0022-538X/81/040115-10$02.00/0 Vol. 38, No. 1
Alignment
of
a
Restriction
Map with the
Genetic Map of
Bacteriophage T4
JIING KUAN YEE AND ROBERT C. MARSH*
Biology Program, University of Texas at Dallas, Richardson, Texas 75080 Received1October 1980/Accepted 31 December 1980
A restriction map of the bacteriophage T4 genome was aligned with the T4
genetic map. Included were the cleavage sites for BamHI, BglII, KpnI,
PvuI,
Sall, and XbaI. The alignment utilized the factthat the T4 genetic map had been
orientedpreviously withrespect to a T2/T4 heteroduplex map. DNA fragments
fromaBgII digestion ofcytosine-containing DNA from a T4dCTPase- denA
denB(rIIH23B)alc mutant werehybridized withfull-length chromosomal strands
of bacteriophageT2,and the heteroduplexeswere examined byelectron
micros-copy. From their lengths and patterns of substitution and deletion loops, the
heteroduplexes formed with6ofthe 13BglIl fragmentscould beunambiguously
identified andpositioned on the T2/T4 heteroduplex map. The ends of the T4 DNAstrands in the heteroduplexesdirectly identified thelocation of 10BglII cleavage sites. The remaining three BglII cleavagesites could be assigned to the
T2/T4 heteroduplexmapbasedontheir relativelocations on the restriction map.
Itwasalsopossibletoidentify thesourceof the DNAstrands(i.e.,T2 or T4) in
fourpreviously unassigned deletion loopsonthe T2/T4 heteroduplex. Among the
BglII fragments identified in heteroduplexes was the fragment containing the
rIIH23B deletion; this deletionwas used as the primary point of reference for
alignment of the T4 restriction map with the T2/T4 heteroduplex map and,
hence, with theT4geneticmap.
Since the isolation ofbacteriophage T4
mu-tantswhich allow thegrowth and packaging of
DNA containing
cytosine
instead of thegluco-sylated
hydroxymethylcytosine
residuesnor-mally
present(15),
T4 DNAhas beenamenabletoanalysis with restriction endonucleases.
Cir-cular
cleavage
mapsof the 166-kilobasepair
T4genomehave been
published
for restrictionen-zymes
BamHI,
BglII, KpnI,
SalI, SmaI,
andXhoI (3, 6,
14),
and maps forBglI, EcoRI,
HindIII,
and PstI have beencompleted
recently
(2,12). In the
accompanying
report,the sites of cleavage by PvuI and XbaI were added to acomputer-fitted,
unified map of theBamHI,
Bglll, KpnI,
andSailI
cleavage
sites(9).
Alto-gether,
morethan250cleavage
sites have beenidentifiedontheT4 genome.
We extend the usefulness of these maps
by
reportinghere an alignmentbetween our
com-puter-fitted restrictionmap and the T4genetic
map. Heteroduplexes wereformed between T4
BglII fragments
andfull-length
T2chromo-somalstrandsand examinedby electron
micros-copy. Fromthepositionsof the ends of the T4
strands relative to known
T2/T4
nonhomolo-gies,theBglll
cleavagesitescould be locatedontheT2/T4heteroduplexmap. This made
possi-ble an
alignment
between the restriction mapandthe T4 genetic map, asthe T2/T4 hetero-duplexmaphad been orientedpreviously with
respect tothe T4geneticmap(7). Thealignment
of therestriction and genetic mapsis fixed by
the
position
ofanrII deletion. Themapsappeartobecorrelated within1to 2kilobases(kb)over
their
entirety.
O'Farrell
et al.(12)
haveindependently
aligned a T4 restriction map with the genetic
map,
primarily
by filterhybridization
ofrestric-tion
fragments
to clonedsegmentsofT4 DNAknowntocarry
specific
genes.Also,
Carlson(2)
has determined a general alignment based on
the
cleavage
patterns ofcytosine-containing
DNAfromT4 alcmutantswhich carryvarious characterized deletions. Our resultsarein
agree-ment.
MATERIALS AND METHODS
Phage and bacterial strains.
Bacteriophage
T456-(amE51dCTPase-)denA(nd28)denB(ArIIH23B)
alc8 and its
restriction/modification-negative
hostsEscherichia coli K803 (supE hsdSrgl
gal
met) andB834 (supE+
ris
mB- galmet) wereprovided by
L.Snyder (15). Originally, this T4 alc mutant was
thoughttocarry the 4.2-kbrIIH23 deletion,but in-steadit hasbeenfoundtocarryalongerrIIdeletion,
definedgeneticallyasextendingfromor neartheend 115
on November 10, 2019 by guest
http://jvi.asm.org/
of therIIAgeneinto theaclocus(8).We refer tothis
deletion asrIIH23B. Byelectronmicroscopyof het-eroduplexes, wefound that the rIIH23B deletion
re-moves5.8 kb and that itsstarting pointatthe endof the rIIA gene isindistinguishable from thestarting point forthe rIIH23 deletion.BacteriophageT2Land E. coliBwereobtained from W. Harm.
Growth of bacteriophage and isolation of DNA. T4 alc quadruple-mutant particles that
con-tainedcytosine-containingDNAweregrownby using firstE. coli K803 and thenE.coliB834,and theDNA
wasisolated by phenolextractionasdescribedinthe
accompanyingpaper(9).T2Lstockswereobtainedby
infectionofexponentially growingE.coli Bwithsingle plaques; Hbrothwasusedasthemedium;itcontains
(per liter)8gof nutrient broth (DifcoLaboratories),
5gofpeptone(Difco),5gofNaCl,and 1gofglucose. The T2Lparticles were purifiedby differential
cen-trifugation and stored in 20 mM Tris-hydrochloride (pH 7.8)-0.5mM EDTA.
T2/T4DNAhybridization.T4 cytosine-contain-ing DNA from the alcquadruplemutantwasdigested
withBglII (NewEngland Biolabs)asdescribed in the accompanying paper (9) and then extracted with phenol. The mixture of T4BglII fragmentswas
hy-bridizedwithfull-lengthT2 chromosomal strands
es-sentiallybythemethod of Davisetal.(4).This method minimizes breakage of the long chromosomal DNA strandsby usingalkalitosimultaneously disrupt phage particles and denaturethe DNA inthesametubeused
forthehybridization. To5x 109 T2 particles and0.2 ,ug of the T4BglII fragments in 45
Pi,
5plof 0.2 M EDTA (disodium salt)-1 MNaOHwasadded. After10minatroomtemperature,the solutionwasadjusted
topH8.5with7
pl
of 1.5 M Tris-hydrochloride (pH 7.8)-0.2 M Tris base, and then 41 to 43pl
of99% formamidewasadded.Renaturationofthe DNAwasallowedtoproceedatroomtemperaturefor2hduring dialysis againstasolutioncontaining0.45MNaCl,0.1 MTris-hydrochloride (pH 8.5), 0.01 M EDTA, and40 to 50% formamide or for5 h without dialysis. The conditionsweredesignedtopermit 50% of the mole-culestoanneal.
Electronmicroscopy. Circular, single-strandedfd DNA (Miles Laboratories, Inc.) containing6,408
nu-cleotides (1) and double-stranded pBR322DNA (ob-tainedfromH.BoyerinE.coliRR1) containing4,362 basepairs (bp) (16)wereaddedtothe T2/T4 hybrid moleculesasinternal length standards. TheDNAwas
spreadontoamonolayer ofcytochromecin the
pres-enceofformamideasdescribedby Davisetal. (4) and thenpickedup oncarbon-coated grids.Thegridswere
rotaryshadowed with Pt and photographed with a
Siemans 1Aelectronmicroscopeatx10,000. Lengths ofDNA molecules weremeasured onprojections of
photographic negatives by using a linear integrator
(NumonicsGraphics).
RESULTS
T2/T4 aicmutantheteroduplex. The use
ofnonhomology loopsasmarkerstolocate the
ends ofBglII fragments ontheT2/T4
hetero-duplexmap, which inturnpermittedan
align-ment of the restriction and genetic maps,
re-quired verification of the published wild-type
T2/T4 heteroduplexmapfor the strains which
weused.Because of their circularpermutation,
T2 and T4 form large circular heteroduplexes
when denatured and reannealed.Measurements
made on suchheteroduplexes formed from T2L
andthe T4alc quadruple-mutantchromosomal
strandsyielded a map of substitution and
dele-tion loops (Fig. 1, outercircle) identical tothe
one constructed by Kim and Davidson (7) for
wild-type DNAs, except for an additional
sub-stitution-type nonhomology labeled G', which
weconsistently observed, and a 5.8-kb deletion
loop that resulted from the rIIH23B mutation
carried by the T4 alc quadruple mutant. As
pointed outby Kim and Davidson (7), this
het-eroduplex map is an idealized map. In individual
heteroduplexes, neighboring loops sometimes
merged to give a bigger loop. In other cases,
large substitution loops underwent additional base pairing, creating several smaller loops;
sometimes loops collapsed or became twisted
andappeared as stretches ofhomologousDNA.
For example, several smaller loopswere often
observedinplaceof the Floop,and the
homol-ogous regions between loops V' and U' or
be-tweenloops H and S often remainedunpaired,
generatingloops larger than normal.
Heteroduplex mapping of the T4 Bgll fragments. The BglII restriction fragments
from atotaldigestion of T4alcmutant
cytosine-containing DNA(Table1)were hybridized with
full-length T2 chromosome strands and exam-inedbyelectron microscopy. Of 50well-spread heteroduplex molecules which were chosen for
measurement,one-halfcould beassigned to
spe-cific T4BglIIfragments.The otherseither could
notbe positioned unambiguously on the
heter-oduplexmap on thebasis of their loop patterns
or did not correspond in length to one of the
knownBglII fragments.Some examples in this
lattercategory were to be expected due to
ran-dom cutting of the ends of T4 chromosomes during phageDNApackaging.
For the initial measurementofthe length of
the T4 DNAstrandin aheteroduplex,the two
arms ofthe substitution loops were averaged,
and the lengths ofthe deletion loops were
di-vided inhalfunlessthe loops were already
as-signedtoT2 or T4DNA byKimand Davidson
(7). In the case of BglII fragments 5 and
7a,
onceheteroduplexeswere correlated with these
fragments,the measured lengths of the T4 DNA
strandswereadjustedupward by
assigrunent
ofpreviously unassigned single-stranded loops to
T4 DNA in order to more exactly match the
knownlengthsofthese fragments.
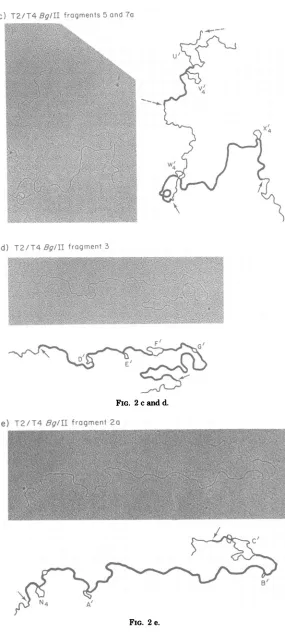
Figure2shows examples of theheteroduplex molecules that could be correlated with BglII
on November 10, 2019 by guest
http://jvi.asm.org/
ALIGNMENT OF T4 RESTRICTION AND GENETIC MAPS 117
0.
-FIG. 1. BgUlI restriction mapofT4(innercircle) alignedwith respecttotheT2/T4heteroduplexmap(outer
circle). The solidarrowsindicateBgUl cleavagesites locatedontheheteroduplexmapbyvisualizationof
heteroduplexes formedbetween T4
BgUII
fragmentsandfull-length T2chromosomal strands. The dashedarrows indicate theBglII cleavagesites assigned to theheterodupkxmap on thebasis oftheirrelative
locationsontherestriction map. The circularheteroduplex mapwasdrawnbyaveragingthelengths ofthe twoarmsofsubstitutionloopsandtakingone-half ofthelengths ofdeletionloops. The letterdesignations
givento someofthenonhomologiesonthe
heteroduplex
mapby KimandDavidson(7)have beenretained;theremainderofthenonhomologiesaredesignatedwithletters that carryprimes.Thesubscripts2and4are
usedtoindicate thesourcesofthesinglestrands in thenonhomologyloops, ifknown(72andT4,respectively).
Theful-length ofthe 5.8-kb rIIH23Bdeletion carried bythe T4 alcmutantisrepresentedas a segment
bisectingbothmaps.Its boundarieswereusedtofixthealignment ofthe maps.
fragments2a,2b,3, 4, 5,and7a.Figure3shows
theaverage
lengths
of theT4DNA strandsandoftheindividualduplexandsingle-stranded
re-gions of all measured heteroduplexes. In the
middle oftheheteroduplex in Fig.2a, a
promi-nent deletion
loop
that corresponds in size to the5.8-kb rIIH23Bdeletion iseasily recognized. About400bp away isthe neighboring deletionloop A,which wasassignedtoT4 DNAbyKim and Davidson (7) because theduplex segments
between loop Aand various rII deletionloops
often open up to form single substitution-type loops,the arms of which can be accounted for
only if loop A is T4 DNA. Missing from the
heteroduplex
inFig.
2aisloop
I',whichshouldbe visible between loopA4 and the left end of
on November 10, 2019 by guest
http://jvi.asm.org/
[image:3.494.54.436.63.458.2]TABLE 1.
BglII
fragments
ofT4alcmutant cytosine-containing DNAFragment Size(bp)a
1 55,840 2a 17,530 2b 17,500 3 14,010 4 12,660 5 10,580 6 7,940 7a 5,540 7b 5,470 8a 4,410 8b 4,350 9 3,330 10 1,180
aSizes, as determinedbyagarosegel
electrophore-sis,wereadjustedslightlybyaleast-squares analysis
designed to yield a best-fit map for the fragments
producedbyBglII and five other restriction endonu-cleases(9).
the molecule.
Apparently
itcollapsed
in thisexample, whichwas chosen for illustration
be-cause basepairingofthe segment between the
rIIH23B and A4 loops had occurred and all
otherregionswerewellspreadduring mounting.
Inother examples of thisheteroduplex, loop I'
was visible eitheras a
deletion-type loop
orasthe
asymmetric
substitution-type loop
showninFig. 3.
Ateach end of theheteroduplex regioninFig.
2aisasingle-strandedtail of T4
DNA;
the single-stranded T2 DNAextends overamuchlonger
distance,
past theedges
of themicrograph.
Cleavage
ofT4alcDNAby BglII
within theHnonhomology region would have generated the 1.32-kbtail ofT4 DNA attheright end of the heteroduplex. Howthe tail ofT4 DNA which
was seen attheleft end of all examples of this
heteroduplex was generated isnot directly
ap-parent. Itcould have resulted fromcleavage by
BglIl
withintheH' deletionloop,
which was notassignedtoT2 or T4 DNA (7).However, as an
examination of the end of the heteroduplex formed with the adjoining
BglII
fragmentshowed (Fig. 2dand 3; seebelow), the H'loop
wasapparentlyT2DNA,andcleavageofthe T4
DNA occurred 560 bp beyond the H' deletion
loop. Thus,the tail of T4 DNAappears to have
resulted from afailure of the T4DNA to base
pair with T2 sequences beyond the H'2 loop,
probably due to insufficienthomology and the
lack of a well-paired neighboring region, as
wouldbe found in acomplete T2/T4
heterodu-plex.
Altogether,
the sequencesassigned
to T4 inthe
heteroduplex
shown inFig.
2a were12.36±0.09kblong (Fig.
3),
indicating
that the T4 DNAstrandwasderived from
BglII
fragment
4,whichwas12.66kblong,asdeterminedbyagarosegel
electrophoresis.
On the BglII restriction map of the T4 alc
mutantDNA, one end of BglII fragment 4
ad-joins the smallestBglIIfragment(fragment10),
andbeyond that is fragment 2b. The
heterodu-plex that corresponds to this fragment is shown in Fig. 2b. It contains the seven nonhomology
loopsfrom K'toO',which extend from near the
end ofBglII fragment 4 in the H substitution
loop in a counterclockwise direction. A small,
eighthloop was present between M' andN',but
it did not occur regularly in T2/T4
heterodu-plexes and thus was notdesignated with a letter.
Given the fact that deletion loops 0 and P
represent T4 DNA (7), the T4 strand in this
heteroduplex was 17.97 ± 0.21 kblong (Fig. 3),
compared with 17.50 kb for BglIIfragment2b
from agarose gels.
The right end of BglIIfragment2b was
gen-eratedbycleavage 190 bp beyond loopO',within
a region of T2/T4 homology. At the left end,
cleavage occurred betweenloopsS andR,
leav-ing space forBglII fragment 10 between
frag-ments 2b and 4 (Fig. 1). Note that inthe
heter-oduplex shown inFig2b thestrand of T2 DNA
terminated within the region of homology
be-tween loops K' and J', leaving the T4 DNA
unpaired over the relatively short distance to thehomology region betweenloops S andR.
Proceeding around the restriction map from
BglII fragment2b, the nextfragmentsfor which
unambiguous
heteroduplexes
were observedwere fragments 7a and 5, both of which are
shownhybridized tothe same T2 DNA strand
inFig.2c.Theintervening fragment 7bisabsent,
andexcept for this particular example, its
cor-respondingsequencealong the T2 DNAwould
have remained undetected, as it contains no
observablenonhomologies. Inagarose gels,
frag-ment7b was 5.47kblong; inFig. 2cthe
corre-spondingT2single-stranded regionwas 5.39 kb
long.
The region to which BglII fragment 7a was
hybridized covered deletionloop V' and
substi-tution loop U'. The 1.39-kb loop V' was not
assigned to either T2 or T4 by Kim and
David-son,butmustbeT4 DNA ifthe measured length
of the T4 strand in theheteroduplexis to match
the length ofBglII fragment 7a. Including V',
the T4 fragment was 5.76 kb long (Fig. 3); in
agarosegels,fragment 7a was 5.54 kb long. The
counterclockwise.end
of fragment 7a could beplaced 1.85 kb beyond loop V'. The other end waslocatedabout 1kbpast the U'substitution
loopin aregion whichnormallyisbase paired in
full-length
T2/T4 hybrids but remainedun-paired inthisheteroduplex.
BglIIfragment5was 10.58 kblongas
on November 10, 2019 by guest
http://jvi.asm.org/
ALIGNMENT OF T4 RESTRICTION AND GENETIC MAPS 119
a) T2/T4
Bg/II
fragment 4H4
b) T2/T4
B_/II
fragment 2bN'
_Wof P4 v ",'IV
FIG. 2. ElectronmicrographsandtracingsofheteroduplexDNAsformedfrom full-lengthT2chromosomal strands and strandsfrom BglIIfragments ofT4.Thearrowsdesignatetheendsofthe T4 DNA strandsand,
thus,BglII cleavagesites.Nonhomologyloopsaredesignated bylettersasdescribed in thelegendtoFig.1.
minedbyagarosegels. Toaccountforthislength
inthecorrespondingheteroduplex inFig. 2c, we
assigned the long arm of substitution loop W'
and the deletion
loop
X'toT4. Altogether, theT4 strandintheheteroduplex thenwas 10.42 ±
0.42 kb
long
(Fig. 3).Bgll
cleavage siteswerelocated2.47kbbeforeloopW'4 and1.85kbafter
loopX'4.
Turning
tothe restriction fragments locatedon the clockwise side of
BglII
fragment 4,ex-amples of the heteroduplexes assigned to
frag-ments 3 and 2a are shown in Fig. 2d and e,
respectively. The heteroduplex in Fig. 2d
con-tained the
readily
recognizedpattern
ofnon-homologies between loops D' and G' and
con-tained a T4 strand with an average length of
14.12±0.13kb(Fig.3). From agarosegels,
BglII
fragment 3 was 14.01 kb
long.
Note that theright endof the
heteroduplex
region
whichad-joined
fragment
4wasfully duplex and,
accord-ingtoits 6.02-kbdistance from
loop G',
termi-nated before the H' deletion
loop,
which was6.23kb from
loop
G'. Thissupports
the earlierassignmentsof
loop
H'toT2 DNA and theBglII
cleavage site at the end offragment
4 to thehomologous
region
beside the H'loop
ratherthantothe DNAwithin the H'
loop.
However,
our
assignment
ofloop
H'toT2 DNAcanonly
be tentative. The left end of the
heteroduplex
withBglII
fragment
3 is located 2.80 kb fromloopD'.
The
heteroduplex
inFig.
2e contained thenonhomologies
C'through
N4,
along
with anunletteredsubstitution
loop
nearN4,
whichwason November 10, 2019 by guest
http://jvi.asm.org/
[image:5.494.104.393.73.450.2]c)
T2/T4B/II fragments5and 7ad) T2/T4 Bg/II fragment 3
_o_~~~~~~~~~~~~~~~~
Ec
FIG. 2cand d.
e)
T2/T4Bg/II
fragment 2aFIG. 2e. 120
B/
on November 10, 2019 by guest
http://jvi.asm.org/
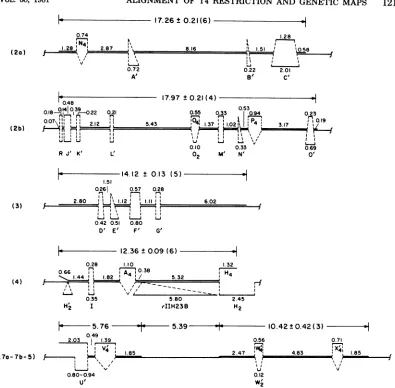
[image:6.494.114.399.43.678.2]17.26+0.21(6)
1.28
t r, '8
JLt1. . ,058
, I
0.22 2.01
B' C'
0.48
0.18074:I0i3 o22 0.21 (2b) i n r- r-
1.1
r-'
Kiu l
R J' K' L'
17.97 t0.21 (4)
0.53
0.55 0.33 0.94 0.23
5.43 1'37M'1102 I,Q19
II I :I II %
Li L-I I L
0.10 0.33 0.69
02 N' 0'o
14.12 0.13 (5)
1.51
0.261 0.57 0.28
r , e
2.80 "11.12' 1.1 6.02
(3) r- r-- rJ
U- L.- L..-j Li
0.42 0.51 0.80
D' E' F' G'
L 12.36 0.09(6)
0.28 1.10 1.32 0.66 20.38
.44' 182' 45.32
(4 )
I\ -._
0.35 5.80 2.45
H2 I rIIH23B H2
2~~~~ ~ ~ ~ ~ ~~~~~04
*- 5.76 5.39 10.42 t0.42( 3) H
0.49
2.03 1.39 0.56 0.71
IIV
(70-7b-5)} H J
~~~
~~18.5
~
2.47 :,} 4.83 \ 6 8'J v u v
0.80-0.94 0.12
U' W2
FIG. 3. MapsoftheT2/T4BglII fragment heteroduplexes. The heteroduplexesarenumbered accordingto
the T4BglII fragment which they contain. Distancesaregiven in kilobases foreach duplex and
single-strandedregion. Theaveragelength±standard deviationfor eachT4strand isgiven above eachmap,and
thenumber of heteroduplexes measured is given inparentheses. The two values given for the length of substitutionlooplFintheheteroduplexwithBglII fragment7arepresentthelengths of thetwoarmsof the
loopinfullT2/T4heteroduplexes.Heretheupper armisnotshown duetothelack of base pairingbeyond the
loopin theheteroduplexwiththeBglII fragment.
onlyoccasionally observed and isnotshown in
Fig. 3. Theaveragelengthof the T4strandwas
17.26 ±0.21kb, comparedwithavalue of17.53
kb forBglll fragment2ameasuredwithagarose
gels. The rightend of theheteroduplex region
terminated 580 bp beyond loop C'. This was
within60bpof thelocationassignedtothesame
BglII cleavage siteby measuringfromloopD'
in theheteroduplexwithBglII fragment3.The
leftend of theheteroduplexwithBglll fragment
2awaslocated1.28kb beforeloop N4,within 170
bp ofthelocationassignedtothisBglII cleavage
site by measuring from loop C" in
heterodu-plexes formed withpartof theneighboring BglII
fragment1 (datanotshown). Several
heterodu-plexes containing this end ofBgll fragment 1
were found, although no unambiguous
hetero-duplexes containingthe entire 56-kbBgll
frag-mentwereobserved.
Alignment of the T4 restriction and
ge-neticmaps.Altogether,theheteroduplexes
be-tween T2 DNA and the T4 BglII fragments
permitted 10 ofthe 13 Bgl cleavage sites on
the T4 alc mutant DNA to be located onthe
T2/T4 heteroduplexmap.Theexactlocation of
thelargerIIH23B deletionontheT2/T4
heter-oduplex map and withinBgJII fragment4 was
also defined.By using this deletionas apointof
0.74 1.28:L
I
0.72
A'
on November 10, 2019 by guest
http://jvi.asm.org/
[image:7.494.51.446.63.451.2]reference and the BglII cleavage sites to
cor-rectly orient theT4BglII restrictionmapwith
respect to theT2/T4heteroduplexmap,thetwo
maps werealigned,asshown inFig.1.Thesmall
differences observed between the locations of thecleavage sitesontherestrictionmap(Fig. 1,
inner circle) and the locations of the cleavage sites on the heteroduplex map (Fig. 1, arrows
pointingtoward theoutercircle) reflect the fact
that the restriction map is a strictly physical
mapof the T4 genome, whereas the
heterodu-plex map was constructed by averaging the
lengthsof the substitution looparmsandhalving thelengths of thedeletion loops. This procedure
was necessary because not all
loops
could beassigned to T2 orT4. Over
long distances,
the contributions fromT2 andT4average out,butovershortdistances,unequalamountsof DNA attributable to T2 or T4 can cause the BglII sites on the heteroduplex map to be shifted relativetothephysical restrictionmap.
AsFig. 1shows,twoofthethreeBgllI
cleav-agesitesnot
directly
locatedby the heteroduplex analysis fallatthe ends offragments thatcon-tain no nonhomology loops, i.e., fragments 8b
and 9. These two neighboring sites could be
locatedontheT2/T4heteroduplexmapby
mea-suring from the
BglII
cleavage site locatedbe-tween
fragments
5and9.The lastBglII
site fallsbetweenfragments8aand6.Itslocationonthe heteroduplexmapcouldnotbe determined
ex-actly duetothepresence of unassigned deletion loops in both of these
fragments;
however, by measuring from theBglII cleavage
site at the end offragment7a,it could be definedaslyingat one end or the other of a 330-bp interval,
which is adistance equal tothe unassigned S' deletion
loop
infragment
8a.The final alignment ofthe BglII restriction mapwith the T4geneticmapis shown in Fig.4.
Also includedarethe cleavage sites forthefive
otherrestrictionenzymesmapped in the
accom-panying report (9). The alignment is based on
theprior orientation of the T2/T4 heteroduplex
map with the T4
genetic
map as described byKim and Davidson(7). The rII locus was used
astheprimary referencepoint for thealignment, with the start of the rIIH23B deletion being
placedattheleftend of the rIIA gene. TherIIA
gene has beenestimatedtocontain 2,300 bp by
electron microscopy ofheteroduplexes formed
withvarious rII deletion mutants (7). This is in
line with length
estimates
based on themolec-ularweightof the rIIA protein (13).
DISCUSSION
BglII
cleavesthecytosine-containing DNA ofT4amE51 nd28
'rIIH23B
alc8
into 13fragments,6of whichwehaveidentifiedinheteroduplexes with full-length T2 chromosomal strands. The ends of these
heteroduplexes
specifiedthe loca-tionsof10of theBglIIcleavage
sitesontheT2/T4heteroduplexmap,providing the framework for (i) theassignment of the remaining 3BglII cleavage sitesonthe
heteroduplex
map,(ii)
the identificationof the source of DNA infourmoreof the nonhomologyloopsontheT2/T4 heter-oduplex map, and (iii) the orientation of the restriction mapwith respect to the T4 genetic
map. The final
alignment
between therestric-tion and
genetic
mapswasfixedby
the location of the startof therIIH23B
deletionatthe end of the rIIAgene.The accuracywith which the ends of the T4
DNA strands in the heteroduplexes with T2 could beidentified, and thus theaccuracywith which the
BglII
restriction sites could beas-signedtothe
T2/T4 heteroduplex
map,canbe gauged byexamining
thosecaseswherehetero-duplexes
werefound forneighboring restrictionfragments (e.g.,
fragments
1and2a orfragments
2a and 3). Inthese twocases, the independent assignment of the restriction sitesvariedbyonly
60and170nucleotidepairs.
This isinlinewiththelimit ofresolution
expected
forheteroduplex
mapping by electron
microscopy.
Because our alignment of the T4restriction
map with the
genetic
map is fixed at the rIIlocus, itsaccuracy overthe
entirety
of themapdepends upon the extent to which the
genetic
map itself represents truephysical
distances along theDNA.Actually,
thecorrelation should bequite good,asthegenetic
map inFig.
3isnot a classical map based only on recombination frequencies, which canvaryfrom region tore-gion within the genome. For many
loci,
the geneticmaprepresentsphysical distances deter-mined bymeasuring
thefrequency with whichtwo
given
genes are carried together on theincomplete
chromosomes found insmallT4par-ticles (10, 11). The positions of the remaining
genes were
interpolated
from recombinationfre-quencies.
The distance inkilobasepairs betweenthe rII and lysozyme genes was measured
di-rectly byelectronmicroscopyofdeletion
heter-oduplexes (7). Thesetwoloci arenearly opposite
one another on the circular genetic map and
thus have served tofix distances along the T4
genetic map at two widely separated points.
Between these points some distortion can be
anticipated, but itshould not be too great.
The location ofthe single BamHI cleavage
site canbe taken as an indication of thedegree
with which the restriction and genetic maps
coincide outside the rII and lysozyme regions.
Our
alignment places
the BamHI site withingene8, verynearitsjuncturewith gene 7. This
on November 10, 2019 by guest
http://jvi.asm.org/
\
P,,r-~~~~~~~A
oe~ ~ ~
FIG. 4. Restriction mapof T4 DNA forBamHI, BglII, KpnI, PvuI, SalI, and XbaI aligned with the T4
genetic map. Theunifiedmapof cleavagesitesforthese restriction endonucleaseswasconstructedbyMarsh
andHepburn (9). Thegeneticmapisanupdatedversionofthe mapofWood and Revel(18), withexpansion
ofthe distancebetween genes30and54in accordancewith the mapofMosig(11). Genes whoseexactlocations areunknownarebracketed.Shadedareasdenote theestimated sitesofgenes,basedonheteroduplexmapping
and the molecularweightsofpolypeptideproducts (18).Thealignmentofthe maps isfixed bythe boundaries
oftherIIH23B deletion carried bythe T4 alcmutantusedtoconstructthe restriction map. The scalejust
insidethegeneticmapshowsthe distance in kilobasepairsaround the map; the totallengthis166kb(9).
is in exact agreement with Wilson et al. (17), who located the BamHI site by digestion ofa
clonedEcoRIfragmentknown to carry genes 7
through9.
Hanggi andZachau (5) have mapped indetail
asegment of the T4 genome that extends from
gene 35throughgene 63. Ouraligrunent places
oneXbaI cleavagesitewithinthe end of gene 34
proximalto gene 35 and another in the middle
ofgene 63, in exact agreement with the locations
determined by Hanggi and Zachau (5). These
authors also identifiedaSaIIsite near theXbaI
site in gene 34, as we did, and showed that a
second
SailI
site lies within thedihydrofolate
reductase
(frd)
gene. Toaccommodate thisas-signment,thefrdgene mustbemoved about 1
kb closertogene 32 than is shown on thegenetic
map inFig.3.Thisshouldrepresent the variance
that canbeexpectedbetween the restriction and
geneticmapsas
aligned
inFig.
3.Usingdifferent
approaches
and different T4mutants, twoother groups of workers have
in-dependently aligned restriction maps with the T4genetic map. Carlson
(2)
hasexamined the 123on November 10, 2019 by guest
http://jvi.asm.org/
[image:9.494.44.444.72.459.2]124 YEE AND MARSH
BglI, KpnI, andSailIcleavage patterns of
sev-eralT4deletionmutants todeterminea
general
alignment
between the maps. O'Farreil et al.(12) have constructed a T4 restriction map which contains the
EcoRI, HindIII,
and PstI cleavage sites and have aligned it with the T4 genetic mapby hybridization of restrictionfrag-ments toclonedsegmentsof the T4genomeand
by correlationof restriction
fragment
sizes with thepattern offragmentsproduced by digestion
of the cloned DNAs. In both cases, there was
agreement with the
alignment
which wefoundfor therestriction and
genetic
maps.ACKNOWLEDGMENTS
This investigation was supported by Public Health Service research grant GM 23608 from the National Institute of Gen-eral Medical Sciences and by Public Health Service Research Career Development Award CA 00490 to R.C.M. from the National Cancer Institute.
LITERATURE CMD
1. Beck, E.,R.Sommer,E.A.Auerswald,C.Kurz,B. Zink, C. Osterburg,H.Schaller,K.Sugimoto,H. Sugisaki, T. Okamoto, and M. Takanami. 1978. Nucleotide sequence of bacteriophage fd DNA. Nucleic Acids Res.5:4495-4503.
2. Carlson,K. 1980. Correlation betweengenetic map and map ofcleavage sites for sequence-specific endonucle-asesSall,KpnI,BgII,and BamHI inbacteriophageT4 cytosine-containing DNA. J. Virol. 36:1-17.
3. Carlson,K., and B. Nicolaisen.1979.Cleavagemap of bacteriophage T4 cytosine-containing DNA by se-quence-specificendonucleasesSalI and KpnI. J. Virol. 31:112-123.
4. Davis, R. W., M.Simon,and N. Davidson. 1971.
Elec-tron microscope heteroduplex methods for mapping regions of base sequence homology in nucleic acids. MethodsEnzymol. 21D:413-428.
5. Hanggi,U.J., andH. G. Zachau. 1980. Isolation and characterization of DNAfragments containing the di-hydrofolate-reductase gene of coliphage T4. Gene
9:271-285.
6. Kiko, H.,E.Niggemann,and W.Ruger. 1979. Physical mapping of the restriction fragments obtained from bacteriophage T4 dC-DNA with restriction endonucle-asesSmaI, KpnI and BglII. Mol. Gen. Genet. 172:303-312.
7. Kim,J.-S., and N. Davidson. 1974. Electron microscope heteroduplex study of sequence relations of T2,T4,and T6 bacteriophage DNAs. Virology 57:93-111. 8. Kutter, E., A. Beug, R. Sluss, L. Jensen, and D.
Bradley. 1975. The production of undegraded cytosine-containing DNA by bacteriophage T4 in the absence of dCTPase and endonucleases II and IV, and its effect on T4-directed protein synthesis. J.Mol. Biol. 99:591-607. 9. Marsh, R.C., and M. L. Hepburn. 1981. Map of restric-tion sites on bacteriophage T4 cytosine-containing DNAfor endonucleases BamHI, BgilI, KpnI, PvuI, Sall, and XbaI. J. Virol. 38:104-114.
10. Mosig, G. 1968. A map of distances along the DNA molecule ofphage T4. Genetics 59:137-151.
11. Mosig,G.1976.Linkage map and genes of bacteriophage T4, p. 664-676. InG. D. Fasman (ed.), Handbook of biochemistry and molecular biology, 3rd ed. Nucleic acids, vol. 2.CRC Press,Cleveland, Ohio.
12. O'Farrell, P., E.Kutter, and M. Nakanishi. 1980. A restriction mapof the bacteriophage T4 genome. Mol. Gen. Genet. 179:421-435.
13. O'Farrell,P.Z.,L. M.Gold,and W. M.Huang.1973. The identification ofprereplicative bacteriophage T4 proteins. J. Biol. Chem. 248:5499-5501.
14. Ruger, W., M. Neumann, U. Rohr, and E. Nigge-mann. 1979. Thecomplete maps of BgIII, Sall and XhoI restriction sitesonT4dC-DNA. Mol. Gen. Genet. 176:417-425.
15. Snyder,L.,L. Gold,and E. Kutter. 1976. Agene of bacteriophage T4 whose product prevents true late transcription on cytosine-containing T4 DNA. Proc. Natl. Acad. Sci. U.S.A. 73:3098-3102.
16. Sutciffe, J.G. 1979. Completenucleotide sequence of theEscherichia coliplasmid pBR322.ColdSpring Har-borSymp. Quant. Biol. 43:77-90.
17. Wilson, G. G., R.L.Neve, G. J.Edlin, and W. H. Konigsberg.1979.The BamHirestriction site in the bacteriophage T4 chromosome is located in or near gene 8.Genetics93:285-296.
18.Wood,W.B., and H. R. Revel. 1976. The genome of bacteriophage T4. Bacteriol. Rev. 40:847-868.
J. VIROL.