Copyright01975 AmericanSocietyforMicrobiology PrintedVol.in15,U.SAA.No.
Products of
Complementation Between
Temperature-Sensitive
Mutants of Simian Virus 40
JANICE YANG CHOU ANDROBERT G. MARTINNational InstituteofArthritis, Metabolism,andDigestiveDiseases, Bethesda, Maryland 20014 Receivedforpublication 27 June 1974
Temperature-sensitive mutants of the D complementation group ofsimian virus 40 exhibit delayed complementation. Analysis ofthe thermal stability,
kinetic profilesintemperatureshiftexperiments, and progeny of complementa-tion have led to the hypothesis that delayed complementation is not true
complementation, but the result of a very low level ofleakiness, followed by
phenotypic mixing of the progeny D mutants. This hypothesis is consistent with the proposal that D mutants are defective in uncoating. In the course of these experiments, it was observed that fresh medium suppresses the growth of D mutants atthe restrictivetemperature.
From complementation analyses, tempera-ture-sensitive mutants of simian virus 40
(SV40) have beendivided into classesA, B, C,
BC, and D (3).
MutantsoftheB, C and BCclasses havebeen
designatedas"late" becauseoftheircapacityto
synthesize viral DNA under restrictive
condi-tions (2, 10). Recent studies suggest that these mutants may map within a single cistron (6;
C.-J. Laiand D. Nathans, Cold
Spring
HarborSymp. Quant. Biol., in press). Therefore, the
complementation between B and C mutants
may represent intra- rather than intercistronic
complementation. One of several possible
ex-planations offered for the existence of the BC
classwasthat the virions of suchmutants were
particularly sensitive to heat inactivation (3).
Data presented in this paper exclude that
possibility.
Mutants of the A and D classes have been
designated as "early" on the basis of their
failuretosynthesizeviral DNA uponincubation at the restrictive temperature (2). The A func-tion isrequiredtoinitiate thereplicationofviral DNA molecules (2, 10). DNA synthesis ceases when cultures infected with mutants of the A group are shifted from the permissive to re-strictive temperature. D function, on the other hand, isapparently required priorto viral DNA replication. When cultures infectedwith D mu-tants are incubated in depleted medium at the permissive temperature, andthenshiftedtothe restrictive temperature, viral DNA synthesis continues unabated, provided the incubation has beenallowedtoproceedfor 10to 20hatthe permissivetemperature (2).Inthe course ofthis investigation, itwasobserved thatthelengthof timerequiredatthe permissive temperatureto
overcomethe D virion's defectwasconsiderably reduced when fresh rather than depleted me-dium was used. Various interpretationsof this observation are discussed.
It has been proposed that D mutants are unableto "uncoat" attherestrictive tempera-ture(9), i.e., thatsomevirioncomponent hasto be removed before expression of the viral ge-nome, and that this component remains associ-ated with the genome when D mutant virions infect monkey cells at40 C. This proposal was based on the observations that (i) D101 DNA was infectiousfor oneround ofreplicationatthe restrictive temperature yielding temperature-sensitivevirions; (ii) D101 virions were noncom-plementing when tested after 3 days at 40 C; and (iii) D101 virions failed to exhibit any known early functions at 40 C although they adsorbed to, and penetrated monkey cells
nor-mally (9). The fact that 10 to 20 h of incubation
indepleted medium was required to overcome. the defect exhibited by D mutants (2) neither supported nor negated the uncoating hypothe-sis.
Newdata suggestthat the D mutants mapin that portionofthegenomewhosetranscription
only commences late, i.e., after viral DNA
synthesis has started (6, C.-J. Lai and D.
Nathans, Cold Spring Harbor Symp. Quant. Biol.,inpress; T. E. Shenk, C. Rhodes, P. W. J.
Rigby,andP. Berg, (Cold Spring Harbor Symp.
Quant. Biol.,inpress). These resultsimplythat
(i) the D product may be synthesized late in infection; (ii)therefore, any defect exhibited by D virionsearlyininfectionprobably results from the introduction ofthedefective D product by thevirionsandnot fromtheearly synthesis of a defective Dprotein(in supportoftheuncoating 127
on November 10, 2019 by guest
http://jvi.asm.org/
CHOU AND MARTIN
proposal); and (iii) D mutants should be referred to as late mutants whose phenotype is ex-pressed early.
An apparent contradiction to the proposal that D mutants are defective inuncoating arose from complementation experiments (3). Al-though D mutants failto exhibit complementa-tion after 3days of co-infection with representa-tives of the other mutant classes (3, 9), com-plementation is observed after 2 weeks of co-infection-"delayed complementation" (3). This paper primarily concerns an investigation oftheprogeny obtained after co-infection with D mutants and temperature-sensitive mutants of the other classes. An hypothesis to account for delayedcomplementation is presented.
MATERIALS AND METHODS
Virus and cells. The virusstocks, mutants, and CV-1 cells used have been described(3).Theprimary AGMK cells were supplied by Flow Laboratories, Inc., Rockville, Md., and the BSC1 cellswerekindly provided by D. LeBlanc, and tested by him to verify thatnoinduction of host DNA synthesiswasobserved afterSV40 infection.
Media, plaque, and slant agar assays. The modi-fied nutrient mix F-12 and modified Eagle media (3E), as well as the plaque assay and slant-agar technique forcomplementation, have been described (3, 8). Depleted medium was the 3E5 (5% serum) medium removed after 4 to 5 days from confluent CV-1 cells.
Thermal inactivation of virus. Stocks of the mutant virions were diluted 50-fold with phos-phate-bufferedsaline, (8), andincubatedat50C.
Kinetics of temperature shift. Confluent mono-layers of CV-1 cells in the 24-well (1.6-cm diameter each) plates of Linbro Chemical Co., New Haven, Conn., wereinfected with themutants at multiplici-ties ofinfection(MOIs) of2 to5,after removal ofthe old medium. The viruswasallowedtoadsorb for2h at 33C, and 1 ml of fresh 3E5 mediumor depleted medium per well was added. At appropriate times, cultures were shifted to 40C. In addition, acontrol was prepared in which the adsorption aswell asthe incubation was carried outat 40C. At 96 h postinfec-tion, all of the cultures were frozen, thawed and titeredinFalcon25cm2flasksat 33C.
Products of complementation. Confluent CV-1 monolayers in the 24-well Linbro plates were pre-pared. The medium was removed, and thetwo mu-tants or mutantand wild-typevirions suspendedin 0.2to 0.3mlof3Ewereallowedtoadsorbat 40C for2 to 3 h. Fresh 3E5 (1 ml/well) was added, and the plateswereincubatedfor 96 h at 40Cwhen wildtype was co-infected with the mutants. The monolayers werealmostfully lysedby thistime.Theplateswere incubatedfor 13daysat 40C whencomplementation between twoSV40mutants wereinvestigated. Com-plete lysis was seen by 5 to 6 days in all but the
co-infected cultures containing Dmutants(whichwere not fully lysed even by 13 days).The cultureswere frozen,thawed and titeredat33C. Individualplaques were picked with asterile Pasteurpipette. The agar plugs weresuspended in 2 ml ofphosphate-buffered saline inscrew-cap tubes and storedat 4C. Totest the plaque isolates when wild-type virions were in-volved in theexperiment, 0.2 ml of each suspension wasaddedtofresh confluentCV-1 monolayers in the 24-well plates. After 2 to 3 hof adsorption at 40 C, medium in agar was added, and the cultures were incubated 13days at 40C with feeding at 3days and7 dayspostinfection. After staining, plaque containing wellswerescored aswild type.
The plaque isolates fromthe crosses between two temperature-sensitivemutants werepickedasabove, andstocksweregrowninthe 24-wellLinbroplatesat 33 C. These stockswerethentested for progeny type by the agar-slant complementation technique (3). This technique measures delayedcomplementation, sothatall mutants, including D mutants, complement by this assay. The progeny were tested for com-plementation in each case against both parental types. No wild-type recombinant virus were found amongthe limited number of progeny examined, as judgedbythe fact thatnoneappearedtocomplement bothmutantparental types. The results withA x B, A x C, and B x Cwerecompletelyunambiguous, but theD x A, D x B, and D x C resultswere occasion-ally difficult to interpret. (The agar-slant technique yields ambiguous results in about 10% ofall crosses
[31).
To distinguish D from B or from C progeny, the followingtechnique,basedonthe fact thatDmutants are unableto synthesizeviral DNA at 40C (2), was employed. Duplicate, confluent monolayers in the 24-wellLinbroplateswereinfected withafewdrops of the progeny virus stocks. After adsorption at the appropriate temperature for 2 to 4 h, 1 ml of 3E5 medium was addedto each well, and incubation at theappropriate temperature wasallowed toproceed for 2days. Next, the medium was removed, and 3E medium lacking serum but containing 1 gCi of
["4C]thymidine per ml wasadded, and the cultures
wereincubated attheappropriate temperature over-night. After salt precipitation of sodium dodecyl sulfate-extracted material (5), supernatant fractions wererecoveredaftercentrifugation, andsamples were precipitated with 10% trichloroacetic acid and counted in a Mark II Nuclear-Chicago scintillation counter.
Distinction between the A and D mutants was based upon the fact that Amutant-infected cultures incubatedat 33C for 2 to 3days and then shiftedto 40C, rapidly lose their ability to synthesize viral DNA (2). Therefore, to distinguish the progeny, duplicate cultures were infected with a fewdropsof progeny virus asabove, but both cultures were incu-batedfor 3 daysatthepermissive temperature. At 3 days, one of the cultures was radioactively labeled
with ["4C]thymidineasabove, the otherwasshiftedto
40Cfor 5 h prior to the addition ofthe radioactive thymidine. Incubation was again allowedtoproceed
128 J.VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
129
overnight at the appropriate temperature and o o o o
counted as above.
RESULTS
Tempeature sensitivity of viral particles.
We previously postulated that D mutants are o
defectiveinuncoatingatthe restrictive temper- x x x x
ature (9). This hypothesis suggests that D virions harbor a defective virionprotein which
in turnmight destabilizethevirions at elevated *
temperatures. Representative strains ofeach of o o o
the classes of viral mutants were therefore x x x x
tested for their ability to withstand thermal m C4
inactivation for 3 hours at 50C in buffered
solution (see above). However, D mutant
vi-rions, like wild-type virions and A mutant | x x
virions, were notinactivated underthesecondi- e gD ue
tions (Table 1). m _ c'
-Virions of the late mutant classes were
in-cluded in this study todeterminewhetherany
correlation might exist between the thermal x x x x
lability of the mutants and their ability to LO c4
complement, i.e., to determine whether BC Q 4 _
mutants were
particularly
sensitive to heat.After 3 h at 50C, B201 was not inactivated,
ii
o o> >whereas the titers ofB204 and B8dropped35- x x x x
and 257-fold, respectively; C219 did not de- X X X eee
crease in titer, whereas C244 exhibited a four- o @
fold loss of infectious units; and BC230 de-
ie
creased10-foldintiter, whereasBC245dropped e o
1,000-fold. Thus, no simple correlation can be x x x x
drawn between the heat
sensitivity
of the late ae,-4
mutant virionsand theirabilitytocomplement. Virions of BC230are inactivated only 10-fold
under conditions where B204 and B8 (both of
which can complement C mutants) are inacti- x x x x
vated35- to250-fold. _
Theseresults areentirelyconsistent with the
notion that the Band Ccistronsencodeone or oo o o
moreviralcapsidproteins and suggest that the 2 _ _ _
failureofBCmutants tocomplementwith Bor . . .x
C mutants is not
simply
the result of theirhypersensitivity toheat inactivation. The
fail-ure to observe heat inactivation of A and D
mutant virionsdoes not, of course, exclude the x x x x
possibility that the proteins encoded by these e
cistrons arealso virion components.
Kinetics of virusyieldintemperatureshift o o o o
experiments. It has previously been argued | x x
bothon the basis of virion inactivationbyheat a X X 00
(as above) and the kinetics of virus yield in __
temperatureshift experiments, that Bmutants
are defective in a capsid-protein component X o o0
(10). To conduct asimilar analysis with the D
3___
___mutants, we have performed experiments
(based on those of Tegtmeyer [10]), involving 8 _ cqc
incubation of mutant-virion-infected cultures
on November 10, 2019 by guest
http://jvi.asm.org/
CHOU AND MARTIN
for varying lengths of time at the permissive temperature, followed by incubation at the restrictive temperature. After 4 days (total incubation time), the cultures were frozen, thawed and titered for virus at the permissive temperature. The profiles for early mutants would be expected to risebefore the profiles of late mutants, assuming, of course, that the shifting ofan infected culturetothe restrictive temperature immediately shuts off further vi-rionsynthesisand/or assembly and thatvirions formed at the permissive temperature remain viable - assumptions which for many reasons
neednotbe valid. Nonetheless, Tegtmeyerand Ozer (11) and Tegtmeyer (10) found that B4 and Bll virionyields increasedinparallel wvith, but were delayed by approximately 12 hfrom, theyields of three A group mutants.
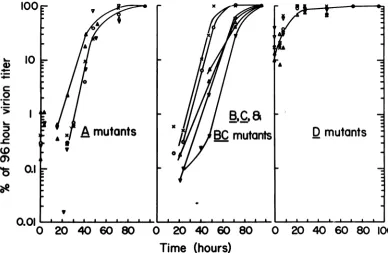
The results oftemperature-shift kinetic ex-periments with the mutants isolated in our laboratory are not entirely consistent with the previous reports (10, 11). All oftheA mutants with the exception of A276 display similar profiles in these experiments (Fig. 1). We
can-notexplain the morerapidappearance of A276 virions except to assume either that A276 is somewhat "leakier" than the otherA mutants and hence is not entirely "shutoff' when the culturesarechangedtothe restrictive tempera-ture, orthat the otherA mutants aresomewhat defective even at the permissive temperature. All of our previous data would tend to argue against the former explanation; first, because thedifferenceinplaque-formingunits (PFU) of A276atthe permissive and restrictive tempera-tures is of the order of 106 (3), and secondly, because DNA synthesisappears to ceasewithin 1 h at the restrictive temperature with this mutant (2). We therefore assume that the A mutants, with the exception ofA276, are par-tially defective evenat 33C.
The kinetic studies of virus yield with the late mutants generate afamilyof curvesrather than auniqueprofile(Fig.1,middle panel). Thiswas not unexpected since suchmutantsmight have different thermal stabilities even at 40C,orbe
differently affected in the synthesis, folding,
turnover, or assembly of capsid protein.
Mu-A mutants
I
20 40 60 80
Time
(hours)
FIG. 1. Kineticsof virus appearanceon temperatureshifttotherestrictive temperatureinfreshmedium. After2hofabsorptionat33C the unadsorbed viruswereremovedbyaspiration,andfresh3E5 mediumwas
addedtoeach culture. Thecultureswereshiftedto40Catthetimesindicated,and all cultureswerefrozenand thawedat96h. The titersarepresentedas apercentageofthe titerobtainedafter96hofincubationat33C. Symbols, left-handpanel:A,A207; V,A209; x,A239;0, A241; T,A255andA,A276.Middlepanel: 0, B201; V,B204;A,BC230; V, BC245; x, C219 andA,C240.Right-hand panel:0,D101;A, D202;V,D222;A,D238; V,D236 and x, D270.
100
L. 10
a)
0I
0 c
(00)
't0.I
D mutants
130 J. VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:4.504.60.448.339.592.2]tants C219 and B201 reach 10% of their final yield by approximately 42 h atthe permissive temperature prior to the temperature shift, whereas mutants BC245, BC230, and C240 do not attain this percentage of their final yield untilapproximately55h. B204 requires67hat the permissive temperaturebeforetemperature shift toreach 10% ofthe yield foundat 96h at the permissive temperature. Whatever the cause(s) forthe family of curves generated by the latemutantsinthese kinetic experiments, it isclear thatvirions of theB, C, and BCmutants
always appear at, or after the A mutants
(compare left-hand and middle panels,Fig.1).
Similar kinetic experiments performed with
the Dmutantsyieldastrikingly differentprofile (Fig. 1). When the virionsareadsorbed (asinall ofthese experiments) for 2h at the permissive temperature and immediately shifted to the restrictive temperature, the yields after 4 days ofincubation are already over 10%of thefinal yields found when the entire incubation is carriedout atthepermissivetemperature! Sim-ilar results were obtained whether the experi-ment was performedatMOIs of2 to5PFU/cell
oratMOIsaslowas0.1PFU/cell.Yet, the titers
ofseveral ofthese mutantsdifferbymorethan 105 at 33 C versus 40 C by plaque assay (3). Controls inwhich theadsorption aswell asthe incubation was carried out at 40 C gave titers that were 10-fold less than those observed at time 0. By contrast, when the same stocks of viruswereusedtomeasurethe induction of host and viral DNAsynthesis,noviral DNA synthe-sis was observed by 24 h at the restrictive temperature,and theinduction of the host DNA
synthesis wasreduced (2).
One notable difference between the experi-ments ofFig. 1 and those involving the induc-tion ofviral DNA synthesis (2) was that after
adsorption, fresh medium was added in these
experiments, whereas serum-depleted medium was used for the latter. We therefore reex-amined the kinetics ofvirion appearance with
depletedmedium.
The kinetic profiles of the D mutants are
markedly affected by carrying out the
experi-ment in depleted medium (Fig. 2). Indepleted
medium, the titer obtained for the sample
shifted to the restrictive temperature
immedi-ately afteradsorptionwasthesame asthe titer
obtained from thecontrol in which the adsorp-tionwas carried out atthe restrictive tempera-ture, andthetiters ofbothsampleswere atleast 100-fold less than when the D mutants were allowed toincubatefor 4daysatthe permissive temperature. Furthermore, itnow took 14 h at
L..
*-1-C 0
L..
0
(0
0)
'l
_1o-10
I0
0.I
0.01
0
20
40 60
80
Time
(hours)
100
FIG. 2. Kinetics ofvirus appearance on tempera-ture shift to the restrictive temperature in depleted medium. The detailsoftheexperimentare asinFig. 1, with the exception that depleted medium was employed.Symbols: 0,D202; x,D270;V, A209 and A,B204.
thepermissivetemperaturepriortothe shift for D202 to reach 10% of the yield observed when the entireincubation wasallowedtoproceedat thepermissivetemperature.The10%markwas only slightly delayed with the other mutants (compareFig. 1).These results with the virions corroborateourpreviousobservation thatby20 h of incubationat the permissive temperature indepleted medium, D mutantinfections have reached a stage such that subsequent incuba-tion attherestrictivetemperature allows DNA replicationtoproceed unabated (2).
Theaboveresults indicate that theexpression of normal D function is required during the initialstages of infection.They donothowever,
distinguish between the possibilitiesthat (i) D
encodesadiffusible proteinsynthesizedearlyin infection;(ii) Dencodesanondiffusibleprotein *131
on November 10, 2019 by guest
http://jvi.asm.org/
[image:5.504.265.450.61.383.2]CHOUAND MARTIN
synthesized earlyininfection; and(iii)late in in-fection D is translated into a structural com-*ponent of thevirion andthattherelease of this protein fromviral DNAuponsubsequent
infec-tion isrequired for the expression of viral func-tions. Possibility (i), seemingly eliminated by the observation that D mutants did not
com-plement after3 days of co-infection with other temperature-sensitive mutants, must again be entertained because Dmutantsdocomplement
onprolonged incubation. Inthiscase, normal D
product shouldact in trans. In the latter cases
(ii) and (iii), complementation experiments at the restrictive temperature between wild type and the D mutantsshould indicate that D
mu-tants are cis acting, i.e., the progeny should
be predominantly of the wildtype.
Products of complementation with
wild-type andtemperature-sensitive mutants. To
determine if Dproductwerecis acting, cultures
were co-infected with wild-type and mutant virions. Theprogenyofsuch co-infections were
plated at the permissive temperature, and plaqueswere pickedatrandom,suspended in2 ml ofphosphate-buffered saline and then tested fortheir ability to grow at the restrictive tem-perature.
Dmutantsarecis acting (Table 2). Compari-sonofthe ratios ofmutant towild-type particles
inthe inputversusprogenyviruspools (Table2,
lastcolumn) indicatedapreferential replication
ofwild-type particlesoverthe Dmutant parti-clesuponco-infectionattherestrictive temper-ature. Inthecontrol experiments withA,B, BC and C mutants, wild-type progeny
approx-imated the input ratio ranging from50% (B204
x wild type) to a sevenfold increase (A209 x
wild type). However, the same ratio was
in-creased 17- and 83-fold when D mutants were
employed. The statistical significance of the
precise increase is less with the Dmutantsthan with the controls (i.e., the ratio might possibly be much higher), since 25 to 50%o of all the
progenyinthe A, B, BC, and C mutant experi-ments were mutant type, whereas only 3of 95
D202 x wild-type progeny and2 of75D270 x
wild-type progeny were of the mutant type. Furthermore, the final yields of the Dmutants
among the progeny (approximately 3% of the
total PFU, or 6 x 106 PFU) were very closeto the inputs for these mutants. Thus, the data suggest that Dmutant replication proceeds, if atall, ata reducedrate upon co-infection with
wild type. Other classes of mutants behave normally upon co-infection with wild type at therestrictive temperature, i.e., they are
com-plemented by wild type and appear at nearly theinput ratio.
Products ofdelayed complementation. The aboveresultssuggestthat Dfunctioncannotbe supplied in trans by a co-infecting wild-type
virus particle. Consequently, one would again
expect D mutants tobe noncomplementing as
they are in the usual assay with CV-1 cells
whereco-infectionis allowedtoproceed for only 3days (3). To determineifthe hostcell hadany
effect on the complementation, we repeated
these experiments with primary AGMK cells (Table 3) andBSC-1 cellswhich donot exhibit stimulation of host DNA synthesis upon
infec-tionby SV40(Table 4),andagain observed that Dmutantsfailedtocomplementothermutants
upon co-infection for 3 days at the restrictive temperature.
Wewerepreviously unabletoaccountfor the complementation observedwith the Dmutants when co-infection wasallowed to proceed for 1 to 2 weeks. Asexplained more fully below,the
observation that Dmutants are cisactingupon
co-infection with wild-type virions led to the
TABLE 2. Productsofcomplementationbetweentemperature-sensitivemutantsofSV40and wild typeatthe restrictive temperature
ParentalVirionsa Progenyvirions
Mutant strain Input(I) No. Progeny (P) RatioI/ratioP
ratioc examined ratioc
A209 2.8x105 1.8 1.2x 107 47 0.27 6.7
A276 2 x105 1.3 1.4x 107 48 0.55 2.4
B204 105 0.63 107 40 0.5 1.3
BC230 3 x10' 1.9 107 29 0.93 2.0
C244 10' 0.63 107 46 0.24 2.6
D202 8x104 0.5 2.4x107 95 0.03 17.
D270 4x 105 2.5 2.2x 107 74 0.03 83.
aEach culture contained approximately 105 cells and received1.6 x
10'
wild-typevirions. bInfectingmutant.cMutant virions towildtype.
132 J.VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
TAm 3. Vinuyields after co-infection ofAGMK cells for 3 days at40 C
PFU/ml Mutants
A209 B201 C219 D202
A209 32 2.4x10 4x 104 200 (185)a (17) (1.8) B201 1.3 x 10' 8x104 1.6 x10'
(22) (1.2)
C219 2.4x10' 1.6x103
(0.7)
D202 80
aNumbers in parenthesis represent the titer di-vided by the sum of the titers of the two parental strains alone.
TABz 4.VirusYields after co-infection ofBSCI cells for3daysat 40C
PFU/ml Mutants
A209 B201 C219 D202
A209 40 7.4x 10' 10' 2x102
(460)a (17) (0.8) B201 1.6x 10' 1.4x10' 3.6x 10'
(18) (2)
C219 6x10' 7x103
(1.1)
D202 2.4x102
aNumbersinparenthesisasinTable 3.
hypothesis that delayed complementation was
the resultofa rareproductiveinfection followed
by phenotypic mixingofthe progeny particles.
A prediction of this hypothesis is that the
progeny ofdelayed complementationbetweena D mutant and any other co-infecting mutant virion should consist predominantly ofD
mu-tant particles - the inverse ofthe result
ob-tained when D mutants co-infect wild-type
in-fectedcultures.
Cultures were co-infected with variousA, B,
orCmutants atMOIsof1to2and D202at an
MOI of approximately 0.2 to 0.5.
(This
wasnecessary since at higher MOIs ofD202, only
D202 progeny wereobserved.) Afterincubation
for 13 days at the restrictive temperature, the
cultureswere frozen, thawed and titeredatthe
permissive temperature. Individual plaques
were picked, and stocks were prepared from
thematthepermissivetemperature. The
prog-eny viral types were then distinguished as
follows; all were testedbythe agar-slant
com-plementation technique previously described
(3). In addition, the D mutants were distin-guishedfrom B and C mutantsbytheirinability
tosynthesizeviral DNAafterincubation atthe
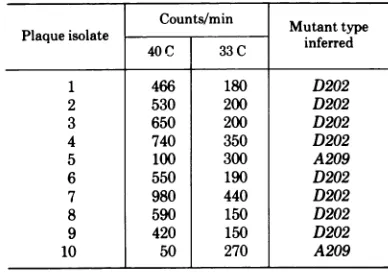
restrictive temperature. The data on 20 of 90 such analyses are presented in Table 5. D mutants were distinguished from A mutants by their ability to synthesize viral DNA upon shifting to the restrictive temperature after 3
days ofincubation at the permissive
tempera-ture (data from the first 10 of 43 progeny
examined in Table6).
The progeny resulting from delayed com-plementation between D mutants and other mutant types are predominantlyof the D class
(Table 7). When combinations ofA, B, and C
mutants were used toco-infect a monolayer of
CV-1 cells at the restrictive temperature, the progeny mutant yields reflected the parental input. This isdemonstrated bythe fact that the ratios ofparentaltoprogeny mutant types was
approximately unity(Table 7). This ratio,
how-ever, varied between3.5and 22inexperiments involving D202, indicating a preponderance of D mutants among the progeny.
DISCUSSION
Examination of the thermal lability of the virions of various temperature-sensitive SV40 mutants revealed that members of the late groups,B, C, and BC,areunstable. No
correla-tion between thermallability and
complemen-tation behavior was observed among the late mutants. The BC mutants do not therefore
TABLE 5. Analysisof representative plaque isolates from the D202 x C219 or D202 x B204
complementationexperiments D202 xC219' D202 x B204a Plaque
Counts/min
MutantCounts/min
Mutantisolate Mtn
type type
40C 33C inferred 40C 33c inferred 1 0' 520 D202 2,150 780 B204 2 400 700 C219 2,120 720 B204 3 0 410 D202 2,550 910 B204 4 29 430 D202 1,720 420 B204
5 0 540 D202 45 300 D202
6 0 175 D202 2,050 750 B204
7 250 820 C219 81 400 D202
8 0 390 D202 1,880 640 B204 9 41 460 D202 2,310 760 B204
10 0 310 D202 63 210 D202
aDataonthe first10of46progeny exami-ned in the D202 x C219 cross and the first 10 of 44 progeny examined in the D202 x B204 cross.
bNumbers represent the counts per minute in the supernatantfractions after extraction and salt precip-itation by the method of Hirt (5). The counts/min of mock-infected controls have been subtracted. They were192count/min at 40 C and 144count/min at 33 C.
15, 133
on November 10, 2019 by guest
http://jvi.asm.org/
[image:7.504.54.246.72.202.2]CHOUAND MARTIN represent a class of mutants which are
exqui-sitelysensitive to thermal inactivation. A and D mutants were not inactivated underthe condi-tionsused.Whereas these results areconsistent with the late mutants corresponding to the majorcapsid protein(s),they donotexclude the possibility that the proteins encoded by the A and Dmutants are also structural components of the virions. Indeed, recent mapping data suggest that D mutants map in the lateregionof the genome (6; C.-J. Lai and D. Nathans, in TABLE 6. Analysis of representative plaqueisolates from the D202 x A209complementation experimenta
Counts/min
Plaqueisolate Mutanttype
40 C 33C inferred
1 466 180 D202
2 530 200 D202
3 650 200 D202
4 740 350 D202
5 100 300 A209
6 550 190 D202
7 980 440 D202
8 590 150 D202
9 420 150 D202
10 50 270 A209
aCounts per minute less mock-infected controlsare
presented as inTable 5.Themock-infected controls had 82counts/minat 40C and84counts/minat 33 C.
Thecountsper minute forstrainD202at40Cand33 Cwere800and360,respectively,and for strainA209 were 70and 390counts/min, respectively.
press;T. E. Shenk, C. Rhodes, P. W. J. Rigby,
and P. Berg, in press).
The kinetic experiments in which mutant-infected cultures were incubated for varying
lengths of time at the permissive temperature before shift to the restrictive temperature dis-tinguish among the early mutants, in addition todistinguishingtheearlymutantsfromlate. D function was required only during the earliest
stagesof virion infection.
Interestingly, serum stimulation seems to partially overcome the defect exhibited by D mutants. Theprofiles of the virion yields in the kineticexperimentsweredisplacedtowardlater
timeswhendepleted mediumwasused in place
of fresh medium.Possibly,someserum-induced
proteaseof the host isnecessaryforthe uncoat-ing of virions, and D mutants are altered in
their susceptibility to the protease. Whatever the mechanism by which fresh medium
over-comes the defect exhibited by D mutants, we
wishtoemphasize that the properterminology
todescribe this phenomenon is "suppression," notleakiness. Thedifference in the titer of D202 at 33C and 40C is greater than 7 x 105 (3).
This means that when a monolayer of 3 x 106
monkeycellswasinfectedatMOI = 1,lessthan
five plaques wereobserved (3), although
statis-tically 10' cells were infected with 5 or more
PFU.The fact that after4daysin fresh medium the viral yield at 40 C was 10% of that at the permissive temperature means that fresh
me-dium can suppress the D mutations.
Suppres-TABLE 7. Productsofcomplementation betweendifferent mutantstrainsof SV40
Parental virions Progenyvirions
Mutant strain 1xMutantstrain2
Progen2
Input(l)
Total NO single X WCInfecting Infecting ratio PF plaques ratiob
Strain PFUa Strain PFU examined
A209 7 x104 D202 2x104 3.5 6x 105 43 0.39 9
C219 1.25 x106 D202 2x 104 6.3 106 46 0.28 22
B204 106 D202 2x 104 5.0 3 x 106 44 1.44 3.5
A209 7 x104 B204 10 0.7 4 x 106 50 0.47 1.5
A209 7x 104 C219 1.25 x 105 0.56 4 x 105 59 0.84 0.67
B204 105 C219 1.25x 105] 0.8 6 x 106 31 2.9 0.3
aThe titers of all of theparental stocks with the exceptionofC219 were checked atthe same time.
bMutant1/mutant2.
cAnotherwayofpresenting the datainthis column would be to point out that with the A mutant as strain 1
and the BorCmutantasstrain2,theI/Pratiosare1.5and0.67(average = 1.1)ascomparedwith avalue of 9 with the Dmutant asstrain 2.Similarly,with theB mutant as strain1and the A or C mutant asstrain 2, the ratios are0.67(1/1.5)and0.3(average =0.49), respectively,ascompared with a value of 3.5 with theD mutant as strain 2. Finally, with the Cmutant as strain 1 and theA orB mutant as strain 2, the I/P ratios are 1.5 (1/0.67)and3.3(1/.3)(average = 2.4)versus aratio of 22with the D mutant as strain 2. Combining the data in thisway, the averageI/Pratio forthe combinations excluding D202is 1.3[(2.4 +1.1 + 0.5)/3],andthe average including D202is 11.5 [(9 + 22 + 3.5)13],or adifference of ninefold.
134 J. VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:8.504.57.251.215.352.2]sion of certain phage mutations by nutrient factors is welldocumented (1).
When cultures were co-infected with D mu-tants and wild type under restricting condi-tions, the progeny were predominantly wild type, suggesting that the D function cannot be complementedin trans.The cisaction ofthe D mutants is most easily explained byone of two mechanisms: either (i) the protein encoded by
the D cistron is a virion component introduced with the viral DNAupon infectionand serving to prevent the expression ofall viral functions until released from the genome; or (ii) the D cistron is transcribed and translated from the genome soon after infection, and the protein synthesized remains associated with the paren-tal DNA. Whereas bacterial models for both mechanisms exist (7; B. Hoffman and M. Le-vine, personal communication), the former seems morelikelyineukaryotic cells where itis presumed that viral transcription and transla-tion are compartmentalized inthe nucleus and cytoplasm, respectively. We have been unable to find anyother plausible explanation forthe cis behavior of the D mutants. Whether the product of the D mutant were required in stoichiometricorcatalyticamounts,the ratio of D mutant to wild-type progeny should remain the same asthe parental ratio, provided the D product were a diffusible substance. While we cannoteliminate the theoreticalpossibilitythat upon co-infection wild-typevirions exclude the
adsorptionof D virions(butnotA,B,etc., even
though D mutants adsorb normally by
them-selves [9]), orthat the DNA of the D mutants contain a temperature-sensitive site, we find these explanationsunlikely.
Since the D cistron might encode a virion component (9) and possibly even one
synthe-sized late in infection (6), we propose the
followingasthebasisfordelayed
complementa-tion: ifDmutants are veryslightly leakysothat
perhaps 1in 10'to106cells infectedat anMOI
of 2 leads to a single round of infection, a low level of progeny D mutant virions will be re-leased into the medium. We have previously demonstrated thatby2 to 3dayspostinfection at the restrictive temperature, 80% of all cells which have been infected witha Dmutantlose their ability to lead to a productive infection upon subsequent incubation at the permissive temperature (9). Thatis, virion-infected cells in which virion replication cannot occur as the result of atemperature-sensitive lesion tendto become "cured" attherestrictivetemperature. Thus, thenewD virionsreleased from the 1 in 10'to101 infected cellswill,withhigh
probabil-ity, encounter cured cells after their release. However,the probability of infection will again beonly 1 in 10'to10,and since the burst size is less than 10' virions per cell, even prolonged incubation will not lead to an extensive infec-tion. When the same event takes place during co-infection withanother mutant, however, the result will be quite different. Only in the rare cell in which the D mutantreplicates will any virus be produced. But because this cell is co-infected with a different mutant, of classX, both will replicate. However, the virions re-leased will be phenotypically mixed, i.e., some ofthe D mutantvirions will haveawild-typeD cistron protein provided bythe other tempera-ture-sensitive mutant. Since most ofthe host cells for the second round of replication are effectively cured, anumber of types of infection arepossible. Single infectionsby the normal D virion, the X virion, and the phenotypically mixed X virion, will all be unproductive. Even most double infections by both mutant types (those in which the D virion contains the defective D-encoded product) will be nonpro-ductive. However, those cells receiving a phe-notypically mixed D virion will be productive for a single round, and those cells receiving a phenotypically mixed D virion and the Xvirion will beproductive and providemore phenotypi-cally mixed D virions.
The net effect will be that "complementa-tion" will not occur until many rounds of replication have taken place.Inaddition, there will beanincreaseinthe ratio of D virionstoX virions but not to an overwhelming degree.
Thosecellsinfectedbyonlyasingle,
phenotypi-cally mixed D virion will go through a single
round of replication and will release normal D virions. But the normal D virions released will
not be capable of efficient reinfection. This
prediction was tested, and D mutants indeed
prevailindelayedcomplementation byabouta
factor of9 (Table 7).
We therefore conclude that delayed com-plementation is nottruecomplementation, but the combined effect of a very low level of leakiness and phenotypic mixing of theprogeny D mutants. Inherent in this conclusion is the assumption that the D-encoded protein is part ofthe normal virion and that the Dvirions are blockedinuncoatingattherestrictive tempera-ture.Whether theDproteinissynthesized early orlate after infection cannot be deduced from these experiments.
While theevidencesupportingourconclusion concerningthe nature ofdelayed complementa-tion is only circumstantial, the datapresented 135
on November 10, 2019 by guest
http://jvi.asm.org/
CHOU AND MARTIN
in conjunction with the otherknown properties ofD mutants (3, 4, 6,9) areentirelyconsistent
with the proposed hypothesis.
ACKNOWLEDGMENTS
We thank J. L. Rosner, M. Singer, R. Saral, and D. LeBlanc for theirhelpful criticisms of the manuscript and Sylvia Bailey for technical assistance.
LITERATURE CITED
1. Ames,G. F., and B. N. Ames. 1965. The multiplication of T4 rnI phage in E. coli K12 (A) in the presence of polyamines. Biochem. Biophys. Res. Commmn. 18:639-647.
2. Chou, J. Y., J. Avila, and R. G. Martin. 1974. Viral DNA synthesis in cells infected by temperature-sensitive
mutantsof simian virus 40. J. Virol.14:116-124. 3. Chou, J.Y., and R. G.Martin.1974.Complementation
analysis of simian virus 40 mutants. J. Virol. 13:1101-1109.
4. Chou, J.Y.,and R. G. Martin. 1974. DNAinfectivity and
theinduction of host DNAsynthesis with
temperature-sensitive mutants of simian virus 40. J. Virol. 15:145-150.
5. Hirt, B. 1967. Selective extraction of polyoma DNA from infectedmousecellcultures. J. Mol. Biol. 26:365-369. 6. Lai, C.-J., and D. Nathans. 1974. Mapping
temperature-sensitivemutantsofsimian virus 40:rescueofmutants
by fragments of viral DNA. Virology 60:466-475. 7. Lindahl, G. 1970. Bacteriophage P2: replication of the
chromosomerequiresaprotein whichactsonlyonthe
genomethat coded for it.Virology 42:522-533. 8. Robb, J. A., and R. G. Martin. 1970. Genetic analysis of
simian virus 40: description of microtitration and replica plating techniques for virus. Virology 41:751-760.
9. Robb, J. A., andR.G. Martin. 1972.Geneticanalysis of simianvirus 40.III.Characterization ofa
temperature-sensitivemutantblockedatanearlystageof produc-tive infection inmonkey cells. J. Virol. 9:956-968. 10. Tegtmeyer, P. 1972. Simian virus 40deoxyribonucleic
acidsynthesis: the viral replicon. J. Virol. 10:591-598.
11. Tegtmeyer, P., and H. L. Ozer. 1971. Temperature-sensi-tivemutantsof simianvirus40:infection ofpermissive cells. J. Virol8:516-524.
136 J. VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/