Vol. 33, No. 3 JOURNAL OFVIROLOGY, Mar.1980,p.1083-1096
0022-538X/80/03-1083/14$02.00/0
Spontaneous
Regression
of Friend Virus-Induced
Erythroleukemia
VI.
Structural and Antigenic Differences Between the Regressing and
Conventional Strains of Virus
PHILIP FURMANSKI,* CLIFFORD LONGLEY, CHRISTOPHER S. BOLLES, DAVID L. HINES,AND
MICHAEL DIETZ
Departmentof Biology, Michigan Cancer Foundation, Detroit, Michigan 48201
The regressing and conventional strains of Friend virus were compared by
neutralizationassays, sodiumdodecylsulfate-polyacrylamidegelelectrophoresis, andtryptic peptide mapping of the individual viralcomponents. Neutralization rates of the two viruses differed in the presence of monospecific anti-gp70 antiserumandserafromregressedorimmunizedmice. Neutralization of regress-ing Friend virus, butnotconventional Friend virus,occurred when the viruses were incubated with anti-pl5(E) and complement. Human serum inactivated conventional Friend virusmorerapidly than regressing Friend virus, probablyas aresult ofvirolysis induced by the reaction of viral p15(E) with human comple-mentcomponent Cl. Structural differences between the viruses were detectedin theirgp7O viralglycoproteinsandp15(E) andp12proteins. Analysis of different stocksand clonalisolatesof theviruses showedthatthedifferences betweenthe gp70 and p15(E), butnotthe p12proteins, were associatedwith the regressing phenotype of theregressing strainof Friend virus.
In contrast to the progressive, chronic, and lethal erythroleukemia caused in miceby con-ventional strains of Friend virus (CFV), the re-gressing strain of Friend virus (RFV) inducesan
erythroleukemia that spontaneously regresses (28, 30). Both viruses induceaninitially identical disease characterizedby massive splenomegaly
and viremia. However, at 30 to 60 days post-virusinoculation, when the mice inoculated with CFVbegintodieduetotheirdisease,the
eryth-roleukemia in the mice inoculated with RFV
spontaneously regresses and, in about half the leukemic mice, the animals return to a
histo-pathologicallyandvirologicallynormalor near-normal state. This characteristic of the RFV strain of Friend virus isstable through routine
sequentialviruspassageandtoviruscloning.
Our previous studies have established that immunological reactivity isanessential compo-nent ofthe process ofregression and that re-gressed mice developpotentantivirus and anti-leukemia cell immunologicalactivity (5, 17, 29; C.S.Johnson, S.Fouchey,and P.Furmanski,J. Natl. CancerInst.,in press). Bothhumoraland cell-mediatedspecificimmunereactivitycan be detected in regressed mice. We have also sug-gestedthat thedifferencebetween the RFV and conventional strains of virus which results in disease regressionis that RFV or its structural components are morehighly immunogenicthan thoseof CFV(14).
These data have ledus tocomparethe struc-turalcomponents of RFV andCFV,both phys-ico-chemically and immunologically. We report here that in neutralization and neutralization kinetics assays RFV and CFV are distinguish-able on the basis ofreactivity with sera from regressed mice, from immunized mice, and with monospecific antisera againstgp70 andp15(E).
Furthermore,theviralgp70 and p15(E) compo-nentshave differenttryptic peptidemaps.
MATERIALS AND METHODS
Viruses. RFVwasmaintainedbyserial passage in
random-bredSwiss/ICRmice. Virusstockswere
pre-pared as20%cell-free spleen homogenatesfrom
leu-kemic mice and stored at -70°C. Unless otherwise
indicated,the CFV used in theseexperimentswasthe
N-tropic,nonregressingFriendvirus,PEN,originally
obtained from Robert J. Eckner,Boston University
School of Medicine, and maintained, as described
above,byserial passage inNIH/PLCRmice.
For use in polyacrylamide gel electrophoresis (PAGE) and tryptic peptide mapping experiments,
virus waspurifiedfromculturefluids of infectedNIH/
3T3 cells. Spleen homogenates were diluted 20-fold
withDulbecco modifiedEaglemedium(DMEM),
fil-tered, and inoculatedontocultures ofNIH/3T3cells
in 75-cm2 plastic tissue culture flasks.The cellshad
beenpretreated for1 hwith 25ug of DEAE-dextran
per ml and then washed with serum-free medium.
Afterinfection for 1 h at 37°C, 20ml of DMEM + 15%
calfserum (K-CBiologicals)was added to eachflask.
Three days after infection, thecellsof each flaskwere
1083
on November 10, 2019 by guest
http://jvi.asm.org/
trypsinized andtransferred to a 490-cm2 plasticroller
bottle(Corning) in 60 ml of DMEM + 15% calf serum.
Thefollowing day, the medium was replaced with 60
ml of DMEM + 5%heat-inactivated fetal calf serum
(K-C Biologicals), and on day 7 postinfection, the
bottles were fedagain with the same medium.
Super-natants for viruspurification was harvested on days
10, 12, and 14 postinfection. The fluids were
centri-fuged at10,000 x g for 10min and then layered over
apadof 20% sucrose in TNE (0.01 M Tris, 0.15 M
NaCl, 0.002 M EDTA, pH 7.4) in an SW 27 tube and
centrifuged at 25,000 rpm for 60min. The pellets were
suspended in TNE buffer, layered onto a 20 to 50%
sucrose gradient in TNE in an SW 50.1 tube and
centrifuged at48,000 rpm for 60min. Virus bands were
removed, diluted with TNE buffer, and rebanded in a
20 to 50% sucrose gradient. The virus bands were
diluted with TNE buffer andpelleted in an SW 50.1
rotor at 48,000 rpm for 60 min. The pellets were
drained, and the tubes were sealed and stored at -700C.
For thepreparation of radiolabeled virus, the
me-dium in theroller bottleswasreplacedwith 30 ml of
leucine-free DMEM containing 5% dialyzed,
heat-in-activatedfetal calfserum and either30uCi of
[3H]-leucine (Amersham Corp.) per ml or 4 MuCi of
["C]-leucine (Amersham) per ml. Supernatant fluids were
harvested and viruswaspurifiedasdescribed above.
Pseudotypes of the Kirsten sarcoma virus
(Ki-MuSV) werepreparedwith RFVand CFV forusein
theneutralization and neutralization kinetics assays.
Cultures ofnonproducerK-NIH cells(obtainedfrom
S. Aaronson, National Cancer Institute[1]) were
su-perinfected with about 500PFU of the appropriate
murineleukemia virus(MuLV). Supernatants of the
infected cultureswereharvestedat7 or 14days
post-inoculation and titrated forfocus-forming activityon
NIH/3T3 cells (see below).Pseudotypes ofMoloney
sarcomavirus(MoMuSV)werepreparedin thesame
way from cultures of a transformed nonproducer
MoMuSV-NIH/3T3 cell line isolated in our labora-tory.
Antisera.Monospecificgoat antiseratoRauscher
virus-derived gp7O, p30, p15, p12, and plOwere
ob-tainedthrough the Office ofProgramResources and
Logistics, Virus CancerProgram,National Cancer
In-stitute. Rabbit antiseratoRauscherp15(E) were
ob-tained from E. Fleissner, Memorial SloanKettering
Cancer Center, and S. Oroszlan, Frederick Cancer
Research Center.Regressedserawerepoolsfrom
in-dividualregressedmice. Immuneserum wasobtained
by inoculationof strain129/Jmice(Jackson
Labora-tories) with RFV; those mice that did not become
leukemicwere reinoculated with virus40dayslater
and then reinoculated again after another 40 days.
Fourteendaysafter the lastinoculation,theanimals
werebled, and theirserum wasseparatedand stored
at-70°C. Humanserum wasobtained from blood of
healthydonors,filtered,andstoredat-70°C.
PAGE. Virusproteinswereseparatedandanalyzed
by sodiumdodecylsulfate(SDS)-PAGEin15%
acryl-amide,15-cmgelsprepared bythe method ofLaemmli
(23). The gelswereelectrophoresed at1mApergel
for 16 h.Alternatively,thehighresolutiongelsystem
describedby Montelarowasusedexactlyasdescribed
earlier (24, 25), except for the equimolar substitution ofB-mercaptoethanolfor themercaptopropionic acid.
When the polypeptides were labeled with 125I (see
below), the gels were frozen, sliced on a Mickle gel
slicer (BrinkmannInstruments), and two 1-mm
seg-ments were counted in a Searle automatic gamma
counter.Gels of3H-or"4C-labeledvirus, or both, were
pulverized in2-mmsegments directly intoscintillation
vials with a Gilson automatic gel fractionator. The
radioactivity wasdetermined in a toluene-Triton
X-100scintillation cocktail with a Packardscintillation
spectrometer. Protein standards ("nI-labeled bovine
serum albumin, ovalbumin, chymotrypsinogen, and
RNase) were included with eachgel run for
determi-nationof molecular weights.
Bromelaindigestion. Intact, purified virus was
digested with purified bromelain as described earlier
(25, 26). Afterdigestionfor3hat37°C, the virus was
collectedbycentrifugationthroughadiscontinuous 20
to50%sucrosegradient andpelleted. Thepelletswere
dissolved in 1%SDS, 1%,B-mercaptoethanol, 0.01 M
phosphate buffer, pH 7.2, heated for 4 min at 100°C, andanalyzedby SDS-PAGE.
Trypticpeptide mapping.A
10-pl
amountofpu-rifiedvirus (1 mg ofprotein/mlin water) was added to
40
pi
of TNE+0.2% Nonidet P-40(NP-40) andincu-batedat4°Cfor30min.One mCi of'25I(Amersham)
wasthenadded,followedby20
pl
ofchloramine T (4mg/ml). After incubationatroomtemperature for10
min, the reactionwasterminated with50
pl
of sodiummetabisulfite (5 mg/ml). The mixturewasdilutedwith
100
pl
of TNE containing 2% NP-40, 1% sodiumde-oxycholate, and 10 mM KI. Phenylmethyl sulfonyl
fluoridedissolved inisopropylalcoholwasthenadded
to a finalconcentration of100,uM. The labeled virus
was dialyzed against TNE containing 0.2% NP-40,
0.05% sodiumdeoxycholate, and 5mMKI. After
di-alysis,phenylmethyl sulfonylfluoridewasagainadded
to 100MM.
Individual virusproteinswereimmunoprecipitated
from the labeled preparation and purified by
SDS-PAGE as follows: the virus suspension was diluted
with TNE containing 0.2% NP-40, 100MLM
phenyl-methyl sulfonyl fluoride, and 1 mg of bovineserum
albumin per ml. Normal goatserumwasadded(2
pi),
and the mixturewasincubatedfor1h at4°C, followed
by the addition of20
pl
ofa10%suspensionofStaph-ylococcusaureus, Cowan strainI,prepared exactlyas
describedpreviously (21).After incubationat4°Cfor
30min, the bacteriawereremovedbycentrifugation
(2,000xg,20min),and thesupernatantwasusedfor
immunoprecipitation. A
2-pl
amountof theappropri-atemonospecificantiserumwasaddedto200
pl
of thelabeled virus, and the mixture wasincubatedat4°C
for 1h. Staphylococcusorganismswereadded,
incu-bated, and removed asdescribed above. The pellet
containing the bacteria and antigen-antibody
com-plexeswasthensuspendedin0.5ml ofTNEcontaining
0.5% NP-40and 0.02% sodiumazide. Thesuspension
waslayeredover1mlofasolution of TNEcontaining
5%sucrose, 0.5% sodiumdeoxycholate,and 3%NP-40.
Aftercentrifugation (2,000xg,10min),thepelletwas
washed with TNE+ 0.5% NP-40and drained. The
complexes were dissociated with 0.01 M phosphate
buffer,pH7.4,2%SDS,and 2%
fB-mercaptoethanol
aton November 10, 2019 by guest
http://jvi.asm.org/
STRUCTURE OF REGRESSING FRIEND VIRUS
100°C for3 min, and the bacteriawere removedby
centrifugation (2,000 x g, 10 min). The supernatant
wasthenremoved and electrophoresedon
high-reso-lutionSDS-PAGEgels,preparedasdescribed above.
After electrophoresis, the gels were frozen and
sliced, and the slices were counted in the gamma
counter. The slices containing the specific, purified,
immunoprecipitatedproteinwerepooled and washed
twice for30mineachat roomtemperaturewith 50%
methanol, 10% acetic acid inwater. The sliceswere
thendried ina vacuum ovenandrehydrated with0.5
mlof0.05M NH4HCO3, pH 8.0, containing 25 Lg of
L-(tosylamido 2-phenyl)-ethyl chloromethyl
ketone-trypsin (269U/mg;WorthingtonBiochemicalsCorp.).
The slices were incubated overnight at 37°C with
continuous gentle shaking. The supernatant was
re-moved andlyophilized.
The tryptic peptides were analyzed by
2-dimen-sional thin-layer electrophoresis/chromatography
(10). Thepeptidesweredissolved in acetic acid-formic
acid-water, 15:5:80, applied to a 0.1-mm
cellulose-coated thin-layerchromatography plate (EM
Labo-ratories), andelectrophoresed at900V for about 35
minat0°C. Theplatewasthen dried and
chromato-graphed in the second dimension in
butanol-pyridine-aceticacid-water,32.5:25:5:20.Theplatewasdried and
exposed to Kodak X-Omat R film.A single Cronex
Lightning PlusX-rayintensifyingscreen wasusedto
enhance theautoradiography(35).
Insomeinstances,sequential immunoprecipitations
werecarriedout onsingle labeled virus preparations.
Differences intryptic peptide mapswereconfirmedby
analysis of admixtures of the tryptic peptide prepara-tions.
Neutralization assays. The KiMuSV
pseudo-typeswerediluted with serum-free DMEMtoafinal
titerof about400focus-forming units per ml. Portions
of0.5 mlwere added to tubes containing0.5 ml of
serialdoubling dilutions of thetestantiserum made in
Dulbeccophosphate-bufferedsaline(GIBCO
Labora-tories).The mixtureswereincubated ina37°Cwater
bath for 1 h and chilled, and 0.1-ml portions were
inoculated onto NIH/3T3 cells in the individualwells
ofamultiwell dish (16-mmwells, COSTAR). Thewells
wereseeded the previous day with10"cells in DMEM
containing 15% calfserumand2,ugofpolybrene
(Ald-richChemical) per ml and washedoncewith DMEM
before inoculation. After the addition of the
virus-antibody mixture, thedisheswereincubatedfor 1 h at
37°Cin 10% CO2in air, andthen 0.4 mlof DMEM
with 15% calf serum was added to each well. The
following day andonday4post-virus inoculation, the
cultures were fed with DMEM containing 5% calf
serum and 1%dimethyl sulfoxide (22). On day 7
post-infection, the cells were fixed and stained with Giemsa.
Fociwerecountedmicroscopically.
For neutralization assays with complement, freshly
reconstituted,lyophilizedguinea pig serum (Colorado
Serum Co.) was addedto the virussuspension to a
final concentration of 5% before admixture with the antiserum.
All neutralization assays were carried out at least in duplicate.
Neutralization kinetics assays. The KiMuSV
pseudotypes of thetestviruseswere incubated with a
single dilution of serum, andatthe timesindicated,
portionswereremoved,diluted,chilled,and tested for
residual virus. The concentration ofserum used in
these assayswasgenerallytwo toeight timeshigher
than the concentration which caused 50%
neutraliza-tion ofthe moresensitivepseudotypeintheendpoint
assaysdescribed above. Portions of0.5ml of theserum
diluted in phosphate-buffered saline were added to
tubes. Anequal volume ofvirus,diluted in DMEMto
contain about 4,000focus-forming units per ml was
thenadded and mixed inaVortexmixer,and0.1 ml
wasimmediatelyremoved withanautomaticpipettor
(zero timesample).Theremainingvirus-antibody
mix-turewasincubated, with occasionalgentle shaking,in
a 37°C water bath. At 10, 20, 40, and 60 min of
incubation, 0.1-ml samples were removed.
Immedi-ately afterremoval, each sample, includingthe zero
time sample,wasaddedto aprechilledtubecontaining
0.9mlof DMEM and mixed inaVortex mixer. The
dilutedsampleswereheldoniceuntil the serieswas
complete, and they werethen inoculated in
quadru-plicate ontomonolayers ofNIH/3T3 cells in 16-mm
multiwellplates for virus assayasdescribed above.
Virus clones. Cultures containing 105
DEAE-dex-tran-treated NIH/3T3 cells in 60-mm dishes were
infected with 100 XC PFU of virus (4). Thecells were
trypsinized and inoculated at 100cells perwell into
the individual wells ofa Microtest II plate (Falcon
Plastics).Whenthecellshad grown toconfluence, the
supernatantswereremoved and frozen, and thecells
in thewellswereUV irradiated and overlaid with XC
cells. Thesupernatants from XC-positive wells (-10
per96-wellplate)werethen usedtopreparestocks in
fresh NIH/3T3 cells (see above) and to prepare
pseu-dotypes in K-NIH cells,asdescribed above.
RESULTS
Immunological analysis of viral
compo-nents by neutralization. Differences in the antigenic properties of RFV and CFVwerefirst determinedby usingneutralizationassays.This approach, although limited to the analysis of external proteins or glycoproteins, relates di-rectly to the major humoral antiviral immune response detectable in regressed but not leu-kemic mice (17, 29; M. Dietz, D. Hines, M. A.
Rich,andP.Furmanski, Proc. Am. Assoc. Can-cerRes. 19:156, 1978) andmight, therefore, be relevantto theprocessofregression.
The effect of monospecific antisera against each of themajor viral polypeptideswastested inneutralizationassaysbothinthepresenceand absenceofcomplement. Aspreviouslyreported (13,32;M.Kende, S. Oroszlan,R.Donahue, and G.Kelloff,Proc. Am. Assoc.Cancer Res. 18:101, 1977), in the absence of complement, neutrali-zation wasobtained withanti-gp7O serum (Fig. 1B), but not with antiserum against p12 (Fig. 1A)orp30, p15,orplO (data not shown). In the absence of complement, anti-pl5(E) did not causeany greater degree of neutralization than did normal rabbit serum(Fig.
10).
1085
VOL. 33,1980
on November 10, 2019 by guest
http://jvi.asm.org/
Theapparentneutralizingactivity of the anti-gp7Oserum wasincreasedtwo- tofourfold inthe presenceofcomplement (Fig. 1B). As expected, noneof theantisera against thegag gene prod-ucts (p30, p15, p12, plO) caused virus neutrali-zation in the presence ofcomplement. In con-trast, anti-pl5(E), which didnothave neutral-izingactivity against RFVorCFV in the absence ofcomplement, exhibitedavery strong neutral-izingeffect in itspresence,butonly against RFV. Atthe dilutions used in theseexperiments,
com-z A
0
-so
Geo
40-o 0
plementalone did not have any effect on either virus, nor did heat-inactivated (560C, 30 min) complement,either in the presence or absence of anti-pl5(E) (datanotshown).
[image:4.504.69.463.193.342.2]To further assess immunological differences between the viruses, neutralizationassays were carriedout, usingserafromregressed mice and strain 129/J miceimmunizedwithRFV as test reagents. Bothserahad potentneutralizing ac-tivity against RFV and CFV which was en-hancedtwo- tofourfoldbycomplement(Fig. 2).
[image:4.504.88.447.409.602.2]DILUTION
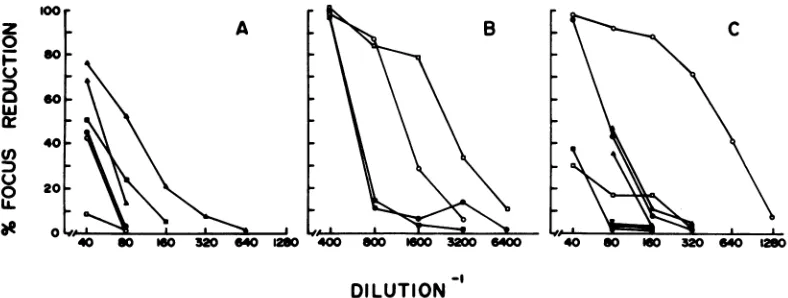
FIG. 1. Neutralization ofRFVandCFV bymonospecificgoatantisera.(A)Neutralization by anti-p12and
normalgoatserum;(B) neutralization by anti-gp7O; (C) neutralizationby anti-pl5E and normalrabbitserum.
Neutralization ofRFVbyantiserum (0, 0); neutralization of CFVby antiserum (0, U); neutralization of
RFVbynormalserum(A, A);neutralizationofCFVbynormalserum(V,V). Neutralization in the absence
of complement,closedsymbols;neutralization inthepresenceofcomplement,opensymbols.
100
z 0
F:
Lu
n a
C-)
0
80
60
40
20
0
40 80 160 320 640 1280 40 80 160 320 640 1280
-I
DILUTION
FIG. 2. NeutralizationofRFV and CFVbymousesera.(A)Neutralizationby regressedmouse serumand
normalmouseserum; (B)neutralizationbyimmunemouse serum.NeutralizationofRFVbyantiserum (0,
0); neutralizationofCFVbyantiserum(0, U;neutralizationofRFVbynormalserum(E, A);neutralization
of CFV by normalserum(V, V).Neutralization in the absenceof complement,closedsymbols;neutralization
in thepresenceof complement,opensymbols.
J. VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
STRUCTURE OF REGRESSING FRIEND VIRUS
Asmallbut consistentdifference in theactivity of these sera against the two viruses was
ob-served.
Immunological analysis ofviral
compo-nentsby neutralizationkinetics. The results
obtained in the experiments described above
wereconfirmed and extended, using
neutraliza-tion kinetics. For these assays, antibody at a
single dilutionwasaddedtothe twoviruses,and
the rate of reduction in focus formation was
1 O.S
1.00
0.5
0.2
0
4-(9
z
z
0
-)
4 0.1
.05
determined. In this way, small differences in
antigenicityorreactivityintheendpointassays
could be magnified andevaluated.
The kinetics of neutralization of RFV and
CFV pseudotypes of KiMuSV differed when
tested withserafromregressed mice,immunized
strain 129/J mice, or monospecific goat
anti-gp70 (Fig. 30). Since neutralization with the
latter serum gave results whichwere the con-verseof the results obtainedwith theregressed
A
_ BD
.02
.01
1.0
0.5
E F
0.2
0.1
.05
.02
F
.01 I , I
0 10 20 40 60 0 10 20 40 60
MINUTES
FIG. 3. NeutralizationkineticsassaysofRFV and CFV.(A)Neutralization of RFV in the absence ofserum
(0),orin thepresenceof normalmouse(O)ornormalgoatsera(A);(B) neutralization of CFV in the absence
ofserum(0), orin thepresenceof normalmouse (U)ornormalgoat(A); (C) neutralization ofRFV (open symbols) and CFV (closed symbols) in thepresenceof regressedmouse serum(0, 0), immunemouse serum
(El, ),oranti-gp7Oserum(A, A); (D)neutralization ofMoMuSVpseudotypes ofRFV andCFV by regressed, immune,oranti-gp70sera(symbolsarethesame asinC);(E)neutralization ofRFV-B23 andCFV-FES-7by regressed, immune,oranti-gp70sera(symbolsarethesame asinC);(F) neutralization of RFV (0,O,A)and CFV(0, Ui A) by three different pools ofserafrom regressedmice (0,O,A).
VOL. 33,1980 1087
on November 10, 2019 by guest
http://jvi.asm.org/
[image:5.504.101.405.160.578.2]orimmune sera,it isunlikelythat thedifferences in ratesof neutralizationweredueto aninherent susceptibility of RFV to neutralization or an excessofcompetingantigenin oneof the virus stocks.
Toeliminatepotential artifacts duetotheuse of KiMuSV focus-forming pseudotypes in the
assaymethod, similar determinationsweremade
usingpseudotypes ofMoMuSV, withthe same results (Fig.3D). Neutralization kineticsassays withthe intact Friend viruscomplexes,using in vivospleen focus fornation for virus titration, also showedasubstantial difference in therate ofreaction of RFV andCFV with anti-gp7O and serafromregressed mice (M. Dietz and P. Fur-manski,unpublisheddata).
Thesamedifferential sensitivitiestothe neu-tralizing antiserawereobtainedwhentwo
inde-pendently derived regressing and conventional virus stocks were compared by kinetics assay (Fig. 3E), and the distinction between RFV and CFV was also obtained when the assays were carriedoutusing three otherpoolsofserafrom regressed mice (Fig. 3F).Finally,KiMuSV pseu-dotypespreparedusingotherindependently de-rivedRFV and CFV stocks andaseriesof clonal isolates of theMuLV's of RFV andaCFV (Fig.
4) also showed thesamedifference in neutrali-zationrates asthe
parental
virusstocks.Neutralization ofvirusbyhumansera.A major difference was detectedin the endpoint neutralizationassays of RFV and CFV with
an-tiserumtop15(E)andcomplement. Neutraliza-tionof MuLV's byhumanserum has been
re-ported and has been showntobe dueto adirect and specific reaction between human
comple-ment component Cl and viral p15(E) in the absence of antibody (2). Complement is acti-vatedby thisreaction,resultinginvirolysis.We therefore used normalhumanserum asanother
reagent tofurther establish differences between the pl5(E)'s of RFV and CFV.
Individualhuman sera were titrated for activ-ity against RFV and CFV. A typical result is shown inFig. 5A.Thekinetics of neutralization of the viruses by this same human serum is shown in Fig. 5B. A clear distinction was ob-served between theratesofreaction of RFV and CFV with the humanserum. Nosignificant neu-tralizationwasdetected ifthe human serum was heat inactivated
(560C,
30min) before use, or if guinea pig complement, which itself does not neutralize MuLV's,wasadded to the heat-inac-tivated humanserum. Thus, theneutralizationof the viruses observed in theseexperiments was likely duetocomplement and its reaction with
anti-p15(E)
asreported(2),andnot to antibody or complement-dependent antibody in the hu-manserum.Thesignificanceof this reactivity with normal human serum lies in the difference it demon-strates in the p15(E)'s of RFV and CFV.
Fur-thermore, since the resultswith human serum are the converse of the results obtained with neutralization of RFV and CFV byrabbit anti-serum to MuLV p15(E) plus complement, the difference inreactivity is probablynotdueto an inherent difference in susceptibilitytovirolysis, differences in virus concentration or the pres-enceofcompeting proteins, anti-complementary
activity, orother artifacts inthe virus prepara-tions. However, the exact role in regression of this difference in the pl5(E)'s remains to be determined.
PAGE analysis of viral proteins. The structural basis for the immunological
differ-1.0
0.5
100
Zs 0
u
0
a
w
In
0 $0
0
N1- 0.2 _
0 10 20 40 60
DILUTION' MIN.JTES MINUTES
FIG. 4. Neutralization (A) andneutralization ki- FIG. 5. Neutralization kineticsofclonal isolates netics(B) ofRFV(0)andCFV(0)inthepresenceof oftheMuLVofRFV(0)andCFV(0)inthepresence
normalhumanserum. ofimmune129/Jmouse serum.
0.1
0.05
lI i, .
0 10 20 40 60
on November 10, 2019 by guest
http://jvi.asm.org/
[image:6.504.73.453.444.646.2] [image:6.504.72.271.482.620.2]STRUCTURE OF REGRESSING FRIEND VIRUS
encesdetected in the neutralizationassays was firstinvestigated by comparing thepolypeptide
composition ofarepresentativeRFVandCFV,
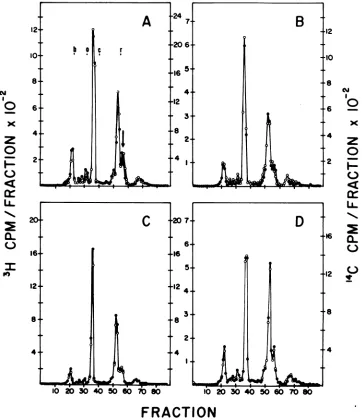
using SDS-PAGE in 15% gels preparedby the method of Laemmli (23). We found no major differences between the viruses; both prepara-tions contained all of the resolvable viral pro-teins and glycoproteins in approximately the same proportions. However, analysis on the same gel of admixtures of these two viruses labeled with different isotopes (Fig.6A)revealed
thattherewas asmall, reproducibledifference inthe mobilities ofone of the smnll viral pro-teins,identifiedasp12onthe basis ofmigration
and coincidence withimmunoprecipitated viral p12 (datanotshown). Reciprocal labeling(Fig. 6B)andelectrophoresis of mixtures of thesame virustypelabeled with eachisotope(Fig. 6C and D) established that the difference observed in the p12 was notduetolabelingor
electropho-reticartifacts. The difference inelectrophoretic
mobility of the p12 proteins of thetwoviruses
24 7
-20
6-16 5
4.
.92
t8
C
l0 20 30 40 50 60 780 4.
2+
'4
1
.
20
7-B
64
*16
54.
12 4
8s
3t
2-.4 Il
'0 -30i 0i-0 6 8
lb 2-0
30 40 50 60 70 '12
10
-8
a
6 _
x 4
0
2 -,
H
0.~
06
20
12 0)
[image:7.504.64.426.194.614.2].4
FRACTION
FIG. 6. SDS-PAGE analysis of RFVandCFV.Symbols:(A)3H-labeledRFV(0), '4C-labeledCFV (0):(B) 'H-labeled CFV(0), 14C-labeledRFV(0);(C)3H-labeledRFV (0),14C-labeledRFV (0): (D)3H-labeledCFV
(0), 4C-labeked CFV(0). Arrow inAindicatesthe positionof viralp12.Migrationofthemolecular weight
markers: b,bovineserumalbumin;o,ovalbumin;c,chymotrypsinogen;r,RNase.
12
10o
8
A
I
N I 0
x
z
0
cr
N.L.
CL
6 4 2Z
20
16+
12+
8+
4+
W--. a
4
1089
VOL. 33,1980
JT
D
8e
on November 10, 2019 by guest
http://jvi.asm.org/
corresponded to an apparent difference in mo-lecular weight of about 600 to 800, as calculated from the relative migration of standard proteins. The high-resolutionPAGE system of Monte-laro et al. (24, 25) was employed to further analyzethepolypeptidecomposition of RFV and CFV. In this gel system the two viruses were
N
0
x
z
0
0
LL
Um.
0
65
4
3
2
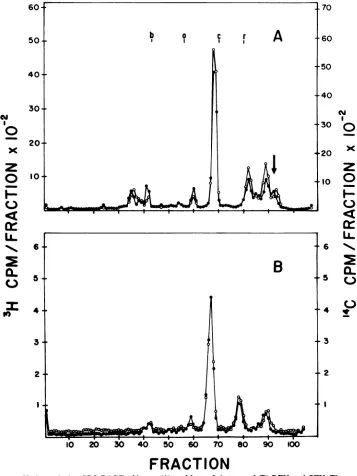
composed of thesame polypeptides in approxi-mately the same proportions, with one of the smallproteins,identified as p12 (25), differing in electrophoreticmobility (Fig. 7A).
Montelaro et al. have previously demon-strated that the proteolytic enzyme bromelain will remove the major external glycoprotein
;70
°
Cr
A
60
-50
.40
_30
O
-20
z
10 0
w6
.6 s%
B
a.)
4 (m
3
2
[image:8.504.88.445.150.626.2]lb
20 30 40 50 60 70 80 90 i00FRACTION
FIG. 7. High-resolutionSDS-PAGEofintact(A)andbromelain-treated(B)RFV and CFV. Thearrowin
A indicates theposition ofviralp12.Symbols: 3H-labeledRFV(0), 14C-labeledCFV(0).Migrationofthe
molecularweightmarkers:b,bovineserumalbumin;o,ovalbumin;c,chymotrypsinogen, r,RNase.
on November 10, 2019 by guest
http://jvi.asm.org/
STRUCTURE OF REGRESSING FRIEND VIRUS
gp7Oand thep15(E) from intact virus particles without affecting the internal (inaccessible)
polypeptides p30, p15, p12,andplO (25).When
RFV andCFVweretreated withbromelain,the gp7O and p15(E) peaks of both viruses were
eliminatedorreduced(Fig. 7B). The other com-ponents, including the peak which showed a
difference inelectrophoreticmobility,the
(inter-nal) p12,remained intact.
Trypticpeptidemappingof viral compo-nents.Tofurtheranalyze the structural differ-ences between RFV and CFV, tryptic peptide
maps werepreparedof eachof the virion
com-ponents, isolated, and purified by
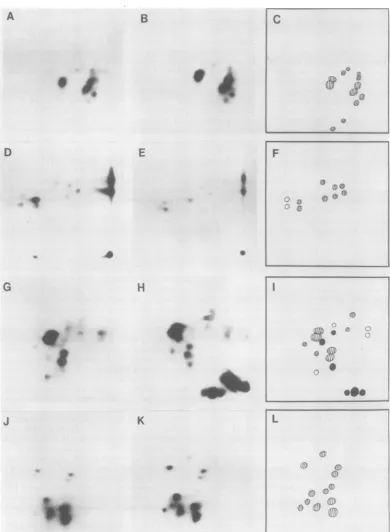
immunopre-cipitation withmonospecificantisera and SDS-PAGE. The tryptic peptide maps for the gag geneproductsareshown inFig.8.
Amongthegaggeneproducts, the p12protein
was significantly different in RFV and CFV, confirming the differences observed in electro-phoreticmobility.Thep30'sandp10'swere iden-tical. Thep15 showedtwoadditional minor pep-tidesintheprotein isolated fromCFV,but be-causetherewerenoreciprocaldifferencesinthe map ofp15 from RFV andno apparent differ-encein molecularweights of theproteins (Fig.
7), these addedpeptidesmayhave been dueto contamination.
Both env gene products, gp70 and p15(E), were different in RFV and CFV (Fig. 9). For gp70, the differencewasdueto ashiftin asingle majorpeptide.Incontrast,thedifferencesinthe p15(E) weremuchmorecomplex, and onlyone ofthe 12distinguishablepeptides could be con-sideredcommon tothetwoviruses.
To determinewhether these structural differ-enceswererelated totheregressingphenotype
of the viruses, tryptic peptide maps were pre-pared from theproteins isolated fromtwo
inde-pendently derived stocks of RFV and CFV (RFV-B-23andCFV-FES-7) andclonal isolates of the MuLV's ofRFVand CFV. Thedifferences observed inthe gp70's ofRFV and CFV were also observedinthe other stocksandclones(Fig. 10). Similar results were obtained with the
p15(E)'s (notshown). Incontrast, thep12 pro-teins of each of these viruses gave identical
tryptic peptide maps, andhence differences in thep12 donotappear tobe relatedtoregression.
DISCUSSION
We havecompared thestructural components of two strains of Friend virus: a conventional strain, CFV, which induces a progressive and lethal erythroleukemia, and the RFV strain, whichinduces an erythroleukemiathat sponta-neously regresses. Two approaches have been used to make these comparisons: physico-chem-ical, based on electrophoretic mobility and
tryp-ticpeptidemapping, andimmunological, based onneutralizationassays.
SDS-PAGE analysis revealed that RFV and CFV both consisted of all of thegenerally ob-servedmajor viral proteins inapproximatelythe same proportions. One polypeptide, p12, ex-hibitedasmall, consistent difference in molecu-lar weight(mobility) in thesetwoviruses.
Furtheranalysis of the structuralcomponents of the viruses was achieved by using tryptic peptide mapping of the isolated viral proteins and glycoproteins. The difference in the p12 detected by electrophoresis was confirmed by this method. In addition, we found that the primarystructureofthe twoenvelope constitu-ents,gp70 and p15(E),differedinRFV andCFV. The p30's and p10's of the two viruses gave identical tryptic peptide maps, and, with the exception oftwo additional minor peptides in theprotein fromCFV, the p15'swerealso iden-tical.
The p12of the MuLV's isaninternal, acidic, RNA-bindingphosphoprotein (33).Itexhibitsa very high degree of type specificity, both in terms ofantigenic structure (34) and function-ally,in termsof its interaction with viral RNAs (31). In additiontodifferences in primary amino acidstructure,changesinthepatternordegree ofphosphorylation might also beassociated with differences observed betweentwoviruses.Itis, therefore, perhaps notsurprising that compari-sonofanRFVandaCFVmight reveal different p12's. However, when additional virus stocks and clonal isolateswereexamined, the difference in p12was not aconsistentfinding,andtherefore isnotrequired for induction ofaleukemia that spontaneouslyregresses.
Both env gene products, pg7O and p15(E), yielded different tryptic peptide maps for the two viruses. The differences observed between the p15(E)'s were extensive, whereasa change in onlyone tyrosine-containing tryptic peptide distinguished thegp7O's. Elderetal. have made adetailedanalysis ofthegp7O's of MuLV'sand havecategorized the virusesonthebasis of the peptidemaps of theirgp7O's (9).Recombinant viruses have alsobeenidentified by analysis of the tryptic peptidemapsoftheirgp7O's (8, 11). The differenceswehaveobserved between RFV and CFV might be the result ofa recombina-tional event which occurred during the genesis of RFV.
It is of interestthat thep30'sofRFV and CFV gave identical tryptic peptide maps in view of reports from severalinvestigators on differences betweenp30'sofcloselyrelatedMuLV's. Buch-hagenetal. reporteddifferencesintryptic pep-tide patterns forp30's isolatedfrom N- and B-tropic viruses derived from a single BALB/c 1091
VOL. 33,1980
on November 10, 2019 by guest
http://jvi.asm.org/
B
d.
0 .:
K
J
0
c
li~~~~i
'I~~~~~~~~~~~~~~~
1")~~~~~~~~~~~~~~~~~~~~~~
M.4t~~~~~~~~~~~~~~~~~~~~~~
.II
0*OR
L
*1,
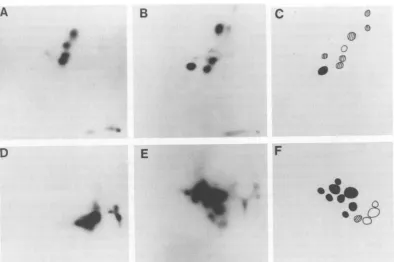
FIG. 8. Tryptic peptideanalysis ofgag geneproducts of RFV and CFV. (A) RFV p3O; (B) CFV p3O; (C) composite ofp30maps;(D) RFVpl5; (E) CFVpl5; (F) composite ofp15maps;(G) RFVpl2; (H) CFVp12;(I) composite ofp12maps;(J)RFVplO; (K) CFVplO; (L) composite ofplOmaps.Electrophoresiswascarriedout from righttoleft; thin-layerchromatography in the second dimensionwasascending from the bottom. In the composites, peptides uniquetoRFVareindicated inwhite,peptides uniquetoCFVareindicated inblack, andcommonpeptidesarestriped.
A
D
E
.4.
I.S
S*
G
H
A a
U.
S' 0
on November 10, 2019 by guest
http://jvi.asm.org/
[image:10.504.72.462.58.590.2]STRUCTURE OF REGRESSING FRIEND VIRUS 1093
A
B
I
D
E
C
9
F
FIG. 9. Trypticpeptide analysis of env gene products of RFV and CFV.(A)RFVgp7O;(B)CFVgp7O; (C)
composite ofgp7Omaps;(D)RFVpl5(E);(E) CFVpl5(E); (F) composite ofpl5(E) maps. Symbolsarethe same
asinFig.8.
A * B C D
*
e
04
0
e_s
F G
Ab,-"W.,,:.At
. _'
.-.
H
9
.._ m
a *.*.
FIG. 10. Tryptic peptideanalysisof thegp7O(A-D) andp12 (E-H)ofRFV-B-23 (A,E),CFV-FES-7 (B, F)
andclonalisolates of theMuLVof RFV (C, G) and CFV (D, H).
mouse(3).Hopkinsetal. showed that
p30's
from NB-tropic MuLV's haddifferentelectrophoretic mobilities than the parent B-tropic virus (18), and Gautsch et al. have found subtledifferences inp30tryptic peptide maps that areassociated with virus tropism (15). We have previouslyshown that RFV and CFV differ in tropism
(RFV is NB-tropic, whereas CFV is N-tropic)
andthatthisdifference
apparently
relatestothe regressingphenotype of the virus (4). Itmightthusbeexpectedthat thep30'sofRFVandCFV would also differ. However, Hopkins et al.
re-VOL. 33,1980
on November 10, 2019 by guest
http://jvi.asm.org/
[image:11.504.47.441.62.324.2] [image:11.504.52.442.369.560.2]ported that there was little to no difference in thep30'sof N and NB viruses, incontrast to the clear differencebetween NB and B isolates (18), and themajor differences observed by Gautsch et al. among the p30's ofalarge series of eco-tropic MuLV's was between N- and B-tropic viruses (15). Itistherefore possible thata change in p30, although relatedtotropism,is not nec-essarily associated with theN to NBconversion, or that RFV and CFVareexceptional with re-spect to their identical p30's. It is also possible that subtle differences in the p30's would not havebeendetected byourmethods.
Neutralization kineticsassays werechosento assessimmunological differences between RFV and CFV.Neutralizationassays,although inher-entlyrestrictedtoviral surfaceconstituents,can yieldinformative results independent of differ-ences in theprimary structure ofthe proteins tested. This is becauseadifference in the struc-tureofanexternalprotein oftwoviruses might notbereflected inadifference inneutralization. This could result if the changes were in an inaccessibleportion of the molecule when incor-porated into whole virusparticles and thus be irrelevanttotheprocessofinfectionor neutral-ization. Likewise, it is conceivable thattwo vi-ruseswhose externalproteinswereidentical in primary structure might differ in reactivity to
neutralizing antibodybecause of conformational or accessibility differences induced by juxta-posed internal or adjacent surface proteins
themselves having differentstructures. Invivo neutralization kineticsassayshave been success-fully usedto distinguishamongdifferent pseu-dotypes of Friendspleenfocus-formingvirus(7).
By using the neutralization assays, we de-tected differences inreactivityof RFV and CFV withanti-gp70 serum, serafrom regressed and immunizedmice,andsera (andhuman
comple-ment) reactiveagainst anti-p15(E).The neutral-izationof RFVbyanti-pl5(E) andcomplement
is likely duetovirolysis(27). The effect of hu-man serumontheviruses,which is dueto viro-lysis mediatedbydirectcomplementactivation by viralp15(E),shows thatCFV alsoexpresses p15(E)onits surfaceand issusceptibletolysis.
Thus, both RFVandCFVapparentlyhavetheir
p15(E)'saccessibleonthe viralsurface,but with distinctantigenicsitesexposed. Fischingeretal. reported that (conventional) F-MuLV was not
neutralizedbyanti-pl5(E)andcomplement (13).
Incontrast, axenotropicMuLVwassusceptible.
It is possible, therefore, that the p15(E) sites exposedonRFVarerelatedtoxenotropicvirus sequences acquired during the genesis of this strain.
Thestudiesreported here have been carried
outprimarily byusing the in vivo-derived Friend viruscomplex,consisting of spleen focus-forming virus andthehelperMuLV, as startingmaterial for preparation of virus stocks in cultures of mousefibroblasts in vitro.With someexceptions (6, 37),however, growth of spleenfocus-forming virus in short-term fibroblast cultures is re-stricted or atleastoccurs atmuch lower levels thanthe MuLV helper.Furthermore,theMuLV specifies the antigenicity of the complex (7). Also, data obtained withthevirus complex were confirned by using clonal isolatesof theMuLV's from both RFV and CFV. Therefore,ourresults on both physico-chemical and immunological differences between RFV and CFV reflect, pri-marilyorexclusively, differences in theirMuLV components. This isjustified by the results of previous studies which established that the re-gressing phenotype of RFV is entirelyafunction of its MuLVhelper (4).
Comparisonof thestructuresof RFV and CFV might be instructivewith regardto the origin of theregressing strain. Our previous studies have established that viruses that inducearegressing disease can be generated from conventional strainsby forcedpassagethroughaspecific sub-line of Fv-1 heterogeneous Swiss/ICRmice(4). In these circumstances, virus tropism changes from N to NB and the resultant virus stock inducesanerythroleukemiathatspontaneously regresses. Recombination between MuLV's has been reported in several systems, and these eventsand thederivative virus strainsare con-sideredto beimportant inthe pathogenesis of thevirus-inducedleukemias (12, 16). The SFFV componentof theFriendvirusleukemias is itself a recombinant between its ecotropic MuLV helper andamouseendogenous xenotropic virus (36), and non-replication-defective recombi-nantsbetween thehelper MuLV of Friend virus andxenotropicviruses have beenreportedafter passage of the isolated helper in NIH/Swiss mice(38).Thus,itistemptingtospeculatethat RFVarose byaspecificrecombinationalevent
involving the helper MuLV component of the
parental CFV and endogenousmouse viral
se-quences to yield a new virus strain with the regressingphenotype.Several ofourfindingsin this study,as noted above,areconsistentwith this hypothesis, although altemative
explana-tions havenotbeenexcluded.
The exact role of any of the structural or
antigenic differences between CFV and RFV in theprogressionorregressionofleukemiacannot
bedeterminedatpresent. This isespeciallytrue
ofthe structural differenceswhich may be
to-tally
unrelatedtothebiological
behavior ofthe viruses. It is ofinterest,however,thatthe viralJ. VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
STRUCTURE OF REGRESSING FRIEND VIRUS 1095
components that exhibit significantdifferences are related toimmunological reactivity against
the virus and leukemia cells which develops duringregression. The two env gene products, gp7O and p15(E), are clearly involved in neu-tralization of viralinfectivity.The enhanced
sus-ceptibility of RFVtoneutralizationby regressed serumandanti-p15(E) antiserum couldhelpto limitdiseaseprogression by preventing reinfec-tion oferythroblastic stemcells. Furthermore,
these sameviralcomponentsarepresentonthe surfaces of infected cells andserve astargetsfor cytotoxic reactions mediatedbytheappropriate
antisera andcomplement. Thus,if the structural and antigenic differences found in the compo-nents of RFV and CFV alter their in vivo
im-munogenicity or immunosensitivity, profound
changes in the dynamics of the disease might
result, including its regression. Further studies willberequiredtoestablish theexact relation-ship betweenthese changesand the biological
properties of the viruses.
ACKNOWLEDGMENTS
Wethank E. Fleissner and S. Oroszlan for theirgiftsof
anti-p15(E)antisera and S.Marcusfor his criticalreadingof themanuscript.
This studywas supported by grant CA-14100 from the National Cancer Institute andaninstitutional granttothe MichiganCancer Foundationfromthe United Foundationof Greater Detroit.
LITERATURE CITED
1.Aaronson,S.A., and W. P. Rowe. 1970. Nonproducer clones of murine sarcoma virus transformed Balb/3T3 cells.Virology 42:9-19.
2. Bartholomew,R.M., A. F. Esser, and H. J.
Muller-Eberhard. 1978. Lysis ofoncornaviruses by human serum.Isolation of the viral complement(Cl)receptor andidentificationaspl5E.J. Exp. Med. 147:844-853. 3. Buchhagen, D. L.,0.Stutman, and E. Fleissner. 1975.
Chromatographicseparation and antigenic analysis of proteins of theoncornaviruses.IV. Biochemical typing of murine viralproteins. J. Virol.15:1148-1157. 4.Dietz, M., S. P. Fouchey, C. Longley, M. A. Rich, and
P.Furmanski. 1977. Spontaneous regression of Friend virus-inducederythroleukemia.I. The role of the helper murineleukemia virus component. J. Exp. Med. 145: 594-606.
5. Dietz,M., P.Furmanski,R.Clymer, and M. A. Rich. 1976.Theeffects of thymectomy and anti-thymocyte serumon spontaneous regression of Friend virus in-ducederythroleukemia.J. Natl. Cancer Inst. 57:91-95. 6. Eckner,R. J.1975.Continuousreplicationof Friend virus complex (spleen focus-forming virus-lymphatic leuke-mia-inducing virus) in mouse embryo fibroblasts.
Re-tention ofleukemogenicity andlossof
immunosuppres-siveproperties. J. Exp. Med.142:936-948.
7. Eckner,R.J., and R. A.Steeves.1972. A classification ofthe murineleukemia viruses. Neutralization of pseu-dotypes of Friend spleen focus-forming virus by type-specificmurineantisera. J. Exp. Med. 136:832-850. 8.Elder, J. H., J. W. Gautsch, F. C. Jensen, R. A.
Lerner, J. W. Hartley, and W. P. Rowe. 1977. Bio-chemical evidencethat MCFmurineleukemiaviruses
are envelope (env) generecombinants. Proc.Natl. Acad. Sci. U.S.A. 74:4676-4680.
9.Elder, J. H., F. C. Jensen, M. L. Bryant, and R. A. Lerner. 1977. Polymorphism of the major envelope glycoprotein (gp70) of murine C-type viruses: virion associated and differentiation antigens encoded by a multi-gene family. Nature (London) 267:23-28. 10. Elder, J. H.,R.A. Pickett, J. Hampton, and R. A.
Lerner. 1977. Radioiodination of proteins in single polyacrylamide gel slices. J.Biol. Chem. 252:6510-6515. 11. Fischinger,P.J., A. E.Frankel,J.H. Elder,R. A. Lerner, J. N. Ihle, and D. P. Bolognesi. 1978.
Bio-logical, immunologicaland biochemical evidence that HIX virus is a recombinant between Moloney leukemia virus and a murine xenotropic C type virus. Virology 90:241-254.
12.Fischinger, P. J., J. N.Ihle,F.deNoronha,and D. P. Bolognesi. 1977.Oncogenic and immunogenic poten-tial of cloned HIX viruses in mice and cats. Med. Microbiol. Immunol. 164:119-129.
13.Fischinger, P.J., W.Schiifer, and D. P.Bolognesi.
1976. Neutralization ofhomologous andheterologous
oncornavirusesby antiseraagainstthep15(E)andgp7l polypeptides of Friend murineleukemia virus. Virology 71:169-184.
14.Furmanski, P., J. Baldwin, R.Clymer, and M. A. Rich. 1975. Spontaneous regression of Friend virus inducedleukemia: coinfection with regressing and con-ventional strains of virus. Science 187:72-73. 15.Gautsch,J.W.,J. H.Elder,J.Schindler,F. C.Jensen,
and R. A.Lerner.1978.Structural markers on core proteinp30of murineleukemia virus:functional corre-lation withFv-1tropism. Proc. Natl. Acad.Sci. U.S.A. 75:4170-4174.
16.Hartley, J. W., N. K.Wolford, I.. J. Old, and W.P. Rowe. 1977. A new class ofmurine leukemia virus associated withdevelopment of spontaneous lympho-mas. Proc.Natl. Acad.Sci. U.S.A. 74:748-792. 17. Hines, D., M.Dietz,M. A.Rich,and P.Furmanski.
1978. Active immunotherapy of Friend virus-induced erythroleukemia and its spontaneous regression. Cancer Immunol. Immunother.5:11-16.
18. Hopkins, N.,J.S.Schindler,and R.Hynes. 1977.Six Ntropic murine leukemia viruses derived from a B-tropic virus of BALB/c have alteredp3O.J. Virol. 21: 309-318.
19. Hunsmann,G.,M.Chariez,V.Moennig,H.Schwarz,
and W.Schiifer. 1976.Properties of mouse leukemia viruses. X. Occurrence ofviral structural antigens on thecell surfaceasrevealed byacytoxicitytest.Virology 69:157-168.
20.Kende, M.,B.Sass,S.Hino,R. M.Donahoe,and G. J.Kelloff. 1978.Type-Cvirusstructuralantigens on the surface of the infected cell asdetermined by a humoralcytotoxicity assay. Int. J. Cancer 21:194-203. 21. Kessler, S. W.1975.Rapid isolation of antigens from cells with a Staphylococcal protein A-antibody absorbent: parameters ofthe interaction ofantibody-antigen com-plexes with proteinA.J.Immunol.115:1617-1624. 22. Klement, V.,and M. 0. Nicolson. 1977.Methods for
assays ofRNAtumorviruses, p. 59-108. In K. Mara-moroach and H. Koprowski (ed.), Methods in virology, vol. VI.Academic Press Inc., New York.
23. Laemmli,V. 1970.Cleavageof structuralproteins during theassembly of the head of bacteriophage T4. Nature (London)227:680-685.
24.Montelaro,R.C.,and R.R. Rueckert.1975. Radiola-belingofproteinsandvirusesinvitro by acetylation withradioactive acetic anhydride. J. Biol. Chem. 250: 1413-1421.
25. Montelaro, R. C., S. J.Sullivan, and D. P. Bolognesi. 1978. Ananalysis of type-C retrovirus polypeptides and VOL. 33,1980
on November 10, 2019 by guest
http://jvi.asm.org/
theirassociations in the virion. Virology84:19-31. 26. Mosser,A.G., R. C. Montelaro,and R. R. Rueckert.
1975.Polypeptide composition of spleennecrosis virus,
areticuloendotheliosis virus. J. Virol. 15:1088-1095. 27. Oroszlan, S., and R. V. Gilden.1978.Immune lysis of
type-C viruses by antiseratopurified viral subunits,p.
90-93InP. Bentvelzen, J. Hilgers and D. S. Yohn (ed.), Advances incomparative leukemia research-1977. El-sevier/North-Holland Publishing Co., Amsterdam. 28. Rich, M. A., R. Clymer, and S. Karl.1969.Spontaneous
regression of virus induced murine leukemia. fl.
Influ-enceof environmental factors. J. Natl. Cancer Inst.43: 571-577.
29. Rich,M.A.,S.Karl,andR.Clymer.1971.Complement fixation in conventional and regression virus induced murineleukemia. J. Immunol. 106:1488-1492. 30. Rich,M.A.,R.Siegler,S.Karl,and R.Clymer.1969.
Spontaneous regression ofvirusinducedmurine
leuke-mia.I.Host-virussystem.J.Natl. CancerInst.
43:559-570.
31.Sen, A.,C. J.Sherr,and G. J. Todaro. 1976.Specific binding of thetype Cviralcoreprotein p12 with purified
viral RNA. Cell 7:21-32.
32. Steeves, R.A.,M. Strand, and J. T.August. 1974. Structuralproteinsofmammalianoncogenic RNA
vi-ruses:murineleukemia virus neutralizationbyantisera prepared against purified envelope glycoprotein. J.
Vi-rol.14:187-189.
33. Stephenson, J. R., S. G. Devare, andF. H.Reynolds. 1978. Translational products of type-C RNA tumor
viruses. Adv.Cancer Res.Volume 27:1-53.
34.Stephenson, J. R., S. R. Tronick, and S. A. Aaronson. 1974.Analysis oftypespecific antigenic determinants oftwostructuralpolypeptides ofmouseRNAC-type viruses.Virology 58:1-8.
35.Swanstrom, R.,and P. R. Shank. 1978.X-ray intensi-fyingscreensgreatly enhance the detection by autora-diography of the radioactive isotopes32P and 125I. Anal. Biochem.86:184-192.
36.Troxler,D.H.,D.Lowry,R.Hawk,H.Young,and E. M.Scolnick.1977.Friendstrain ofspleen focus-form-ing virus isarecombinant betweenecotropic murine
typeC virus andtheenvgeneregion of xenotropictype Cvirus. Proc. Natl. Acad. Sci. U.S.A. 74:46714675. 37.Troxler,D.H.,W. P.Parks,W. C.Vass,and E. M.
Scolnick. 1977. Isolation ofafibroblastnonproducer cell linecontainingthe Friend strain of thespleen focus-formingvirus.Virology76:602-615.
38.Troxler, D. H., E. Yuan,D. Linemeyer, S.Ruscetti,
and E. M.Scolnick.1978.Helper-independentmink
cellfocus-inducingstrains of Friendmurine type-C
vi-rus:potential relationshiptothe origin of
replication-defective spleenfocus-forming virus. J. Exp.Med.148: 639-653.
on November 10, 2019 by guest
http://jvi.asm.org/