0022-538X/78/0028-0434$02.00/0
Copyright 0 1978 AmericanSocietyforMicrobiology Printed in U.S.A.
Binding
of
Tryptophanyl-tRNA
to
the Reverse
Transcriptase
of
Replication-Defective
Avian
Sarcoma Viruses
AMOS PANET,'* GILDAWEIL,' ANDROBERT R.FRIIS2
Department of Virology, TheHebrew University-HadassahMedicalSchool, Jerusalem, Israel,'and Institut
fur
VirologiederJustus-Liebig-Universitat,
Giessen,
WestGermany2
Received forpublication6 June 1978
The ability ofreverse transcriptasetobindto[3H]tryptophanyl-tRNA andto
functionas DNApolymerase wascompared for five temperature-sensitive
mu-tants of avian sarcoma virus. Both activities ofthe reverse transcriptase were
found to be heat labile in LA 335 andLA 336 as compared with the wild-type
parents. For theothermutantviruses, LA 338, LA 343, and LA 672,grownatthe permissivetemperature,thereversetranscriptasewasnearlyasheat stableasfor
thewild-type parents intermsof tRNA binding andDNA polymerase.LA 338, LA343, and LA 672 showed characteristic defects in theirreverse transcriptase
whenpropagatedatthenonpermissivetemperature;namely, tryptophanyl-tRNA binding and DNA polymerase activities were coordinately decreased in these
virions. The reduced enzymatic activities were not entirely due to an inactive
reversetranscriptasepresentin thevirions, however, but rather loweramountsof
enzymeprotein incorporated into the virions contributedtotheeffect, according
toassaysofreversetranscriptase antigen by radioimmune competition.
Three activities can be independently mea-sured withpurifiedreversetranscriptaseofavian sarcomaviruses(ASV), namely,DNA polymer-ase(1, 20), RNaseH(13),andtryptophan-tRNA binding (16). Bindingof theprimer tryptophan-tRNA isviewedas aninitiationfunction in pro-viral-DNAsynthesis, whereas DNA polymerase activity with the polyadenylate oligodeoxy-thymidylate [poly(A) oligo(dT)] template is consideredanelongationfunction (19, 23).
Severalreplication-defective ASV have been investigated for theirreversetranscriptase activ-ities(12, 24, 25). Early replication temperature-sensitive (ts)mutantsLA 335and LA 337 were shown to contain heat-labile reverse transcrip-tase for both DNA polymerase and RNase H, thus indicating that these enzymatic activities arecodedby theviral genome. RSV,a,a noncon-ditional defective mutant, has been shown to completelylack the reversetranscriptase protein (8, 15).
In thepresent study, we haveexamined the DNA-polymerase activity and the binding to tryptophanyl (trp)-tRNA of reverse transcrip-tase from five conditional ASV mutants, two mutantscarryinganearly replicationlesion and three mutants defective in a late replication function.
MATERIALS AND METHODS Materials.[3H]tryptophan,11Ci/mmol,and
[3H]-dTTP, 46 Ci/mmol, were obtained from the
Radi-ochemicalCentre,Amersham, England; Sephadex G-100 was a Pharmacia Fine Chemicals product. Rat serumspecificforpurified avian myeloblastosis virus
(AMV) reverse transcriptase was a kind gift from R. Nowinski. Theimmunoglobulin G (IgG) fraction was
purified asdescribed byWatson et al. (26). Chicken liver tRNAwasprepared according to Rogg et al. (18). Chickenliveraminoacyl-tRNAsynthetases were pur-ifiedby using the procedure of Nishimura and Wein-stein (14). AMV reverse transcriptase was purified fromsucrosegradient-banded virus as previously de-scribed(17).
Preparationof[3H]trp-tRNA.The reaction mix-ture(1ml)contained:20mMTris-hydrochloride, pH
7.5; 10mM MgCl2; 10mM KCl;2mM ATP;0.5mM CTP;2mMdithiothreitol;160t,Ciof[3H]tryptophan;
1 mg of chicken liver tRNA; and aminoacyl-tRNA synthetase preparation (260,ug). After30 min of in-cubationat37°C,sodiumacetatebuffer(pH5.0)was addedtoa 0.1Mconcentration; EDTAwasaddedto 25mM,and thetRNA wasextracted three times with an equal volume ofphenol saturatedwith 0.1 M so-diumacetate (pH5). The tRNAwasprecipitated by
adding2volumes ofethanol,and theprecipitatewas washed twice by resuspending in 66% ethanol-33% sodium acetate (0.1 M), pH 5. The final pelletwas dissolved in 10 mM sodium acetate (pH5)-0.1 mM EDTA.A maximumof0.4to0.5%ofthe crude chicken liver tRNA was routinely charged with [3H]trypto-phan.
DNApolymerase assay. Reaction mixtures (100
p1) contained: 50 mM Tris-hydrochloride, pH 8.3; 6 mM MgCl2; 10 mM dithiothreitol; 10 Lg of bovine serum albumin; 10 MM [3H]dTTP (specific activity, 450cpm/pmol); 0.5 ,g ofpoly(A) oligo(dT); and 20-434
on November 10, 2019 by guest
http://jvi.asm.org/
werefor 30minat37°C. Reactionswerestopped with
5% trichloroacetic acid, and radioactive precipitates
were collected on glass fiber filters. A unit is the
amountofenzymeneededtocatalyzethe incorpora-tion of1pmolof[3H]dTMPin 1minunder standard conditions.
Binding assay. Reaction mixtures (100 Ml)
con-tained: 50 mM Tris-hydrochloride, pH 7.5; 60 mM NaCl;6mM MgCI2; 10 Mgof bovine serumalbumin;
0.2%Triton X-100;5mMdithiothreitol; 16,000cpmof
[IHItrp-tRNA
(unfractionated) (1.3 pmolof [3H]tryp-tophan);andpurifiedvirusasindicated.After 10min ofincubationat22°C,themixtureswereloadedonto Sephadex G-100 columns (0.6 by22 cm) and eluted with buffer A (0.1 M potassium phosphate, pH 7.1; 10% glycerol; 0.1% Triton X-100; 10 mM mercapto-ethanol). Fractions of 0.18 ml were collected; 20-MLsamples were taken for assay of DNA polymerase
activity,and therestwerecounted for[3HItrp-tRNA
in toluene scintillation fluid containing30% Triton
X-100.
Cells and viruses. Chicken embryo cells were
preparedfromfertile whiteleghorn embryosobtained from Heisdorf and Nelson Farms, Redmond, Wash., and tested for the expression ofchick helperfactor according to the procedure of Friis et al. (5). Eagle Dulbecco-modified medium supplemented with 10% tryptosephosphate broth, 5% calfserum, and 1% di-methyl sulfoxidewasusedforvirusstockpreparation.
Chickenhelper factor-negative chicken embryo cells
wereinfectedwiththevarious virusesin thepresence
of 2 MgofPolybrene(Aldrich Chemicals) perml(21), andcultures were maintained withregular transfers
for 10to14daysuntil 30to40 dishes(90 mm)of each wild type or mutantvirus stockhad been prepared. Harvests of virus were then made, usually at 24-h intervals, but,asreportedbelow forts-672,
occasion-allyat 3-h intervals,from cultures maintained
sepa-rately at 35and 42°C. Cultures maintained at42°C
wereharvestedina warmroom on a warmtable held
at42°C. Pooled culture supematantswerefrozen at -70°Cuntilavolume of 3to5 literswasavailable,at whichtime viruswaspelletedinaSpincotype19rotor (19,000rpmfor 2h).All thevirusestested inthis work
were purifiedunder identical conditionsby gradient
purificationoncontinuous 20to50%sucrosegradients.
Biologicalcontrolswereperformedwithallvirus
prep-arationstodemonstrate thatthestocks exhibited the typical temperature-sensitive properties of the
mu-tantsand thecorrectsubgroup.
Thefollowing mutants obtained from the Prague strain (PR) ofRoussarcoma virus (RSV) havebeen used:tsLA335,subgroupC(10);tsLA338, subgroup C (27); ts LA 343, subgroupC (27); and tsLA 672, subgroup A (3). For control purposes, wild-typePR RSV,subgroup C (wt PR-C), has beenused. TS LA 336(formerlycalledts149)(6, 22)wasalsoinvestigated
and its parent, wild-type ASV strain Bratislava 77, subgroupC (wt B77), wasused for controlpurposes.
RESULTS
Binding of trp-tRNA to reverse
tran-scriptase usingdisruptedvirions. In
previ-ousstudies, the interaction of the primer
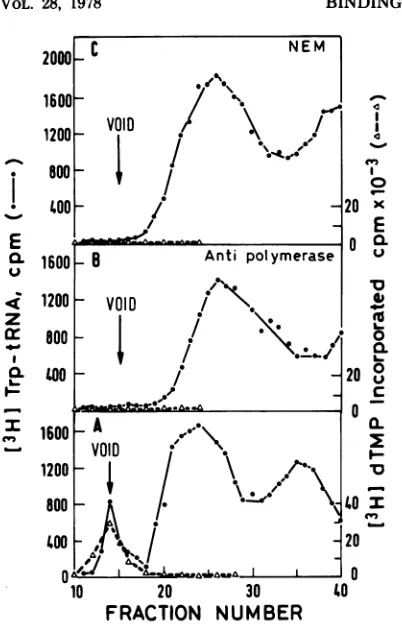
[32P]-tRNAtP and AMV reverse transcriptase has been demonstrated, using purifiedenzyme prep-arations (7, 16). To be able to measure tRNA complex formation with the enzyme of ASV temperature-sensitive mutants, we sought con-ditions for binding in crude virionextracts. Fig-ure 1demonstratesthat[3H]trp-tRNA bindsto reversetranscriptase in detergent-disrupted vir-ions aswellas to a purified AMV reverse tran-scriptasepreparation. Complexformationis ev-identby the comigration of virion DNA polym-erase and part of the [3H]trp-tRNA, with a sedimentation coefficient of 9Songlycerol gra-dients. Under the same conditions, unbound purifiedreversetranscriptase (not shown) hasa sedimentationcoefficient of 7.2S(150,000 molec-ular weight), similar to that of the protein marker IgG (16).
Toexclude the possibility that[1H]trp-tRNA
binds indetergent-disrupted virionstoproteins otherthan thepolymerase and formsacomplex withasedimentation coefficient similartothat of the reverse transcriptase, we investigated whether known inhibitors oftheviralDNA po-lymerase activity also inhibit tRNA complex formation in disrupted virions. In these experi-mentsbindingof [3H]trp-tRNAwasassayedby
gelchromatography onSephadex G-100, where theenzymecomplex migrates inthe voidvolume (fractions 13 to 15), while unbound [3H]trp-tRNAelutesasthe secondpeak (fractions20to 30)and free[3H]tryptophan,adeacylation prod-uctfromtRNA, is retardedtofractions30to 40 (Fig. 2). Preincubation of detergent-disrupted virions withmonospecific antibody, produced in rats against purified reverse transcriptase (26), orwith N-ethylmaleimide (NEM), apotent in-hibitor ofDNApolymerase and tRNA binding activities in purifiedenzyme preparations (15), abolished both activities in crudeextracts (Fig. 2).These resultssuggestthat of all viralproteins only thereversetranscriptase is able to forma specific and stable complex with trp-tRNA.
Inthiswork[3H]trp-tRNA rather than [32P]-tRNAtr"Pwasusedtoquantitate complex forma-tion withreversetranscriptase, andwefind sev-eral advantages in using the aminoacylated tRNAasasubstrate:(i)specific tagging of crude tRNA preparationwith [3H]tryptophan is easy and does notrequireuseof two-dimensionalgel electrophoresis, which is necessary for the puri-fication of
[32P]tRNA"rP
from a mixture of tRNA's; (ii) the isotopein[3H]trp-tRNA is sta-ble as compared with the in vivo labeled[32P]-tRNA, and specific activities of [3H]trp-tRNA
preparations are accurately assessed. The one
disadvantageinusing[3H]trp-tRNAisthe rela-tiveinstabilityof theaminoacylesterbond. We therefore ran glycerol gradient centrifugations
on November 10, 2019 by guest
http://jvi.asm.org/
436 PANET, WEIL, AND FRIIS
en
E
60 °
o
x 0
oE
o40
c--I
<a
20l
'a.4w
0 c
40
a
I-I
1
10
20
FRACTION
NUMBER
FIG. 1. Binding of trp-tRNA in disrupted virions. Bindingof [3H]trp-tRNAwasconductedasdescribed in
thetext,except50mMpotassiumphosphate buffer, pH6.0,wasusedinstead of Tris buffer. After incubation
for10minatroomtemperature,internal protein marker, humanIgG (0.6mg),wasadded,andthemixtures
werecentrifuged for 30 minat5,000xgtoremoveprecipitates. Theclearsupernatantswerelayeredon5-ml
glycerol gradients (5to25%, vol/vol) in 50 mMpotassium phosphate(pH6.0),0.1MKCI,0.1%TritonX-100, 10mM/-mercaptoethanol, and 20% ethylene glycol. Gradientswerecentrifuged for23hat49,000rpminan
SW50.1rotorandfractionated. DNA polymerase activityand trp-tRNA bindingwereanalyzedasdescribed
in thetext, andthelocation of IgG proteinmarkerwasdetermined by assaying the proteincontentinthe fractions after trichloroacetic acid precipitation,accordingtoLowryetal.(11). (A)wtPR-A virions,340pg;
(b)purified AMVreversetranscriptase,100enzymeunits.
and gel fitration columns under conditions
which minimized deacylation of aminoacyl-tRNA,namely, low-pH phosphate buffer.
In all experiments described in this
commu-nication, saturating amounts of [3H]trp-tRNA
were added, and thereforeonly 15% of [3H]trp-tRNA bound to the enzyme (Fig. 2A). Under
conditions of suchtRNA molar excess,any
re-duction of the enzyme-binding activity should
bedetectable,thusenablingustomeasure
bind-ing activityperagiven amount of virus. When
anexcessofvirus,rather thantRNA,wasused in the binding reactions, as much as 35% of
[3H]trp-tRNA bound to the enzyme in crude
virus extracts (results not shown). A similar percentage of binding has been previously
re-ported, using a purified preparation of
[32P]-tRNAtrP (16). No structural differencebetween bound and unbound fractions has as yet been
detected, and it could be that conformational
isomersoftRNAt'rPexplain the two
fractions.-trp-tRNAbindingin LA 335 and LA336 mutants. Temperature-sensitive mutants LA 335 and LA 336 have been shown to be early replication-defective mutants which contain
heat-labile reversetranscriptase (6, 10, 12, 25).
The enzyme in these mutants is defective for
both DNApolymeraseandRNase H(12,24,25). The results shown inTable 1demonstratethat
for both mutants, DNA polymerase activity, measured with poly(A) oligo(dT) template, is
heat labileascomparedtothewild-type viruses
RSV PR-C and ASV B77. Thus, in agreement with theresults of Vermaetal. (25), preincuba-tionat45°C resultedin50%inactivation ofthe
polymerase activity with detergent-disrupted virions of LA 335 and LA 336 after4and2min, respectively.
Since the specific binding of the primer tRNA11' represents yet another activity of the
enzymeprobably needed for initiation of DNA
synthesis onthe viral RNAtemplate, it wasof
interesttocomparethe heatlabilityof the
bind-ing site tothat of the polymerasesite. [3H]trp-tRNA binding activity was estimated, after
preincubation ofdisrupted virionsat45°Cfor 30
min, by gelfiltration of thecomplexformed.In
thecontrolreactions,reversetranscriptaseofwt
PR-C, LA 335, and LA 336 was shown to be
active in binding,as is evidentby the
co-chro-7
300
I 1
200
E
100
0. u
.ci
400
z
E
300
"
200
I
100
I"
n
J. VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:3.499.109.400.74.307.2]i
800
_
_
400_
20 x/
~~~~~E
E
-- -A-a0CXL
1600 -B
Anti polymerase
'<
1200-VOID;
z / 0
a
1.00
/O
20 °F o' ~~~~~~c
A-A
~~~~~0
a.
C 1600 A
A.
L
VOID
*
.
1200 ~
800 '.
401
400
,
-120O
0 ~ 0
10 20 30 tO
FRACTION NUMBER
FIG. 2. Inhibitionofreversetranscriptasebinding activityindisruptedvirions. VirionsofwtPR-A(340 pg)werepreincubated inbindingreactionmixtures,
with 0.1 mMdithiothreitol:(A)withoutadditions;(B)
with ratIgGmonospecificforAMVreverse transcrip-tase(12pg);(C)with 2 mMN-ethylmaleimide.After
20min, dithiothreitol(10mM) was addedtodestroy
excessNEM,followed byadditionof[3H]trp_tRNA
(16,000 cpm). The mixtures were chromatographed afteranadditional 10minonSephadex G-100 col-umnsasdescribedin thetext.
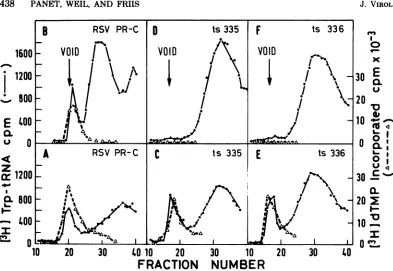
matography of[3H]trp-tRNAand the DNA po-lymerase activityonSephadexcolumns(Fig. 3A, C, and E). Preincubation at high temperature
hardlyaffected thewtPR-Cactivities,whereas both trp-tRNA binding and DNA polymerase
activitiesin mutants LA 335 and LA 336 were completelyinactivated (Fig. 3B,D,andF).
Heat sensitivityof DNApolymerase and
tRNA binding activities with LA 338, LA 343, and LA 672. The three temperature-sen-sitive mutants LA 338, LA 343, and LA 672 exhibit a commonproperty;all carrylate repli-cation defectswhichdetermine that virions pro-duced at the nonpermissive temperature are
noninfectious (3, 27). Other important differ-ences emerge between LA 338and LA 343,on
the one side, and LA 672on the other. LA338
and LA 343clearlycarry severalgeneticdefects
sensitive ASV DNApolymerases
Half-life (min) for DNA polymer-Virus aseactivity after preincubation at
45°C"
wtPR-C 80
LA 335 4
LA338/350C 40
LA338/42°C 60
wt B77 60
LA336 2
"Mixtures (100
pl)
contained 50 mMTris-hydro-chloride,pH 8.3,6mMMgCl2,10,ug of bovineserum albumin, 0.02% Triton X-100, 5 mMdithiothreitol, and virions asfollows: B77 and wt PR-C, 32 enzyme units; LA 336, 25.4 enzyme units; LA 338 grown at 35 or 42°C, 33 enzyme units; LA 335,30enzymeunits. The mixtures wereincubated at45°C, and 10-Mul samples
weretaken into ice. Residual DNApolymerase activity was assayed by adding 90,l of assay mixture as described in thetext.
(2, 9), oneof which affects transformation inde-pendent of virus replication. The other is re-sponsible for, in addition to the late defect in replication, an early defect, first described by Wyke and Linial (27) as a transient essential function for the establishment of transforma-tion. Recentexperiments with LA338 (Molling
and Friis, manuscript in preparation) indicate that this early defect, which affects replication, is indeed only a separate phenotype caused by a reversetranscriptase lesion, perhaps thesame whichdetermines the late defect. Hence,for LA 338, a reverse transcriptase lesion probably
causes both an early and a late defective phe-notype. LA 338harvestedat35°C also exhibited in ourexperiments asignificantly reduced
half-life compared withwtPR-C after incubation at
45°C. LA 672 apparently has only a single ge-netic defect,thataffectingreversetranscriptase (4). Mostinterestingwith LA672is the fact that thereversetranscriptasedefect resultsonlyina
late defective phenotype; hence, cells infected withstocksof LA672preparedatthepermissive
temperaturedonotexhibitany temperature
sen-sitivityin the initial establishment of infection. The noninfectious particles of these mutants havethesamestructuralpropertiesasinfectious
virus, and all themajorstructural proteinsare
present in normal amounts (4). We have studied twoproblems with these mutants. (i) Does the reversetranscriptasecontaina temperature-sen-sitivelesion which effects the initiationof DNA synthesisasmeasuredbythetRNAprimer bind-ingactivity,and (ii)is the reducedDNA
polym-eraseactivityin virionsproducedatthe
nonper-is.sive
temperature (42°C) due to an inactive enzymeor to areducedamountofactivereverse401%
.4
1
1
4
%-O
on November 10, 2019 by guest
http://jvi.asm.org/
[image:4.499.48.249.56.368.2] [image:4.499.80.445.59.371.2]438
1600
VOID
VOID
VOID
U~~~
I \,
I
I^1^^
IN
11200
1
3
0
EE
0
20
/
10
a
-~~
4L-O000CA.
A
RSV(PR-C
a-C
ts335E
ts336z
gx
1200
I
30
.c
I-
80-20
X
ff00sP-
/10.-0
oa
10
20
30
40
10
20
30
10
20
30
40
FRACTION
NUMBER
FIG. 3. Inactivation oftrp-tRNA binding activity in LA 335 and LA 336. (B, D, and F) Virions were
preincubatedinbindingmixtureathightemperature(450C)for30min, and[3HJtrp-tRNAwasthen added.
(A, C,andE)Bindingwasconductedwithoutpreincubationasdescribed in thetext. (AandB)wtPR-C(330 pug); (CandD)LA335(325pig);(EandF)LA 336(330
pg).
transcriptase assembled into virions?
The observation ofVerma etal. (25) that in vivo proviral DNA synthesis is reduced when infection with LA338is carriedoutatthe
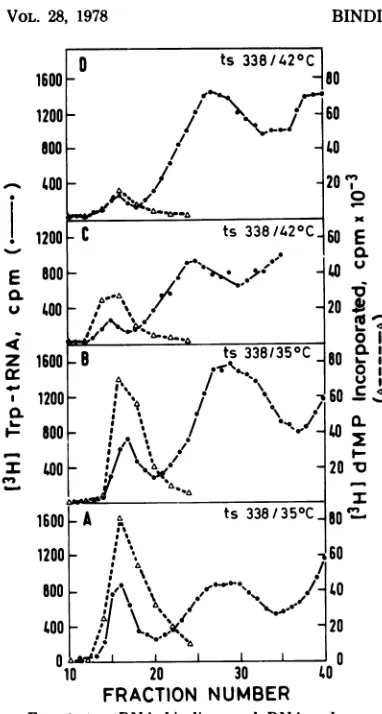
non-permissive temperature, even though in vitro DNA polymerase activity is heat stable, prompted us toinvestigate whether the initia-tion of DNAsynthesis,asmeasuredbythe trp-tRNAbindingactivity,istemperaturesensitive inthis virus. The results shown inFig.4indicate that thetRNAbindingactivityofreverse
tran-scriptase from LA 338virions, grown at either thepermissive(350C)orthenonpermissive tem-perature (420C),isnotthermolabile. Disrupted virionswere preincubated athigh temperature (45°C) or left at
40C
for 30 min, followed by addition of [3H]trp-tRNA and analysis by gelfiltration. The enzyme in LA 338grown at the permissive temperature andlater preincubated
at
450C
canbindasmuch[3H]trp-tRNAasthe same enzyme which was kept at40C
(Fig. 4A andB). Similarly, trp-tRNA binding to nonin-fectiousvirions,producedat420C,
is heatstable,and preincubation at 45°C for 30 min did not
affect thebinding activityof thatreverse
tran-scriptase (Fig. 4CandD). Preincubationof dis-rupted virions at 45°C resulted, however, in a
small decrease in the total amount of DNA
polymerase eluted from the Sephadex column (see alsoTable 1).
To investigatethe biochemical nature of the latereplication lesion in LA338, wequantitated the DNApolymerase and tRNAbinding activi-ties in virions madeatboth35and
420C.
Agiven amount of virionsproduced at350C
bind four-fold more trp-tRNA as compared with virions madeatthenonpermissive temperature(420C)
(Fig. 4A andC; Table2). Asimilarly enhanced (three-tofourfold) specificactivity of DNA po-lymeraseisnoticed in virionsproduced at350C
ascomparedwith those madeat420C.
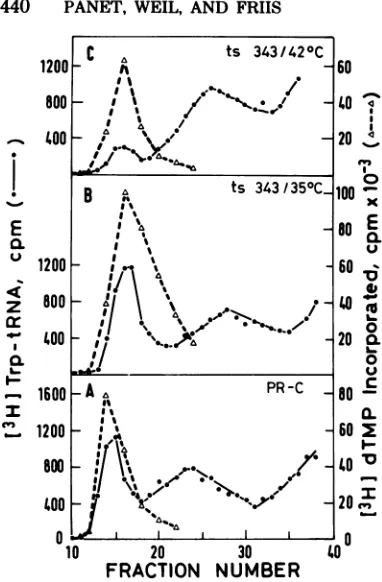
The total DNApolymeraseand[3H]trp-tRNAbinding ac-tivities eluted from the columns (Table2)were calculatedfromtheresults showninFig.4Aand Cbyintegratingtheareaunder the voidvolume peaks.LA 343 wasalsoanalyzedforitsreverse tran-scriptase properties (Fig. 5). The virions pro-ducedatthenonpermissivetemperature
(420C)
show a fourfold-lower specific activity for trp-tRNA binding compared with virionsgrown at350C,
and the DNApolymerase specific activityin the noninfectious virus was reduced by a factor of2.5.Wild-type PR-C producedateither
42 or
350C
areboth infectious and havespecific activities for tRNAbindingand DNAon November 10, 2019 by guest
http://jvi.asm.org/
[image:5.499.62.455.54.325.2]E
-i
z cr
-W
I
CL
IE
Q
I E
V
o0I
0 1
I
o I U I
O a.
P-10 20 30 40
FRACTION NUMBER
FIG. 4. trp-tRNA binding and DNA polymerase activities in LA 338. Binding reactions were
con-ductedasdescribed in thetext.(A)LA338,grownat 35°C (330pg), and(B) thesamevirions were
prein-cubated in thebindingmixturefor30min at45°C before[3Hltrp-tRNA wasadded. (C) LA 338grown
at42°C (330pg)and(D)thesamevirionswere
prein-cubated, for30 min at45°C, before ['HJtrp-tRNA
wasadded.
asesimilartothemutantvirus LA 343produced
at 350C (Fig. 5A). The results of these two
independentassaysfortRNAbinding and DNA polymerase suggest that the late
temperature-sensitive lesions in LA 338 and LA 343 affect the amounts ofactive reverse transcriptase in
virions producedatthe nonpermissive temper-ature.
LA672alsoappears tobe defective ina late
stageofreplication in the synthesisorassembly
ofafunctionalreversetranscriptase (4). We have
compared the tRNA bindingaswellastheDNA
polymerase activities in infectious virions
pro-duced at 350C and noninfectious virionsmade at420C. Suchastudy givesusinformationasto
whether the noninfectious virions of LA 672
activities and phenotypesofLA672,LA 338, and LA 343propagated at permissive and nonpermissive
temperatures
DNApo- [' fl
lymer- ]trp- Relative Ei-Virus ase" (en- binding'tRNA amt of ciencyof
realica-zyme (pmol) antigen'
tionrl
LA338/350C 16 0.64 0.36 1
LA338/420C 5 0.15 0.19 0.08
LA343/350C 23 0.94 0.5 1 LA343/420C 9 0.22 0.14 0.07 LA672/350C 4.0 0.55 0.45 1 LA672/420C 0.7 0.15 0.09 0.0025
aTotal DNApolymeraseactivityeluted from Seph-adex G-100columnswas calculated bysumnmingthe enzyme activity in the void volume(usuallyfractions 13 to 18inFig. 4, 5, and 6).
b[3H]trp-tRNA bound is the sum of radioactivity
co-chromatographedwith theenzyme in the void vol-ume(usuallyfractions 13 to 18). Virion protein input wasaround 330,ug (see legends to Fig. 4, 5, and 6).
'Relative amount of reverse transcriptaseantigen in virions wasderived from amount (micrograms) of viralprotein needed to restore 20 or 30% of the DNA polymerase activity in the immunoassay (Fig. 7). At 20 to30% of enzyme activity the competition immu-noassay is linear, and therefore the amount of
com-petingvirus needed torestore thisactivity is inversely linear to the virus content of reverse transcriptase
antigen.The amount ofantigeninwtPR-Aor PR-C wasadjustedto 1unit, and the relative amounts of ts mutants wereaccordingly corrected.
dEfficiency ofmutant replication was calculated from the titers of infectious virus released to the medium at 35 and 42°C. The focus assay wascarried
outunder standard conditions (2, 3).
contain enzyme protein which has lost DNA polymerase activity but still retains its tRNA
binding activity. The results illustrated in Fig.
6Aindicate thatinfectious virusproducedatthe
permissivetemperature (35°C) containsa high
tRNA binding activity. An equal amount
(mi-crograms of protein) of noninfectious virions
(produced at 42°C) binds threefold less trp-tRNA but exhibits a sixfold reduction in DNA
polymerase activity as compared with LA 672 producedat35°C(Fig.6; Table2). The standard LA 672virions usedinthisstudywereharvested every 24 h and then purified. To exclude the possibility that the virions produced at 42°C were inactivated due to the high temperature afterreleasefromcells,wealsoinvestigatedthe reverse transcriptaseinrapidly harvested (3h)
virus. A rapidly harvestedvirus grown at42°C
(Fig.6C)behavessimilarlytothe24-h-harvested virus (Fig. 6B), and its polymerase and tRNA binding activities are both reduced compared
on November 10, 2019 by guest
http://jvi.asm.org/
[image:6.499.46.237.50.407.2]I* >,/\.z,,
|*~
I B ,, ts
3431350C_
10E
1600 , PR-C _
80 E1200 . 60
-zSoo I 40V
0I- -Io
1600
A~
PRC 02.1200 -601
800 /\ 40~
400 j Y20c,
10 20 30 40
[image:7.499.63.254.56.347.2]FRACTION
NUMBERFIG. 5. trp-tRNA binding and DNA polymerase activities in LA 343. Binding of[3HJtrp-tRNA was carriedout asdescribed in thetext. (A)PR-C grown at420C(300pg);(B)LA343grownat35°C (325ug);
(C) LA 343 grown at420C (327 pg).
with virusgrownatthepermissivetemperature,
35°C (Fig.6A). Allof these results indicate that
qualitatively the lesion in LA 672 is similarto
the late temperature-sensitive function in LA 338and LA343.
Quantitationofreverse
transcriptase
an-tigen in
temperature-sensitive
ASVmu-tants.Thefmdingof reducedDNApolymerase
andtrp-tRNAbindingactivities in ASVmutants
LA 338, LA 343, and LA 672 grown at the nonpermissive temperature could beexplained
in either oftwoways: (i) packaging ofenzyme into virions is a temperature-sensitive process, and virions produced at 420C contain less
re-verse transcriptase protein;or (ii) enzyme syn-thesized in thecellsathigh temperatureis de-fective, and,as aresult, virionscontain normal amountsofenzymeproteinwithlowenzymatic activities. To differentiate between these two
possibilities, wemeasured the relative amounts
of reverse transcriptase antigen in virions by using a competition immunoassay (15).
Deter-gent-disrupted virions are reacted with NEM,
which inhibits in a nonreversible fashion the DNApolymeraseactivitybutdoesnotaffect the
0
E
0.
z cr
-W
Q
L-: U
I
IC
v-x
E
a
u
-0 .0. 0 CL
a-~0
I 10
10 20 30 40
FRACTION NUMBER
FIG. 6. trp-tRNA binding and DNA polymerase activities in LA 672. Conditionsfor binding and DNA polymeraseactivityaredescribed in thetext.(A) LA
672,grownat35°C,harvestedat24-h intervals(345
pg); (B) LA 672,grown at42°C, harvested at 24-h intervals (345pg); (C) LA 672,grownat42°C, har-vestedevery3 h(360 pg).
antigenic properties of thereversetranscriptase,
andexcessNEMisdestroyed bydithiothreitol.
Increasingamountsof theNEM-treatedvirions
arethen reacted withalimitedamountof
anti-reverse transcriptase IgG. The ability of the
mutantvirusestoabsorb the monospecific IgG
canthenbeestimated by adding purified active reversetranscriptase toreactwith the free IgG, and finally the residual reverse transcriptase
activity is estimated with poly(A) oligo(dT) as
template primer for DNA synthesis (Fig. 7). Virions whichcontainalargeamountofreverse
transcriptase antigen will absorbmore
antibod-ies, and as a result less ofthe purifiedreverse
transcriptase willbeinactivated inthe final
re-action. The neutralization inhibition method used hereprovedtobe asaccurateasthe
stan-dard competition radioimmunoassay technique
00
_
10~~~~~~~~
D0 _ .I \
."e
n
/
I)o-
0A:
001on November 10, 2019 by guest
http://jvi.asm.org/
[image:7.499.267.458.73.396.2]C=
A
BC
P-
C.n~~~~~~~~~~~~~~~~P-_=
80.
.44 60 -
.0.o0*"
or I.- O de00
uJ0
-A:
40200
0
20o-
0
-- //20
0 .
-37°C,
thetueswee6rasf72/o4°C
20~~~~~~~~~~~~~~~~~~~~~~~~~.
ain mixure A4,uI cotinn 1mMTi-yrclid(p8.0)10 mM KCI, . go bvneu
ts
338/
420C
.
al i pe
m0
20
40
0
20
40
0
20
4t
[image:8.499.53.443.58.236.2]RSV-NEM
(pg
protein)
FIG. 7. Neutralization inhibitionofDNApolymerasein RSVtsmutants. The reactionswereconductedin
foursteps. (i)DNApolymeraseactivity in virionswas inactivated by incubationwith 2 mMNEM, 50mM
Tris-hydrochloride (pH7.5), 1.0 mgofbovine serumalbuminperml, and1.0% TritonX-100.After15mm at 37"C, the tubes were transferred to40C and dithiothreitol (7mM) was addedto destroyexcessNEM. (ii) Increasingamountsofvirus-NEMwerereactedfor20minat370Cwithanti-reversetranscriptaseIgG (3 pg)
in mixtures (45 gil) containing 10mM Tris-hydrochloride (pH 8.0), ffXJ mMKCI, 0.5 mg ofbovine serum albumin per ml, 10%glycerol, and 0.1% Triton X-100. (iii) ActivepurifiedAMvVreverse transcriptase, 1.5
units/S
[LI,
was added, and incubation was continuedforanother 20 min at370C. (iv) The residual DNApolymerase activitywasdeterminedby addingamixture(50Md) ofall thecomponentsneededfor assaying
DNApolymerase activity (see text), andthe incubation time was20min. Under the above conditions, the
enzymaticactivitywaslinearforatleast 40min. Thepercentageofresidual DNA polymerase activity was calculatedbycomparisonwith control reactions in duplicate tubes that contained no immune IgG (taken as 100%).
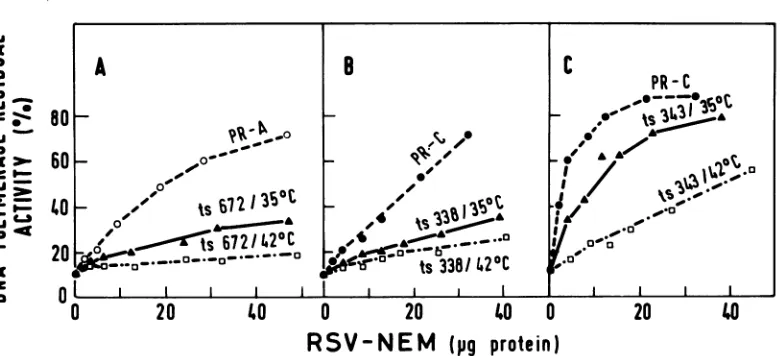
for reverse transcriptase (15). Thus, torestore 20% of theprobe DNA polymerase activity, 55
,ug
of LA 672 virions grown at 42°C, 11 ,tg of virionsgrown at350C,and5,ug
ofwtPR-A virus were needed (Fig. 7A). These results indicate that LA672virusgrownat420C contains about fivefold less enzyme protein as compared with thesamevirusgrownatthepermissive temper-ature(350C).LA338and LA343 mutantsshow a similarlack, and virionsproducedatthenon-permissivetemperaturecontaintwo-tofourfold lessreversetranscriptaseascompared with virus madeat35°Candsomesixfold lessenzymethan thewtPR-C (Fig. 7BandC;Table2).
All of these results indicate that the three conditional mutantsanalyzed, LA 672, LA 338, andLA 343,contain reducedamountsofreverse
transcriptase protein per total virus proteins
whenharvested atthenonpermissive tempera-ture.Itisinterestingtonote,however,thateven
the infectious virus mutants produced at the permissivetemperaturealways contain less
en-zyme protein than the wild-type parents. Whereas the reduced incorporation of reverse
transcriptaseinto virions canexplainlower DNA polymerase and trp-tRNA binding for LA 338 and LA 343 grown at
420C,
LA 672 grown at420C
probablyincorporatessignificantamountsofdamagedenzyme,since the trp-tRNA binding andDNApolymerase activities for these virions grownat
420C
are notparallel.DISCUSSION
Inthepresentstudywe describe the interac-tion betweenthe tRNAprimerfor DNA
synthe-sisand thereverse transcriptase of five temper-ature-sensitive mutants of ASV. The finding that trp-tRNA binding activityin LA 335 and LA336 is thermolabile provides the best proof that this function is virus coded like the DNA polymerase and the RNaseHactivities (24, 25). The active sites for these three activities are apparentlyapartof thesamegeneproduct since they are coordinately affected by single muta-tions. Biochemical studies, however, show only littleoverlappingof the three active sites. Thus, each of the three activities can be separately inhibited by different chemical reagents (Go-recki andPanet, manuscriptinpreparation).
Our findings with the late replication-defec-tive mutants indicate that in terms of DNA polymeraseandtrp-tRNAbinding, reverse
tran-scAiptaseis either not atall (LA 343 and LA 672) or only slightly (LA 338) thermolabile. In fact, the lesions responsible for the phenotype
on November 10, 2019 by guest
http://jvi.asm.org/
442 PANET, WEIL, AND FRIIS
served need not necessarily be in the genetic
elementcodingfor theviralreversetranscriptase
(pol) butcould be locatedin anunknown func-tion essential for correct virus assembly. More
probably, however, defective reverse transcrip-tasemoleculesarethemselvesimpededinsome kind of self-assembly process, and the actual defectsarecausedbylesions inpol. Thefindings
presented inFig.7andTable 2, inany case, go alongwayinexplainingthe lack ofinfectivityof virusisolated from cultures maintainedat42°C.
Certaincomplications remain, however,if the late defective mutants are to be explained
en-tirelyonthe basis of lack ofreversetranscriptase incorporation. First,there is theinterestingfact thatwhereasspecificactivityof DNA
polymer-aseobserved with LA672isolated from cultures at42°C was reduced sixfold(andmore insome experiments), trp-tRNA binding by the same viruspreparationswasless reduced. Thisfinding
was consistently observed in repeated experi-ments.Hence,for LA672,the fractional
comple-mentofreversetranscriptase,whichdoes incor-porateintovirionsgrown at42°C,seems tohave retained more trp-tRNA binding activity than DNA polymerase activity. This would point
quite definitely to a pol lesion. Furthermore,
analyses of recombinants made between the avian leukosisvirus, RAV-6, and LA672 (Joho, Stoll, Friis, andWeissmann,unpublished obser-vations) showed that T, RNase-resistant
oligo-nucleotidesdirectlyassociatedwith the temper-ature-sensitive phenotype were those which mapped in theregion defined ascodingforpol. Likewise, LA 338 can be considered a likely
mutant inpol. A
significantly
shorter half-life for inactivation ofthereversetranscriptasewasnoted with thismutant (Table 1),regardlessof whether virus preparationshad beengrown at 35 or 42°C. Since this mutant shows an addi-tionalveryleaky, early, transientdefect (27), it may well be speculated that there is a slight
(leaky) early pol-related defective phenotype.
Evidence for this earlydefect can be found in thethermolabilityreferredtoabove and in sim-ilar recent experiments showing temperature-sensitive behavior of LA 338 in terms of both DNApolymeraseand RNaseHactivities
(Moll-ing andFriis,manuscriptinpreparation). The amountofenzyme innoninfectious viri-onsofLA 338, LA343, and LA 672 produced at the nonpermissive temperature is three- to six-foldlower thanin the samevirusesproducedat the permissivetemperature. The amount of re-verse transcriptase per wtASV virion was cal-culated by radioimmunoassay to be 30 to 70 molecules (15). Asixfold reduction would leave 5 to 12 enzyme molecules in mutant virions
produced atthe nonpermissive temperature. It isinteresting thatthereductionininfectivity of mutantviruspreparationsgrown atthe nonper-missive temperature is 100- to 500-fold. This disparity between loss ofenzyme and reduced infectivitymaybeseen asevidence that infectiv-ity requires more thanjust a few molecules of reverse transcriptase per virion, or it may be interpretedtosuggestthat, indeed, LA 338,LA 343, and LA 672 incorporate notjust reduced amounts ofreverse transcriptase, but also con-tain molecules of lowerspecific activity.
ACKNOWLEDGMENTS
Theencouragement ofN.Goldblum isgratefully acknowl-edged. This workwasinitiated whileoneofus(A.P.) was the recipient ofanEMBO short-termfellowship.
This workwassupported bytheStiftung Volkswagenwerk and the Israel Academy ofSciences, and in part by the Sonderforschungsbereich47(Virologie)ofthe Deutsche For-schungsgemeinschaft.
LITERATURE CITED
1. Baltimore, D. 1970. RNA-dependent DNA polymerase in virions of RNA tumor viruses. Nature (London) 226:1209-1211.
2. Blair, D.G., W. S. Mason, E. Hunter, and P. K. Vogt.
1976.Temperature sensitive mutants of avian sarcoma
viruses:geneticrecombination betweenmultipleor co-ordinatemutants and avian leukosis viruses. Virology 75:48-59.
3. Friis, R.R., and E. Hunter.1973. Atemperature sensi-tive mutant of Rous sarcoma virus that is defecsensi-tive for replication. Virology 53:479-483.
4. Friis, R. R., W. S. Mason, Y. C. Chen, and M. S. Halpern.1975.Areplication defective mutant of Rous sarcomavirus which fails to make a functional reverse transcriptase.Virology 64:49-62.
5. Friis, R. R., H. Ogura, H. Gelderblom, and M. S. Halpern.1976.Thedefective maturation of viral prog-eny withatemperaturesensitive mutant of avian sar-comavirus.Virology 73:259-272.
6. Friis, R. R., K. Toyoshima, and P. K. Vogt. 1971. Conditional lethal mutants of avian sarcoma viruses. I. Physiologyof ts 75and ts 149. Virology 43:375-389. 7. Grandgenett,D.P., A. C. Vora, and A. J. Faras. 1976.
Different states of avian myeloblastosis virus DNA polymerase and their binding capacity to primer tRNA"'. Virology75:26-32.
8. Hanafusa, H.,D.Baltimore,D.Smoler, K. F. Watson, A.Yaniv,and S.Spiegelman. 1972.Absence of po-lymeraseprotein in virions of alpha type Rous sarcoma
virus.Science177:1188-1191.
9. Hunter, E., and P. K. Vogt.1976.Temperature sensitive
mutantsof avian sarcomaviruses: genetic
recombina-tion with wild type sarcoma virus and physiological analysis ofmultiple mutants. Virology 69:23-34. 10.Linial,M., andW.S. Mason. 1973.Characterization of
twoconditional early mutants of Rous sarcoma virus. Virology 53:258-273.
11. Lowry, 0. H.,N.J.Rosebrough, A. L. Farr, and R. J. Randall. 1951. Protein measurement with theFolin phenol reagent. J. Biol.Chem.193:265-275.
12. Mason, W.S.,R. R.Friis,M.Linial, and P. K. Vogt.
1974. Determinationofthe defective function in two
mutantsofRoussarcomavirus.Virology 61:559-574.
13. Moelling, K.,D.P.Bolognesi, H. Bauer, W. Busen, H. W.Plassmann,and P. Hausen.1971.Association J. VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
theRNAmoiety of RNA-DNA hybrids.Nature (Lon-don) New Biol. 234:240-243.
14. Nishimura,S., and B. I. Weinstein. 1969. Fractionation of rat liver tRNA. Isolation oftyrosine, valine,serine andphenylalanine tRNA and theircoding properties. Biochemistry 8:832-842.
15. Panet,A.,D.Baltimore,and T.Hanafusa.1975. Quan-titationof avian RNA tumor virus reversetranscriptase byradioimmunoassay.J. Virol. 16:146-152.
16. Panet,A.,W.H.Haseltine,D.Baltimore, G. Peters, F.Harada,and J. E.Dahlberg.1975.Specific binding oftryptophan-transfer RNA to avian myeloblastosis virusRNA-dependentDNApolymerase (reverse tran-scriptase). Proc. Natl. Acad. Sci. U.S.A. 72:2535-2539. 17.Panet,A.,L.M.Verma,and D. Baltimore. 1974. Role of thesubunits of the avian RNAtumorvirusreverse transcriptase. ColdSpringHarborSymp. Quant.Biol. 39:919-923.
18. Rogg,H.,W.Wehrli,and M. Staehelin.1969.Isolation ofmammalian transfer RNA. Biochim.Biophys. Acta 195:13-15.
19.Taylor, J. M. 1977. An analysis of therole of tRNA species asprimersfor thetranscriptioninto DNA of RNA tumor virus genomes. Biochim. Biophys. Acta 473:57-71.
20. Temin,H.M., and S.Mizutani. 1970. RNAdependent DNA polymerase in virions of Rous sarcoma virus.
21. Toyoshima, K., and P. K. Vogt. 1969. Enhancement aind inhihition of avian sarcoma viruses wypolycations
andpo)lyanions. Virology38:414-42(i.
22. Toyoshima, K., and P. K. Vogt. 1969. Temperature
senisitiveimiutantsofan aviansarcoma vitus. Virology
39:9310-9:31.
2:3. Verma,I. M. 1977. T'he reverse transcriptase. Biochim. Biophvs.Acta 473:1-38.
24. Verma, I. M., W. S. Mason, S. D. Drost, and D. Baltimore. 1974. D)NA polvmerase activityfrom two ternperatUre-sensitivemutantsof Rous sarcoma virus is therniaolaile. Natture (IAndon) 251:27-;31.
25. Verma, I. M., H. E. Varmus, and E. Hunter. 197(6. Characterizationofearlyt.emperaturesensitivemutants ofaviani sarconma viruses: biological properties,
ther-molahility of reversetranscriptaseinvitio and synthesis
ofviralD)NA ininfected cells. Virology 74:16-29. 26. Watson,K.F., R.Nowinski,A.Yaniv, and S.
Spie-gelman. 1972. Serological analysis of the
deoxyribo-nucleic acid polymerase of avian oncornaviruses. 1. P'reparationandcharacterizationofnaospeeific anti-serunm withp)UrifieddeoxyrihonuLcleicaci(dpolymerase.
J.Vitol.10:951-958.
27. Wyke, J. A., andM.Linial.197:3.Temperature-sensitive aivian sarcomaVirtLses: at physiological comparison of twenty mutants. Virology53:152-1 61.
on November 10, 2019 by guest
http://jvi.asm.org/