Heterogeneity and Evolution
Rates
of
Delta Virus RNA
Sequences
FUMIO IMAZEKI, MASAOOMATA,* ANDMASAO OHTO
FirstDepartmentof Medicine, Chiba University School of Medicine, Chiba 280,Japan
Received 24 May 1990/Accepted14August1990
To investigate the geographical divergenceofdelta virus RNAsequences,868 nucleotides(nt), including the
deltaantigen-coding region,weredetermined in isolates fromtwoJapanese patients,MandS, by polymerase
chain reaction and directsequencing andcomparedwith threepreviously reportednucleotidesequences. The sequenceobtained for hepatitisdelta virus RNAfrom patientMwasapproximately92% identicaltosequences
previously obtained fortwoother strains of hepatitis deltavirus,whereasthesequenceofhepatitisdeltavirus RNA obtained from patient S was approximately 81% identical to the previously sequenced strains. This suggeststhat deltaagentinJapanhasaheterogeneousoriginand the delta virus RNAsequencefromJapanese patientS is the mostdivergentdelta virus isolateyetanalyzed.Tostudythe evolutionrateof delta virusRNA,
viral isolates obtained 3 and 4yearsapartfromeachoftwopatientswerealsosequenced.Itwasestimated that the substitution rateof viral RNAwas0.57 x 10-3ntpersiteper yearinpatientMand 0.64 x 10-3ntper
siteper year inpatient Sfor the deltaantigen gene.
Delta agentwas firstdiscovered by Rizzettoetal. (20)as a novel antigen localized in the nuclei of hepatocytes in
patients infected with hepatitis B virus. This agent was
experimentally transmitted to chimpanzees (21) and
wood-chucks (17) in association with hepadnaviruses. Hepatitis delta virus (HDV) is a 36-nm particle containing acircular RNAgenomeof 1.7 kilobases (2, 4, 9, 19). HDV isendemic
throughout the world; itoccurs eitheras aresult of superin-fection of the hepatitis B virus carrier state or as an acute
coinfection, and delta virus infection is known to cause severe liver damage (24). Approximately, 1% ofJapanese
hepatitisB viruscarrierswereinfectedwith deltaagent(14).
However, there was no information on the nucleic acid
structureofdeltaagentderived from Japanese patients. Recently, complete nucleotide sequences of delta virus
RNAwerereported from three laboratories. Wangetal. (27, 28) analyzed an isolate from the serumofa chimpanzee to
which the serumofanItalian patient wastransmitted, Kuo
etal.(11)examinedanisolate from the liver ofawoodchuck towhich theserumofthesamepatientwastransmitted, and Makino et al. (13) analyzed an isolate directly from the serum ofan Americanpatient. In thisreport, partial RNA
sequences 868nucleotides long, includingthe delta antigen-coding regions of isolates fromtwoJapanese patients,were determined to investigate the geographical divergence of viral RNA, and viral isolates obtained 3 and 4 years apart from each oftwopatients were also examined tostudy the evolution of delta virus RNA.
MATERIALS AND METHODS
Virus isolates. Two HDV isolates were obtained on two differentoccasions from Japanese patient M, a 25-year-old malewithchronic active hepatitis.Japanese M-1was avirus
isolate obtained on 20 September 1986, and Japanese M-2 wasobtained on 6 July 1989. Similarly, two HDV isolates were obtained from Japanesepatient S, a 39-year-old male
withchronic active hepatitis. JapaneseS-1isa virus isolate
obtainedon 18July 1983, and Japanese S-2wasobtainedon 6August 1987. The seraof both patients were positive for
*Corresponding author.
hepatitis B surface antigen and hepatitis B eantibody, and the patients were born and grew up in Japan and had no
history ofbloodtransfusion,intravenousdruguse,or homo-sexualactivity.
Extraction of HDV RNA. Serumsamples (100,ul)obtained
from the patients were incubated into a 400-,u solution
containing 10 mM Tris hydrochloride (pH 7.5), 10 mM
EDTA,0.2% sodiumdodecyl sulfate,and 1mgofproteinase
K per ml at 50°C for 3 h. HDV RNA was then extracted twice with 1 volume of phenol-chloroform, followed by
ethanol precipitation in the presence of salt and carrier dextran.
cDNAsynthesisand PCR. Samples (30to50
[L)
ofserum weredenaturedat65°Cfor 3minandreversetranscribed in 20,ul ofasolutioncontaining50 mMTrishydrochloride (pH8.3),100 mMKCl,10 mMMgCl2,10 mMdithiothreitol,50 U of human placental RNase inhibitor (Takara Shuzo Co., Kyoto, Japan), 1 mM deoxynucleoside triphosphate, 1 ,uM
antisense primer,and 25 U ofreverse transcriptase(Takara ShuzoCo.)at42°Cfor 60min. cDNAproductswereusedas
templates for polymerase chain reactions (PCR) (22). PCR wereperformedin 100 pl ofasolution containing 10mM Tris
hydrochloride (pH 8.4), 50 mMKCl,2.5 mMMgCl2,0.2mg
ofgelatinper ml, 1 ,uM senseprimer, and 4U ofTaqDNA
polymerase (New England BioLabs, Inc., Beverly, Mass.)
overlaid with 100 pul of mineral oil withan automatic
ther-mocycler.The thermalprofileinvolved35cycles of denatur-ationat95°Cfor 2min, primer annealingat42°C for2min,
and extension at 70°C for 2 min. Oligonucleotide primers
were synthesized by a DNA synthesizer (380B; Applied
Biosystems, Inc., Foster, Calif.) and purified with high-performance liquid chromatography. Synthetic primersused for cDNAsynthesisandPCRarelisted in Table 1.Theyare basedonthe nucleotidesequenceinformation ofWangetal.
(27, 28) or sequencing data obtained with isolates from
patients M and S. PCR products were electrophoresed on 8%polyacrylamide gel, andthegelwas stained with
ethid-ium bromide,followedby confirmationthat thebandwasof the estimated size.
Directsequencing.PCRproductsweredirectly sequenced bidirectionally by using the appropriate 32P-end-labeled
se-quencing primerand thedideoxy-chain termination method
5594 0022-538X/90/115594-06$02.00/0
Copyright © 1990,American Society for Microbiology
on November 10, 2019 by guest
http://jvi.asm.org/
TABLE 1. Syntheticoligonucleotides used for cDNA synthesis and PCR'
* ~~~~~~~~~~~~~~Nucleotide
Primer Sequence position
positions DELP4 5'-CGGGGGCGGCTTCGTCCCCA-3' 1110-1091
DELP7 5'-TAATGGCGAATGGGAC-3' 754-769
DELP25 5'-GAGGTTGACCGAGGAAGACG-3' 1206-1187 DELP3 5'-GGTCGACAACTCTGGGGAGA-3' 961-980 DELP24 5'-CCGGACCTAGGAAGAGGC-3' 1351-1334 DELP30 5'-TTGGGGACGAAGCCGCC-3' 1090-1107 DELP11 5'-AGCAGTCTCCTCTTTACAGA-3' 1657-1638 DELP9 5'-TCTTGTTCTCGAGGGCCTTC-3' 1265-1284 DELP37 5'-AAGGAAAAGAAGAGTAGCCG-3' 1182-1163 DELP34 5'-TGGGTCCCCCTGATGTCCAG-3' 1024-1043
"Thenotationis that ofWangetal.(27,28).Thesynthetic oligonucleotides for DELP4, DELP7, DELP25,DELP3,DELP24, DELP9, and DELP11are based onthe information ofWang et al. (27, 28), andthose for DELP30,
DELP37, and DELP34 are based on data obtained from Japanese patients M andS.
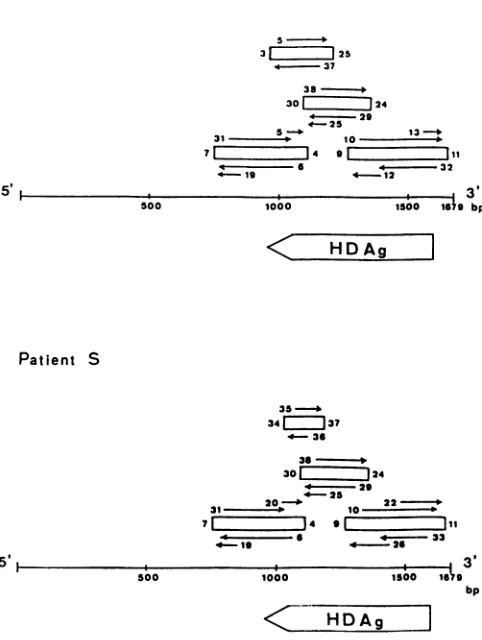
Patient M
5
-31 125
4.~. 37
38
301[ 124
*~. 29
e-25
31 -5 ---13
--71 1~4 9
< - 6 *4 32
19 4-12
5'
500 1000 --4---'1500 169 bp3'
<~ HDAg
Patient S
afterpurificationwith a Centricon 30 (Amicon Div. of W. R.
Grace & Co., Danvers, Mass.) (25). The oligonucleotide
primers used for sequencing are listed in Table 2, and the
strategies used for PCR and sequencing of the HDV
ge-nomes ofpatients M and S are shown in Fig. 1.
Analysis ofsequence identity. HDV RNA sequences ob-tained with isolates from the Japanese patients were com-paredwith those reported previously and analyzed with the
SDC Genetyx system (Software Development Co., Tokyo,
Japan).
Nucleotide sequence accession numbers. The nucleotide
sequence data reported here will appear in the DDBJ,
EMBL, and GenBank nucleotide sequence data bases under
accession numbers D90190, D90191, D90192, and D90193.
RESULTS
cDNAsequence of HDV RNA and comparison of sequence
identities. Eight hundred sixty-eight nucleotides, from
posi-TABLE 2. Syntheticoligonucleotides used for directsequencing"
Nucleotide
Primer Sequence poiton
positions DELP5 5'-TCGGCTGGGAAGAGTATATC-3' 991-1010 DELP6 5'-AGTCCCGGAGTCCCCCTTC-3' 1080-1062 DELP10 5'-TTCGTCGGTGATCCTGCCTC-3' 1286-1305 DELP12 5'-AGAAGGATAAGGATGGAGAG-3' 1420-1401 DELP13 5'-TCTAACTTCTTTCTTCCG-3' 1516-1533 DELP19 5'-CGCGGTCCGACCTGGGCATC-3' 874-855 DELP20 5'-GGAGTACACTCGAGGAGTGG-3' 961-980 DELP22 5'-TTCTTCCTCGAGTTTCTTGA-3' 1458-1477 DELP25 5'-GAGGTTGACCGAGGAAGACG-3' 1206-1187 DELP26 5'-AGGAAGAAAATCCCTGGCTG-3' 1465-1446 DELP29 5'-TTCACCGACAAGGAGAGG-3' 1319-1302 DELP31 5'-CAGAACTCTCTCTAGATTCC-3' 771-790 DELP32 5'-AAAGAGTAAGAGTACTGAGG-3' 1637-1618 DELP33 5'-AAAGAATAGAGAGAACTGAG-3' 1637-1618 DELP35 5'-CTCCCCCGTCCGAGAGAAGG-3' 1047-1066 DELP36 5'-CCCGCGGGTTGGGGATGTGA-3' 1161-1142 DELP38 5'-GGTTCACATCCCCAACC-3' 1139-1155
aThe notation is that ofWangetal.(27,28).Thesyntheticoligonucleotides for DELP5, DELP6, DELP10, DELP12, DELP13, DELP19, DELP20, DELP25, DELP29,and DELP31 are based on the information ofWangetal.
(27, 28), and those for DELP22, DELP26, DELP32, DELP33, DELP35, DELP36,andDELP38 are based on data obtained fromJapanesepatientsM andS.
35
-34 E 37
-4-- 36
36-4
301 124
4-~29
4--25
20 -. 2
4-.-6 4.. 33
4-l . we
5'
S00 1000 1300 I6>9
bp
FIG. 1. Strategies used for PCRandsequencing of the genomes of HDV isolates from patients M and S shown relative to the 1679-nucleotide full-length HDV genome and delta antigen-coding frame. The notation is that of Wang et al. (27, 28). The open rectangles indicatecDNA segmentsamplified by PCR. The numbers onboth sides of the rectangles indicate the synthetic oligonucleo-tidesusedforPCR (Table 1). The arrows indicatetheorientations and regions sequenced. The numbers by the arrows indicate the syntheticprimers used for direct sequencing (Table 2). HDAg, HDV antigen; bp,basepairs.
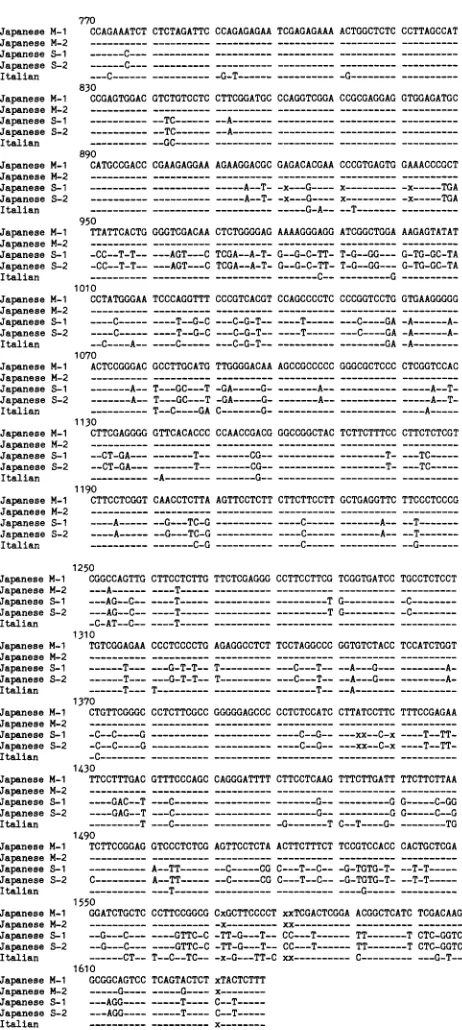
tions770 to1637,asdesignated by Wang et al. (27, 28),were
determinedinJapanesevirusisolatesM-1, M-2,S-1,andS-2 (Fig. 2). Theopen readingframe of deltaantigen, the only knownprotein of HDV, is included in this region. ThecDNA sequence of HDV RNA from patient S showed not only single-base changes but six-base deletions and four-base insertions from positions 770 to 1637 compared with the sequence reported by Wang et al. (27, 28), whereas only single-base changes werefound in HDV RNAfrom patient
M(Fig. 2).
Thepercent identities of nucleicacids in thisregionamong
five virus isolates are shown in Table 3. The highest nucle-otide identity (98.8%), as expected, wasobserved between two isolates from chimpanzee-adapted (27, 28) and
wood-chuck-adapted (11) HDV genomes from an Italian patient (Table 3). The cDNA sequenceidentitybetweenthe
Italian-derived (27, 28) and American-derived (13) HDV genomes
was89% in the entire region and 92.4% in the regionfrom
positions 770 to 1637. Similar degrees of cDNA sequence identity (90.8 to92.3%)wereobserved between HDVRNA frompatientM and HDV RNAs derived from the Italianand American patients (Table 3). In contrast, the cDNA
on November 10, 2019 by guest
http://jvi.asm.org/
[image:2.612.54.294.96.215.2] [image:2.612.51.294.495.673.2]770
Japanese M-1 CCAGAAATCTCTCTAGATTC CCAGAGAGAA TCGAGAGAAAACTGGCTCTC CCTTAGCCAT Japanese M-2 --- ---JapaneseS-1 ---C---.--__
---Japanese S-2 ---C---.-.---__-
_--_---Italian ---C---G-T---G---
---830
JapaneseM-1 CCGAGTGGACGTCTGTCCTC CTTCGGATGC CCAGGTCGGA CCGCGAGGAG GTGGAGATGC
JapaneseM-2 ---
---JapaneseS-1 ---TC--- --A---
---Japanese S-2 ---TC---A--- ---Italian ---GC---
---890
JapaneseM-1 CATGCCGACC CGAAGAGGAA AGAAGGACGC GAGACACGAACCCGTGAGTG GAAACCCGCT JapaneseM-2 --- --- ---JapaneseS-1 ---A--T--x---G---- x---x---TGA
Japanese S-2 --- ---A--T--x---G----x---x---TGA
Italian ---G-A--
--T---950
Japanese M-1 TTATTCACTGGGGTCGACAA CTCTGGGGAG AAAAGGGAGG ATCGGCTGGA AAGAGTATAT JapaneseM-2 --- ---
---JapaneseS-1 -CC--T-T---AGT---C TCGA--A-T- G--G-C-TT- T-G--GG--- G-TG-GC-TA
JapaneseS-2 -CC--T-T-- ---AGT---C TCGA--A-T-G--G-C-TT-T-G--GG---G-TG-GC-TA
Italian ---C---G
---1010
JapaneseM-1 CCTATGGGAA TCCCAGGTTTCCCGTCACGT CCAGCCCCTCCCCGGTCCTG GTGAAGGGGG Japanese M-2 ---
---Japanese S-1 ----C--- ----T--G-C ---C-G-T-- ----T--- ---C----GA -A---A-Japanese S-2 ----C--- ----T--G-C---C-G-T---T--- ---C----GA -A---A-Italian --C----A-- ----C--- ---C-G-T---GA
-A---1070
Japanese M-1 ACTCCGGGACGCCTTGCATG TTGGGGACAAAGCCGCCCCC GGGCGCTCCC CTCGGTCCAC JapaneseM-2 --- ---JapaneseS-1 ---A-- T---GC---T-GA---G---A-- --- ---A--T-Japanese S-2 ---A--T---GC---T-GA---G---A---
---A--T-Italian ---T--C----GA C---A---
----A---1130
Japanese M-1 CTTCGAGGGG GTTCACACCCCCAACCGACG GGCCGGCTACTCTTCTTTCC CTTCTCTCGT Japanese M-2 ---
---Japanese S-1 --CT-GA---T-- ---CG---T- ---TC---JapaneseS-2 --CT-GA---T---CG---T- ---TC---Italian --- -A---G---
---_-1190
JapaneseM-1 CTTCCTCGGT CAACCTCTTA AGTTCCTCTTCTTCTTCCTTGCTGAGGTTC TTCCCTCCCG
Japanese M-2 ---__
-Japanese S-1 ----A---G---TC-G---C--- ---A--
--T---JapaneseS-2 ----A--- --G---TC-G---C--- ---.A- --T---Italian ---C-G---C---
--G---1250
Japanese M-1 CGGCCAGTTG CTTCCTCTTGTTCTCGAGGGCCTTCCTTCGTCGGTGATCC TGCCTCTCCT JapaneseM-2 ---A---T--- ---Japanese S-1 ---AG--C-- ----T---T G--- -C---JapaneseS-2 ---AG--C-- ----T---TG---
-C---Italian -C-AT--C-- ----T--- ---
---1310
JapaneseM-1 TGTCGGAGAACCCTCCCCTGAGAGGCCTCTTCCTAGGCCCGGTGTCTACCTCCATCTGGT
JapaneseM-2 --- ---JapaneseS-1 ---T--- ---G-T-T-- T--- ---C---T-- --A---G---
---A-JapaneseS-2 ---T--- ---G-T-T-- T--- ---C---T-- --A---G--- ---A-Italian ---T---T---T-- --A---
---1370
Japanese M-1 CTGTTCGGGC CCTCTTCGCCGGGGGAGCCCCCTCTCCATCCTTATCCTTC TTTCCGAGAA
Japanese M-2 --- ---JapaneseS-1 -C--C----G.---C--G-- ---xx--C-x
----T--TT-Japanese S-2 -C--C----G.---C--G-- ---xx--C-x
----T--TT-Italian -C.--- ---1430
Japanese M-1 TTCCTTTGACGTTTCCCAGC CAGGGATTTTCTTCCTCAAG TTTCTTGATT TTCTTCTTAA Japanese M-2 --- ---Japanese S-1 ----GAC--T ---C---G---GG---C-GG Japanese S-2 ----GAG--T ---C---G---GG---C--G
Italian ---T ---C---G---TC--T----G- ---TG 1490
JapaneseM-1 TCTTCCGGAGGTCCCTCTCG AGTTCCTCTA ACTTCTTTCTTCCGTCCACC CACTGCTCGA JapaneseM-2 --- ---JapaneseS-1 --- A--TT--- --C---CGC---T--C---G-TGTG-T- --T-T---Japanese S-2 C---A--TT---C---CGC---T--C---G-TGTG-T- --T-T---Italian ---T---G---
---1550
JapaneseM-1 GGATCTGCTCCCTTCCGGCG CxGCTTCCCCT xxTCGACTCGGAACGGCTCATC TCGACAAGAG
JapaneseM-2 ---x---xx--- ---Japanese S-1 --G---C---GTTC-C-TT-G---T--CC---T---TT---T CTC-GGTCG-Japanese S-2 --G---C---GTTC-C -TT-G---T--CC---T--- TT---T CTC-GGTCG-Italian ---CT--T--C--TC-- -x-G---TT-Cxx---C---
---G-T----1610
Japanese M-1 GCGGCAGTCCTCAGTACTCTxTACTCTTT
JapaneseM-2 ---G---G----x
Japanese S-1 ---AG .---T----
C--T---Japanese S-2 ---AGG---T---- C--T---Italian ---
x---FIG. 2. Nucleic acid sequence alignment of five virus isolates
from two Japanese patients and an Italian patient (27, 28) at
nucleotide positions 770 to 1637. An x indicates deletion of the
sequence. The designations are thoseof Wangetal. (27, 28). The
Japanese M-1 isolate obtained from patient M was isolated on20
September 1986; the Japanese M-2 isolate obtained from patientM
wasisolatedon6July1989;the Japanese S-1isolate obtained from
patient S was isolated on 18 July 1983; the Japanese S-2 isolate
obtained from Japanesepatient Swasisolatedon6August 1987; the
[image:3.612.70.301.87.602.2]Italianisolate obtained fromanItalianpatientwasreported byWang etal.(27, 28).
TABLE 3. Comparisonofnucleicacidsequenceidentity inthe region between positions 770and 1637
%Identitywith:
Isolate Italian Italian Japanese
(chimpanzee)' (woodchuck)b American' M_jd Italian (wood- 98.8 100
chuck)b
Americanc 92.4 92.2 100
JapaneseM_1d 92.3 92.2 90.8 100
JapaneseS-V' 81.4 81.7 81.0 80.2
" Chimpanzee-adapted HDVgenomefrom anItalianpatient,reported by
Wang et al. (27, 28).
bWoodchuck-adapted HDV genome from the same Italian patient,
re-ported byKuo etal.(11).
' Obtainedfrom anAmerican patient,reported byMakino et al.(13). dObtained fromJapanesepatientM(virus isolatedon 20September 1986).
eObtainedfromJapanesepatient S (virus isolatedon 18July 1983).
quences of isolates from patient S showed rather weak
identities (80.2 to 81.7%) with the other isolates (Table 3). Deltaantigenwascomposed of 214 amino acids (aa) in the reports of Wang et al. (27, 28) and Makino et al. (13). However, the delta antigen of Japanese patients M and S
was supposed tobe 195 aa because of a G-to-Asubstitution
atposition 1012,asin the reportfromKuo etal. (11)(Fig. 2).
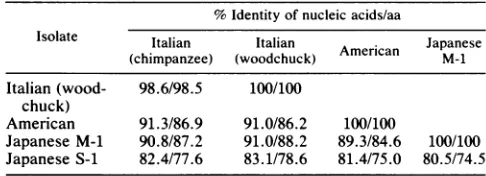
A comparison of nucleic acid and aa sequence identities
amongdeltaantigen-codingframesof HDV isolates is shown in Table 4. The percent identities of the aa of deltaantigen predicted by the nucleic acid sequences were 85 to 88%
among isolates from the Italian patient, the American pa-tient, and JapanesepatientM(Table 4). When the aaof delta
antigenderived from Japanesepatient S were compared with thosefrom the Italianpatient(27, 28), the Americanpatient
(13), and Japanese patient M, there was only 75 to 79%
sequenceidentity.
Evolution of HDV RNA. The sequences of two virus
isolates obtained 3 and4years apartfrom patients MandS,
respectively,werecompared. Of 868 nucleotides, therewere
substitutions at positions 1253, 1264, 1615, and 1625 in the isolates frompatientMandpositions 1436, 1488, and 1490 in theisolates from patient S (Fig. 2). Two of four nucleotide substitutions in the isolatesfrom patient Mand all three in theisolatesfrom patient S occurred within the delta
antigen-coding region,which is 585 nucleotides long. The
substitu-tion rate of HDV RNA for the delta antigen gene was
calculated(5)as0.57x 10-3nucleotides per site per year for
patientMand 0.64 x 10-3 nucleotides persite per year for
patient S. Thefour-of-five nucleic acid substitutions in the deltaantigen-codingregion occurred at position 1 or 2ofa
codon, and this resulted infour aminoacidchanges.
DISCUSSION
Five groups have reported the molecular cloning and
sequencing of either partor all of the HDV genome (9, 11, 13, 23, 27, 28). Three of them determined the complete nucleotide sequence of thevirus and, hence, determined the open reading frame of delta antigen (11, 13, 27, 28). We obtained partial sequences of delta virus RNAs from two Japanesepatients,including the delta antigen-coding region, and compared them with the previous reports (11, 13, 27, 28). The percent identities of the nucleic acids from positions 770to1637were91to92%amongthe three deltavirus RNA sequences,namely, thatofanisolate from an Italian patient reported by Wanget al. (27, 28), that of an isolate from an
on November 10, 2019 by guest
http://jvi.asm.org/
[image:3.612.324.565.94.186.2]TABLE 4. Comparison of nucleic acid and aa sequence identities in the delta antigen-coding region'
%Identity of nucleic acids/aa
Isolate Italian Italian A Japanese
(chimpanzee) (woodchuck) American M-1
Italian (wood- 98.6/98.5 100/100 chuck)
American 91.3/86.9 91.0/86.2 100/100
Japanese M-1 90.8/87.2 91.0/88.2 89.3/84.6 100/100 Japanese S-1 82.4/77.6 83.1/78.6 81.4/75.0 80.5/74.5
aNucleotides954 to1598in the American andchimpanzee-adaptedItalian
isolatesand nucleotides 1011 to 1598inthe woodchuck-adapted Italian isolate
and theJapanese M-1andS-1 isolateswerecompared because ofa stop codon atnucleotides1011 to 1013.
American patient reported by Makino et al. (13), and that of an isolate from Japanese patient M. There was only 80% identity of nucleic acid sequences between Japanese patient S and the other three patients. Thus, the nucleic acid sequenceobtained from patient S was the most divergent of those studied. Several subtypes of delta virus may exist in
the world, and HDV in Japan may have a heterogeneous
origin.
When the aa and nucleic acid sequences in the delta
antigen-codingregions of five virus isolates were compared, the percent identities of aa were lower than those of nucleic
acids (Table 4). The nucleic acid substitutions in the delta
antigen-codingregion occurred more frequently at positions 1and 2 of a codon than at position 3. Nucleotide
substitu-tions atposition 1 or 2usually change the codingat the aa level, while nucleotide substitutions at position 3 usually do not.Ingeneral,coding at the aa level is more conserved than
atthenucleic acid level in RNA viruses like human influenza A virus (1), hepatitis C virus (10), and retroviruses (5). In
these viruses, most of the nucleotide substitutions within a
subtype occur at position 3 in a codon. HDV may be a
uniqueRNAvirus in this regard.
Therateofevolution of HDVwasestimated to bearound 10-3 nucleotides per site per year, and this value is almost thesame asthatof the nonstructural (NS) gene of influenza Avirus(3) and the envelope gene of human T-cell
lympho-tropicvirus type III(6); 10to
102-fold
higher than that of the surface, core, X, and polymerase genes of hepatitis B virus (16);and106-fold
higher than that ofmostDNA genomes(5, 7). This highevolutionrate of HDVseems to bepartly duetolack of theproofreading enzymes like otherRNAviruses (7). The HDV genome may existas amixture of viruses with
many microheterogeneities of the nucleotide sequence in
oneindividual. The nucleotide sequence reported byWang
et al. actually contains an ambiguity arising from clonal
heterogeneity of single nucleotides at 16 different positions (27, 28), and such aresult makes the calculations of
evolu-tion rates, such as thoseoffered here, seem inappropriate.
However,weusedamethodof directsequencingafter PCR,
instead of ordinarycDNA cloning,toestimatetheevolution
rateofthe HDVgenomeonthebasis oftwopairs of isolates. Although these results apply only to changes in which the averagevalueofthenucleotideat apositionwaschanged,it is possible to determine the nucleotide sequence ofa virus genomewhich is dominant inone individual and, hence, to
know its transition based on two pairs of virus isolates. In
previousreports of estimates of the evolutionrates of virus
genomes, nucleotide sequences of the NS gene of human
influenza A viruses were determined by direct RNA
se-quencingofpurified viral RNA (3)or nucleotide sequences
of sequentially isolated human T-cell lymphotropic virus type III in one individual were determined by molecular
cloning and only one clone for each virus isolate was
analyzed and compared (6). Because virus genomes may
exist as a heterogeneous population in one individual, it
seemsimportanttodetermine the nucleotide sequence of the virus isolate which isdominant,eitherby sequencingmany clones after ordinary
cloning
orby directsequencing
afterPCR,as wedid,tocalculate and estimate the evolutionrates
of genomes.
Recently, Luo et al. reported that RNA duplex
unwin-dase,which deaminatesadenosineandconvertsittoinosine, mightbe involved innucleic acid substitutions in the HDV genome(12). Theeffect isreplacement ofAwith G on one
strand and T with C on the complementary strand. They
found thatAwasreplaced byGat
position
1012,which leadsto translation of the large 214-aa delta antigen,
during
replication
ofthe HDV genome. Inourstudy,
4and 3of868nucleotides were substituted in
patient
M over a 3-yearperiod and in patient S over a 4-year
period, respectively.
Conversion ofA to G orT toC was found in two of four nucleotidechangesinpatientMandoneof three inpatientS. This value, three of seven, is
higher
than thatexpected,
although conversion of A to G at
position
1012 was notdetected in eitherpatient. Theseresults suggest that unwin-daseplays some importantrole innucleic acid substitutions of the HDV genome in
humans,
but not all of thechanges
followedtheproposed
rule. Morestudy
isneeded,
becauseaminorpopulation of virus genomesmight be missed
by
ourmethod ofdirect sequencingafterPCR.
Analysis ofHDV RNA sequences showed 5 to 11 open
readingframeslargerthan 100aawithin HDV RNA and the
complementary anti-HDV RNA strand(11, 13, 27,
28).
Thedelta
antigen
is theonly viralprotein
knowntobeexpressed
during HDV infection in vivo and encoded from theantige-nomic strand ofHDV. Wang et al. (27, 28) predicted two
delta antigen coding frame
species
214 and 195 aalong
because of theambiguity arisingfrom clonal
heterogeneity.
Makinoetal.(13)andKuoetal. (11)
predicted only
the214-and 195-aa
species,
respectively.
In ourstudy,
only
the 195-aa deltaantigen-coding
framespecies
waspredicted
inHDVisolates from bothJapanese
patients.
HDV is consideredtobe
directly
cytopathic
(18),but the mechanism ofpathogenesis
remains to be elucidated.Re-cently, Negro et al. found that extensive
complementarity
between human 7SL RNA and
antigenomic
HDV RNAexists andthey
presented
thehypothesis
thatanantigenomic
form of HDV may anneal to the human 7SL RNA and
formation of
hybrids
maybeafactor in disease(15).
Indeed,
nucleotides 858 to 899 in HDV sequences from Japanese
patients
M and S were very well conserved(in fact,
100%matched in this 42-nucleotide segment;
Fig. 2),
like those fromanItalianpatient
andanAmericanpatient (13, 27, 28).
HDV has some characteristic structures in common with
viroid or virusoid (27). The virusoid consensus sequence,
GATTTT,
which may beimportant
forreplication
ofviru-soids,
exists in HDV from Japanesepatients
M and S atnucleotides 1454 to 1459, as
pointed
out in HDV isolates froman Italianpatient
andanAmericanpatient (13, 27,
28).
Wanget al. determined the
immunogenic
domains of HDVantigen by using
thesynthetic hexapeptides
spanning
the entire214-aaresiduesof theprotein (26). They
reported
that the domainsrecognized by
antibodies present in sera fromhumanchronic carriers of HDV included residues 2to
7,
63 to74, 86to 91,94to 100, 159to172,
174to195,
and 197to 207. When theseregions
werecompared
among five viruson November 10, 2019 by guest
http://jvi.asm.org/
Italian1 Italian2
American
JapaneseM-1
Japanese S-1
Italian Italian
American
Japanese M-1
Japanese S-1
Italian Italian American
JapaneseM-1
Japanese S-1
10 20 30 40 50 60
MSRSESRKxNRGGREEILEQW VAGRKKLEELERDLRKTKKK LKKIEDENPWLGNIKGILGK ---R--xD- ----D--- -S--- ---L--- I--L-ED--- ---I---KGxk- A---Q--- -D--- ---I--- I--L-E---- ---V---Q--T-RGR- -T---T--K- ITA---A--- -K---R-T I--L-E---- ----V--IxR 70 80 90 100 110 120 KDKDGEGAPPAKRARTDQME VDSGPRKRPL RGGFTDKERQ DHRRRKALEN KKKQLSAGGK ---KL-M---- I-A--- --- -R----S---.-.---.T---S--- --- -R---A---G--- ---P--- ---G---H KS---E--- ---130 140 150 160 170 180 NLSKEEEEEL RRLTEEDERRERRVAGPPVG GVIPLEGGSR GAPGGGFVPS LQGVPESPFS
--- --- --N--- ---N
---S--R.---K---K- ---I---S--_-N--- ---M---A --- K---S-- --N---P- ---N M---T I---D---E-K---KR-- D-N-SR--P- ---Q
MA---190 200 210 Italian RTGEGLDIRG NRGFPWDILF PADPPFSPQS CRPQ Italian ---
-Q---American --- SQ.--- ----Japanese M-1 ---VT-
-L---Japanese S-1 ---
TQ---FIG. 3. Alignment of theaasequencesof the deltaantigengenes of five virus isolates. The single-letter aa code is used. An x
indicates deletion of thesequence.The Italian'sequenceis that ofa
chimpanzee-adapted HDVgenomefromanItalianpatient, reported by Wangetal. (27,28); the Italian2sequenceis thatofa woodchuck-adapted HDV genomefrom the same Italianpatient, reported by Kuoetal.(11);the Americansequence wasreportedby Makinoet al.(13); the Japanese M-1sequenceobtained fromJapanese patient M wasisolated on20 September 1986; the JapaneseS-1 sequence obtained fromJapanese patient Swasisolatedon18July1983.
isolates, residues 63to72, 160to 169,174to 179,and 181 to 187were completely conserved (Fig. 3). These results sug-gest that sequences in these functional domains were well conserved in each of the virus isolates.
We used the PCR technique instead of ordinary cDNA
cloningtoexaminethevariationinHDV andtoestimate the evolution rates ofgenomes. This method saves time, and very small amounts of infected serum are needed to
deter-mine the amplified cDNA sequences (about 30 to 50 ,ul of serum per PCR in our study). The nucleotide substitutions
shown in our study might be due to an artifact of PCR
amplification (8, 22), but they were reproducible and were confirmed by bidirectional sequencing. This method is also usefulfordiagnosisof delta virus infectionbycDNA ampli-ficationby using synthetic primerslocated inveryconserved
regions of HDV sequences.
In conclusion, the sequence obtained for hepatitis delta virus RNAfrompatientMwasapproximately92% identical
to sequences previously obtained for two other strains of
hepatitis deltavirus, whereas thesequenceofhepatitisdelta
virusRNAobtained from patient Swasapproximately 81%
identicaltothepreviously sequenced strains and 80% iden-tical to the sequence obtained from patient M. These data suggestthatdeltaagentinJapan hasaheterogeneous origin. The delta virus from patient S may be adifferent strain of deltaagent. Complete sequencing of delta virus RNA from
our patients and comparison with other sequences derived
from different areasof the world may elucidate the signifi-cance of theobserved heterogeneity.
ACKNOWLEDGMENTS
Thisstudywassupported inpartbyJapaneseMinistry of Educa-tiongrants(B)58480215, (C)60570310, and(I)59010029.
LITERATURE CITED
1. Air, G. M. 1981. Sequence relationships among the hemagglu-tinin genes of 12 subtypes of influenza A virus. Proc. Natl. Acad. Sci. USA 78:7639-7643.
2. Bonino,F., B. Hoyer, J. W.-K. Shih, M. Rizzetto, R. H. Purcell,
and J. L. Gerin. 1984. Delta hepatitis agent: structural and antigenic properties of the delta-associated particle. Infect. Immun.43:1000-1005.
3. Buonagurio, D. A., S. Nakada, J. D. Parvin, M. Krystal, P. Palese, and W. M. Fitch. 1986. Evolution of humaninfluenzaA viruses over 50 years: rapid, uniform rate of change inNSgene. Science 232:980-982.
4. Chen, P.-J., G. Kalpana, J. Goldberg, W. Mason, B. Werner,J. Gerin, and J. Taylor. 1986. Structure and replication of the genomeof thehepatitisdelta virus. Proc. Natl.Acad. Sci. USA 83:8774-8778.
5. Gojobori, T., and S. Yokoyama. 1985. Ratesofevolution of the retroviral oncogeneof Moloney murine sarcoma virusand of its cellularhomologues.Proc.Natl.Acad. Sci. USA 82:4198-4201. 6. Hahn, B. H., G. M. Shaw, M. E. Taylor, R. R. Redfield, P. D. Markham, S. Z. Salahuddin, F. Wong-Staal, R. C. Gallo, E.S. Parks, and W. P. Parks. 1986. Genetic variation inHTLV-III/ LAV over time in patients with AIDS or at risk for AIDS. Science 232:1548-1553.
7. Holland, J., K. Spindler, F. Horodyski, E. Graban, S. Nichol, and S. VandePol. 1982. Rapid evolution of RNA genomes. Science215:1577-1585.
8. Keohavong, P., and W. G. Thilly. 1989. Fidelity of DNA polymerases in DNAamplification. Proc. Natl.Acad.Sci.USA 86:9253-9257.
9. Kos, A., R. Dijkema, A. C. Arnberg, P. H.vanderMeide,and H. Schellekens. 1986. The HDV possesses a circular RNA. Nature (London) 323:558-560.
10. Kubo, Y., K. Takeuchi, S. Boonmar, T. Katayama, Q.-L.Choo, G. Kuo, A. J. Weiner, D. W. Bradley, M.Houghton, I.Saito, and T. Miyamura. 1989. A cDNA fragment of hepatitis C virus isolated from an implicated donorofpost-transfusion non-A, non-Bhepatitis in Japan.Nucleic Acids Res.17:10367-10372. 11. Kuo, M. Y. P., J. Goldberg, L. Coates, W. Mason,J. Gerin, and
J.Taylor. 1988. Molecularcloningofhepatitisdelta virus RNA from an infected woodchuck liver: sequence, structure, and applications. J. Virol. 62:1855-1861.
12. Luo, G., M. Chao, S.-Y. Hsieh, C.Sureau,K.Nishikura, and J. Taylor. 1990. A specific base transition occurs on replicating hepatitis delta virus RNA. J. Virol. 64:1021-1027.
13. Makino, S., M. Chang, C. Shieh, T. Kamahora, D. Vannier, S. Govindarajan, and M. Lai. 1987. Molecular cloning and
se-quencingof a human hepatitis delta virus RNA. Nature (Lon-don) 329:343-346.
14. Mori, J., Y. Ito, M. Omata, 0. Yokosuka, and K.Okuda. 1986. Deltainfectionin Japan: immunohistological and immuno-elec-tron microscopic study of delta antigen in liver tissue. J. Gastroenterol. Hepatol. 1:33-38.
15. Negro, F., J. L. Gerin,R. H. Purcell, and R. H. Miller. 1989. Basis ofhepatitis delta virus disease? Nature(London)341:111. 16. Okamoto, H., M. Imai, M. Kametani, T. Nakamura, and M. Mayumi. 1987. Genomicheterogeneity ofhepatitis Bvirusina
54-year-old woman who contracted the infection through materno-fetal transmission. Jpn. J. Exp. Med. 57:231-236. 17. Ponzetto, A., P. J. Cote, H. Popper, B. H. Hoyer, W. T.London,
E. C. Ford, F. Bonino, R. H. Purcell, and J. L. Gerin. 1984. Transmission of the hepatitis B associated delta agent to the easternwoodchuck. Proc. Natl. Acad. Sci. USA81:2208-2212. 18. Popper, H. 1984. Pathological observations on delta agent infection, p. 381-384. In G. N. Vyas, J. L. Dienstag, and J. H. Hoofnagle (ed.), Viral hepatitis and liver disease. Grune & Stratton, Orlando, Fla.
19. Rizzetto, M. 1983. Thedelta agent. Hepatology 3:729-737. 20. Rizzetto, M., M. G. Canese, J. Arico, 0. Crivelli, F. Bonino,
C. G. Trepo, and G. Verme. 1977. Immunofluorescence detec-tion of a newantigen-antibody system(delta/anti-delta) associ-ated to the hepatitis B virus in the liver and in the serum of HBsAgcarriers. Gut 18:997-1003.
21. Rizzetto, M., B. Hoyer, M. G. Canese, J. W.-K. Shih, R. H.
Purcell, and J.L.Gerin. 1980. Delta agent: association ofdelta agent with hepatitis B surface antigen and RNA in serum of delta-infected chimpanzees. Proc. Natl. Acad. Sci. USA 77: 6124-6128.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:5.612.64.308.72.229.2]22. Saiki, R. K., D. H. Gelfand, S.Stoffel, S. J. Scharf,R.Higuchi, G. T. Horn, K. B. Mullis, and H. A. Erlich. 1988. Primer-directed enzymaticamplification of DNA withathermostable DNApolymerase. Science 239:487-491.
23. Saldanha, J. A., H. C. Thomas, and J. P. Monjardino. 1987. Cloning and characterization ofadelta virus cDNAsequence derived fromahumansource.J. Med. Virol. 22:323-331. 24. Smedile, A., P. Farci, and G. Verme. 1982. Influence of delta
infectiononseverity ofhepatitis B. Lancet ii:945-947. 25. Tada,M., M.Omata,and M.Ohto. 1990. Analysis ofras gene
mutations in hepatic malignant tumors by polymerase chain reaction anddirect sequencing. Cancer Res. 50:1121-1124.
26. Wang, J.-G.,R. W. Jansen, E. A. Brown, and S. M. Lemon. 1990. Immunogenic domains of hepatitis delta virus antigen: peptide mapping ofepitopes recognized by human and wood-chuckantibodies. J. Virol. 64:1108-1116.
27. Wang, K.-S., Q.-L. Choo, A. J. Weiner, H.-J. Ou, R. C. Najarian, R. M. Thayer, G. T. Mullenbach, K. J. Denniston, J. L. Gerin,andM. Houghton. 1986. Structure, sequence and expressionof the HDVgenome. Nature(London)323:508-513. 28. Wang, K.-S., Q.-L. Choo, A. J. Weiner, H.-J. Ou, R. C. Najarian, R. M. Thayer, G. T. Mullenbach, K. J. Denniston, J. L. Gerin, andM. Houghton. 1987. Structure, sequence and expression of the HDVgenome. Nature(London) 328:456.