Copyright © 2001, American Society for Microbiology. All Rights Reserved.
Different Levels of T-Cell Receptor Triggering Induce Distinct
Functions in Hepatitis B and Hepatitis C Virus-Specific
Human CD4
⫹
T-Cell Clones
HELMUT M. DIEPOLDER,1,2* NORBERT H. GRUENER,1J. TILMAN GERLACH,1,2
MARIA-CHRISTINA JUNG,1,2EDDY A. WIERENGA,3ANDGERD R. PAPE1,2
Institute for Immunology, University of Munich, 80336 Munich,1and Department of Medicine II, Klinikum Grosshadern,
University of Munich, 81377 Munich,2Germany, and Laboratory of Cell Biology and Histology,
University of Amsterdam, Amsterdam, The Netherlands3
Received 12 January 2001/Accepted 24 May 2001
CD4ⴙT cells play a major role in the host defense against viruses and intracellular microbes. During the
natural course of such an infection, specific CD4ⴙT cells are exposed to a wide range of antigen concentrations
depending on the body compartment and the stage of disease. While epitope variants trigger only subsets of T-cell effector functions, the response of virus-specific CD4ⴙT cells to various concentrations of the wild-type
antigen has not been systematically studied. We stimulated hepatitis B virus core- and hepatitis C virus NS3-specific CD4ⴙ T-cell clones which had been isolated from patients with acute hepatitis during viral
clearance with a wide range of specific antigen concentrations and determined the phenotypic changes and the induction of T-cell effector functions in relation to T-cell receptor internalization. A low antigen concentration induced the expression of T-cell activation markers and adhesion molecules in CD4ⴙT-cell clones in the
absence of cytokine secretion and proliferation. The expression of CD25, HLA-DR, CD69, and intercellular cell adhesion molecule 1 increased as soon as T-cell receptor internalization became detectable. A 30- to 100-fold-higher antigen concentration, corresponding to the internalization of 20 to 30% of T-cell receptor molecules, however, was required for the induction of proliferation as well as for gamma interferon and interleukin-4 secretion. These data indicate that virus-specific CD4ⴙT cells can respond to specific antigen in a graded
manner depending on the antigen concentration, which may have implications for a coordinate regulation of specific CD4ⴙT-cell responses.
Virus-specific CD4⫹T cells are thought to play a major role
in successful viral clearance in acute hepatitis B and hepatitis
C (1, 3, 7, 31). In memory CD4⫹ T-cell clones a relatively
constant threshold number of T-cell receptors (TCRs) has to
be triggered to induce CD4⫹T-cell activation as determined
by cytokine secretion (43). It is unclear, however, whether this threshold is regularly reached in vivo during different stages of viral infections. Previous studies of the T-cell response during viral hepatitis B and C have used saturating doses of antigen in vitro to detect antigen-specific proliferation or cytokine secre-tion. In contrast, during the natural course of viral hepatitis, the antigenic load varies by several orders of magnitude within a given compartment (e.g., peripheral blood) and even more between different compartments (e.g., blood and liver). There
is evidence from a study on autoreactive human CD4⫹T cells
that, similar to what has been described for altered peptide ligands, low concentrations of specific peptide can induce par-tial T-cell activation (19). It is unknown whether this is also
true for virus-specific CD4⫹ T cells that have been isolated
from a real disease situation and to what extent the antigen concentration influences the induction of different effector functions. For example, tiny amounts of residual viral antigens
may be able to promote long-term CD4⫹T-cell memory
fol-lowing resolution of acute hepatitis B and acute hepatitis C (13, 30, 38). In these patients there is no evidence of tissue
damage, suggesting that this low level stimulation of CD4⫹T
cells does not induce inflammatory responses. A detailed un-derstanding of different levels of T-cell activation may contrib-ute to our understanding of the coordinate regulation of the immune response in spontaneous viral clearance as well as to the development of T-cell vaccines for the treatment of chronic viral hepatitis.
During the last several years, considerable progress has been made in the understanding of the molecular basis of T-cell
activation. (i) CD4⫹T cells recognize antigens in the form of
8- to 12-amino-acid peptides bound to autologous HLA class II molecules (2). Despite a rather low affinity of the TCR for the
peptide-HLA complex, specific CD4⫹T cells can respond to
antigen-presenting cells displaying as few as 50 to 100 specific peptide-HLA complexes (39). However, full T-cell activation has been estimated to require the triggering of approximately 8,000 TCRs (43). This seeming paradox could be explained by the observation that T cells interact with antigen-presenting cells for a prolonged time and that a single specific peptide-HLA complex may serially trigger up to a few hundred TCRs (39). (ii) Evidence has accumulated that the TCR is not just an on-off switch but may be able to transmit qualitatively distinct signals into T cells (15, 26, 27, 34). Minor modifications within the amino acid sequence of a specific peptide can lead to inactive peptides, weaker or stronger agonists and antagonists (32), or so-called altered peptide ligands (APL) (11). While
* Corresponding author. Mailing address: Medizinische Klinik II, Klinikum Grosshadern, Marchioninistr. 15, 81377 Munich, Germany. Phone: 49-89-70 95 22 22. Fax: 49-89-70 00 95 40. E-mail: helmut [email protected].
7803
on November 9, 2019 by guest
http://jvi.asm.org/
APL selectively induce certain but not all effector functions or even T-cell anergy (36, 37), antagonistic peptides do not acti-vate specific T cells but inhibit stimulation by the wild-type peptide (32). Although different patterns of phosphorylation of TCR subunits have been described after stimulation with APL, the molecular basis for this distinct TCR signaling is still in-completely understood (16, 21, 26, 27, 32). (iii) A high affinity between peptide and major histocompatibility complex (MHC) or a high antigen dose may promote the differentiation of naive
CD4⫹T cells into Th1 cells, whereas a low affinity between
MHC and peptide or a low antigen concentration favors the development of a Th2 cytokine profile (4).
In this study, we investigated the response of virus-specific
memory CD4⫹ T cells to various concentrations of specific
peptides and correlated the induction of effector functions with the level of TCR triggering. To this end, we used hepatitis B virus (HBV) core (HBc) and hepatitis C virus (HCV)
NS3-specific CD4⫹T-cell clones which had been isolated during the
early phase of acute hepatitis from patients who subsequently cleared the infection. Our data demonstrate that virus-specific
CD4⫹T-cell clones can respond to low antigen concentrations
with up regulation of activation markers and adhesion mole-cules. A 30- to 100-fold-higher antigen concentration, which corresponds to the triggering of 20 to 30% of TCRs, is required to induce cytokine secretion or proliferation. The relevance of these findings for the coordinate regulation of the virus-spe-cific T-cell response in acute and chronic viral infections is discussed.
MATERIALS AND METHODS
Peptides. HBc-derived peptides (HBV amino acids [aa] 81–105 [SRDLV VSYVNTNMGLKFRQLLWFHI], HBV aa 145–158 [ETTVVRRRGRSPRR], and HBV aa 146–159 [TTVVRRRGRSPRRR]) and an HCV NS3-derived pep-tide (HCV aa 1248–1261 [GYKVLVLNPSVAAT]) were synthesized by Multiple Peptide Systems, San Diego, Calif., or Chiron Mimotopes, respectively; all pep-tides were purified by high-pressure liquid chromatography to⬎95%.
Specific CD4ⴙT-cell clones.HBc-specific CD4⫹T-cell clones (summarized in
Table 1) were isolated from an individual with acute hepatitis B 2 weeks after the onset of hepatitis, just after the peak of aminotransferase levels and disappear-ance of serum HBV DNA. The minimal epitopes for these T-cell clones were defined previously as aa 93 to 103 (MGLKFRQLLWF; T-cell clones G9, G40, and G42) or aa 145 to 155 (ETTVVRRRGRS; T-cell clones G27, and G61) (8). All HBc-specific clones were HLA-DRB1ⴱ1401 or HLA-DRB1ⴱ1302 restricted. HCV NS3-specific T-cell clone L11 (HLA-DRB1ⴱ1101 restricted) was isolated from an individual with acute hepatitis C shortly after viral elimination, 6 weeks
after the onset of acute hepatitis. The minimal epitope was previously deter-mined as aa 1251 to 1259 (VLVLNPSVA) (6).
CD4⫹T-cell clones were cultured in 96-well U-bottom plates (Costar,
Cam-bridge, Mass.) at a density of 1⫻104to 5⫻104/well in RPMI 1640 medium
(Gibco, Grand Island, N.Y.) containing 2 mML-glutamine, 1 mM sodium pyru-vate, 100 U of penicillin per ml, 100g of streptomycin per ml, 10% human AB serum, and 20 U of recombinant interleukin-2 (IL-2) (kindly provided by Boehr-inger, Mannheim, Germany) per ml. The CD4⫹T-cell clones were stimulated
every 3 to 5 weeks with irradiated allogeneic peripheral blood mononuclear cells (PBMC) (5⫻104/well) and 2g of phytohemagglutinin (HA16 [Murex
Diag-nostics, Dartford, United Kingdom]) per ml and were routinely used for exper-iments 3 to 4 weeks after the last restimulation, when expression of CD25 was back to baseline.
Proliferation assays.Specific CD4⫹T-cell clones were stimulated with
differ-ent concdiffer-entrations of specific peptide (0.0001 to 100g/ml) in the presence of irradiated autologous PBMC or the following HLA-matched lymphoblastoid cell lines (kindly provided by D. J. Schendel, Geselschaft fu¨r Strahlenforschung, Munich, Germany) (45): HO301 (DRA1ⴱ0102, DRB1ⴱ1302, DRB3ⴱ0301, DQA1ⴱ0102, DQB1ⴱ0605, DPA1ⴱ0201, DPB1ⴱ0501) for T-cell clones G9, G40, and G42; TEM (DRAⴱ0101, DRB1ⴱ1401, DQA1ⴱ0101, DQB1ⴱ05031, DPA1ⴱ01, DPB1ⴱ0401) for T-cell clones G27 and G61; and SPO010 (DRA1ⴱ0101, DRB1ⴱ1101, DRB3ⴱ0202, DQA1ⴱ0102, DQB1ⴱ0502, DPA1ⴱ01, DPB1ⴱ02012) for T-cell clone L11. After 4 days, T-cell cultures were labeled by incubation for 16 h with 2Ci of [3H]thymidine (specific activity, 80 mCi/mmol
[Amersham, Little Chalfont, United Kingdom]). The cells were collected and washed on filters (Dunn, Asbach, Germany) using a cell harvester (Skatron, Sterling, Va.), and the amount of radiolabel incorporated into DNA was esti-mated with a beta counter (LKB/Pharmacia, Uppsala, Sweden). Triplicate cul-tures were assayed routinely, and the results are expressed as mean counts per minute (cpm).
FACS analyses.Triple immunofluorescence staining was performed on T-cell clones 16 to 24 h following antigenic stimulation with the following combinations of antibodies: CD25-fluorescein isothiocyanate (Coulter, Hialeah, Fla.), HLA-DR–phycoerythrin (PE), CD54-PE, CD69-PE (Becton-Dickinson, Hamburg, Germany), and CD4-Tricolor (Medac, Hamburg, Germany). FACS analysis was performed with a FACScan instrument (Becton-Dickinson) as described previ-ously (17). Expression of surface molecules was determined as median fluores-cence intensity. A typical histogram for the expression of CD25 in response to different antigen concentrations is shown in Fig. 1.
Lymphokine assays.Specific T-cell clones were stimulated (105cells/100l)
with specific peptide in the presence of HLA-matched lymphoblastoid cell lines in a 1:1 ratio. Supernatants were collected after 24 h and stored at ⫺80°C. Secretion of IL-4 and gamma interferon was measured by sandwich enzyme-linked immunosorbent assay techniques as described previously (41, 42). Cyto-kine secretion by lymphoblastoid cell lines alone was always below the level of sensitivity.
Anergy induction.T-cell clone G27 was used for anergy induction 3 to 5 weeks after the last stimulation with phytohemagglutinin. A total of 104clone cells/well
were stimulated with specific peptide (HBc aa 145 to 155 or aa 146 to 155) at various concentrations (0.1, 1.0, 10, or 100g/ml) in the presence of HLA-matched lymphoblastoid cell lines (3⫻104/well). After 24 h the cells were
[image:2.612.50.553.84.194.2]washed twice in phosphate-buffered saline and restimulated with HBc aa 145 to TABLE 1. Summary of HBV- and HCV-specific CD4⫹T-cell clones
Clinical dataa T-cell clone Time point of isolation
(wk after onset) Specificity Cytokine profileb,c
Acute, self-limited hepatitis B; male;
58 yr old; follow-up, 36 mo G9 2 MGLKFRQLLWF
b(HBc aa 93–103) Th0
G40 3 MGLKFRQLLWF (HBc aa 93–103) Th1
G42 3 MGLKFRQLLWF (HBc aa 93–103) Th1
G27 2 ETTVVRRRGRSb(HBc aa 145–155) Th1
G61 3 ETTVVRRRGRS (HBc aa 145–155) Th0
Acute, self-limited hepatitis C; female;
64 yr old; follow-up, 16 mo L11 4 VLVLNPSVA
c(HCV NS3 aa 1251–1259) Th1
aSummary of clinical data in references 6 and 8.
bEpitope mapping and cytokine determinations are given in reference 9. The deduced amino acid sequence of the specific viral strain was previously determined and found to be identical to that of the peptides used in this study (8).
cEpitope mapping and cytokine determinations are given in reference 6. The deduced amino acid sequence of the infecting viral strain was identical to that of the peptide used in this study (unpublished results).
on November 9, 2019 by guest
http://jvi.asm.org/
155 (10g/ml). Proliferation was determined after 4 days by measuring [3
H]thy-midine incorporation.
RESULTS
Low antigen concentrations induce CD25 and adhesion mol-ecule expression in the absence of cytokine secretion or pro-liferation in virus-specific CD4ⴙT-cell clones.During the
nat-ural course of acute HBV or HCV infection, virus-specific
CD4⫹T cells are exposed to highly variable concentrations of
viral antigen. To investigate how virus-specific CD4⫹ T-cell
clones respond to different concentrations of specific antigen in
vitro, we stimulated HBc- and HCV NS3-specific CD4⫹T-cell
clones, which had been isolated during viral clearance from patients with acute hepatitis, with specific peptide at
concen-trations varying from 100 pg/ml to 100g/ml in the presence of
homozygous HLA-matched lymphoblastoid cell lines. CD3
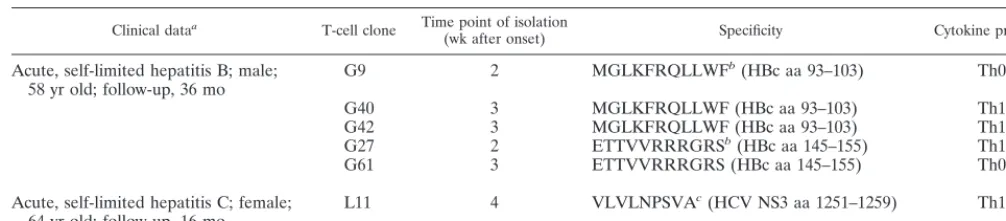
and CD4 down regulation were used as substitute markers for TCR internalization (29, 44). The first detectable markers of T-cell activation were CD25 and CD54 (intercellular cell ad-hesion molecule 1 [CAM-1]) up regulation, increasing the cell size and cluster formation (Fig. 2); similarly, HLA-DR and CD69 expression increased (data not shown). An increase in the levels of these markers could be observed as soon as a decrease in the levels of TCR components CD3 or CD4 oc-curred (Fig. 2A and B). Typically, cytokine secretion and pro-liferation required 10- to 100-fold-higher antigen concentra-tions than did induction of CD25 (Fig. 2A to C) and CD54 expression (Fig. 2D). Whereas the expression of CD25 and adhesion molecules increased as soon as TCR internalization became detectable, a decrease in CD3 expression of 20 to 30% was required for T-cell proliferation and cytokine secretion.
Expression of CD25 and adhesion molecules is functional.
To test whether the expression of CD25 was functional, we
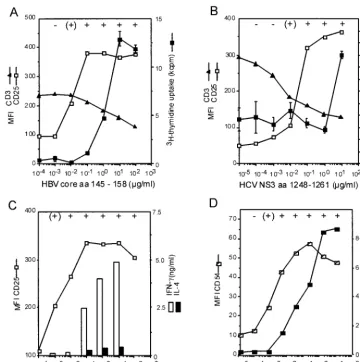
stimulated the CD4⫹T-cell clones G27, G40, and G61 with a
suboptimal concentration of specific peptide and increasing concentrations of recombinant IL-2. Dose-dependent prolifer-ation was induced in the peptide-stimulated cells but not in unstimulated cells (Fig. 3).
Intercellular adhesion was judged microscopically as forma-tion of cell clusters and correlated with the expression of ac-tivation markers and CD54 (ICAM-1). For all clones, cluster formation became detectable with similar kinetics to the ex-pression of CD54, demonstrating the functional exex-pression of adhesion molecules at low antigen concentrations (Fig. 2).
Induction of anergy in ThI clones requires full T-cell acti-vation.T-cell anergy has been discussed as a mechanism in-volved in self tolerance but also in the pathogenesis of chronic
infections. Stimulation of virus-specific CD4⫹T cells with low
antigen concentrations induced CD25 expression but no cyto-kine secretion, while the response to exogenous IL-2 was main-tained. These characteristics have frequently been referred to as the anergic phenotype (5, 33). Also, other researchers have claimed that suboptimal stimulation may induce anergy in
CD4⫹T cells (14). We therefore determined the threshold for
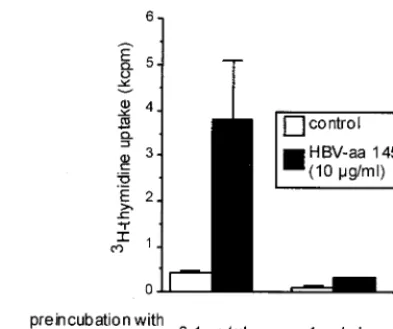
anergy induction in an HBV-specific CD4⫹T-cell clone (G27),
which had previously been shown to be prone to anergy induc-tion following stimulainduc-tion with high concentrainduc-tions of specific peptide (8). We found that for G27 the threshold for anergy induction corresponded to the threshold for induction of pro-liferation (Fig. 4). Stimulation with antigen concentrations that did not induce proliferation left the T-cell clone fully respon-sive to a higher antigen concentration.
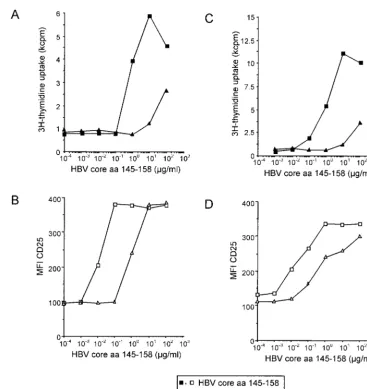
T-cell activation does not depend on antigen affinity but on the kinetics of TCR internalization.For T-cell clones G27 and G61, which are specific for the minimal epitope HBV aa 145 to 155, a low-affinity ligand which lacks the amino-terminal glu-tamic acid at position 145 (aa 146 to 159) was discovered, and its effect was compared to that of stimulation with a peptide containing the complete minimal epitope (aa 145 to 158). The low-affinity peptide aa 146 to 159 required 100-fold antigen concentration for the induction of CD25 expression (Fig. 5A and C) and proliferation (Fig. 5B and D) compared to the complete epitope, aa 145 to 158. Notably, independent of the antigen used for stimulation of clone G27, for any given level of TCR internalization, a similar T-cell response was observed (Fig. 6A) and a similar correlation between TCR
internaliza-FIG. 1. Representative histogram showing CD25 expression of a CD4⫹T-cell clone at different antigen concentrations. HBc-specific
CD4⫹ T-cell clone G40 has been stimulated with different antigen
concentrations inducing no (A), half-maximal (B), and maximal (C) expression of CD25. Cells were gated for CD4 expression to exclude the lymphoblastoid cells that were used as antigen-presenting cells. The dotted line represents unstimulated control cells.
on November 9, 2019 by guest
http://jvi.asm.org/
tion and the induction of different T-cell functions was found for other clones, as shown, for example, for the HCV NS3-specific T-cell clone L11 (Fig. 6B).
DISCUSSION
CD4⫹ T cells play a major role in host defense against
viruses and other intracellular pathogens. The importance of a
vigorous CD4⫹ T-cell response for a favorable outcome of
such infections has been shown in many animal models and can be inferred in humans from the occurrence of opportunistic infections in patients with T-helper- cell deficiency states (24, 25, 28). On the other hand, an uncontrolled or overshooting T-cell response may lead to unnecessary and detrimental tissue damage, as seen, for example, in fulminant viral hepatitis or in
postinfectious autoimmune disease. It is therefore obvious that a coordinate regulation of the immune response is required to achieve control of the infection on the one hand and to avoid excessive tissue damage on the other.
Here we show that virus-specific CD4⫹T cells can respond
to low antigen concentrations by up regulation of activation marker molecules and adhesion molecules without secretion of cytokines or proliferation, which require 30- to 100-fold-higher antigen concentrations. What is the relevance of this finding
for the virus-specific CD4⫹T-cell response? For CD8⫹
[image:4.612.122.483.71.433.2]cyto-toxic T cells (CTL) it has been shown that only tiny amounts of antigen are required to induce the lysis of target cells whereas at least 100-fold-higher levels are required for cytokine secre-tion (40). For CTL, it appears biologically reasonable that
FIG. 2. Induction of activation markers and effector functions in virus-specific CD4⫹ T-cell clones in relation to TCR internalization.
Expression of CD25 and CD54 and T-cell clustering occurred at about 100-fold-lower antigen concentrations than did proliferation or cytokine secretion. Whereas for proliferation a certain threshold of TCR internalization had to be reached, expression of CD25 occurred as soon as TCR internalization became detectable. (A and B) This is shown for the relationship between CD3, CD25, and proliferation with the HBc-specific CD4⫹
T-cell clone G27 (A) and the HCV-NS3-specific CD4⫹T-cell clone L11 (B). (C and D) The secretion of gamma interferon (IFN-␥) and IL-4 in
relation to CD25 expression is shown for HBc-specific CD4⫹T-cell clone G61 (C), and the relation between CD54 and proliferation is shown with
the HBc-specific CD4⫹T-cell clone G9 (D). All experiments were at least repeated twice, and similar results were obtained with autologous PBMC
and HLA-matched lymphoblastoid cell lines as antigen-presenting cells and were also valid for additional T-cell clones (G40 and G42). For all T-cell clones, the formation of cell clusters was evaluated microscopically: cluster formation was judged as absent if T cells were evenly distributed throughout the well, weak if some clusters could be identified whereas other cells were still outside the clusters, and strong if all the cells in the well were concentrated in clusters (see below). Cluster formation correlated strictly with the expression of CD25 and CD54. Symbols:⫺, absent; (⫹), weak;⫹, strong.
on November 9, 2019 by guest
http://jvi.asm.org/
specific CTL are extremely sensitive to low antigen concentra-tions, e.g., for killing of virally infected cells, but that their numbers expand only in the presence of higher levels of
anti-gen. For CD4⫹T cells, several immunological situations can be
conceived in which a differential regulation of T-cell activation might be required. (i) The interaction between TCR and MHC-peptide complex is of low affinity and is not sufficient to maintain intercellular contact between an antigen-presenting cell and a specific T cell. The early induction of adhesion molecules by triggering of a few TCRs could contribute to stabilization of the cellular interaction and facilitate further triggering of TCRs to achieve full T-cell activation. (ii)
Con-sidering a circulating specific CD4⫹T cell, it is conceivable that
at some distance from the inflammatory site the CD4⫹T cell
is partially activated by low antigen concentrations. This partial activation may promote T-cell adhesion and IL-2 responsive-ness without resulting in too early a secretion of inflammatory cytokines at a site still distant from the center of the inflam-matory process. Following migration to the focus of disease, a higher antigen concentration may then trigger full T-cell
acti-vation and secretion of cytokines. Thus, a graded CD4⫹T-cell
response may contribute to the translocation of specific CD4⫹
T cells to the center of an inflammatory process and avoid tissue damage outside the disease focus. (iii) After various viral infections, a memory T-cell response is maintained for many years and is probably related to the presence of low levels of residual viral antigen. Partial T-cell activation may promote T-cell survival without the secretion of inflammatory cytokines, which might otherwise induce ongoing tissue damage.
What could be the implications of these observations for the pathogenesis of chronic hepatitis B and C? Most studies of the
virus-specific CD4⫹T-cell response so far have used high
an-tigen concentrations (1 to 10g of protein or peptide antigens
[image:5.612.333.530.71.235.2]per ml) and have determined either proliferation or cytokine secretion (either in the supernatant or by the enzyme-linked immunoslot assay). By this approach is has clearly been shown for both HBV and HCV that viral clearance is associated with
FIG. 3. Response to exogenous IL-2 in three CD4⫹T-cell clones
stimulated with a suboptimal antigen concentration. (A and B) T-cell clones G27 (A) and G61 (B) were stimulated with 0.1g of specific peptide (aa 145 to 158) per ml, which induces high expression of the IL-2 receptor CD25 but no proliferation. (C) Similarly, T cell clone G40 was stimulated with 1.0g of peptide aa 81 to 105 per ml. Exogenous IL-2 induces proliferation (solid squares), whereas no pro-liferation occurred in the absence of peptide stimulation (open squares).
FIG. 4. Anergy induction in HBV aa 145 to 155-specific CD4⫹
T-cell clone G27. Prestimulation with a proliferation-inducing peptide concentration (1g/ml) leads to unresponsiveness to subsequent re-stimulation with high antigen concentrations (10g/ml). In contrast, prestimulation with 0.1g/ml, which induces maximum expression of CD25 but no proliferation, leaves the T-cell clone fully responsive to subsequent stimulation with higher peptide concentrations (10g/ml).
on November 9, 2019 by guest
http://jvi.asm.org/
[image:5.612.58.285.79.627.2]a strong virus-specific CD4⫹T-cell response, whereas in
pa-tients with chronic hepatitis B or C a virus-specific CD4⫹T-cell
response is either weak or absent. However, for chronic
hep-atitis C there are preliminary data suggesting that if CD4⫹T
cells are analyzed for antigen-induced CD25 expression,
virus-specific CD4⫹T cells can be detected in the absence of
anti-gen-specific proliferation or cytokine secretion (H. M. Diepolder, N. H. Gruener, J. T. Gerlach, M.-C. Jung, R. M. Hoffmann, R. Zachoval, and G. R. Pape, Abstr. 6th Int. Symp. Hepatitis C Relat. Viruses, p. 65, 1999). A similar observation
has been made with HCV-specific CD8⫹T cells using HLA
class I tetramers: in the peripheral blood of patients,
HCV-specific CD8⫹ T cells were identified that did not produce
cytokines and had a low potential for cytotoxicity (18, 23). Future studies are needed to clarify whether these
virus-spe-cific CD4⫹T cells in chronically infected patients are of lower
affinity or may have down regulated the TCR density, which
could explain why full T-cell activation including cytokine se-cretion cannot be induced.
For both HBV and HCV, a high rate of viral mutations has been described and viral escape from immune responses is considered to be a major factor in the pathogenesis of chronic viral hepatitis. TCR triggering depends on the trimolecular interaction between the HLA molecule, the peptide, and the TCR and is therefore influenced by both the binding affinity of the peptide and HLA molecule and the binding affinity be-tween the peptide-HLA complex and the TCR. It has previ-ously been shown that single amino acid changes in a specific peptide can lead to APL. APL induce only incomplete T-cell activation, where certain effector functions are triggered but others are not (10, 12, 35). For two HBc-specific T-cell clones (G27 and G61 [Fig. 5]), we identified a low-affinity peptide ligand which lacks the amino-terminal amino acid of the
min-imal epitope. At standard concentrations (10g/ml), this
pep-FIG. 5. Induction of activation markers and effector functions with high- and low-affinity ligands. CD4⫹T-cell clones G27 (A and B) and G61
(C and D) have been stimulated with various concentrations of peptide aa 145 to 158, which contains the minimal specific epitope (squares), and the low-affinity variant aa 146 to 159 (triangles). Induction of proliferation (A and C) and CD25-expression (B and D) required a 100-fold higher concentration of the low-affinity peptide aa 146 to 159.
on November 9, 2019 by guest
http://jvi.asm.org/
[image:6.612.119.486.68.457.2]tide induced the expression of CD25 and intercellular adhesion but induced no proliferation. Interestingly, when the degree of TCR internalization was studied over a broad range of antigen concentrations, for any level of TCR internalization the variant peptide induced exactly the same response as did the wild-type peptide with regard to phenotype and proliferation. These data support the hypothesis, as previously suggested (19, 20, 22), that if TCR triggering is determined by TCR internalization, it is the quantity rather than the quality of TCR triggering that determines the T-cell response. Moreover, these findings imply that if the consequences of viral mutations on specific T-cell responses are studied, a broad range of antigen concentrations must be used to define the relevance of an individual mutation. A previous study that looked at the hierarchy of T-cell re-sponses to increasing doses of specific peptide found that two
CD4⫹T-cell clones specific for the same peptide of the
au-toantigen myelin basic protein differed significantly with regard to the sequence of T-cell effector functions that were induced (19). No such differences in effector function hierarchy were
found in our six different CD4⫹T-cell clones from two patients
with different viral infections, suggesting that the hierarchy of
T-cell functions may be more constant in virus-specific CD4⫹
T cells.
It should be noted that these observations have been made
in CD4⫹ T-cell clones which have been expanded in vitro.
Obviously the cloning procedure itself leads to a selection of T cells, and it is therefore never completely clear whether the
clones are representative of the circulating specific CD4⫹ T
cells in vivo. Also, some effector functions may change during in vitro culture. Nevertheless, our findings were very consistent in a set of different T-cell clones from different patients. Also, the durations of in vitro culture of individual clones were quite different, ranging from several weeks to years, and no signifi-cant differences were noted between these clones. Moreover,
even if the behavior of the CD4⫹ T-cell clones that we
ob-served cannot be generalized to all primed CD4⫹T cells in
vivo, it is likely to be part of a preexisting program that be-comes active under certain circumstances. This important question can be addressed as soon as disease-specific HLA class II tetramers are available, which will allow the labeling of
specific CD4⫹T cells in freshly isolated PBMC without prior
stimulation with antigen.
In conclusion, our results suggest that CD4⫹ T cells from
patients with self-limited acute hepatitis B or hepatitis C can respond in a graded manner to different antigen
concentra-tions. If this observation can be generalized to specific CD4⫹T
cells in vivo, it may have implications for the regulation and
trafficking of specific CD4⫹T cells in general. Our results also
suggest that a better knowledge not only of viral sequence variations but also of viral antigen concentrations in vivo may be required for a more complete understanding of the immu-nopathogenesis of viral infections.
ACKNOWLEDGMENTS
This work was supported by the Wilhelm-Sander-Stiftung (grant 94.072.3) and the European Commission (contract QLRT-PL1999-00356) within the 5th Framework Program.
We thank Antonio Lanzavecchia, Istituto di Ricerca in Biomedicina, Bellinzona, Switzerland, and Annegret de Baey, Micromet GmbH, Munich, Germany, for critical discussion; Alessandro Sette and Scott Southwood, Epimmune Inc., San Diego, Calif., for determinations of HLA-peptide-binding affinities; and Jutta Do¨hrmann, Carmen Amsel, and Carola Steiger for excellent technical assistance.
REFERENCES
1.Cerny, A., and F. V. Chisari.1999. Pathogenesis of chronic hepatitis C: immunological features of hepatic injury and viral persistence. Hepatology
30:595–601.
2.Chien, Y. H., and M. M. Davis.1993. How alpha beta T-cell receptors ‘see’ peptide/MHC complexes. Immunol. Today14:597–602.
3.Chisari, F. V., and C. Ferrari.1995. Hepatitis B virus immunopathogenesis. Annu. Rev. Immunol.13:29–60.
FIG. 6. Correlation of the degree of TCR internalization with the expression of CD25 (open symbols) and proliferation (solid symbols). (A) For HBV aa 145 to 158-specific CD4⫹T-cell clone G27, the correlation of TCR internalization with the induction of T-cell effector functions was
studied using the wild-type epitope (aa 145 to 158) and a low-affinity truncated variant (aa 146 to 159). Independent from the epitope used, for any given level of TCR down regulation, a similar degree of CD25 expression and proliferation, respectively, was observed. For both peptides, the individual measurements could be approximated by the same sigmoid-shaped curve. (B) A very similar correlation between TCR internalization and the induction of proliferation and CD25 expression is shown for the HCV NS3-specific CD4⫹T-cell clone L11.
on November 9, 2019 by guest
http://jvi.asm.org/
4.Constant, S., C. Pfeiffer, A. Woodard, T. Pasqualini, and K. Bottomly.1995. Extent of T cell receptor ligation can determine the functional differentiation of naive CD4⫹T cells. J. Exp. Med.182:1591–1596.
5.DeSilva, D. R., K. B. Urdahl, and M. K. Jenkins.1991. Clonal anergy is induced in vitro by T cell receptor occupancy in the absence of proliferation. J. Immunol.147:3261–3267.
6.Diepolder, H. M., J. T. Gerlach, R. Zachoval, R. M. Hoffmann, M. C. Jung, E. A. Wierenga, S. Scholz, T. Santantonio, M. Houghton, S. Southwood, A. Sette, and G. R. Pape.1997. Immunodominant CD4⫹T-cell epitope within
nonstructural protein 3 in acute hepatitis C virus infection. J. Virol.71:6011– 6019.
7.Diepolder, H. M., R. M. Hoffmann, J. T. Gerlach, R. Zachoval, M. C. Jung, and G. R. Pape.1998. Immunopathogenesis of HCV infection. Curr. Stud. Hematol. Blood Transfus.62:135–151.
8.Diepolder, H. M., M. C. Jung, E. Wierenga, R. M. Hoffmann, R. Zachoval, T. J. Gerlach, S. Scholz, G. Heavner, G. Riethmuller, and G. R. Pape.1996. Anergic Th1 clones specific for hepatitis B virus (HBV) core peptides are inhibitory to other HBV core-specific CD4⫹T cells in vitro. J. Virol.70:
7540–7548.
9.Diepolder, H. M., G. Ries, M. C. Jung, H. J. Schlicht, J. T. Gerlach, N. H. Gruener, W. H. Caselmann, and G. R. Pape.1999. Differential antigen-processing pathways of the hepatitis B virus e and core proteins. Gastroen-terology116:650–657.
10. Evavold, B. D., and P. M. Allen.1991. Separation of IL-4 production from Th cell proliferation by an altered T cell receptor ligand. Science252:1308–1310. 11. Evavold, B. D., J. Sloan-Lancaster, and P. M. Allen.1993. Tickling the TCR: selective T-cell functions stimulated by altered peptide ligands. Immunol. Today14:602–609.
12. Evavold, B. D., J. Sloan-Lancaster, B. L. Hsu, and P. M. Allen.1993. Separation of T helper 1 clone cytolysis from proliferation and lymphokine production using analog peptides. J. Immunol.150:3131–3140.
13. Gerlach, J. T., H. M. Diepolder, M. C. Jung, N. H. Gruener, W. W. Schraut, R. Zachoval, R. Hoffmann, C. A. Schirren, T. Santantonio, and G. R. Pape.
1999. Recurrence of hepatitis C virus after loss of virus-specific CD4(⫹) T-cell response in acute hepatitis C. Gastroenterology117:933–941. 14. Girgis, L., M. M. Davis, and B. F. de St Groth.1999. The avidity spectrum
of T cell receptor interactions accounts for T cell anergy in a double trans-genic model. J. Exp. Med.189:265–278.
15. Grakoui, A., S. K. Bromley, C. Sumen, M. M. Davis, A. S. Shaw, P. M. Allen, and M. L. Dustin.1999. The immunological synapse: a molecular machine controlling T cell activation. Science285:221–227.
16. Grakoui, A., L. F. VanDyk, S. F. Dowdy, and P. M. Allen.1998. Molecular basis for the lack of T cell proliferation induced by an altered peptide ligand. Int. Immunol.10:969–979.
17. Gruber, R., C. Reiter, and G. Riethmuller.1993. Triple immunofluorescence flow cytometry, using whole blood, of CD4⫹and CD8⫹lymphocytes ex-pressing CD45RO and CD45RA. J. Immunol. Methods163:173–179. 18. Gruener, N. H., F. Lechner, M.-C. Jung, H. M. Diepolder, T. Gerlach, G.
Lauer, B. Walker, J. Sullivan, R. Phillips, G. R. Pape, and P. Klenerman.
2001. Sustained dysfunction of antiviral CD8⫹T lymphocytes after infection
with hepatitis C virus. J. Virol.75:5550–5558.
19. Hemmer, B., I. Stefanova, M. Vergelli, R. N. Germain, and R. Martin.1998. Relationships among TCR ligand potency, thresholds for effector function elicitation, and the quality of early signaling events in human T cells. J. Im-munol.160:5807–5814.
20. Itoh, Y., B. Hemmer, R. Martin, and R. N. Germain.1999. Serial TCR engagement and down-modulation by peptide:MHC molecule ligands: rela-tionship to the quality of individual TCR signaling events. J. Immunol.
162:2073–2080.
21. Kersh, E. N., G. J. Kersh, and P. M. Allen.1999. Partially phosphorylated T cell receptor zeta molecules can inhibit T cell activation. J. Exp. Med.
190:1627–1636.
22. Lanzavecchia, A., G. Lezzi, and A. Viola.1999. From TCR engagement to T cell activation: a kinetic view of T cell behavior. Cell96:1–4.
23. Lechner, F., D. K. Wong, P. R. Dunbar, R. Chapman, R. T. Chung, P. Dohrenwend, G. Robbins, R. Phillips, P. Klenerman, and B. D. Walker.
2000. Analysis of successful immune responses in persons infected with hepatitis C virus. J. Exp. Med.191:1499–1512.
24. Maloy, K. J., C. Burkhart, G. Freer, T. Rulicke, H. Pircher, D. H. Kono, A. N. Theofilopoulos, B. Ludewig, U. Hoffmann-Rohrer, R. M. Zinkernagel,
and H. Hengartner.1999. Qualitative and quantitative requirements for CD4⫹T cell-mediated antiviral protection. J. Immunol.162:2867–2874. 25. Maloy, K. J., C. Burkhart, T. M. Junt, B. Odermatt, A. Oxenius, L. Piali,
R. M. Zinkernagel, and H. Hengartner.2000. CD4(⫹) T cell subsets during virus infection. Protective capacity depends on effector cytokine secretion and on migratory capability. J. Exp. Med.191:2159–2170.
26. Matsushita, S., and Y. Nishimura.1997. Partial activation of human T cells by peptide analogs on live APC: induction of clonal anergy associated with protein tyrosine dephosphorylation. Hum. Immunol.53:73–80.
27. Nishimura, Y., Y. Z. Chen, T. Kanai, H. Yokomizo, T. Matsuoka, and S. Matsushita.1998. Modification of human T-cell responses by altered pep-tide ligands: a new approach to antigen-specific modification. Intern. Med.
37:804–817.
28. Oxenius, A., R. M. Zinkernagel, and H. Hengartner.1998. CD4⫹T-cell induction and effector functions: a comparison of immunity against soluble antigens and viral infections. Adv. Immunol.70:313–367.
29. Padovan, E., G. Casorati, P. Dellabona, S. Meyer, M. Brockhaus, and A. Lanzavecchia.1993. Expression of two T cell receptor alpha chains: dual receptor T cells. Science262:422–424.
30. Penna, A., M. Artini, A. Cavalli, M. Levrero, A. Bertoletti, M. Pilli, F. V. Chisari, B. Rehermann, G. Del Prete, F. Fiaccadori, and C. Ferrari.1996. Long-lasting memory T cell responses following self-limited acute hepatitis B. J. Clin. Investig.98:1185–1194.
31. Rehermann, B., and F. V. Chisari.2000. Cell mediated immune response to the hepatitis C virus. Curr. Top. Microbiol. Immunol.242:299–325. 32. Ruppert, J., J. Alexander, K. Snoke, M. Coggeshall, E. Herbert, D.
McKen-zie, H. M. Grey, and A. Sette.1993. Effect of T-cell receptor antagonism on interaction between T cells and antigen-presenting cells and on T-cell sig-naling events. Proc. Natl. Acad. Sci. USA90:2671–2675.
33. Schwartz, R. H.1996. Models of T cell anergy: is there a common molecular mechanism? J. Exp. Med.184:1–8.
34. Sette, A., J. Alexander, and H. M. Grey.1995. Interaction of antigenic peptides with MHC and TCR molecules. Clin. Immunol. Immunopathol.
76:S168–S171.
35. Sloan-Lancaster, J., B. D. Evavold, and P. M. Allen.1993. Induction of T-cell anergy by altered T-cell-receptor ligand on live antigen-presenting cells. Nature363:156–159.
36. Sloan-Lancaster, J., B. D. Evavold, and P. M. Allen.1994. Th2 cell clonal anergy as a consequence of partial activation. J. Exp. Med.180:1195–1205. 37. Sloan-Lancaster, J., A. S. Shaw, J. B. Rothbard, and P. M. Allen.1994. Partial T cell signaling: altered phospho-zeta and lack of zap70 recruitment in APL-induced T cell anergy. Cell79:913–922.
38. Takaki, A., M. Wiese, G. Maertens, E. Depla, U. Seifert, A. Liebetrau, J. L. Miller, M. P. Manns, and B. Rehermann.2000. Cellular immune responses persist and humoral responses decrease two decades after recovery from a single-source outbreak of hepatitis C. Nat. Med.6:578–582.
39. Valitutti, S., S. Muller, M. Cella, E. Padovan, and A. Lanzavecchia.1995. Serial triggering of many T-cell receptors by a few peptide-MHC complexes. Nature375:148–151.
40. Valitutti, S., S. Muller, M. Dessing, and A. Lanzavecchia.1996. Different responses are elicited in cytotoxic T lymphocytes by different levels of T cell receptor occupancy. J. Exp. Med.183:1917–1921.
41. Van der Meide, P. H., M. Dubbeld, and H. Schellekens.1985. Monoclonal antibodies to human immune interferon and their use in a sensitive solid-phase ELISA. J. Immunol. Methods79:293–305.
42. van der Pouw Kraan, T., I. Rensink, and L. Aarden.1993. Characterisation of monoclonal antibodies to human IL-4: application in an IL-4 ELISA and differential inhibition of IL-4 bioactivity on B cells and T cells. Eur. Cytokine Netw.4:343–349.
43. Viola, A., and A. Lanzavecchia.1996. T cell activation determined by T cell receptor number and tunable thresholds. Science273:104–106.
44. Viola, A., M. Salio, L. Tuosto, S. Linkert, O. Acuto, and A. Lanzavecchia.
1997. Quantitative contribution of CD4 and CD8 to T cell antigen receptor serial triggering. J. Exp. Med.186:1775–1779.
45. Yang, S. Y. E., E. Milford, U. Ha¨mmerling, and B. Dupont1989. Description of the refernece panel of B lymphblastoid cell lines for factors of the HLA system: the B cell panel designed for the Tenth International Histocompat-ibility Workshop, p. 11–19.InB. Dupont (ed.), Immunobiology of HLA. Springer Verlag, New York, N.Y.