made, and the other part included a HindIII site. To make truncations to Rep68/78 amino acid 150, 200, 251, 300, 352, or 400, the upstream primer was complementary to the region encoding Rep68/78 amino acids 3 through 9. To make trunca-tions to Rep68/78 amino acid 150, 200, 251, or 300, the PCR product was trimmed withPstI andHindIII and used to replace thePstI-HindIII fragment of pMBP-Rep68⌬, which contains a portion of therepgene. To make truncations to amino acid 352 or 400, the PCR product was trimmed with BamHI and HindIII and used to replace the correspondingBamHI-HindIII fragment of pMBP-Rep68⌬.
To construct plasmids encoding MBP fusion proteins trun-cated to Rep68/78 amino acid 442, 447, 460, 466, or 476, the upstream primer was complementary to the region encoding Rep68/78 amino acids 357 to 364, which is just upstream of a SalI site. The PCR product was trimmed withSalI andHindIII and used to replace the SalI-HindIII fragment of pMBP-Rep68⌬, which contains a portion of therepgene.
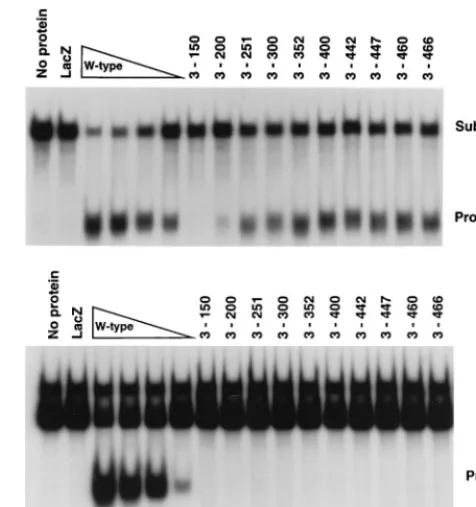
Figure 3A shows that endonuclease activity was detected with a mutant protein containing amino acids 3 to 200 of Rep68/78, but a protein containing amino acids 3 to 150 had no detectable activity on an unfilled hairpin. This confirms that the first 200 amino acids of Rep68/78 contain all of the essen-tial components for cleavage. Our C-terminal truncation mu-tants which were truncated to Rep68/78 amino acid 466 or less lacked the ability to nick the filled hairpin substrate (Fig. 3B). This is consistent with previous data using truncated Rep pro-teins produced in a rabbit reticulocyte in vitro translation sys-tem (51).
Covalent attachment assays. Snyder et al. showed that Rep68 can become covalently attached to the 5⬘phosphate of a T residue at the nicking site in AAV hairpin DNA and that this linkage involves a tyrosine residue (37). MBP-Rep68⌬has also previously been shown to possess the ability to covalently attach to AAV hairpin DNA (8). It is unclear whether it is biologically important that Rep68/78 remain covalently at-tached to the DNA or if this attachment simply represents a trapped intermediate in the cleavage process. There have been reports of Rep78 being tightly associated with virus particles (21, 29), but no function has been determined for these at-tached Rep proteins. In any case, this covalently attaching tyrosine should be an important component of the endonucle-ase active site.
Our previous analysis suggested that Y152 is the Rep68/78 active-site tyrosine (48). This was based on the observation that of the three tyrosines conserved between AAV Rep68/78 and the human herpesvirus 6 Rep homologue (42), the deduced product of the only gene known to be able to substitute for the AAVrepgene in AAV replication (43), only mutation of Y152 eliminated the ability of MBP-Rep68⌬to nick AAV hairpin DNA with a double-strandedtrs(48). The Y152F mutant also was helicase positive, indicating a specific defect in the endo-nuclease function. Tyrosine 156 of Rep68/78, however, aligns best with the covalently linking tyrosine of the NS1 protein of
minute virus of mice (MVM), the only parvovirus nonstruc-tural protein for which the active-site tyrosine has been deter-mined (27).
In our previous report, we had technical difficulties with the production of our Y156F mutant (48). This first batch of the Y156F protein had a large amount of a low-molecular-weight protein contaminant which apparently interfered with our analysis. Subsequent protein preparations from bacteria carry-ing the same expression plasmid also had this low-molecular-weight contaminant. For the present work, we reconstructed the plasmid encoding the Y156F mutant protein. Protein prep-arations from bacteria carrying the reconstructed Y156F plas-mid showed no such contaminant (data not shown).
Since it is possible that Y152 and Y156 both participate in the endonuclease reaction, we also made a double mutant (Y152F/Y156F). Two tyrosines separated by three amino acids have been reported to be involved in the DNA cleavage reac-tion performed by theX174 gene A protein (47). It should be noted that the MVM NS1 protein has a second tyrosine resi-due right next to the one which forms the covalent attachment (27, 35). We also individually mutated the other tyrosines in the first 200 amino acids of the Rep68/78 moiety, in the context of the full-length MBP-Rep68⌬protein.
Plasmids encoding the Y121F, Y152F, and Y175F mutant MBP-Rep proteins have been reported previously (48). Plas-mids encoding the Y5F, Y88F, Y156F, and Y152F/Y156F mu-tant proteins were made by an overlap extension PCR method (12). Briefly, two pairs of primers were used to direct synthesis of mutant DNA fragments. These fragments, which
over-FIG. 3. Rep68/78 amino acids 3 to 200 are sufficient fortrsendonuclease activity on unfilled hairpin DNA.trsendonuclease assays were performed (as detailed in the text) with the truncated fusion proteins indicated above each lane. Samples contained either no protein, 1.0g of MBP-LacZ (LacZ), or 1.0g of the fusion protein with the indicated truncation. The wild-type (W-type) lanes contained 1.0, 0.5, 0.25, or 0.125g of MBP-Rep68⌬. All reaction mixtures were incubated at 37°C for 60 min, boiled, and resolved by nondenaturing PAGE (6% polyacrylamide). The positions of the substrate and released cleavage product are indicated on the right. Each sample contained 25,000 cpm of 5⬘32 P-end-labeled unfilled (A) or filled (B) AAV terminal repeat hairpin DNA.
on November 9, 2019 by guest
http://jvi.asm.org/
[image:4.612.313.551.73.326.2]lapped by at least 25 bp, were gel purified and subsequently used for overlap extension PCR using 5⬘and 3⬘flanking prim-ers. The resulting amplified product was gel purified and di-gested with appropriate restriction endonucleases, generating a fragment which was substituted for the corresponding frag-ment within the parent plasmid, pMBP-Rep68⌬(7, 12). In the cases of the Y5F and Y88F mutations, aBglII-SacII fragment containing parts of themalE(MBP) andrepgenes was gener-ated. In the case of the Y156F mutation, aPstI-BamHI frag-ment comprised of a portion of therep gene was generated. The Y152F/Y156F mutation was made using the same primers as the Y156F mutation, but the initial PCR template was pMBP-Rep68⌬/Y152F (48) instead of pMBP-Rep68⌬. PCRs were carried out using the thermostablePfuDNA polymerase, which has a high fidelity (1.3⫻ 10⫺6 errors/base pair/cycle) (Stratagene, La Jolla, Calif.). The presence of mutations in the repcoding region was confirmed by DNA sequencing.
[image:5.612.319.547.68.442.2]For our covalent attachment assay, we needed a 3⬘ -end-labeled hairpin DNA in which thetrswas still single stranded. We therefore partially filled the AAV hairpin, using only32 P-labeled A and unP-labeled G nucleotides (Fig. 1). This partially filled hairpin DNA was then purified by nondenaturing PAGE (6% polyacrylamide). Gel-purified, partially filled, 3⬘32 P-end-labeled AAV terminal repeat hairpin DNA (25,000 cpm) was incubated in the presence of MBP-Rep68⌬or mutant proteins under the standard trs endonuclease assay conditions. Some samples were incubated for an additional 10 min in the pres-ence of 1g of proteinase K at 37°C. A one-fourth volume of 5⫻ SDS gel-loading buffer (125 mM Tris-HCl [pH 7.5], 5% [wt/vol] SDS, 50% [vol/vol] glycerol, 0.25% [wt/vol] bromophe-nol blue, 1% [vol/vol]-mercaptoethanol) was then added to each sample. The reaction products were then boiled for 7 min and resolved on a 24-cm-long SDS–5% polyacrylamide gel. The gel was then soaked for 30 min in a fixing solution con-taining 20% methanol, 10% acetic acid, and 5% glycerol; dried; and autoradiographed.
Figure 4 shows that in the presence of MBP-Rep68⌬, there is a radiolabeled species which migrates between the 147- and 241-kDa protein standards in SDS-PAGE. This species mi-grates slower than the predominant bands seen with the MBP-LacZ or no protein negative controls. This species does not form in the absence of MgCl2, which is known to be required fortrsendonuclease activity, and is eliminated by proteinase K treatment. Based on these observations, we conclude that this band represents the previously described covalent complex between MBP-Rep68⌬ and the hairpin DNA (8). Covalent attachment of MBP-Rep68⌬to the partially filled hairpin did not require ATP. The Y5F, Y88F, Y121F, and Y175F proteins covalently attached to the hairpin comparably to the wild-type MBP-Rep68⌬.
Only mutation of Y156 completely disrupted the ability of the protein to covalently attach to the partially filled hairpin substrate. The data implicate Y156 as an active-site tyrosine. The Y152F protein showed much less covalent attachment than the wild-type protein. It is unclear if this means that Y152 can function as a secondary site of covalent attachment or if its mutation disrupts the ability of Y156 to attach. The nearby K146A-D149A-E150A mutation also resulted in a protein that is defective in nicking (Fig. 2) and therefore is presumably defective in covalent attachment as well.
Endonuclease and helicase assays on the Y156F protein.We next wished to examine thetrsendonuclease (on unfilled hair-pin) and DNA helicase activities of the Y156F protein. Our previous analysis (48), with the contaminated Y156F protein preparation, showed no detectabletrsendonuclease (on a filled hairpin substrate) or DNA helicase activity. The new Y156F
protein preparation, used in the present study, lacks endonu-clease activity (Fig. 5A).
Helicase assays were performed under the conditions devel-oped by Im and Muzyczka (13), with modifications described by Kyo¨stio¨ and Owens (22). The DNA helicase substrate, which consisted of a radiolabeled 26-mer annealed to single-stranded M13 DNA, was prepared as described previously (13). The new Y156F protein preparation has helicase activity comparable to that of MBP-Rep68⌬(Fig. 5B), indicating that the Y156F mutation does not cause a global disruption of the protein’s structure. The Y152F/Y156F double mutant per-formed identically to the Y156F mutant (Fig. 5).
Elucidation of the mechanism of Rep68/78 endonuclease activity is key to understanding Rep-mediated preferential
in-FIG. 4. Covalent attachment assays of wild-type and mutant MBP-Rep68⌬
fusion proteins. Covalent attachment assays were performed as detailed in the text. Each sample contained 25,000 cpm of 3⬘32P-end-labeled partially filled AAV terminal repeat hairpin DNA. Samples contained either no protein, 3.0g of MBP-LacZ (LacZ), or 3.0g of the mutant fusion protein indicated above each lane. The descending wild-type (W-type) lanes contained 3.0, 1.5, 0.75, or 0.375g of MBP-Rep68⌬. Contents of other lanes are as follows: W-type w/o ATP, 3.0g of MBP-Rep68⌬in a reaction mixture without ATP; W-type w/o Mg2⫹, 3.0g of MBP-Rep68⌬ in a reaction mixture without magnesium;
W-type⫹Proteinase K, 3.0g of MBP-Rep68⌬in a reaction mixtures which was incubated for an additional 10 min in the presence of 1g of proteinase K at 37°C. Reaction products were resolved by SDS-PAGE (5% polyacrylamide). The positions of the free substrate and covalently attached product are indicated on the right. Molecular weight markers are indicated on the left.
on November 9, 2019 by guest
http://jvi.asm.org/
tegration and replication of the AAV genome. The use of partially single-stranded substrates should accelerate determi-nation of the portions of Rep68/78 directly involved in DNA cleavage.
We thank Catherine McKeon, Nancy Nossal, and Robert Craigie for critical reading of the manuscript. We also thank Ramani Wonderling and Scotty Walker for technical assistance.
REFERENCES
1.Balague´, C., M. Kalla, and W. W. Zhang.1997. Adeno-associated virus Rep78 protein and terminal repeats enhance integration of DNA sequences into the cellular genome. J. Virol.71:3299–3306.
2.Bordo, D., and P. Argos.1991. Suggestions for “safe” residue substitutions in site-directed mutagenesis. J. Mol. Biol.217:721–729.
3.Carter, B. J.1990. Adeno-associated virus helper functions, p. 255–282.InP. Tijssen (ed.), Handbook of parvoviruses, vol. I. CRC Press, Inc., Boca Raton, Fla.
4.Carter, B. J., E. Mendelson, and J. P. Trempe.1990. AAV DNA replication, integration, and genetics, p. 169–226. InP. Tijssen (ed.), Handbook of parvoviruses, vol. I. CRC Press, Inc., Boca Raton, Fla.
5.Chejanovsky, N., and B. J. Carter.1989. Mutagenesis of an AUG codon in the adeno-associated virus rep gene: effects on viral DNA replication. Vi-rology173:120–128.
6.Chejanovsky, N., and B. J. Carter.1989. Replication of a human parvovirus nonsense mutant in mammalian cells containing an inducible amber sup-pressor. Virology171:239–247.
7.Chiorini, J. A., M. D. Weitzman, R. A. Owens, E. Urcelay, B. Safer, and R. M. Kotin.1994. Biologically active Rep proteins of adeno-associated virus type 2 produced as fusion proteins inEscherichia coli. J. Virol.68:797–804. 8.Chiorini, J. A., S. M. Wiener, R. A. Owens, S. R. M. Kyo¨stio¨, R. M. Kotin,
and B. Safer.1994. Sequence requirements for stable binding and function of Rep68 on the adeno-associated virus type 2 inverted terminal repeats. J. Virol.68:7448–7457.
9.Davis, M. D., R. S. Wonderling, S. L. Walker, and R. A. Owens.1999. Analysis of the effects of charge cluster mutations in adeno-associated virus Rep68 protein in vitro. J. Virol.73:2084–2093.
10. Giraud, C., E. Winocour, and K. I. Berns.1994. Site-specific integration by adeno-associated virus is directed by a cellular DNA sequence. Proc. Natl. Acad. Sci. USA91:10039–10043.
11. Hauswirth, W. W., and K. I. Berns.1977. Origin and termination of adeno-associated virus DNA replication. Virology78:488–499.
12. Ho, S. N., H. D. Hunt, R. M. Horton, J. K. Pullen, and L. R. Pease.1989. Site-directed mutagenesis by overlap extension using the polymerase chain reaction. Gene77:51–59.
13. Im, D. S., and N. Muzyczka.1990. The AAV origin binding protein Rep68 is an ATP-dependent site-specific endonuclease with DNA helicase activity. Cell61:447–457.
14. Im, D. S., and N. Muzyczka.1989. Factors that bind to adeno-associated virus terminal repeats. J. Virol.63:3095–3104.
15. Im, D. S., and N. Muzyczka.1992. Partial purification of adeno-associated virus Rep78, Rep52, and Rep40 and their biochemical characterization. J. Virol.66:1119–1128.
16. Koonin, E. V.1993. A common set of conserved motifs in a vast variety of putative nucleic acid-dependent ATPases including MCM proteins involved in the initiation of eukaryotic DNA replication. Nucleic Acids Res.21:2541– 2547.
17. Koonin, E. V., and T. V. Ilyina.1993. Computer-assisted dissection of rolling circle DNA replication. BioSystems30:241–268.
18. Kotin, R. M., R. M. Linden, and K. I. Berns.1992. Characterization of a preferred site on human chromosome 19q for integration of adeno-associ-ated virus DNA by non-homologous recombination. EMBO J.11:5071–5078. 19. Kotin, R. M., J. C. Menninger, D. C. Ward, and K. I. Berns.1991. Mapping and direct visualization of a region-specific viral DNA integration site on chromosome 19q13-qter. Genomics10:831–834.
20. Kotin, R. M., M. Siniscalco, R. J. Samulski, X. Zhu, L. Hunter, C. A. Laughlin, S. McLaughlin, N. Muzyczka, M. Rocchi, and K. I. Berns.1990. Site-specific integration by adeno-associated virus. Proc. Natl. Acad. Sci. USA87:2211–2215.
21. Kube, D. M., S. Ponnazhagan, and A. Srivastava.1997. Encapsidation of adeno-associated virus type 2 Rep proteins in wild-type and recombinant progeny virions: Rep-mediated growth inhibition of primary human cells. J. Virol.71:7361–7371.
22. Kyo¨stio¨, S. R. M., and R. A. Owens.1996. Identification of mutant adeno-associated virus Rep proteins which are dominant-negative for DNA heli-case activity. Biochem. Biophys. Res. Commun.220:294–299.
23. Linden, R. M., E. Winocour, and K. I. Berns.1996. The recombination signals for adeno-associated virus site-specific integration. Proc. Natl. Acad. Sci. USA93:7966–7972.
24. McCarty, D. M., T. H. Ni, and N. Muzyczka.1992. Analysis of mutations in adeno-associated virus Rep protein in vivo and in vitro. J. Virol.66:4050– 4057.
25. McCarty, D. M., J. H. Ryan, S. Zolotukhin, X. Zhou, and N. Muzyczka.1994. Interaction of the adeno-associated virus Rep protein with a sequence within the A palindrome of the viral terminal repeat. J. Virol.68:4998–5006. 26. Mendelson, E., J. P. Trempe, and B. J. Carter.1986. Identification of the
trans-acting Rep proteins of adeno-associated virus by antibodies to a syn-FIG. 5. The Y156F protein lackstrsendonuclease activity but retains DNA
helicase activity. Assays were performed (as detailed in the text) with MBP-Rep68⌬(W-type) or the mutant fusion proteins indicated above each lane. MBP-LacZ (LacZ) was included as a negative control. Samples contained either no protein or the amount of each protein (in micrograms) indicated below each lane. (A)trsendonuclease assays. Each sample contained 25,000 cpm of 5⬘
32P-end-labeled unfilled AAV hairpin DNA. All reaction mixtures were incu-bated at 37°C for 60 min, boiled, and resolved on a nondenaturing 6% polyacryl-amide gel. The positions of the substrate and released cleavage product are indicated on the right. (B) DNA helicase assays. Each sample contained 25,000 cpm of32P-labeled partial duplex substrate. The sample marked “Boiled” con-tained no MBP fusion protein and was heated to 100°C for 5 min. All other reaction mixtures were incubated at 24°C for 35 min and resolved on a nonde-naturing 6% polyacrylamide gel. The positions of the substrate and released 26-mer product are indicated on the right.
on November 9, 2019 by guest
http://jvi.asm.org/
[image:6.612.62.280.69.533.2]thetic oligopeptide. J. Virol.60:823–832.
27.Nu¨esch, J. P. F., S. F. Cotmore, and P. Tattersall.1995. Sequence motifs in the replicator protein of parvovirus MVM essential for nicking and covalent attachment to the viral origin: identification of the linking tyrosine. Virology 209:122–135.
28.Owens, R. A., M. D. Weitzman, S. R. M. Kyo¨stio¨, and B. J. Carter.1993. Identification of a DNA-binding domain in the amino terminus of adeno-associated virus Rep proteins. J. Virol.67:997–1005.
29.Prasad, K. M. R., and J. P. Trempe.1995. The adeno-associated virus Rep78 protein is covalently linked to viral DNA in a preformed virion. Virology 214:360–370.
30.Ryan, J. H., S. Zolotukhin, and N. Muzyczka.1996. Sequence requirements for binding of Rep68 to the adeno-associated virus terminal repeats. J. Virol. 70:1542–1553.
31.Samulski, R. J., L. S. Chang, and T. Shenk.1987. A recombinant plasmid from which an infectious adeno-associated virus genome can be excised in vitro and its use to study viral replication. J. Virol.61:3096–3101. 32. Samulski, R. J., A. Srivastava, K. I. Berns, and N. Muzyczka.1983. Rescue
of adeno-associated virus from recombinant plasmids: gene correction within the terminal repeats of AAV. Cell33:135–143.
33. Senapathy, P., J. D. Tratschin, and B. J. Carter.1984. Replication of adeno-associated virus DNA. Complementation of naturally occurringrep⫺
mu-tants by a wild-type genome or anori⫺mutant and correction of terminal
palindrome deletions. J. Mol. Biol.179:1–20.
34.Shelling, A. N., and M. G. Smith.1994. Targeted integration of transfected and infected adeno-associated virus vectors containing the neomycin resis-tance gene. Gene Ther.1:165–169.
35.Skiadopoulos, M. H., and E. A. Faust.1993. Mutational analysis of con-served tyrosines in the NS-1 protein of the parvovirus minute virus of mice. Virology194:509–517.
36.Smith, R. H., and R. M. Kotin.1998. The Rep52 gene product of adeno-associated virus is a DNA helicase with 3⬘-to-5⬘polarity. J. Virol.72:4874– 4881.
37.Snyder, R. O., D. S. Im, and N. Muzyczka.1990. Evidence for covalent attachment of the adeno-associated virus (AAV) Rep protein to the ends of the AAV genome. J. Virol.64:6204–6213.
38. Snyder, R. O., D. S. Im, T. Ni, X. Xiao, R. J. Samulski, and N. Muzyczka. 1993. Features of the adeno-associated virus origin involved in substrate recognition by the viral Rep protein. J. Virol.67:6096–6104.
39. Snyder, R. O., R. J. Samulski, and N. Muzyczka.1990. In vitro resolution of covalently joined AAV chromosome ends. Cell60:105–113.
40. Srivastava, A., E. W. Lusby, and K. I. Berns.1983. Nucleotide sequence and organization of the adeno-associated virus 2 genome. J. Virol.45:555–564. 41. Straus, S. E., E. D. Sebring, and J. A. Rose.1976. Concatemers of alternating
plus and minus strands are intermediates in adenovirus-associated virus DNA synthesis. Proc. Natl. Acad. Sci. USA73:742–746.
42. Thomson, B. J., S. Efstathiou, and R. W. Honess.1991. Acquisition of the
human adeno-associated virus type-2repgene by human herpesvirus type-6. Nature (London)351:78–80.
43. Thomson, B. J., F. W. Weindler, D. Gray, V. Schwaab, and R. Heilbronn. 1994. Human herpesvirus 6 (HHV-6) is a helper virus for adeno-associated virus type 2 (AAV-2) and the AAV-2 rep gene homologue in HHV-6 can mediate AAV-2 DNA replication and regulate gene expression. Virology 204:304–311.
44. Trempe, J. P., E. Mendelson, and B. J. Carter.1987. Characterization of adeno-associated virus rep proteins in human cells by antibodies raised againstrepexpressed inEscherichia coli. Virology161:18–28.
45. Urabe, M., Y. Hasumi, A. Kume, R. T. Surosky, G. J. Kurtzman, K. Tobita, and K. Ozawa.1999. Charged-to-alanine scanning mutagenesis of the N-terminal half of adeno-associated virus type 2 Rep78 protein. J. Virol.73: 2682–2693.
46. Urcelay, E., P. Ward, S. M. Wiener, B. Safer, and R. M. Kotin.1995. Asymmetric replication in vitro from a human sequence element is depen-dent on adeno-associated virus Rep protein. J. Virol.69:2038–2046. 47. van Mansfeld, A. D. M., H. A. A. M. van Teeffelen, P. D. Baas, and H. S.
Jansz.1986. Two juxtaposed tyrosyl-OH groups participate inX174 gene A protein catalysed cleavage and ligation of DNA. Nucleic Acids Res.14:4229– 4238.
48. Walker, S. L., R. S. Wonderling, and R. A. Owens.1997. Mutational analysis of the adeno-associated virus Rep68 protein: identification of critical resi-dues necessary for site-specific endonuclease activity. J. Virol.71:2722–2730. 49. Walker, S. L., R. S. Wonderling, and R. A. Owens.1997. Mutational analysis of the adeno-associated virus type 2 Rep68 protein helicase motifs. J. Virol. 71:6996–7004.
50. Ward, P., E. Urcelay, R. Kotin, B. Safer, and K. I. Berns.1994. Adeno-associated virus DNA replication in vitro: activation by a maltose binding protein/Rep 68 fusion protein. J. Virol.68:6029–6037.
51. Weitzman, M. D., S. R. M. Kyo¨stio¨, B. J. Carter, and R. A. Owens.1996. Interaction of wild-type and mutant adeno-associated virus (AAV) Rep proteins on AAV hairpin DNA. J. Virol.70:2440–2448.
52. Weitzman, M. D., S. R. M. Kyo¨stio¨, R. M. Kotin, and R. A. Owens.1994. Adeno-associated virus (AAV) Rep proteins mediate complex formation between AAV DNA and its integration site in human DNA. Proc. Natl. Acad. Sci. USA91:5808–5812.
53. Wonderling, R. S., S. R. M. Kyo¨stio¨, and R. A. Owens.1995. A maltose-binding protein/adeno-associated virus Rep68 fusion protein has DNA-RNA helicase and ATPase activities. J. Virol.69:3542–3548.
54. Wonderling, R. S., and R. A. Owens.1997. Binding sites for adeno-associated virus Rep proteins within the human genome. J. Virol.71:2528–2534. 55. Wu, J., M. D. Davis, and R. A. Owens.1999. Factors affecting the terminal
resolution site endonuclease, helicase and ATPase activities of adeno-asso-ciated virus type 2 Rep proteins. J. Virol.73:8235–8244.
56. Zhou, X., I. Zolotukhin, D. S. Im, and N. Muzyczka.1999. Biochemical characterization of adeno-associated virus Rep68 DNA helicase and ATPase activities. J. Virol.73:1580–1590.