Vol.47, No. 3 JOURNALOFVIROLOGY,Sept.1983,p.586-599
0022-538X/83/090586-14$02.00/0
Copyright © 1983, American Society forMicrobiology
Polyomavirus Origin
for
DNA
Replication Comprises
Multiple
Genetic Elements
WILLIAM J. MULLER, CHRISTOPHERR. MUELLER, ANNE-MARIEMES,ANDJOHN A. HASSELL*
Departmentof Microbiology and Immunology, McGill University, Montreal, Quebec H3A 2B4, Canada
Received10March 1983/Accepted 7June1983
To define the minimal cis-acting sequences required for polyomavirus DNA replication (ori),weconstructed a number ofpolyomavirus-plasmid recombinants and measured theirreplicative capacity after transfection of apermissive mouse cell line capable of providing polyomavirus largeTantigen intrans(MOP cells). Recombinant plasmids containing a 251-base-pair fragment of noncoding viral DNA replicate efficiently in MOP cells. Mutational analyses of these viral sequences revealed that they can be physically separated into two genetic elements. One of these elements, termed the core, containsan adenine-thymine-rich area, a32-base-pair guanine-cytosine-richpalindrome, andalargeTantigen binding site, andlikely includesthesitefrom which bidirectionalDNAreplication initiates. The other, termed ,B,is locatedadjacenttothecore nearthe late region and isdevoid ofoutstanding sequencefeatures. Surprisingly, anothersequence element nameda,located adjacent to,Bbut outside the borders of the 251-base-pair fragment, can functionally substitute for 1. This sequence too contains no readily recognizedsequencefeatures andpossesses noobvioushomologytothe,B element. The three elements togetheroccupy acontiguous noncodingstretch of DNA no more than 345 base pairs in length in the order a, 1B, and core. These results indicate that the polyomavirus origin for DNA replication comprises multiplegenetic elements.
Polyomavirus, oneofthepapovaviruses, has served as a model to study mechanisms of transcription and DNA replication in mammali-ancells. Theutilityof thevirus derives from the small size of its genome and its almost total relianceon cellularproteins for replication. The elucidation of theprimary structure ofthe viral DNA,theidentification of the viralproteins,and the construction of physical maps of the viral mRNAs(9, 15, 22, 38) have heightenedinterest in the virus not onlyas atool to study cellular processes, but also as a vector for the introduc-tion and expression of foreign DNA into mam-malian cells(18, 32).Althoughmuchinformation has been accumulated about the coding se-quencesofthe virus and theproteins they speci-fy, much less is knownaboutregulatoryregions withintheviral DNA and theirinteraction with viral and cellular proteins. We have begun to define the sequences required in cis for viral DNA replication andtranscription as aprelude to help understand the mechanisms of gene expression, DNA replication, and their regula-tion in mammalian cells. Here we report our findings concerning those noncoding sequences required in cis for polyomavirus DNA replica-tion. Forconvenience ofexpressionwereferto
thesecis-actingsequences asori throughout the text.
Genetic and biochemical analyses of polyoma-virus DNA replication have revealed that effi-cient viralreplication requiresapermissive host cell, a viralgene product (large Tantigen), and cis-acting viral sequences. Polyomavirus DNA replicates efficiently in cells of murine origin, but poorly or not at all in cells derived from other species (6). Restricted replication of
polyomavirus DNAin nonmurine cells isdueto
the absenceofpermissive factors rather than the presence of inhibitors (1). The nature of these permissive factors has not yet been elucidated, although cell-cell hybridization studies suggest that they are encoded by the genome of the permissivecell(2). Anotheressentialingredient forefficient DNAreplication islarge Tantigen. Thisproteinisencodedby the viral genome and issynthesized early during infection before viral DNA replication commences. Genetic studies have revealed thatlargeTantigenisrequiredfor the initiation ofeach new round of viral DNA synthesis (8).The latterlikely requiresthe phys-ical interactionoflargeTantigen withori. Two largeTantigen binding siteshavebeenmapped on the viral genome (34), andone of these lies
586
on November 10, 2019 by guest
http://jvi.asm.org/
near sequences from which DNA replication initiates. Finally, the cis-acting sequences that comprise ori are required for the initiation of viral DNA replication. The boundaries of ori have not been precisely defined.However, stud-ies of evolutionaryvariants of polyomavirus (16, 24) and viable deletion mutants (3, 17, 27, 47) have shown that ori resides within noncoding sequences that include the site of initiation of DNA synthesis. To more precisely locate the limits of ori we have employed a strategy that measures thereplicational capacityof polyoma-virus recombinant plasmids in a line of mouse cells (MOP cells) that synthesize theviral large T antigen. MOP cell lines were established by the transformation of NIH 3T3 cells with a
hybridtranscription unit composed of the simian virus 40(SV40) early promoter and the polyoma-virus early region. These cells produce all of the polyomavirus early proteins, large, middle,and
small Tantigen,in apermissivecellular environ-ment (W. Muller, M. Naujokas, and J. A.
Has-sell, datanotshown). We measured the replica-tional capacity of a series of virus-plasmid recombinants and deletion derivatives in one such MOP cell line. Ouranalysesrevealed that oriiscomposedof threesetsofsequences. Two of these sequence elements, termed alpha (a) and beta (3), are functionally redundant and exert their effect in conjunction with a third
element called the core region. Although these elements lie closetogetherin the viralgenome, deletion analyses revealed that theyare physi-callydistinct.
MATERIALSANDMETHODS
Mammaliancells. MOP-8 cellsweregrownon plas-tic dishes, with Dulbecco modified Eagle medium (DMEM)supplemented with10o(vol/vol) calfserum
andantibiotics, and maintainedat37°C inahumidified
CO2atmosphere. Cells werepassaged by trypsiniza-tion after reaching confluence and were replated at
approximately104 cells percm2.
Preparation ofDNAandits modification.
Recombi-nantplasmid DNAs were isolated from bacteria and
purifiedby CsCl density gradientcentrifugation (19).
Digestionsof these DNAs with restriction
endonucle-ases were performed in accordance with the condi-tionsspecified by their manufacturers.
Reaction of DNA with the Klenow fragment of
DNApolymeraseI wasperformed byincubating1 ,g of DNA in a volume of 50 ,ul with 10 mM Tris-hydrochloride (pH 7.6), 5 mM MgCl2, 1 mM dithio-threitol, the appropriate deoxynucleotide triphos-phates (0.1 to 1.0 mM), and 5 U of the Klenow
fragmentof Escherichiacoli DNA polymeraseIfor1h at 15°C. The reaction was terminated by sequential phenol and chloroform-isoamyl alcohol (24:1, vol/vol) extraction, and the DNA was isolated by ethanol precipitation. Digestion of DNA with the Bal 31
nucle-ase and ligation with T4 ligase were carried out as describedpreviously (30).
Construction of recombinant plasmids. The recombi-nantplasmids that servedassubstrateformutagenesis
or replication were derived from molecules whose construction has been described previously (19, 30). We utilize the abbreviation pd (plasmid deleted) to
denote those recombinant molecules whose plasmid
sequencesarederived frompML-2 DNA(25).
Wher-ever possible we have tried to name recombinant plasmidsaccordingtotheircomposition.Forexample,
pPBR2 refers to plasmid (p), polyomavirus (P), BamHI (B)toEcoRI (R),isolate 2. The parentplasmid for all themutantsdescribed herewaspPBR2, which iscomposed of the2,220-base-pair(bp)polyomavirus
BamHI (nucleotide 4632)-to-EcoRI (nucleotide 1560) fragment and the large BamHI-to-EcoRI DNA
frag-mentof the plasmid pMK16.1 (30). pPBR2was modi-fiedby the insertion ofaHindIIIlinkerattheunique BglI site within polyomavirus sequences (nucleotide
87) to create the plasmid pPin7. This was
accom-plished by linearizing pPBR2 DNA with BglI,
remov-ing the 3-bp 3' projections with the exonucleolytic
activity of the Klenow fragment of E. coli DNA polymerase I, andligatingasyntheticHindIlllinkerto
theblunted ends. AfterHindlll cleavage, ligation, and transformation ofE. coli, the plasmid DNAs from individual colonies werescreened, andone that
con-tained a HindlIl site in place of the Bg[I site was
isolated and named pPin7. This mutant DNA was
sequencedacrossthe siteof insertion of the linkerby
thechemical method(28). A 3-bp stretch of viral DNA (nucleotides 91, 92, and 93) has been removed and replaced with 10 bp of linker DNA. Both pPBR2 and pPin7 served as substrates for the isolation of viral DNAfragmentsthatwerecloned withinpML-2 DNA. pdPBR2 was constructed by transfer of the
poly-omavirus sequences frompPBR2topML-2.Thiswas
accomplishedby cleavage of bothspecieswith BamHI andEcoRI andligationof the viral sequencesto the pML-2 vectorsequences. The resulting recombinant plasmid, pdPBR2, contains the viral sequences from nucleotide 4632to1560cloned between the EcoRI and BamHIsites ofpML-2DNA.
pdPBBg(H) was constructed by ligating the viral BamHI-to-HindIII fragment from pPin7 to pML-2 DNAdigested with thesesameenzymes.pdPBBg(H) contains viral sequences from nucleotide 4632to90.
pdPP1(B)Bg(H) was derived from pdPBBg(H) by cleavagewithBamHI and extension of the 3'-recessed termini byincubation with the Klenowfragmentof E.
coliDNApolymeraseIand thefourdeoxynucleotide triphosphates.These blunt-endedmoleculeswerethen
partially digested with PvuII to remove the viral sequence between nucleotide numbers 4632 (the
BamHIsite) and 5130 (aPvuIIsite). Circularization of the blunt ends by ligation with T4 ligase led to the isolation of pdPP1(B)Bg(H). This procedure resulted intheplacement ofaBamHIsite at nucleotide 5131 of
theviralinsert. Sequencing of the virus-plasmid junc-tions with pdPP1(B)Bg(H) confirmed that the viral sequences between nucleotides 5131 and 90 are con-tained within this recombinant as aBamHI-to-HindIII fragment.
pdPPl(B)R1wasconstructedby ligation oftheviral
HindIll-to-EcoRI sequences from pPin7 (the se-quencesbetween nucleotides 90 and 1560) between the
HindlIl and EcoRI sites of pdPP1(B)Bg(H) DNA. pdPP1(B)R contains those polyomavirus sequences
on November 10, 2019 by guest
http://jvi.asm.org/
588 MULLERETAL.
between nucleotides 5131 and 1560 and a 10-bp HindlIl linker between nucleotides90 and94.
pdPBP1wasderived from pdPBR2 by digestionwith
BamHI and extension of the 3'-recessed ends after
incubationwiththe Klenowfragmentof E. coli DNA
polymerase I and the fourdeoxynucleotide triphos-phates. Thesemoleculeswerethen cleaved withPvuII
to generate partial products, circularized with T4
ligase, andusedtotransform E. coli. Afterscreeninga
number of colonies, we identified several that
con-tained a recombinant plasmid from which viral
se-quences had been deleted between the BamHI site
(nucleotide 4632) and the PvuII site at nucleotide
position 5262. ABamHI site wascreatedatthenew
junctionbetween viral andplasmidsequencestoyield pdPBP1,which carries viralsequencesbetween nucle-otides 5265 (a BamHI site) and 1560(an EcoRI site) withinpML-2 DNA.
TherecombinantplasmidpdPBHpcontainsthe viral
BamHI (nucleotide 4632)-to-HphI (nucleotide 152)
DNA fragment between the pML-2 BamHI and HindIl sites. The BamHI site wasrestored, but the
HphI and HindlIl sites were destroyed during the construction ofpdPBHp. Inshort, pPBR2DNAwas digested withHphI, and the 3' projections were
re-moved with the Klenow fragment of E. coli DNA
polymeraseI.The DNAfragmentswerethenreacted with BamHI, and the fragment containing viral se-quencesenclosedbythe BamHIsite(nucleotide4632)
and the HphI site (nucleotide 152) was isolated by
agarosegel electrophoresis. This viral fragment was then ligated to pML-2 DNA that had been cleaved sequentially withHindIll and BamHI. Before cleav-agewithBamHI,the vectorDNAwasdigested with HindIII and the 3'-recessed ends were extended by
reaction with the Klenowfragment of E. coliDNA polymerase I and the fourdeoxynucleotide triphos-phates. Ligationof thevectorand viralsequencesto
each other resultedin the formation ofpdPBHp.The predicted sequences about the virus-plasmid joints
wereconfirmedby sequencingthem.
Isolation ofmutantrecombinant plasmids.The mu-tantseries thatcarryunidirectional deletionsfrom the
BamHI site in pdPBR2 DNAwere, with two
excep-tions, constructed by Bal 31 nuclease digestion of
BamHI-linearized pdPBR2DNA. After reaction with
thenuclease,asyntheticBamHIlinkerwasligatedto
theendsof themolecules,andtheywerethen cleaved with BamHI and EcoRI. The BamHI- and EcoRI-terminated viralsequenceswerecloned between these same sites in pML-2 DNA. Each mutant DNA was sequenced bythe chemical methodto determine the endpoint of the deletion (28). Two mutants in this series, pdPP1(B)R1 and pdPBP1 (see Fig. 1), were constructedasdescribed previously.
The mutantDNAs that bear deletions within viral
sequencesfrom theEcoRIsite inpdPBR2DNAwere constructedasdescribedpreviously,withone excep-tion. pdPBS1 wasisolated after cleavageofpdPBR2
DNA withSstIandEcoRI. The SstI 3'projectionsand theEcoRI 3'-recessedendswereremoved and
extend-ed with the Klenowfragmentof DNApolymeraseI in
thepresenceofdATP, dGTP, and TTP.
Intramolecu-larligationacrossthese ends resultedin the creation of anEcoRI siteatthejunction. pdPBS1containsthose
viralsequencesbetween nucleotides4632 and 569as a BamHI-to-EcoRIfragmentwithinthelarge
BamHI-to-EcoRI segmentofpML-2DNA.
J.VIROL.
Mutants with deletions internal to viral sequences were constructed as described below. pdPdll-8 was isolated afterdeletion of sequences about theunique
BglI site in pPBR2 with Bal 31 nuclease (18). The BamHI-EcoRI viral DNAfragmentbearing the dele-tionwassubsequently cloned between the BamHI and EcoRI sites of pML-2 to yield pdPdll-8. Sequences
betweennucleotides1and 148havebeendeleted from theviralinsert.
pdPdl300 was derived from pdPBR2 by partial cleavage with PvuII followed by intramolecular liga-tion with T4ligase. Among the bacteria transformed withthis DNAwereseveralthat contained a recombi-nant plasmid with a deletion of the PvuII fragment
between nucleotides 5130 and 5265. pdPdl3O4 and pdPdl326 were isolated after partial cleavage of pdPdl300 with PvuII, followed by digestion with Bal 31 nuclease. Theeroded, linear DNA was then circular-ized with T4ligase and used to transform E.coli. After screening a number ofrecombinant plasmids from independentcolonies, we identified several with dele-tions about the PvuII site at the deletion junction (5130/5265) in pdPdl300 DNA. Sequencing across the deletions oftwo mutants revealed that pdPdl304 is deleted between nucleotide positions 5130 and5277, whereas pdPdl326 is deletedbetween nucleotides 5126 and5265.
ThepdPB500dl300mutants(see Fig. 3) were
assem-bled by digesting pdPBBg(H) DNA (see Fig. 2) to completion withBamHI and PvuII. The largest DNA fragment, composed of pML-2 DNA and viral se-quences between nucleotide positions 5265 (a PvuII site) and 90 (a former BglI site, now aHindIII site), waspurified by agarose electrophoresis and ligated to the gel-purified BamHI-to-PvuII fragments of viral DNAfromthepdPB500 mutant series (Fig. 1).
Theinversion mutants (see Fig. 4) were constructed by ligation of the viral BamHI(4632)-to-PvuII (5130) fragment ofpPBR2 DNA toBamHI-digested pdPBP1 DNA(Fig. 1). After the BamHI cohesive ends ofboth specieswerejoined, themolecules were reacted with the Klenow fragment of DNApolymerase I and the four deoxynucleotide triphosphates to fill in the re-maining BamHI ends. The DNA wasthen circularized by ligation across the filled-in BamHI and PvuII termini with T4 ligase and used to transform E. coli. Afterscreening and characterizing the plasmid DNA from a number ofcolonies, we used two, pdPBP5-1 andpdPBP1-5, for the experiments reported here. An identical strategy enabled us toclone the viral BamHI (4632)-to-PvuII (5130)fragmentinbothorientations at theBamHI site ofpdPP1(B)Bg(H)(Fig. 2) to generate therecombinantsplasmids pdPP1P5-1 and pdPPlP1-5 (Fig. 4).
DNA transfection. CsCl gradient-purified, super-coiled plasmid DNAs were transfected into MOP-8 cells by a modification (39) of the DEAE-dextran transfectiontechnique (29). A60-mmdishcontaining
7.5x105MOP-8 cells was washed with 5 ml of serum-freeDMEM, and the cellswerethenincubated with1
ml of DMEM supplemented with 250 ,ug of DEAE-dextran(molecularweight, 500,000) per ml and 2 ,ug of plasmid DNA at 37°C for 4 h in a humdified CO2 incubator. The cellswerethenwashed twice with5ml of serum-free DMEM and maintained in 5 ml of
DMEMcontaining10% calfserum.
Inseparateexperimentswehavevaried the amount of DNAused fortransfection between0.02and 2.0 ,ug
on November 10, 2019 by guest
http://jvi.asm.org/
perdish and found that thephenotype ofthe mutant
plasmids does not change. We decided to routinely
employ 2.0 ,ug of DNA per dishtoinsure that we could detect replication of recombinantplasmids that were
severely impeded in this capacity.
DNA replication assay. At 72 h posttransfection, low-molecular-weightplasmidDNAwasisolated from MOP-8cells byusingthe Hirt(21) extraction proce-dure. After the sedimentation of
high-molecular-weightDNA,0.5 ml of the clearedlysatewasdiluted in 4.5 mlof TE buffer(10 mMTris-hydrochloride, pH
8.0, 1 mM EDTA) and extracted once with
buffer-saturated phenol and once with chloroform-isoamyl
alcohol(24:1). Nucleic acidswereprecipitated bythe addition of 2.5 volumes of cold ethanol and storage
overnight at -20°C. Precipitates were collected by centrifugationandsuspendedin 50 ,ul of TE buffer. A 15-,ul portion of the DNA sample was sequentially
digested with BamHI andDpnIrestriction
endonucle-ases.
ThedigestedplasmidDNAsweresubjectedto elec-trophoresis through 1.0% (wt/vol) agarose gels, and the DNAfragmentsweretransferredtonitrocellulose filters by the Southern (40)technique. The nitrocellu-losefilterswerehybridizedto32P-labeled, nick-trans-lated pdPBR2 DNA (1 x 108to 5 x 108 cpm/,lg)by employing dextran-sulfate (46). After washing, the filters were dried andautoradiographedfor 3 to12 h with Kodak XAR-5 film and Du Pont Lightning-Plus intensifying screens. We estimate thatby72 h post-transfection there are from 500 to 2,000 replicated plasmidmolecules percell.However, notallof these cells take up and replicate the recombinant plasmid
DNA, andthereforethe number ofcopiesofreplicated
DNA pertransfected cell is likelyto be higherthan this. Each recombinant DNAspecieswasassayed in duplicateonatleast three separateoccasions,and the duplicate sampleswereprocessed separately. We de-tected only minor variations in the recovery of
repli-cated DNA between the duplicates, and the
pheno-typesof themutantsdidnotvarysignificantlyamong the variousexperiments.
RESULTS
Limits of thepolyomavirus sequences required incisforDNAreplication. Todefine the borders ofori weconstructedanumber of polyomavirus DNA-plasmid recombinants and measured their
replicationalcapacity inaMOP cellline
(MOP-8; W. Muller, M. Naujokas, and J. A. Hassell, data not shown). We employed pML-2 as a vectorin these experimentsbecause,likeLusky and Botchan (25), we observed that cis-acting sequences within pBR322 DNA inhibited the replicationofpolyomavirus-pBR322 DNA mole-cules in MOP-8 cells (data not shown). These "poison" sequences have been deleted from pML-2 DNA (25). To assess the replicative capacity of the various plasmids, we employed an assay that allows viral recombinant plasmid DNApropagated in mammalian cells to be dis-tinguished from plasmid DNA that has been propagated in E. coli (33). This assay takes advantage ofthe factthat plasmid DNA isolated
from a DNA-adenosine methylase-positive
(dam')
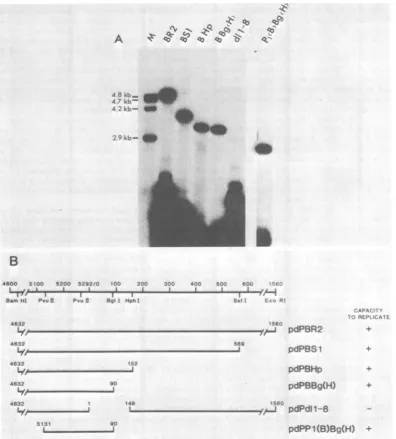
bacterial host is no longer methylatedafter multiple rounds of replication within a mammalian cell. Byusing the restriction endo-nuclease DpnI, which cleaves only methylated DNA, and another appropriate one-cut restric-tion endonuclease, the recovered, low-molecu-lar-weight, replicatedDNA canbevisualized as aDpnI-resistant linearspeciesabove the unrep-licated, DpnI-cleaved, input DNA after South-ernblot transfer and hybridizationto a suitable viralDNA probe. Application of this methodto measure the replication of various polyoma-virus-pML-2 recombinant molecules in MOP-8 cells revealed that allofthecis-actingsequences necessary for replication resided within the small BamHI(nucleotide 4632)-to-EcoRI (nucle-otide 1560) fragment of polyomavirus DNA (pdPBR2; Fig. 1A). We employ the nucleotide numbering scheme proposed by Soedaetal.(38) and report the boundaries of DNA fragments from the clockwise direction on the circular polyomavirus genome. This 2,220-bp fragment includes the region from whichDNAreplication initiates.
Tomoreprecisely delineate the boundaries of thepolyomavirus ori weconstructedaseries of mutantsthatcontain progressive deletions with-in viral DNA sequencesoriginating from either the BamHI site (nucleotide 4632) orthe EcoRI site(nucleotide1560)in pdPBR2 DNA and mea-sured theircapacity to replicate in MOP-8 cells as described above. The structure and replica-tional competence of the various DNAs are shown in Fig. 1. Polyomavirus recombinant
plasmids possessing deletion endpoints up to
nucleotide number 5131 were foundtoreplicate within the MOP-8 cell lineatlevelscomparable tothose ofthe parent plasmid pdPBR2. Howev-er, further deletions extending to nucleotides 5182, 5190, and 5265 rendered these recombi-nant plasmids replication defective (Fig. 1). On the basis of these results, the leftward limit of thepolyomavirus ori can be positioned between nucleotides 5131 and 5182.
To establish the rightward boundary of ori, deletions were created from the EcoRI site to-wardsthe BamHIsitethrough viralsequencesin
pdPBR2 DNA(Fig. 2). Thestructure and
repli-cative capacity of these recombinant plasmids
areillustrated in Fig. 2. All ofthe recombinant plasmids, including one [pdPBBg(H)] with a 1,470-bp deletion from the EcoRI site, were capable of autonomous replication in MOP-8 cells (Fig. 2). These data demonstrate that the rightwardboundary oforiis locatedtotheleftof nucleotide90inpolyomavirusDNA. Toposition
this boundary more precisely we employed a derivative ofpdPBR2, termed pdPdll-8, which bears a deletion betweennucleotides 1 and 148 (18).Measurementofthecapacityof thismutant VOL.47, 1983
on November 10, 2019 by guest
http://jvi.asm.org/
0- 01
1 1 K Q 'CO
A
<1Q) 2 "I
4~4
4>
~
"r Q, qz~'0'o'
B
8 b
4.2
kK.) Ob*
L kL
.48 .zo. b\ _
-H- im_ _
.~_.~ ~
a3
::
Us
D
4600 4700 4800 4900 5000 500 5200 5292/0 100 200 '560
I 1A I 11 .j n
Sc!X PvuE Pvun
BarnHI 4632
l
47812 482;
4834
4841
sCOos
5039
13
5182
5i190
?
5 265
i Hn'
t/,M
pdPBR2
-_1_j
pdPB52O
V/ pdPB51O
-, A-j PdPB5O5
~
pdPB552CAPACITY TO REPLICATE
I* pdPB515
*0-A
pdPBS03
91v,j pdPP1(B)Ri
.+1
,Aj pdPB502
e pdPB523
',oji pdPBP I
FIG. 1. Deletionanalysis of theleftwardboundaryof thepolyomavirusori.(A, B,andC)A2-jig sampleof
polyomavirus recombinantplasmid DNAwastransfected into MOP-8cells,andreplication wasassayed 72h
posttransfectionasdescribedinthetext.After simultaneousdigestionwith BamHIandDpnI, theplasmidDNAs weresubjectedtoelectrophoresis througha1%agarosegel andtransferredtonitrocellulose filters.Replicating
recombinantplasmid DNAcanbe visualizedas aDpnI-resistant band comigrating with thelinearizedparental plasmid afterhybridization andautoradiography. Thesmearthatappearsat thebottom of theautoradiogram represents the closelyspacedfragments of DpnI-cleaved recombinant plasmid DNA that hasnotreplicated.
Becauselarge quantitiesof DNAwereusedtotransfectthe cells, onlyasmall fraction of the totalinputDNAis replicated in theseMOP-8 cells. Theautoradiograms shown in each panelaretaken fromthesamefilm,which was clipped to allow comparisons to be made between adjacent lanes. The laneslabeled M contain DNA
fragmentsofknownsequence.The size ofeachfragmentisshownadjacenttoitspositioninthe autoradiogram. (D) Schematic illustration of the structures ofdeletion mutants relative to known restriction endonuclease
cleavagesitesonthe polyomavirusgenome.The nucleotidenumberingis accordingtoSoedaetal.(38). Because no attempt was made to quantitate the precise level ofDNA replication, the replicational phenotypes are designatedaseither+ or-.
590
0."
%"IV
R-
'0o
C
*).Cj1,6
on November 10, 2019 by guest
http://jvi.asm.org/
[image:5.492.55.448.33.564.2]48kb_... a
4 7 kb-W 4 7
kb-42kb- U_
,29kb-
ft
S
B
4600 5100 5200 5292/0 100 200
1 , L 1 1. .. I I
I, , I I I
Bam Hi Pvull PvuO1 Bgtf HohT
4632
LI
300 400 50C 600
I I I -, I
Sstl [ CC Pi
1560
Iy,i" pdPBR2
4632
I .1
569
152 4632
LI
4632
1 ,.
pdPBSI
pdPBHp
pdPBBg(H)
90
4632 1
5131 90
148 51/
-.,A.1 pdPdl1-8
pdPP1(B)Bg(H) +
FIG. 2. Deletionanalysis of the rightward boundaryofthepolyomavirusori.(A)Thereplicativecapacitiesof
thepolyomavirus recombinant plasmidswere determinedasdescribed forFig. 1. (B)The physicalmaps and
replication phenotypes of the viral deletionmutantsareillustratedasdescribedforFig.1.
DNAtoreplicate revealed that itwasdefective
(Fig. 2). This observation and previous results permitustodefine therightward boundary of ori between nucleotides 1 and 90. On the basis of thesemeasurementswepredictthatone
bound-aryof ori is between nucleotide positions 5131
and5182and that the otherboundaryislocated between nucleotide positions 1 and 90. This prediction was tested by cloning a 251-bp frag-mentofthepolyomavirus genome between the PvuII siteatnucleotide5131 and theBglI siteat
nucleotide90as aBamHI-HindIIIfragment(see
above) within pML-2 DNA and measuring its
replicational potential in MOP-8 cells. The result (Fig. 2) demonstrates that this viral recombinant plasmid, pdPPl(B)Bg(H), is capable of autono-mous DNA replication in MOP-8 cells.
There-fore, the 251-bp viral DNA segment situated betweennucleotides 5131 and 90 contains all the
necessarygenetic information required in cis for
polyomavirus DNA replication.
Polyomavirus ori comprises multiple genetic elements. Todetermine whether the viraloriisa
single contiguous element or is composed of multiple sequence motifs, we constructed
dele-tions within oriand then measured the
replica-92
-}i-.AfA.-.- T
'U,~LfP. (ATE
-4-4-1 47,
on November 10, 2019 by guest
http://jvi.asm.org/
[image:6.492.51.448.70.510.2]592 MULLER ETAL.
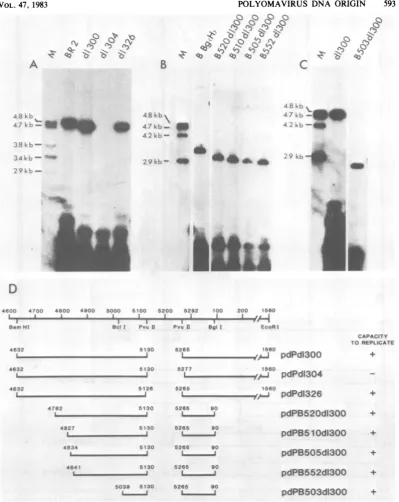
tional capacity ofthederivedmutantplasmids in MOP-8 cells.To carry outtheseexperimentswe planned to introduce deletions about thePvuII siteatnucleotideposition 5265 in pdPBR2DNA. For avariety ofreasons ourinitial experiments led to the isolation of deletion mutants which lacked allof thesequencesbetweennucleotides 5130 and 5265 (sites of cleavage for PvuII) as well as sequences thatflanked these positions. As acontrol, wetested these recombinants for theircapacitytoreplicate andweresurprisedto discover that twoof theaforementioned DNAs werefully capable ofautonomous replication in MOP-8 cells (Fig. 3: pdPdl300 and pdPdl326).
These two DNAs lack sequences thatspan the region between nucleotides5126and 5265 (Fig. 3). Thesequencesmissing in these recombinants hadpreviously been identifiedasformingpartof ori(Fig.1).Theseconflicting observations could bereconciledifwe assumethatsequences else-where within the viral genome (the BamHI-EcoRI fragment) could functionally substitute for those deleted inpdPdl300 andpdPdl326. To pursuethishypothesis we attemptedtoidentify
these sequences by deleting viral DNA from pdPdl300. Deletion of viral sequences between nucleotide positions 90(aBglIsite)and 1560 (an EcoRI site) frompdPdl300 ledto the formation
of a replication-competent plasmid (data not
shown). This observation suggested that the functionally redundant sequences were located between nucleotides 4632 and 5126 inpdPdl300
DNA(Fig.3AandD).Toexamine this
possibili-ty weconstructeda setof deleted derivatives of pdPdl300thatbearlesions whichoriginateatthe
BamHI site
(nucleotide
4632) and terminate atvarious positions within viral DNA (Fig. 3). These deleted recombinants were constructed
by cloning the BamHI-to-PvuIIfragments from
the B500 series of mutants (Fig. 1) within
pdPBBg(H) DNA (Fig. 2) which had been
cleaved tocompletion with BamHI and PvuII.
Theresulting recombinantswerethentested for
their capacity to replicate within MOP-8 cells. The results arepresentedinFig. 3B and C and summarized in Fig. 3D. Theydemonstrate that viral sequences between nucleotides 5039 and 5130 canfunctionally substitute forthose locat-edbetween nucleotidepositions 5131 and 5265. Wehavenamedthesecis-actingsequences thea
and 13elements, respectively.Theaelementand the13elementtogetherareincapableofforminga
functionalorigin(Fig. 2;pdPdll-8). Yetanother sequence motif located between nucleotides 5265and90isrequiredtoformori.This conclu-sion is supported by the isolation ofa mutant,
pdPdl304, which carries a deletion of 12 bp
between nucleotides5265 and 5277 andwhichis replication defective(Fig. 3A and D).
On the basis of these deletion studies we
postulate that thepolyomavirusoricomprisesat leastthreedistinct genetic elements. These ele-mentsincludetwo upstream,functionally equiv-alentregions mappingoneithersideofthe PvuII site at nucleotide 5130, termed the a and 13 elements. In addition, an area of the viral genomecontainingonehigh-affinity largeT anti-genbindingsite,a32-bppalindrome, anda tract ofeight adenine-thymine (A:T) residues desig-nated thecore region is required incisfor viral DNAreplication. Furthermore, neither thecore region alonenortheupstream aand,Belements, alone or together, is capable of autonomous DNA replication in MOP-8 cells (data not shown). This latter observation suggests that a replicationally active recombinant plasmidmust maintain in cis the coordinate presence of at leastoneof theupstream elements(a or13) and anintactcore regiontoremain active in replica-tion.
Inversion of viral sequences containing the a element results in acis-actingdefect inviral DNA replication.Once the approximate boundaries of the sequence elements comprising oriwere es-tablished, we examined the requirement for a positional relationship between these elements. Tothis enda498-bprestriction fragment encom-passing the a element was inserted in both orientationsatthe BamHIrestriction endonucle-asesite of the viralrecombinant pdPBP1, which possesses only the polyomavirus core region (Fig. 4A). One of the resulting recombinant plasmids, pdPBP5-1, which contains the a ele-mentinits native orientation relativetothecore
region, replicated at levels indistinguishable
from those of the pdPdl300 viral recombinant (Fig. 4). This occurred despite the insertion of4
bp at the junction between the a and core
elements. However, inversion of the 498-bp fragment in front ofthe coreregion,asoccursin pdPBP1-5, did not restore replication compe-tence tothe pdPBP1 recombinantplasmid (Fig.
4). It is noteworthy that the inversion mutant pdPBP1-5 not only alters the orientation of a relative to the core region, but also effectively
places 400 bp between the a element and the core region (Fig. 4). To ensure that the pheno-type oftheinversion mutant wasnotduetothe inadvertent generation of sequences inhibitory to DNAreplication,thesame498-bp restriction fragmentbearingawasinserted inboth orienta-tionsatthe BamHI site ofa replication-compe-tent plasmid, pdPP1(B)Bg(H), possessing both the ,B and core regions. The replicative capaci-ties of the resulting recombinants, pdPPlP5-1
andpdPPlP1-5,wereunalteredregardless of the
orientation ofthe upstream a element (Fig. 4). From these results it isapparent that the inver-sion ofahas no detrimentaleffectonthe replica-tionalcapacity ofaviralrecombinantpossessing
J. VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
0~ INl c '
2'
-A
4'7k - w"
r; k., - q
B
C-N,,
q,%
ct Q,Q2)
4
Q0b
;
oo-,
vX
e
48et\ 47kb- *
42;Wb -_0
29kb- fl
23'5
10`1
C
i.,/.; '.
J. .r;_,
- r, _ _
-4
_~ k:-;
4/I_
I
D
4600 4 700 48Cru 490C; 5000 5DoC 5Z200 5292 .00 200 560
1 I
Barn -l1 Bc'I Pvu E Pvu 0I BgiI EcoR
4632 5130 5265 pPI0
i i I . pdPdl300--Vhj
CAPACITY TOREPLICATE
5130 5277 156C
_j I IA
pdPdl3O4
5126 5265
I 1.
15CiO
#-
pdPdl326
+5i130 5265 90
__j L--liiw---i.
5130 5265 90
-J i
48'4 5130
1-841 5130
1 _...j
5265 90
L J._
5265 90
I __J
5039 5130 5265 90
I I L ___
pdPB52Qdl300
pdPB51OdI300
pdPB505dI300
pdPB552dl300
[image:8.492.46.446.58.560.2]pdPB503dl300
FIG. 3. Internaldeletion analysis of the polyoma virus ori. (A, B, and C) The replicativecapacities of the viral deletionmutantswereassessed asdescribed for Fig.1.The recombinantplasmids pdPBR2, pdPBBg(H), and pdPdI300wereincluded in(A), (B), and (C), respectively, for comparativepurposes. (D) Thestructuresofthe deletionmutantsandtheirassociatedreplicational propertiesareillustrated asdescribed for Fig. 1.
an intact and core region. It is therefore
unlikely that the inversion of the a element in front of the core region leads to the chance
creation ofaninhibitorysequencedetrimentalto
DNA replication. Instead it seems more likely
that either the orientation or the spacing (or
both)of the aand core regionsrelativetoeach
other is critical tocreate afunctionalorigin.
DISCUSSION
Thepolyomavirus originfor DNAreplication comprises several genetic elements. These
in-4632
46 32
47B2
4 827
+
+
+ VOL.47, 1983
on November 10, 2019 by guest
http://jvi.asm.org/
594 MULLER ET AL.
*e.
F
K
C,4v kb
"I .o">b>>
e..,,vA;
vKb
It
W
a:~W
4600 4700 43am H1
4632,.
5 130 L
4800 4900 5000 5100 5200 5292
I4 I I I
I I I----V
Bc Pvu O PY:} F B
100 200 t560
weI, Fo.r RI
'
j
~~~~~~~~T
1560 pdPBP5-1
5 30 5 265
CAPAC IT1
CoREPLICATE
L.
s. -v 156C PdPBPI-5
4632 9285
4632 l
4832_ 51
190
pdPP1P5-1 +
pdPPIP1-5 +
FIG. 4. The areplicational elementdemonstrates an orientationorpositiondependence. (A) The replica-tionalcapacities of the viralrecombinants plasmidsweredeterminedasdescribedfor Fig. 1. The recombinant
plasmid pdPdl300 is included for comparative purposes. (B) The structures of polyomavirus recombinant plasmids and theirassociated replication phenotypesareillustratedasdescribed for Fig.1.
clude two functionally equivalent but unique
setsofviral sequences,designatedaand p,that
are situated next toa third element termed the core.Thesimultaneouspresenceof eitheraorP
andanintactcoreregion is requiredtoform ori. All three elements together occupy a contigu-ous, noncoding stretch of the viral genome no morethan345bp in length, between nucleotides
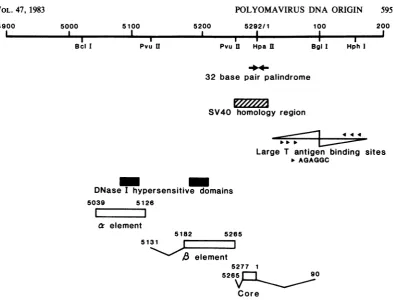
5039 and 90. The sequences that comprise ori and other features resident in this area of the
viralgenome aresummarized in Fig. 5.
Thecoreregion iscomposed ofnomorethan
117 bp situated between nucleotides 5265 and 90. Located within these borders are several
unusualsequencefeatures.They includean8-bp
stretch of A:T residuespositioned between nu-J. VIROL.
B
5130 L
s
~~~~~~~~~~~~~~~~~~~~~~~~~--t3G0v, .5s3
190
T
on November 10, 2019 by guest
http://jvi.asm.org/
5292/1
I I
Pvun1 Hpa I1
100
I I
Bgl I Hph I
-.4-32 base pair palindrome
SV40 homology region
4 44
Large T antigen binding sites * AGAGGC
DNase I hypersensitive domains
5039 5126
L element
5182 5265
5131
element 5277 1
5265 90
Core
FIG. 5. Physicalfeaturesnearthe polyomavirus ori. The viral large T antigenbinding sites arepositioned according to Pomerantz et al. (34). Repeats of the nucleotide sequence -AGAGGC- are designated by
arrowheads. The 32-bp region ofdyadsymmetryisindicatedbyboldarrows.Domains ofDNase I hypersensitiv-ityareshownaccordingtoHerbomeletal. (20). Thevariousreplicational elementsareshownatthe bottom of
thefigure. The borders of the elementsaredenoted byaV-shaped line, where theyareknown.
cleotides 5268 and 5275, a guanine-cytosine
(G:C)-rich 32-bp palindrome (between nucleo-tides 5281 and20), andalarge Tantigen binding
site that resides within the borders of nucleo-tides 5292 and 90 (binding site II; 12, 34). Re-moval of the entire tract ofeight consecutive A:Tbp,asoccursinpdPdl3O4 (Fig.3),orfourof
the eight A:T residues (26), renders the DNA replication defective. These observations
sug-gestthat the A:Ttractisanessential component
of the core region. Interestingly, this sequence
feature is conserved in the DNA oftwo other papovaviruses, SV40 and BK virus, and it is knowntobeacomponentof theSV40 origin for DNA replication (5). There is also evidence to
supportthecontentionthat thepolyomavirus 32-bp palindromeisrequired for ori function. Dele-tion of9 bpbetween nucleotides 5277 and 5287 frompolyomavirus DNA renders that DNA in-capable of replicating (26). This deletion re-moves6bp that constituteoneborder of the
32-bp palindrome. However, removal of 2
contiguous bp within the palindrome does not
alter the replicational properties of the viral DNA, and such molecules can serve as the
genetic material of viable viruses (26). A 27-bp
palindrome that is related in sequence to the
polyomavirusinverted repeat isfoundwithinthe
originofSV40. Moreover, genetic analyseshave
demonstrated that the 27-bp palindrome is an
essential element of theSV40 origin (37). Locat-edadjacenttothe32-bp palindrome in
polyoma-virus DNA are sequences which serve as a
binding site for large T antigen (Fig. 5). This
large T antigen binding site is composed of
sequences between nucleotide positions 5292 and 90(12,34). Because the rightwardborderof thecoreregion has been mapped between
nucle-otides 1 and 90, it is presently not possible to
know whether thisbinding site constitutes part
of the core region. Additional mutants with
lesions in thisareawillberequiredtodetermine unambiguously whether large T antigen binding site II ispartof thecoreregionornot. Nonethe-less, it isprobablynotcoincidental that all viable deletion mutants of polyomavirus isolated to
date retain sequences thatinclude large T
anti-gen binding site 11(3, 4, 17, 26, 27, 47). The
minimal cis-actingsequences required for SV40
DNA replication also include sequences that
serveas bindingsitesforlargeTantigen(5, 14, 30, 42), and thereisawealth ofgeneticevidence VOL.47, 1983
4900
L 5000 5100
BclI
5200
Pvu E
200
on November 10, 2019 by guest
http://jvi.asm.org/
[image:10.492.53.448.51.352.2]596 MULLER ET AL.
that supports thecontention that SV40 large T antigen must interact with ori sequences to initiate DNA replication (14, 36, 37). Taken together, these observationssuggestthatan A:T stretch, a G:C-rich region of dyad symmetry, and a large T antigen binding site are features that not only are shared among papovaviruses butalsoarefunctionallyimportant for the initia-tion of viral DNA replicainitia-tion and constitute domains of thecoreregions of these viruses.
cis-acting viral sequences in addition to the core region are needed to form a functional polyomavirus ori. One such element that, to-gether with the core region, can form ori has beentermed
P.
TheP
elementis located within thepolyomavirus PvuII-4 DNA fragment and is no larger than134bp. One of its borders maps between nucleotides 5131 and 5182, and the other is situated near nucleotide 5265 (Fig. 5). We have recently discovered that either the insertion ofalinkerDNA atnucleotide number 5265orthedeletionofsequences between the,B and thecoreregionshasnoeffectonthereplica-tional capacity of the resulting recombinants (datanotshown). This result demonstrates that although the
1P
element andcore are contiguous within the viral genome they are nonetheless physically distinct genetic elements. There arefewreadily recognizedsequencefeatures within
theborders of,B.Aregion of potential secondary structure canbedrawn thatinvolves base
pair-ing between 43 of the 82 to 134 nucleotides of one of the DNA strands that make up this element(45). Itmaybenoteworthy thata
repli-cation-defective viral recombinantpossessinga deletion that extends to nucleotide 5182 (pdPB502; Fig. 1) destroys this region of
puta-tivesecondarystructure, suggestingthat itmay be of regulatory significance. The
1B
element resides withinan areaof theviralgenomethatis tandemlyduplicatedand mutatedby base substi-tutions in variants of polyomavirus which, un-like the wild-type virus, are capable of produc-tively infecting the F-9 embryonal carcinoma cellline (35). The observation that coinfection of F-9 embryonal carcinoma cells with wild-type and mutant strains ofpolyomavirus yields only mutant virus progeny, despite the presence ofcomplementing large T antigen (inthe infected
cell), indicates that the sequence modifications in thesemutantslikely allowthem to overcome a cis-acting defect in viral DNA replication pres-entinwild-typeDNA(11).
Although the
1P
element isimplicated in viral DNAreplication,it canbe deletedfrom the viral genomebyPvuIIcleavage with little phenotypic effectonthereplicational capacity oftheresult-ing viral recombinant. An adjacent region of viral DNAbetween nucleotides 5039and 5126, designateda, canfunctionallysubstituteforthe
13 element.
Recently,
Tyndall
et al.(44)
have described the replicational properties ofa virus-recombinant plasmid possessing an analogous 134-bp deletion betweenthePvuII sitesat nucle-otides 5130 and 5265. They found that removal of thesesequenceshadapronouncedinhibitory effectonthereplicational capacity of the deletedrecombinant plasmid DNA and rendered
poly-omavirus nonviable. We therefore measured the
replicational capacity of the PvuII-4 deletion
mutant DNA (pdPdl300; Fig. 3) in competition with another recombinant plasmid DNA
(pdPBS1; Fig. 2) which contains all three
seg-ments, namely, a, 13, and core. The results revealed that bothspeciesreplicated with equal efficiency (data not shown). Moreover, Luth-man et al. (26) have recently described the construction of a viable deletion mutant of
polyomavirus(d11024)from which all but 8bpof
the 134-bp PvuII-4 fragmenthas been deleted. Their results and ours suggest thatthis area of theviralgenome (betweennucleotides 5130 and 5265) is dispensable for viral gene expression
and DNA replication (see also reference 18). Thedifference between these results and those of Tyndall et al. (44) may be due to as yet undetected differences in the virus strains or perhapsto mutations in the various clonedviral DNAs. In addition, the amounts of large T
antigen within COP cells (44) and MOP-8 cells
undoubtedly differ, and this parameter may in-fluence theextentofreplication ofmutantviral genomes. Whatever the explanation, there is little doubt that the a element can functionally
substitute for 1, quantitative differences not-withstanding. It isinterestingthatTyndalletal. (44) have isolated unidirectional and bidirec-tionalmutantsabout the PvuIIsiteatnucleotide
position5130(Fig.5). Many of the bidirectional
mutantsfail toreplicate, whereas the unidirec-tionalmutantsthataredeleted towardeither the
early or late transcription units do replicate.
These resultsarereadilyexplained bythemodel we have presented. The bidirectional mutants werereplicationally inactive becausesequences thatcompriseaand13weredeleted.Bycontrast, the unidirectional deletion mutant DNAs bear lesionsthat removeeithera or13 andare
there-forereplication competent.
Variantsofpolyomavirushavebeendescribed that replicate efficiently in PCC4 embryonal carcinoma cells. These variants possess
com-monsequencerearrangements thatresult inthe
duplication of viralsequences between
nucleo-tides 5072 and 5135 (the borders ofa are be-tween 5039and 5126)andthe deletion ofthose sequences between nucleotides 5185 and 5215 (the latter map within
13;
23). These sequence modifications likely delete the aelement while duplicatingthe 13 element. Because these virusJ. VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
variants, capable of productive infection of PCC4 cells, retain the capacity to replicate in secondaryculturesofmouseembryo cells, these results too suggest that a sequences can func-tionally substitutefor the f3 element.
Theactivity of theaelement is dependenton its position or orientation relative to the core sequences. Insertion ofa 498-bp fragment that containsainfrontof thecoreregion in its native orientation resulted inaDNAmolecule capable ofautonomous replication in MOP cells. How-ever, insertion of the 498-bp a-containing frag-ment in the opposite orientation relative tothe core did not result in a replication-competent plasmid. Control experiments demonstrated that thiswas notduetotheinadvertentcreationofa
sequencethatretardsreplicationlike that which
occursinpBR322DNA(25). Because theaand core sequences arealso moredistantly separat-ed in the inversion mutant in comparison with their native state, it is not possible to state whether theorientation orspacing ofarelative to the core is important for origin function. Whatever thenatureof thedefect, the inversion mutant illustrates that the mere presence ofa and the core onthe sameDNA molecule isnot sufficient for its replicationin MOP cells.
The function of the a and
1B
elements is not known. The viral sequences that span nucleo-tides 5021 to 5265 (a and 13 together map be-tween nucleotides 5039 and 5265) have been reported to enhance transcription of the rabbit,B-globingeneinhumanandmousecells(7). It is
temptingtosuggest thataand ,B arecomposed ofsequences thatserve adual function inboth DNA replication andgene expression. For ex-ample, itispossible thatDNA mustbeanchored to the nuclear matrix or some other cellular constituenttobeefficiently expressedand
repli-cated. Ifspecificsequencesarerequiredtoform such complexes, then these might be located within theaand 13elements. However,until the sequences required for gene expression and those required for DNA replication are more precisely defined, such models should be con-sidered speculative.
The a and 13 elements share little if any sequence homology, yet they apparently
func-tion in thesamemanner.Thisimplieseither that different sequences within each element are functionally importantor thattheseregions can assume similarsecondary ortertiary structures. Withinlytically infected cells atlatetimes, both the a and ,B elements are relatively free of nucleosomes and hypersensitive to DNase I digestion (Fig. 5; 20).Itispossible that thea and 13 sequences serve as sites of recognition for cellular proteins required forDNA replication. It is tempting to speculate thatcellular permis-sive factors interact with sequences present
within these elements. Such arole isconsistent with the sequence changes that occur within these regions when polyomavirus is adapted to replicate within F9 andPCC4 embryonal carci-nomacells.
The multielement organization of the poly-omavirus ori resembles the structure of the SV40ori. In SV40 DNA the functionalorigin is composed ofa core segmentanalogous tothat found in polyomavirus and an adjacent region consisting of two perfect and one imperfect
G:C-rich,21-bprepeats(5). Theefficient
replica-tion of SV40-plasmid recombinant DNAs in COS cells(13)requires thecoreandatleastone of theserepeats(5). Incontrast tothe polyoma-viruscoreregion, theSV40core,whenlinkedto plasmidsequences, iscapable of replicating au-tonomously in COS cells. The auxiliary region containing the 21-bp repeats has an enhancing
effectonreplicationefficiencythatis dependent onthenumber ofcopies of the 21-bprepeats(5, 10). Interestingly, an identical 21-bp sequence hasbeen reported toresidewithin the inverted repeattermini ofsimian adenovirus 7 DNA (43). Moreover, sequences of related primary struc-ture areconserved within the terminal, inverted repeatsof the DNA ofmanydifferentserotypes of human adenovirus (41). Although it is not known whether these adenoviral sequences are elements of the functional replication origin,
theirlocationnearthe sites of initiation of viral DNA replication is suggestive ofarole in this process. Taken together, these observations suggest to us that a multielement organization
may be a common feature of viral and, by
analogy, cellularreplicationorigins.
ACKNOWLEDGMENTS
Wethank MonicaNaujokas and CathyCollins-Maynardfor
excellent technical assistance, Mike Botchan for a gift of pML-2 DNA,FenellaRedway forconstructingsomeof the recombinant plasmids used in this study, and Dal Briedis for his comments on the manuscript.
This research was supported by funds from the National Cancer Institute(NCI) and the Medical Research Council of
Canada. W.J.M., C.R.M., and A.-M.M. are supported by
researchstudentships from the Medical ResearchCouncil, the NaturalSciences andEngineering Research Council of Cana-da,and theNCI, respectively. J.A.H. is a research scholar of the NCI.
LITERATURE CITED
1. Basico,C., Y. Matsuyo, and H.Green. 1970. The interac-tion ofpolyoma virus with mouse-hamster somatic hybrid
cells.Virology 41:295-299.
2. Basfico,C., and R. Wang. 1971.Susceptibility to superin-fection ofhybridsbetweenpolyoma "transformed" BHK and "normal" 3T3 cells. Nature (London) New Biol. 230:105-111.
3. Bendig, M. M.,andW. R. Folk.1979. Deletion mutants of
polyomavirus defininganonessentialregion between the
47,
on November 10, 2019 by guest
http://jvi.asm.org/
598 MULLER ET AL.
origin ofreplication and theinitiation codon for early
proteins.J. Virol.32:530-535.
4. Bendig, M. M., T. Thomas, and W. R. Folk. 1980. Regula-tory mutantsof polyoma virus defective in DNA
replica-tionandsynthesis of early proteins. Cell 20:401-409. 5. Bergpma, D.J., D. M. Olive, S. W.HartzeUl, and K. N.
Subramanian.1982.Territoriallimits andfunctional anat-omy ofsimian virus 40 replication origin. Proc. Natl. Acad.Sci. U.S.A. 79:381-385.
6. Black, P. H. 1964. Studies on the genetic susceptibility of cells topolyoma virus transformation.Virology 24:179-185.
7. DeVlliers, J., and W. Schaffner. 1981. A small segment of
polyoma virus DNA enhances the expression of the
cloned13-globingene over adistanceof 1400 basepairs. Nucleic Acids Res. 9:6251-6264.
8.Francke, B., and W. Eckhart. 1973. Polyoma gene func-tionrequired forviral DNAsynthesis. Virology 55:127-135.
9.Friedmann, T., A. Esty, P. Laporte, and P. Deininger. 1979.The nucleotide sequence and genomeorganization
of the polyoma early region: extensive nucleotide and
aminoacidhomologywith SV40. Cell 17:715-724. 10. Fromm, M., and P. Berg. 1982. Deletionmappingof DNA
regions requiredfor SV40early regionpromoterfunction invivo.J. Mol.Appl. Genet.1:457-481.
11. Fujimura,F.K., andE.Linney.1982.Polyomamutants thatproductively infectF9embryonal carcinomacells do not rescue wild-type polyoma in F9 cells. Proc. Natl. Acad.Sci. U.S.A.79:1479-1483.
12. Gaudray, P., C. Tyndall, R. Kamen, and F. Cuzin. 1981. Thehigh affinity bindingsiteonpolyomavirus DNA for virallarge-T protein. Nucleic AcidsRes. 9:5697-5710. 13. Gluzman, Y. 1981.SV40-transformedsimian cells support
thereplicationofearlySV40mutants.Cell 23:175-182. 14. Gluzman, Y., R. J. Frisque, and J. Sambrook. 1979.
Origin-defective mutants ofSV40. ColdSpring Harbor
Symp. Quant.Biol. 44:293-300.
15. Griffin,B. E. 1982.Structureandgenomic organization of SV40 and polyomavirus, p. 61-123. InJ. Tooze (ed.), DNA tumorviruses,2nd ed.ColdSpringHarbor Labora-tory,ColdSpringHarbor, N.Y.
16. Griffin, B. E., and M. Fried. 1975. Amplificationof a
specific region of the polyoma virus genome. Nature
(London)256:175-179.
17. Griffin, B. E., and C. Maddock. 1979. New classes of
viable deletion mutants in the early region of polyoma virus. J. Virol. 31:645-656.
18. HasseUl,J. A., C. MueUler,A.-M.Mes, M. Featherstone, M.Naujokas, B. Ponerantz, and W. Muller. 1982. The constructionofpolyomavirusvectors:functionsrequired
for gene expression, p. 71-77. In Y. Gluzman (ed.),
Eukaryotic viralvectors.ColdSpringHarborLaboratory,
ColdSpringHarbor,N.Y.
19. Hassell, J. A., W. C. Topp, D. B. Rifkin, and P. E. Moreau. 1980.Transformationofratembryofibroblasts
byclonedpolyomavirusDNAfragments containing only
part of theearly region. Proc. Natl. Acad. Sci. U.S.A. 77:3978-3982.
20. Herbomel, P., S. Saragosti, D. Blangy, and M. Yaniv. 1981.Finestructureoforigin-proximalDNAase
I-hyper-sensitiveregioninwild-typeand E.C. mutantpolyoma.
Cell 25:651-658.
21. Hirt,B.1967.Selective extraction ofpolyomaDNAfrom infectedmousecell cultures. J. Mol. Biol. 26:365-369. 22. Kamen, R.,J. Favaloro, J. Parker,R.Treisman,L.Lania,
M.Fried, and A.Mellor. 1980.Comparisonofpolyoma
virus transcription in productively infectedmousecells and transformed rodent cell lines. Cold SpringHarbor Symp.Quant.Biol.44:637-651.
23. Katinka, M.,M.Kanin,M.Vasseur,and D.Blan. 1980.
Expressionofpolyoma earlyfunctions inmouse
embryo-nalcarcinoma cellsdependsonsequence rearrangements in thebeginningof the lateregion.Cell 20:393-399. 24. Lund, E., M. Fried, and B. E. Griffin. 1977. Polyoma
virusdefective DNA's. I.Physical maps of a related set of defective molecules (D76, D91, D92). J. Mol. Biol. 117:473-495.
25. Lusky, M., and M. Botchan. 1981. Inhibition ofSV40
replicationin simiancellsby specific pBR322DNA se-quences. Nature(London)293:79-81.
26. Luthman, H., M. G.Nilsson,andG. Magnusson. 1982.
Non-contiguous segmnents of the polyoma genome re-quiredin cisfor DNAreplication.J.Mol. Biol. 161:533-550.
27. Magnusson, G., and P. Berg. 1979. Construction and
analysis of viable deletionmutantsofpolyomavirus. J. Virol. 32:523-529.
28. Maxam, A., and W. Gilbert. 1977. Anew method for
sequencingDNA. Proc. Natl.Acad. Sci. U.S.A. 74:560-564.
29. McCutchan, T.,andJ.S.Pagano.1968. Enhancement of
infectivityof simianvirus40deoxyribonucleicacid with
diethylaminoethyl-dextran.J. Natl. Cancer Inst. 41:351-357.
30. Mes,A.-M.,andJ.A.Hasseil.1982.Polyomavirus middle
T-antigen is requiredfortransformation. J. Virol. 42:621-629.
31. Myers, R. M., and R. Tjian. 1980. Construction and
analysis of simian virus 40 origins defective in tumor antigen bindingand DNAreplication.Proc.Natl. Acad. Sci. U.S.A. 77:6491-6495.
32. O'Hare,K.1981.Replicationofpolyoma plasmid recom-binants inmousecells. J.Mol. Biol. 151:203-210. 33. Peden,K.W.C., J.M. Pipas,S.Pearson-White,andD.
Nathans.1980. Isolation ofmutantsofananimalvirus in
bacteria. Science209:1392-13%.
34.Pomerantz,B.J., C. R.Mueller,andJ.A.Hassell. 1983.
Polyomavirus large T antigen binds independently to multiple, unique regionson theviral genome. J. Virol. 47:600-610.
35.Sekikawa, K., and A. J. Levine. 1981. Isolation and characterization ofpolyomahostrangemutantsthat
repli-catein nullpotential embryonal carcinoma cells. Proc.
Natl.Acad.Sci. U.S.A. 78:1100-1104.
36. Shortle, D., R. F. Margolskee, and D. Nathans. 1979.
Mutationalanalysisof theSV40replicon: pseudo rever-tantsofmutantswithadefectivereplication origin.Proc. Natl. Acad.Sci.U.S.A. 76:6128-6131.
37. Shortle, D.,and D.Nathans.1979.Regulatorymutantsof simian virus 40: constructedmutantswith base
substitu-tions at the origin of DNA replication. J. Mol. Biol. 131:801-817.
38.Soeda, E., J.R.Arrand,N.Smolar, J.E.Walsh,and B. E. Griffin. 1980. Coding potential andregulatory signalsof thepolyomavirus genome. Nature(London)283:445-453. 39. Sompayrac, L. M., and K. J. Danna. 1981. Efficient infectionofmonkeycells with DNAof simian virus 40. Proc.Natl. Acad.Sci.U.S.A. 78:7575-7578.
40. Southern,E. M. 1975. Detection ofspecific sequences amongfragmentsseparated byagarosegel electrophore-sis.J. Mol. Biol.98:503-515.
41. Stillman, B. W.,W. C. Topp, andJ.A. Engler. 1982. Conserved sequencesatthe origin of adenovirus DNA
replication.J. Virol. 44:530-537.
42. Subrananan,K.N.,and T.Shenk. 1978.Definition ofthe boundaries of theoriginof theoriginof DNAreplication insimian virus 40.Nucleic AcidsRes.5:3635-3642. 43. Tolun,A.,P.Alestrom,andV. Petterson.1979.Sequence
of the inverted terminalrepetitionsfrom different adenovi-rus:demonstration ofconservedsequences andhomology between SA-7 termini andSV40DNA.Cell 17:705-713. 44. Tyndall, C., G.Lamantia, C. M. Thacker, J. Favaloro,
and R. Kamen. 1981. A region of the polyoma virus genome between the replication originand late protein codingsequences isrequired incis for bothearlygene expressionand viralDNAreplication.Nucleic AcidsRes. 9:6231-6249.
45. Vasseur, M.,M.Katinka,P.Herbonel,M.Yaniv,and D. Blangy. 1982.Physicalandbiologicalfeatures ofpolyoma
J. VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
virus mutants ableto infect embryonal carcinoma cell lines. J. Virol. 43:800-808.
46. Wahl, G.M., M. Stern,andG. R. Stark.1979.Efficient transfer of large DNA fragments from agarose gels to diazo benzyloxy methyl-paper and rapid hybridization using dextran sulfate. Proc. Natl. Acad. Sci. U.S.A.
76:3683-3687.
47. WeUs, R. D.,M. A.Hutchinson,andW.Eckhart. 1979. Isolation andcharacterizationofpolyomavirusgenomes with deletions between the origin ofviral DNAreplication and the site of initiation of translation in theearlyregion. J.Virol. 32:517-522.
on November 10, 2019 by guest
http://jvi.asm.org/