Vol.29, No. 2 JOURNALOFVIROLOGY, Feb.1979,p.799-804
0022-538X/79/02/0799/06-0000$02.00/0
Monomer and Multimer Covalently Closed
Circular Forms of
Rous
Sarcoma Virus DNA
GERARD GOUBIN AND MIROSLAV HILL*
Departmentof Cellular and Molecular Biology and Equipe de Recherche No. 148 du Centre National de la RechercheScientifique,Institute
of
Cancerology
andImmunogenetics, Villejuif,
FranceReceived for publication 21 August 1978
Covalently closed circular moleculesof viral DNA synthesized in virus-infected
cells are composed mainly of monomers sedimenting at 22 to 27S in neutral
sucrosegradients.These monomers are detected by annealing with
complemen-taryDNAortransfectionassay.However, 11%oftheinfectious circlessediment
faster thanmonomers. There isa peakat 32S whichmaycorrespondto dimer
molecules. Traces of infectivity (about 3%) found between32Sand65S suggest
thepresenceof higher oligomers.Inalkalinesucrose gradients,covalently closed
monomers arefound at 64 to 71S.Infectivity ofthese monomers isreduced by
alkalitreatment toless than one-tenth, and, perhaps for thisreason, noinfectious
dimersorhigheroligomersareobserved. It has been shown thatupon
resedimen-tation the dimers of 95S can be separated from monomers and detected by
hybridization.
One of the DNA intermediates synthesized
duringthe lifecycleof thetypeC retrovirus has
a closed circular conformation (7, 26) and is
thoughttoarise fromalinear duplex of6 x 106
daltons (23).Giannietal. (6) firstmeasured the
size of the DNA circles and found that, in the
case of murine leukemia virus, the contour
length is characteristic of a molecule with a
molecular weight of about5.5 x106. In the case
of avian sarcoma virus, similar contour length
measurementrevealed thepresenceof different
molecular speciescorresponding in size to
non-defective, transformation-defective (td), and
de-fective genomes containing deletions of from
one-thirdtotwo-thirds of the viralinformation,
respectively (11). Sedimentation coefficients of
genome-sizeformIDNA in neutral and alkaline
sucrose gradients were 30S and 68 to 70S for
murineleukemia virus (7), 24 to 29S and62 to
65S foranaviansarcomavirus (10, 11),and 21S
and32S foranavian reticuloendotheliosisvirus
(5),respectively.
Supercoiled self-replicating circles of
mito-chondrial (22) or simian virus 40 (SV40) (20)
DNA,forexample, have been shownto contain
asmallpercentage of double- and multi-length
DNAforms.These forms are supposed to result
from errorsin replication, from recombination,
or from both (1, 20, 22). Retrovirus DNA was
never showntobeself-replicating. However, in
thispaper weshowthatlargerforms resembling
dimeric and oligomeric circular molecules are
found aswell asthemonomericforms.
Schmidt-Ruppin strain ofRous sarcoma virus
(RSV), subgroup D (SR-D), no. 304, and its td
derivativeno.300used in this workwereisolated
earlier(14). Viral DNAwaspurifiedfrom
virus-infected chicken cells 30 h after infection as
described elsewhere (13). Briefly, chicken
em-bryo fibroblasts (CEF; C/E phenotype,positive
for chickhelper factor)weregrown inmonolayer
cultures. About24hafterseeding, the cultures
were infected, in the presence of2 ,ug of
Poly-breneperml, withSR-Dor tdSR-Dat a
multi-plicity of 1 to 5 focus-forming units or 1 to 5
infectious units per cell, respectively. Thirty
hourslater the cellswerefractionatedaccording
toHirt (17), and the nucleic acidswerepurified
from the Hirtsupernatantand bandedto
equi-librium in a cesium chloride-ethidium bromide
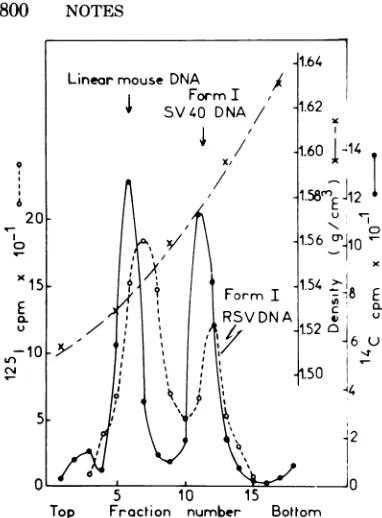
densitygradient asshown inFig. 1.ViralDNA
was detected by
"2'I-labeled
RSV DNA probeandfound in twobands atdensities
character-istic ofsupercoiled (form I)andnonsupercoiled
(forms II and III) DNAmolecules. The
differ-enceindensityof thesetwomolecularspeciesof
viral DNA and the corresponding density
markersseen in Fig. 1 wassupposed tobe due
to the different guanine plus cytosine contents
(N.Stedman,R.Mariage,G.Goubin,J.Hillova,
and M.Hill,J.Gen.Virol.,inpress).Thisfigure also showsthatform IRSVDNAcanbereadily
recovered if gradient fractions containing the
denser (but notthe lighter) part ofthe form I
SV40DNA bandarepooled.
Inallexperiments describedhere,suchapool
ofgradient fractionswassubjected to asecond
equilibrium cesium chloride-ethidium bromide 799
on November 10, 2019 by guest
http://jvi.asm.org/
800 NOTES
_'1X~~~~~~~~~~~
-1.60 A4i20-U
o aV ~~~~~~~~~~1.56ca1~
[image:2.505.69.260.61.320.2]15- rm
1.54Bo-tto
FIG.1.~~~~Iqiiruceiu~Frchoieehiimbo
RSV RSVDNA wU
(p 7.)(.11S)toehrih05 5lo2ehdu
4'
15201~~6
U-10idY2 mgm) inenldniy akr t1C labee'-r)IS4anliermueDA)ad
solidCsCltoa
finaldenstyof18gm.Ci50
5
5 10
150
Top Fraction number Bottom
FIG. 1. Equilibriumicesiumpchioride-ethidium
bro-midedensity gradient centrifugation of supercoiled
RSV DNA. About 1.5x 10' CEFwereinfected with SR-D for 30 h and thenlysed, and thenucleic acids
werepurified fromthe Hirtsupernatantasdescribed
(13). One-quarter of the nucleic acidswasdissolved in 3.5 mlof0.015MNaCs-0.0015 M sodium citrate (pH7.0) (0.1xSSC) togetherwith0.5mlofethidium bromide (2 mg/ml), internal density markers ("C-labeledform ISV40and linearmouse DNAs), and solidCsClto afinaldensityof1.582g/cm'.
Centrif-ugation wascarried outin a Spinco Ti 50 rotor at
40,000rpm at150C for 70 h.Fractionsof0.3mlwere
collected from the top, and 0.2-ml samples were
weighedinacalibratedmicropipettetodetermine the
density. Samples of25
ILI
were deposited on glassfiber filters and dried, and the radioactivity was
measured in a Nuclear Chicago liquidscintillation spectrometer.Inhybridizationexperiments, 50.2g of
carrier rat thymus DNA in 1 ml of CsCl solution
(density1.5g/cm3) wasaddedtoeachfraction.Then
ethidium bromide was removed by treatment with
isopropanol-water (9:1), andthefractions were
di-alyzedagainst0.1xSSC. NaOHwas addedtoafinal
molarityof0.3Mi thefractionswereheatedat80°C for 2h, cooled, andneutralizedwith 2N HColMf Tris solution, and the DNA wasprecipitated with ethanolandspundownbycentrifugation. The DNA
pelletwasdissolved in 50
jil
of0.6MNaCl-0.02MTris-hydrochloride (pH 7.5)-0.01 M EDTA solution
containing 2,000 cpm of "'L1labeled genomic RSV RNA (specific activity20x 106 to80X 106CPM/tg) prepared accordingto Commerford (3) with
modifi-cations(27).Hybridizationswerecarriedout at680C for72h. Then thesamplesweremixed with0.5mlof
2x SSC solution containing 100 pg of RNase A (Worthington; heated at 80°C for 10min) per ml, incubated at 37°C for 60 min, and trichloroacetic
densitygradient centrifugation, and formIRSV
DNA was recovered again from fractions
con-taining the heavier part of form I SV40 DNA
density marker. It was estimated (profiles not
shown) that after this purification procedure
formIRSVDNA wasvirtually devoid (lessthan
1.5%) of form II and III RSV DNA
contami-nants. Contamination by chromosomal DNA
molecules carrying an infectious provirus was
estimated to be even lower, not exceeding
0.02%. Thisdegree of contamination was
calcu-lated from the fraction of chromosomal DNA
foundafter Hirt fractionation in the Hirt
super-natant and from the specific infectivity of the
DNA in the Hirt pellet. We conclude that if
linear DNAmolecules sedimenting fasterthan
supercoiled circles of6 x 106 daltons
contami-nateformIRSVDNApreparations, theycannot
be detected by hybridization and transfection
techniques.
Figure2showsthe sedimentationprofile ofa
purifiedform IRSV DNA ina neutralsucrose
gradient.RSV DNAwasdetectedby annealing
with 3H-labeled complementary DNA (cDNA)
and foundtosedimentmainly ina peak of 25S
correspondingtosupercoiled circles of about 6
x
10'
daltons (2). A shoulder seen in Fig. 2 at20Smaycontainopencircles of thesamesize.A
detectable amountof the RSVDNA, however,
sedimented much faster than 25S witha small
(andreproducible in two experiments) peakat
47 to58S. The evidence derived from transfec-tionassays was more compelling. It wasfound
that only about 89% of the infectious material
consisted of22 to 27S monomers. Asmallpeak
ofinfectivitywas observed in fraction 8 (32S),
consistent with the presence of dimeric circles
(2). The material in this peak and its leading
edge(fractions9and10),thoughhardlyresolved
byhybridization, representedabout8% of
infec-tivity. The rest of the infectivity (about 3%)
sedimented faster than32S,as seeninfractions
(except fractions14and20) between theleading
edge of thepeakand thebottomof thegradient.
In one experiment (not shown), gradient
frac-tions were collectedfrom the bottom and
ana-lyzed as in Fig. 2.Theinfectivity profileinthis
gradient resembledthat in Fig. 2,providing
di-rect evidence that the infectivityseeninlower
partsofthegradientinFig.2didnotarisefrom
contaminationbymonomerssituated in the top
acidprecipitated, andthe amountof acid-insoluble
[125IIRNA
-DNAhybrids was measured in a Packard gamma scintillation spectrometer. Background counts(65 cpm) determinedin control samples con-taining["26I]RNA andcarrier DNA only were sub-tracted.J.
on November 10, 2019 by guest
http://jvi.asm.org/
NOTES 801
9 11325
I12
°°16 o
,,IR~
L-
cSm
n
..
N~~~~~~~ 0
5 1
DN in a-netaursgaiet uecie N
~~~~~~~~
2
~~~~~~~~~~.0
Dwa puiidfo bu . 0 iu-netdCE
j6 {20
q 1522
andtwice banded in cesium chloride-ethidium
bro-mide densitygradients as shown in Fig. 1. Pooled
gradientfractionscontainingthis DNAtogetherwith
GC-labeledform I SV4o DNAdensitymarkerand10 pg ofyeast tRNA were extracted with
isopropa-nol-water,dialyzed against O.lx SSC, and ethanol
precipitated. Theprecipitate wasdissolvedin 0.2 ml
of 0.lx SSCanddepositedon the top of a linear
10-mlgradient(5 to 20% sucrose in 0.1 MNaCI-0.01M
This-hydrochloride [pH7.41-0.001 MEDTA [NTE] solution) and a 0.5-ml cushion of 60% sucrose in NTE.
Centrifugation was carried out in a SpincoSW41
rotor at39,000rpm for 3 h at4°C.Fractions of 0.6 ml
were collected from the top, and the density was
measuredasinFig. 1. Forhybridization, 0.1-ml
sam-plestogetherwith 20 pg ofcarriercalfthymus DNA
weretreatedwithNa2H, ethanolprecipitated, then
dissolvedin the hybridization buffer containing7(W)
cpmof3H-labeled tdPR -RSV-CcDNAprepared
ac-cordingto Taylor et al.(25) andhybridizedat68wC
for 17 has inFig. 1.Afterhybridizationthesamples
weremixedwith2mlof 0.03 M sodiumacetate (pH
4.5)-0.2 MNaCI-0.0i3MZnSO4solutioncontaining
10 pg ofsonically disrupted and alkali-denatured
calf thymusDNAand 41.l ofSlsingle-strand-specific nuclease(24)andincubatedat50°C for2h. The Sl-resistant material([3H]cDNA-DNA hybridsand self-annealed "4C-labeled SV40 DNA)wastrichloroacetic
acid precipitated and assayed for radioactivity. Background tritium counts (8 cpm), determined in
controlsamples containing[3H]cDNA and carrier DNAonly, weresubtracted. Theposition ofthe 14C-labeledformISV40 DNA,usedas aninternal sedi-mentation markerof 21S,isgiven byan arrow.
Sed-imentationcoefficients ofRSVDNAwerecalculated 12
:io
0
'-8
x
a-6
(NI ("4
2
12
oC
-o
4) N
E
L 0
C
0
Z
L
'-2 FormI SV40 DNA
53S
643
1.20
I~
~~~~~~~~~~~
,r1'
/1.124
.11.:
/6ois,
11.06
Top Fractionnumber Bottom
FIG. 3. Sedimentation ofthe supercoiled tdSR-D DNA in an alkaline sucrosegradient. Supercoiled DNAwaspurified fromabout 7.5x108virus-infected CEFas inFig. 2, and deposited togetherwith 3H-labeledformISV40DNA(53S)onthetopofalinear 10-mlgradient (5to20%sucrose)in0.3 MNaOH-0.6 MNaCl-0.01 MEDTA)anda0.5-mlcushionof60% sucroseinthesamesolution.Centrifugationwas car-riedoutinaSpincoSW41rotorat30,000rpmfor1.5 hat4°C.Gradientfractionswerecollectedand an-alyzedasdescribedinFig. 2,except that32P-labeled PR-RSV-CcDNA (700 cpm)wasused,and hybridi-zationswerecarriedoutat68'C for48 h. For
trans-fectionassay, a 0.5-mlportion ofeachfraction was neutralized with 1.1 M HCl-0.2MTris. Then carrier DNAwasadded,and the DNAwasprecipitatedwith ethanol. Thisprecipitatewasdissolved and titrated forinfectivitytoend-pointdilutionasinFig.2.Other
conditionsasinFig.2.
accordingtoMartinand Ames(21). Infectivityassays wereperformed using the calciumphosphate tech-nique (9) asdescribed(15).A 0.5-mlsample of each fractionwasprecipitatedwithethanol in thepresence of10pgofcarriercalf thymusDNAand 0.3 MNaCl. Then the DNAprecipitatewasdissolved in0.1xSSC, seriallyfourfold (or threefold)diluted in N-2-hydrox-yethylpiperazine-N'-2-ethanesulfonicacid (HEPES)-buffered saline,supplementedwithcarrierDNAtoa
finalconcentrationof10pg/ml,andassayedateach dilution inthree(or two) CEF cultures.After2or3 weeks, the cultures were examinedfor the reverse transcriptaseactivityinthe culturemedium(16).The infectivityintermsof infectiousunitswasdetermined by end-point dilution from the fraction of positive cultures (12) and normalized for 109 virus-infected CEF.
VOL. 29,1979
on November 10, 2019 by guest
http://jvi.asm.org/
[image:3.505.47.247.62.316.2]5 10 15 Top Fraction number Bottom
6
i
54
0
"- 4
x
E
0-U
a-CV)
31
2
1
Form I SV 40 DNA B
53$
71$
V
1I
1.12lI
Ii I I
b 110E
6~~~~~~~10
** I C
I s I I
\0w,8I 0
Q __
5
10
15
Top Fraction number Bottom
Form I SV40 DNA 53S
95$ 74S
I
E.o8
EU
1.06
-1.04 :
0
.I II
-II
- ;,I.
I'
.I 0
II
II 0 0
'o.
Il Dill
Ii 'I , I.I
., . I. II
Ii
I I.
\ p
5 10 15
Top Fraction nunber Bottom
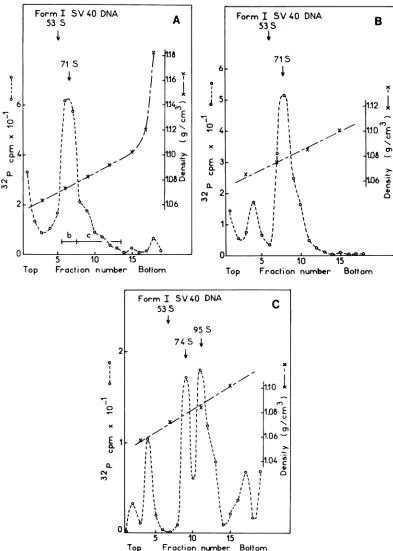
FIG. 4. Sedimentation analysis of the supercoiled SR-D DNA in alkalinesucrosegradients. Supercoiled
DNAwaspurified fromabout 4x108virus-infected CEF and, in (A), sedimented together with the3H-labeled formISV40DNAthroughanalkalinesucrosegradientasinFig. 3exceptthat the time of centrifugationwas
3h. Virus-specificDNA wasdetectedin 0.2-mlportions of gradient fractions by annealing with32P-labeled
PR-RSV-C cDNA(800 cpm)at68°C for40 h. Therest(0.4 ml) of the gradientfractions, in regions shown by
bars b andc,werepooled, neutralized, supplemented with 10pgofyeasttRNA, and ethanol precipitated. The precipitates b andc werecollectedby centrifugation,dissolved in 0.2 ml of 0.1x SSC, addedto2,000cpmof
3H-labeled formISV40DNA,andsedimentedthrough alkalinesucrosegradients in (B) and (C),respectively, asabove, exceptthat this time the60%osucrosecushionswereomitted and thegradientswerecentrifuged for
3.5 h. Gradientfractionswereanalyzed forthepresenceof virus-specificDNAbynucleicacidhybridization
asin(A), exceptthat theradioactivity oftheinput 32P-labeledPR-RSV-C cDNAprobeamountedto500cpm.
Other conditionswere asinFig.2.
6
0
x E
Q4
a-CV)
Form I SV 40 DNA A
53 $
l
1 118
71 S
l
1
< 116I'
~
E112
'l
/
cI
~~~~~~110-X C
b- c~~0I
I
1080AA)t0bs ~~~~1!06
z / ~~b
\ / t~~~~~~~~~~'
b
c0'-b- b
c°os~~~~~~
2 9
0
x
E
a-U
a-C',
cV)
o
2
11
on November 10, 2019 by guest
http://jvi.asm.org/
[image:4.505.67.460.36.587.2]NOTES 803
fractions. We conclude thatatneutral pH form
IRSV DNA is composed of 25Smonomersand
also oflarger forms resembling dimers. These
forms sediment at 32S. Traces of infectious
ma-terial found between 32S and 65S suggest the
presenceofhigheroligomers.
Large covalently closed circles of RSVDNA
werefurther studied under alkaline conditions
to seewhetherthey denatureintosingle strands
or, alternatively, in the absence of alkali-labile
bonds, intoacompactrandom coil.
Sedimenta-tion inan alkalinesucrosegradientis shownin
Fig.
3. The monomer form of tdSR-D DNAsedimentedat64S. This correspondstothe
sed-imentation coefficient ofa form I DNA of6 x
106 daltons under denaturing conditions (2).
Transfection assays show that the monomers
are infectious, though their infectivity was
re-duced to less than one-tenth ofthat found at
neutral pH in Fig. 2. Infectious material
sedi-menting faster than denatured monomers was
found onlyat125S (fraction8ofFig. 3), andno
infectious material wasfound in lowerparts of
thegradient. This could be due toreduction of
thespecific infectivity of covalently closed viral
DNA inalkalinegradients.TheSi-resistant
ra-dioactivity found in fraction 11was not
consid-ered to be relevant to the virus-specific DNA
sincenosuchradioactivitywasfound in further
experiments (Fig. 4A).
To show the alkali resistance of large DNA
formsmoreconvincingly, different size classesof
the denaturedform I SR-D DNAcomposed of
71S monomers and putative 95 to 160S
oligo-mers (Fig. 4A), respectively, wereresedimented
under thesameconditionsinalkalinesucrose as
shown in Fig. 4B and C. In Fig. 4B monomers
resedimentedatabout thesameposition of 74S.
The peak at about 20S shows that they were
partly converted into single-stranded circular
and linear forms of about 3 x 106 daltons. On
the other hand, resedimentation of the 95 to
160S material (Fig. 4C) separatedDNAforms of
about95S frommonomers of74S. Further evi-dence that the peak at 95S represents a
dis-cretely sedimenting specieswas obtainedwhen
the experiment shown in Fig. 4 was repeated
under thesameconditions.Againapeak ofDNA
formssedimenting faster (92S) than the
mono-mers wasresolved.The differencebetween the sedimentation coefficients of monomers of 6 x 106daltons and theirdimers,predicted from the
data obtainedinearlier studieswithotherDNAs
(2), wouldresemble that observed inthesetwo
experiments.
The demonstration in this paper oflarge
cir-cular
fonrs
of RSV DNA raises furtherques-tions. Aretheseforms composedofgenomic or
both genomic and subgenomic submolecules?
Are these molecules unicircular or circular
catenated? Finally, are they generated from
monomersbyadouble recombination(4, 18)or, although this isunlikely, byaberrantreplication
(8, 19)ofviral DNAmoleculesorrecombination
of viral and cellular DNAmolecules?It is
con-ceivable that recombinationeventsinviralDNA
circles would explain the generation of
recom-binant virusesindouble-infected cells.
We thank DominiqueStehelin for kindly providing cDNA probesandMonique Kalekine for excellent technical assist-ance.
Thisinvestigationwassupported by Institut Nationalde la Santeetde la RechercheMedicale,contractsA.S.R.no.1and A.T.P. 28.76.60, and Delegation Generale a la Recherche ScientifiqueetTechniquecontract76.7.1656.01.
LITERATURE CITED
1. Benbow, R. M., M.Eisenberg,and R. L. Sinsheimer. 1972.Multiplelength DNA molecules of bacteriophage .>X174.Nature(London) New Biol. 237:141-144. 2. Clayton, D.A.,and J.Vinograd. 1967.Circulardimer
and catenate forms of mitochondrial DNA in hunman leukaemic leukocvtes. Nature (London) 216:652-657. 3. Commerford,S. L. 1971. lodinationofnucleic acids in
citro.Biochemistry 10:1993-1999.
4. Flory,P.J., Jr.,and J.Vinograd. 1973. 5-bromodeox-yuridinelabelingofmonomeric and catenated circular mitochondrial DNA in HeLa cells. .J. Mol. Biol. 74: 81-94.
5. Fritsch, E., and H. M. Temin. 1977. Formation and structure ofinfectious DNA ofspleennecrosis virus. J. Virol. 21:119-130.
6. Gianni,A. M., J. R.Hutton, D.Smotkin, and R. A. Weinberg. 1976.Proviral DNA ofMoloney leukemia virus: purification and visualization. Science 191: 569-571.
7. Gianni,A.M.,D.Smotkin,andR. A.Weinberg.1975. Murineleukemiavirus:detectionofunintegrated dou-ble-strandedDNA forms of the provirus. Proc. Natl. Acad. Sci. U.S.A.72:447-451.
8. Goebel, W.,and D. R. Helinski. 1968.Generation of' higher multiplecircularDNA forms in bacteria. Proc. Natl. Acad. Sci.U.S.A. 61:1406-141:3.
9. Graham, F. L., and A. J. van der Eb. 1973. A new techniqueforthe assay ofinfectivityofhuman adeno-virus 5 DNA.Virology52:456-467.
10. Guntaka,R.V.,B. W. J.Mahy,J. M.Bishop, andH. E. Varmus. 1975. Ethidium bromide inhibits appear-anceofclosed circularviral DNA and integration of virus-specificDNA in duck cells infected by avian sar-comavirus. Nature(London)253:507-511.
11. Guntaka,R. V., 0. C.Richards, P. R.Shank, H.-J. Kung,N. Davidson,E.Fritsch,J. M.Bishop,and H. E. Varmus. 1976.Covalently closedcircular DNA ofavian sarcoma virus:purification fromnuclei of in-fectedquailtumorcells andmeasurement byelectron microscopy andgelelectrophoresis.J. Mol. Biol. 106: 337-357.
12. Hill, M.,and J. Hillova. 1976. Genetic transformation of animal cells with viral DNA of RNA tumor viruses. Adv. CancerRes. 23:237-297.
13. Hill, M.,N.Stedman,R.Mariage, G.Goubin,J. Hil-lova, andM. Roussel. 1978. Unintegratedand inte-grated viral DNA in Rous sarcoma virus-infected chickencells,p.155-177.In S. Barlati and C. de Giuli-Morghen (ed.), Avian RNA tumor viruses. Piccin, Padua.
14. Hillova, J.,D.Dantchev,R.Mariage,M.-P.Plichon,
VOL. 29,1979
on November 10, 2019 by guest
http://jvi.asm.org/
andM. Hill. 1974.Sarcoma and transformation-defec-tive virusesproducedwith infectious DNA(s) from Rous
sarcomavirus(RSV)-transformed chickencells.
Virol-ogy62:197-208.
15. Hillova, J., M. Hill, G. Goubin, and D. Dantchev.1975. InfectivityofRoussarcomacellDNA: comparison of twotechniques of transfectionassay. Intervirology 5: 367-374.
16. Hillova, J.,M.Hill,and M.Kalekine.1976.Inability of the nondefective Roussarcoma provirus to generate,
upontransfection,atransformation-defective virus.
Vi-rology 74:540-543.
17. Hirt,B. 1967.Selective extraction ofpolyoma DNA from infectedmouse cellcultures.J. Mol.Biol. 26:365-369. 18. Hudson, B., andJ.Vinograd.1967.Catenated circular DNA molecules in HeLa cell mitochondria. Nature
(London)216:647-652.
19. Jaenisch,R., and A.J. Levine. 1973. DNAreplication of SV40-infected cells. VII. Formation ofSV40 cate-nated and circular dimers. J. Mol. Biol. 73:199-212. 20. Kelly,T.J., Jr.,and D.Nathans. 1977. Thegenomeof
simian virus 40. Adv. Virus Res.21:85-173.
21. Martin,R. G.,and B. N. Ames. 1961. A method for determining the sedimentation behavior ofenzymes:
application to proteinmixtures. J. Biol. Chem. 236:
1372-1379.
22. Nass, M. M. K. 1974. Structure, synthesis, and transcrip-tionof mitochondrial DNA in normal, malignant, and drug-treated cells,p.261-307. InK. W. McKerns (ed.), Hormones andcancer.Academic Press,NewYork.
23. Shank, P. R., andH. E.Varmus.1978.Virus-specific DNAin the cytoplasm of aviansarcomavirus-infected cells isaprecursortocovalently closed circularviral DNAin the nucleus. J. Virol. 25:104-114.
24. Sutton, W. D. 1971. A crude nuclease preparation suitable for use inDNA reassociation experiments. Biochim.
Biophys. Acta240:522-531.
25. Taylor, J. M., R. Illmensee, and J. Summers.1976. Efficient transcription of RNA into DNA by avian
sarcomaviruspolymerase.Biochim.Biophys. Acta442: 324-330.
26. Varmus,H.E.,R. V.Guntaka,C. T.Deng, and J. M. Bishop. 1974. Synthesis, structure, and function of avian sarcoma virus-specific DNA in permissive and
nonpermissivecells.ColdSpringHarbor Symp. Quant.
Biol. 39:987-996.
27. Woo,S. L.C.,J. M.Rosen,C. D.Liarakos,Y. C.Choi, H.Busch,A. R.Means, and B. W. O'Malley. 1975. Physical andchemicalcharacterization of purified oval-buminmessengerRNA.J. Biol. Chem.250:7027-7039.
on November 10, 2019 by guest
http://jvi.asm.org/