0022-538X/89/031354-06$02.00/0
CopyrightX) 1989,American Society forMicrobiology
Human Gamma
Interferon and Tumor Necrosis
Factor Exert
a
Synergistic
Blockade
on
the
Replication of Herpes Simplex Virus
ELENAFEDUCHI, MIGUEL A. ALONSO, ANDLUIS CARRASCO*
CentrodeBiologla Molecular, Consejo Superiorde Investigaci6nes Cientificas, UniversidadAutonoma deMadrid, CantoBlanco,28049 Madrid, Spain
Received 15 August1988/Accepted 1 December 1988
Thereplication of herpessimplexvirus type 1(HSV-1)isnotinhibited in either HeLaorHEp-2cellstreated withhuman alphainterferon(HuIFN-a), particularlywhenhigh multiplicitiesofinfectionareused.However,
HuIFN--y partially inhibits HSV-1 translation in HEp-2 cells infected at low multiplicities. Under these conditions, the transcriptionofgenesa22, TK,and-yOisgreatlydiminished. The combinedaddition ofhuman tumornecrosis factor(TNF)andHuIFN-'ytoHEp-2cells exertsasynergisticinhibition of HSV-1 translation. Cells treated with bothcytokinescontinuesynthesizing cellularproteins, even20h after HSV-1 infection. As little as 10 U ofIFN-,y perml blocked HSV-1 DNAreplication, provided that TNFwas also present inthe medium.AnalysesofHSV-1genetranscriptionsuggest thatthe action of bothTNF andIFN--yblockedastep
thatcomesatorpriortoearlyHSV-1geneexpression. Thisearly stepinHSV-1replicationinhibitedbyTNF andIFN--yoccurs after virus attachmentand entry intocells, since the internalization of radioactive HSV-1 virion particles was not blocked by the presence of the two cytokines. Therefore, we conclude that the synergistic action of TNF plusIFN-yaffectsastep in HSV-1replicationthatcomesafter virusentrybutbefore
oratthe transcriptionofimmediate-earlygenes.
Theresponseof animal virusestotheinhibitoryeffects of interferon (IFN) greatly dependson a numberof variables.
The IFN species used to block virus replication is an
important factor, since some viruses, adenoviruses for
ex-ample, are inhibited by IFN-y, but notby IFN-a (14). The virus-cellsystemis another factor of interest, becausenotall animalviruses studiedareequallysensitive toIFN (11, 19). Ingeneral, viruses containingRNAmoleculesasthegenome aremore pronetobe inhibited byIFNthanDNA-containing viruses (11, 19). The replication ofvesicularstomatitisvirus
orpoliovirusisvery sensitive toall IFNspecies assayed in the different celltypes used (11, 19).On the otherhand,the translation of reovirustype3isnotinhibitedby human alpha interferon (HuIFN-a) in HeLa cells, but it is blocked by murinealpha-beta interferon
(MuIFN-a-P)
inmousecells(9,18). Theresistance of reovirus replicationtoIFN isnotdue toreversion of theantiviral state(9).Thisbehaviorcontrasts withthat ofmostDNA-containing animal viruses studied (8, 22). Vacciniavirus andadenovirusesowetheirresistanceto IFNtothefact that theyareableto revertthe antiviralstate established in cells treated with IFN (8, 22). For
herpesvi-rusesthesituation ismorecomplex, and theresponsetoIFN largely dependsonthe systemunder study. Herpes simplex virus type 1 (HSV-1) replicates well in HeLa cells treated with HuIFN-a; neither DNA replication nor viral protein
synthesis is inhibited in these cells (4, 17), but the virions made incellstreated with IFN-aareunabletoreplicate(17). Chatterjee et al. (4) found a reduction in the amount of glycoproteins D andB of HSV-1 andadrastic inhibition of
thereleaseofmatureextracellular particles. Morerecently, itwasfound that the transcriptionof HSV-1 earlygeneswas
inhibited in HeLa cells treated with very high doses (4,000
U/ml) ofHuIFN-aA and infected at low multiplicity (20). The use ofanother cell line, Chang cells, shows that high doses(100to1,000U/ml) of HuIFN-aor
HuIFN-P
doblockthe growth ofHSV-1. Consequently, no viral proteins
ap-pearlatein infection when Chang cellsaretreatedwith 300
or 1,000 U of HuIFN-, per ml (3). HSV-1 replication is sensitivetoIFNinmouseand humanmacrophages (5, 6,15, 25). Thus, primary cultures ofsplenic mouse macrophages
treated with
MuIFN-a-P
do not express immediate-earlygenesof HSV-1(15). TreatmentofmacrophageswithIFN--y
in combination with IFN-aor
IFN-P
resultedinasynergistic inhibition of HSV-1 growth (5). Therefore, the behaviorof HSV-1in humanor mousecells treated withavarietyofIFN preparations is different.Thesearch for differentcytokinecombinations whichare moreeffective inblockingHSV-1replicationinhumancells mayleadtoanimprovementof theavailable chemotherapy againstthisimportanthumanpathogen. In thisregard,anew cytokineknown astumornecrosisfactor (TNF) has shown some promise, used either alone or in combination with
different IFNs against a numberof animal viruses (13, 30). TNF is acytokinewithpleiotropic effects (1, 21). TNF isa 17,000-dalton polypeptide synthesized by macrophages in
response to several agents such as lipopolysaccharide or
tubercle bacilli (1, 21). This protein is best known for its antitumoreffects in vivo and for its invitrocytotoxicitytoa
number of transformedcell lines (1, 21). In addition, TNF also stimulates the growth of normal fibroblasts (26, 28), inhibits the synthesis oflipoprotein lipase (1), and activates polymorphonuclear leukocytes (23). Recently, an antiviral activityof TNFwasdescribed(13,30).TNFalone showsan
antiviral effect in several cell lines against a number of
animal viruses, such as vesicular stomatitis virus,
enceph-alomyocarditis virus, adenovirustype2, orHSV-2 (13,30).
Moreinterestingly,theantiviraleffects of TNFareenhanced
synergistically by IFN-y (30). These results prompted an
investigationtoclarifytheaction ofTNFandIFN--y against HSV-1 in human fibroblasts, and the results are presented
here.
*Corresponding author.
1354
on November 10, 2019 by guest
http://jvi.asm.org/
MATERIALS ANDMETHODS
Cells and viruses. HEp-2 and Vero cells were grown in petri dishes (Nunc, Roskilde, Denmark) containing 10 mlof Dulbecco modified Eagle medium(E4D) supplemented with
10%calf serum (GIBCOLaboratories, Grand Island, N.Y.) and incubated at 37°C ina5% CO2atmosphere.
HSV-1 was grown on Vero cells in E4D medium
supple-mented with 2% calf serum (E4D2). The fraction obtained
afterremoval of cell debris by low-speed centrifugationwas
used as the source of virus. Virus preparations were titrated
by plaque assay.
Plasmids. The 2-kilobase (kb) PstI (a22 gene), 2.4-kb
EcoRI (TK gene), and 3-kb HindlIl (-yO gene) restriction
fragments obtained from HSV-1 genome digestion and cloned in pBR322, pACYC184, and pACYC177,
respec-tively, were generously provided by E. Tabards (Madrid,
Spain).
IFNs and TNF. Human lymphoblastoid interferon,
HuIFN-(Ly)-a (1.7 x 106U/ml),was agenerousgift fromN.
Finter (Wellcome Research Laboratories, Beckenham,
England). Recombinant human gammainterferon,
rHuIFN--y (2 x 107 U/ml), and recombinant human tumor necrosis
factor, rHuTNF-a (6 x 107 U/mg), were generously
pro-vided by G. R. Adolf(Ernst-Boehringer-Institut fur
Arznei-mittel-Forschung). Humanfibroblastinterferon,
HuIFN-P
(5x 104U/ml), was from Lee Biomolecular Research Labora-tories (San Diego, Calif.).
Conditions of infection. HEp-2cells, grown in96-well,
60-or100-mm-diameterdishes,wereinfectedwith HSV-1atthe
multiplicity of infection (MOI)indicated in eachexperiment
in the Results section. After 1 h ofincubation at 37°C, the
medium was removed, and E4D2 was added. The time of
virusadditionwasconsidered -1h, andzerotimewastaken
tobethe time when thevirus was removed. IFN and TNF
treatment was done 20 to 24 h before infection at the
concentration described in the figure legends, in E4D2.
Medium with IFN and TNFwasremoved,and theinfection
was doneasdescribed above.
Analysis of proteins by sodiumdodecyl sulfate-polyacryla-mide gelelectrophoresis. HEp-2 cellsgrownin 96-well dishes
were incubated with 50 ,ul of methionine-free medium and
1.25 ,uCi of
[35S]methionine
(1.45Ci/mmol;
AmershamCorp., Arlington Heights, Ill.) for 1 or2 h at thetime after
infection indicated in each experiment. Cell monolayers
were dissolved in50 ,u ofsample buffer (62.5 mMTris
[pH
6.8],
2% sodium dodecyl sulfate, 0.1 Mdithiothreitol,
17%glycerol, 0.024% bromophenol blue). Samples were heated
at90°C for5min, and 10 ,ulwas
applied
to a15%polyacryl-amide geland runovernight at 100V/20 cm.
Fluorography
was carried out in 20% (wt/vol) 2,5-diphenyloxazole in
dimethyl sulfoxide. Thegelswere finally driedandexposed
with Kodak films (10).
RNA isolation and analysis. HEp-2 cells were grown in 100-mmdishes and infected at5PFU per cell. At the times
indicated,cellswerelysedinabuffer
containing
10 mMTrishydrochloride (pH 7.8), 1 mM EDTA, 150 mM
NaCl,
and0.65% NonidetP-40. Afternucleiwereremoved
by
centrif-ugationat2,000 xgfor5min, supernatantsweremixed with
anequal volumeof buffer
containing
20 mM Trishydrochlo-ride(pH 7.8), 350 mM
NaCl,
20 mMEDTA,and1%sodiumdodecyl sulfate. Samples wereextracted three times witha
mixture ofphenol and chloroform (24:1,
vol/vol),
and theRNAwasprecipitatedwith ethanol
(7, 12).
Dotblotanalysis
with32P-labeled nick-translatedHSV-1TKDNA,a22
DNA,
and
lyO
DNAwasperformed
aspreviously
described(27).
DNA isolation and
analysis.
HEp-2
cells were grown in 60-mmdishes and infectedwith HSV-1at30PFU/ml. At 16h
postinfection
(p.i.),
cells werecollected,
and DNA wasisolated
essentially
as describedpreviously (2).
DNA wasimmobilizedonto nitrocellulosemembranes with a
microfil-trationapparatus
(Bio-Rad
Laboratories, Richmond,
Calif.)
and
subjected
tohybridization
to nick-translatedTK DNA.Preparation
of radiolabeled viralparticles.
HEp-2
cellsgrownin 100-mmdishesandinfectedwith 10 PFUofHSV-1
percellwere labeledwith 50
,uCi
of[35S]methionine
permlfrom 10 to 24 h
p.i.
Cells werescraped,
and the fractionobtainedafter removalofcelldebriswas
centrifuged
over a30%dextrancushionat
40,000
rpmfor3 hat4°C
in a65Ti rotor(Beckman Instruments, Inc.,
Fullerton,
Calif.).
Thepellet
wassuspended
in Dulbecco modifiedEagle
mediumand usedasradiolabeled virions tomeasurevirus entry. Virusentry.Theentryof virus into cellswasmeasured
by
estimating
thetrichloroaceticacid-precipitable
radioactivity
in the cell
monolayer
at different times afterinfection withradiolabeled virus
(see
above)
at37°C.
Thevaluesobtainedat
4°C
were taken as a measure of virusadsorption.
Attheindicated
times,
the medium was removed and cellmono-layers
on 24-well dishes were washed withphosphate-buffered saline-Dulbecco modified
Eagle
medium and0.0025%
trypsin-0.08%
EDTA in Dulbecco modifiedEagle
medium,
precipitated
with 5% trichloroaceticacid,
washedwith
ethanol,
solubilized with0.1% NaOH-1% sodiumdo-decyl
sulfate,
and counted in anIntertechnique
scintillationspectrometer.
RESULTS
Effects of different IFNs on HSV-1
replication.
Previousexperiments
indicated thatHSV-1 was abletoreplicate
andsynthesize
proteins
inhumanfibroblasts(HeLa
cells)
treatedwith IFN-a
(4, 17).
Toanalyze
thesensitivity
of HSV-1replication
toHuIFN-a,
HuIFN-P,
andHuIFN--y,
HEp-2
cells were treated with a
high
concentration(400
U/ml)
ofdifferent human IFNs and were infected with HSV-1 at
several
multiplicities.
As found inprevious studies,
thesynthesis
ofHSV-1proteins
is very resistant to treatment with HuIFN-a(Fig.
1).
Thisphenomenon
isdependent
upon the MOI used. When cells are infected at lowMOI,
theexpression
ofsomeviralproteins
isblocked.Theincrease inMOIovercomestheinhibition ofHSV-1
protein
synthesis
incells treated with IFN-a. It is
surprising
to find thatIFN-fbehavesasIFN-yand that both IFNs showa
higher
activity
against
HSV-1 than does IFN-a. At low MOI most ofthe HSV-1proteins
are not evident whenanalyzed
by
sodiumdodecyl
sulfate-polyacrylamide gel
electrophoresis,
whereas athigh MOI,
viralprotein
synthesis
is similar to that of control cells not treated with IFN. These results reinforcethe idea that HSV-1
replication
isquite
resistant toIFN,
particularly
toIFN-a,
asfarastranslation is concerned. To compare the effect of IFN-a andIFN-,y
on thetran-scription
of HSV-1 genes, theexperiment
shown inFig.
2Bwasconducted. TotalRNAfrominfected cellstreatedornot
treated with IFN was extracted at 4 and 8 h
p.i.
andhybridized
withprobes
derived froma22, TK,
andyO
HSV-1 genes(29).
Inhibitors ofprotein synthesis
suchas cyclohex-imide block theexpression
ofP
and y genes but not thetranscription
of the a class(Fig.
2A).
On the otherhand,
inhibitors ofviral DNA
replication
such asphosphonofor-matehavean
inhibitory
effectonry geneexpression,
butthey
do not block the
transcription
of a and,3
genes. HSV-1 expresses the-yO
and TK genes at control levels inHEp-2
on November 10, 2019 by guest
http://jvi.asm.org/
mm, 5 moL2 3
IFN c.3 ic
{40)u/mi
MW r10-3151
126- _
6
^
...
I.
-mu__
foSW 4 -*-- -Actin
-4
26-
-18- -_
1I
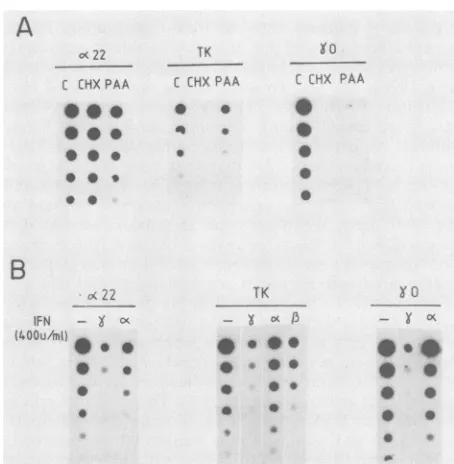
...-FIG. 1. Effect ofIFNs onproteinsynthesisin HEp-2 cells infected with HSV-1. Cells grown in 96-well dishes were treated with IFNs 20 hbeforeinfection at the MOI indicated. Metabolic labeling was carried out for 2 h at 15 h p.i. with[35S]methionine,andproteins wereanalyzed asdescribed inMaterials and Methods. C, Control without IFN. The numbers indicate molecular size in kilodaltons.
A
O<22 TK Yo
C CHXPAA C CHXPAA C CHX PAA
0 * 0
* *
B
B(22
IEN - X c
(400u/mtH
*
* *
0
0
~~0
0
TK
X CK
* 0 *
*t 0
* *
0 0
*
* 0 0 1,
FIG. 2. Effects of different IFNsonthe transcription ofdifferent
HSV-1genes.Cellswereinfected with5 PFUof HSV-1percell, and mRNAwasextractedat4 hp.i. for the a22geneblots andat8hp.i. fortheTK and-yOgeneblots. (A) Effect of cycloheximide(CHX) (5
X l0-5 M) and phosphonoaceticacid (PAA) (250 ,ug/ml) on
tran-scription of HSV-1 immediately-early (a22), early (TK), and late
(-yO) genes. (B) Effect of IFNs on transcription of HSV-1 genes.
Cellsgrownin 100-mm disheswere treated with IFN 24 h before
infection.C, Control; -,control withouttreatment.
cells treated with 400 U ofHuIFN-a per ml andinfected at low MOI of HSV-1(Fig.2B). Thesynthesisof the (x22 gene, however, ispartiallyinhibited by HuIFN-a, suggesting that under theseconditionsIFN-aalsohassomeinhibitory effect
on theexpression of earlygenes. The sensitivity of HSV-1
gene expression to IFN-y is clearly apparent when the
transcription of the three genes is analyzed (Fig. 2). An
inhibition ofmore thaneight times in the expression ofa22
was observed, andconsequently, the expression of the TK and -y0genes was also blocked. These results suggest that
the replication of HSV-1 in human fibroblasts can be
par-tially inhibitedby IFN-ywhenalow MOI is used.Therefore,
HSV-1 in this system somehow resembles the situation in mouse macrophages treated with MuIFN-,. These experi-ments clearly reinforce the idea that the final outcome of HSV-1 replication in agivencell typedepends not only on
thespecies ofIFNused but alsoonothervariables, suchas
theconcentrationofIFN,theMOI of theexperiment,andso
on (3, 11, 14, 19).
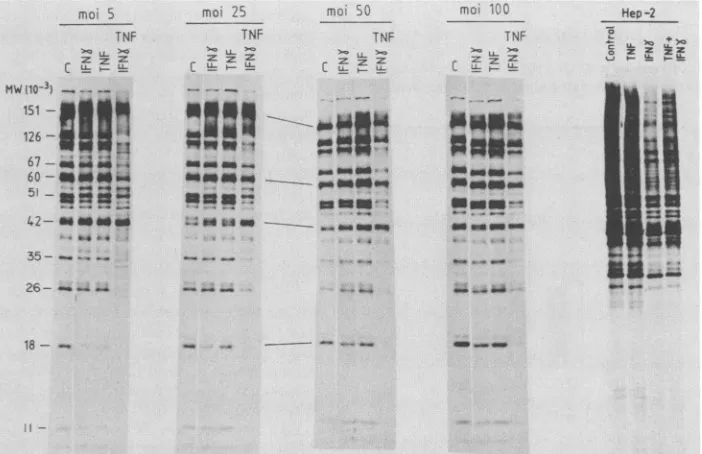
Synergistic effect ofTNF andIFN-yagainst HSV-1 replica-tion. The combined addition ofhuman TNF and HuIFN--y leadsto asynergistic inhibitionof HSV-1 replication (1, 30). TreatmentofHEp-2 cells with TNF alone, even at
concen-trations of 100 ng/ml, had no effect on HSV-1 protein
synthesis (Fig. 3). In agreement with the results shown in
Fig.1and2,neither IFN-a norIFN-yatlowconcentrations
(10U/ml) prevented the appearanceof HSV-1 late proteins.
Strikingly, the combination ofTNF and IFN--y fully
sup-pressed thesynthesisof HSV-1 proteins (Fig. 3), indicating
a synergistic action. This effect was not observed when
IFN-ot was added, further suggesting that the activities of IFN-a and IFN-y against DNA-containing viruses are not similar. Since the MOI used was an important factor
deter-mining the resistance or sensitivity of HSV-1 late-protein
synthesisto IFN--y, weanalyzed the effect of this factor on
HSV-1 translation (Fig. 4). Again, HSV-1 proteins were
on November 10, 2019 by guest
http://jvi.asm.org/
[image:3.612.178.464.71.314.2] [image:3.612.78.305.381.613.2]HSV-1 (16
hpi)
I1TNF
100ng/mI
IFN (1OU/ml) - a Y - CY
151-_ ef A
126
-
110-60- e4
saW-51 - __fnhI _w
42 -__ma_iiA
35 -_
26- ---
-
1811
-FIG. 3. Effectof IFN-a,IFN-y, and TNF treatment onprotein synthesis in HEp-2 cells infected with HSV-1. Cells were grown in 96-welldishes and treated with TNF (100 ng/ml), IFN-a (10U/ml), andIFN--y (10 U/ml) 24 h before infection with 25 PFU per cell. Cells were labeled at 16 to 18 h p.i. with [35P]methionine, and proteins wereanalyzed as described in Materials and Methods. -, Control without teatment. The numbersindicate molecular size in kilodaltons.
moi 5
TNF
>CLLv>
C ~L -U
moi 25
TNF
Z r :Z
c l
IFN (u/mi)
TNF lOOng/mt
O 10 1001000 0 10 100 1000
.
. *0 0**.
A
o o 0* 0 * . 0
-0* 0
* S
* .
FIG. 5. Synergistic effect of IFN--y andTNFon thereplicationof viral DNA. Cells were grown in60-mm dishes, treated with TNF andIFN-yattheindicatedconcentrations for 24 h, and infected with HSV-1 at an MOI of 25 PFU per cell. At 16 h p.i., total DNA was extracted andsubjected tohybridization to a nick-translated EcoRI TK fragment.
more sensitive to
IFN-y
and TNF when a low MOI was employed. However, therewasstillasignificant inhibition in the expression of several HSV-1 proteins, even when high MOIs of HSV-1were analyzed.To analyze the effect of TNF and IFN--y on viral DNA replication, total DNA from infected cellswasextracted 16 h afterinfection. Theamountof HSV-1 DNAwasquantitated byhybridization with a TK probe. Figure 5 shows that 10 U ofIFN--yperml alonedidnotdiminish theamountof HSV-1 DNApresent in infected cells.However, 100 and 1,000 U of
IFN-y per ml inhibited replication of the HSV-1 genome
four- andeightfold, respectively. Although by itself TNF is notinhibitory to HSV-1 DNA replication, the combination ofTNF and
IFN-y
clearly suppresses viral DNA replication. These results agree well with the findings obtained on viral protein synthesis and suggest that the step blocked by TNF plusIFN-y
is prior to HSV-1 DNA replication.TNF
r r- Z
MW110-31
67-^
60
"a;"i
"^_;42
-flu""
42- "..s - _
35-... -- 5--- _ __
MTO![
TNF
z -
-_
i P-U
___ra
__
Po_low
som _.
wa. __
___-soso
Xm
,..
a-26- -_
-18-
-Hep-2
>o
a z zz
'Ii
.-It
IIM"4o
a-_
I1
-FIG. 4. Synergistic effect ofIFN--y andTNF treatment onprotein synthesisin HEp-2cells infected with HSV-1. Cellswere grownin 96-well dishes and treatedwithTNF(100ng/ml)andIFN--y (10U/ml)for 24 hbeforeinfection.Cellswerelabeledfor2 hwith[35S]methionine, andproteinswereanalyzedasdescribedin Materials andMethods.C, Control.Thenumbers indicate molecularsize inkilodaltons.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:4.612.99.252.75.348.2] [image:4.612.360.510.76.195.2] [image:4.612.133.484.466.693.2]c2o2
TNF
r TNF IFN9IFN4
RNA
Spg I* *0 *
* * *
0 * #
TK
TNF
C TNF IFNX IFN9
RNA
s9g *
-.0
a 0
X Il
-tF rTNF IFNhIFN6 RNA
S"g 0
*-* *@e
00
FIG. 6. Synergistic effect of TNF and IFN-yontranscription of HSV-1genesin HEp-2 cells. TNF (100 ng/ml)and IFN (10 U/ml)
were added 24 h before infection. Conditions of infection, RNA extraction, and hybridization were as described in Fig. 2. C,
Control.
The transcription of HSV-1 genes ax22, TK, and -yO was
assayedasdescribed above.TNForIFN-y usedseparately
hadnegligible effectsonthetranscriptionofearlygenes cx22 (<x class)and TK(pclass), althoughboth of them when used aloneclearly inducedsomeinhibition(Fig. 6). However, the
combinationofTNFplus IFN--y preventedtheexpressionof early (ot22andTK)and late(-yO)genes.The conclusion from these results is that TNFplusIFN-yblocksanearlystepin HSV-1 replication that comes either at or prior to the transcription of earlygenes.
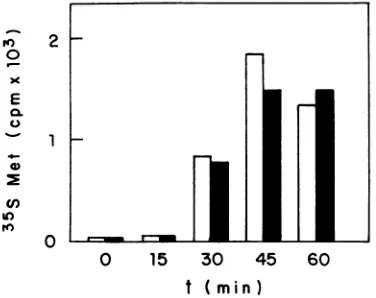
Todetermine whetherveryearly steps in HSV-1 replica-tion,suchasHSV-1bindingandentryinto HeLacells,were
affected by TNF and IFN-y, HSV-1 virions were labeled with [35S]methionine, and theentryof these virions into cells
was monitored at different times. HSV-1 entered control cells andHEp-2cells treated with TNF and IFN with similar kinetics andtoasimilarextent(Fig. 7).Theamountsof viral particles adsorbed touninfected cells at0°C with and with-out treatmentwith TNFandIFN--ywerealso similar(results
notshown). These results lead us to suggestthat HSV-1 is internalized in cells treatedwith the twocytokines and that thestepblockedbythemoccursafterentryandbeforeorat the level oftranscription of HSV-1 earlygenes.
DISCUSSION
The antiviral mode of action of HuIFN-cx against herpes-viruses isdifferent from that against other DNA viruses in
to0
x
E
0.
C.). 0)
(I
I')
to 2
1
0
0 15 30 45 60
t (min)
FIG. 7. Effect ofIFN--y and TNF on HSV-1 entry intoHEp-2
cells. Entryofradiolabeledvirions into untreatedcells(white bars)
orcells treated with 10 U ofIFN--ypermland 100ng of TNF per ml
(blackbars) wasmeasuredasdescribedinMaterials andMethods,
at thetimeindicated.
several respects. First,theproductionofinfectiousparticles
is significantly blocked by HuIFN-ot, but viralprotein
syn-thesis is notaffected (4, 17). The inhibition of virus release (4) and the production ofnoninfectious viral particles (17)
have been documented as the mode of action ofHuIFN-ax
against HSV-1 in human fibroblasts. Our present results
point out the differences that exist between the different
human IFN species with regard tothe inhibitoryeffects on HSV-1 in HEp-2 cells. Thus, IFN--y by itself shows some inhibition ofviral translation andtranscription, particularly
whenalowMOIis used.HigherdosesofHSV-1areableto overcome the inhibition by IFN. This experiment is of interest because it might helpto explain conflictingreports
fromdifferentgroups(4, 6,15, 17). Ourexperimentsarealso
consistent with the finding that high
IFN-P
doses preventprotein synthesisinHSV-1-infectedChangcells(3),because
in theHEp-2 system heredescribed, theaction ofIFN-, is similar to that ofIFN--y rather than to that of IFN-ot. Our
findingslendadditional support tothe view that human and
mouse IFNs show different effects against HSV-1.
There-fore, it is not surprising to us that MuIFN-a-, blocks
immediate-early gene transcription in mouse macrophages
(5).TheconclusionthatcanbemadeabouttheactionofIFN
againstHSV-1is thatit variesaccordingtothe system under
study ineachlaboratory (3, 4, 6, 17, 20).
Therefore,
HSV-1isratheruniqueinthisregardanddiffers frompicornaviruses
andrhabdoviruses, whichare moreconsistentlyinhibitedby
highdosesofIFN in thedifferentsystems
analyzed
(11,19).
HSV-1 is also at variancewithother DNA
viruses,
suchasvaccinia, because it is blocked by the combined action of
TNF and low doses of IFN--y, whereas vaccinia virus
translation continues to control levels in cells treated with
both
compounds (results
notshown).
Anotherinteresting
aspect ofthe present work is that TNF
by
itself has noinhibitory effectsonHSV-1. Thisresultdoesnotsupportthe
idea that TNF stimulates the
production
ofIFN-P2
in ourHEp-2cells,unlesswe assumethat
IFN-P2
plusTNF hasnoeffect on HSV-1.
Experiments
are now in progress in ourlaboratoryto
clarify
thispoint.
The mode of action of TNF plus IFN-y against HSV-1 resembles theinhibitionby IFN-a-1 of HSV-1 in themouse system. Inthemurinemacrophage model, ablockadeinthe
transcription oftheimmediate-earlygeneswas observed(5,
6). We have also shown here that the
expression
oftheimmediate-early gene aL22 is diminished in our system.
Keepingin mind that HSV-1 is internalizedincells,thereare
several possibilities to explainthe decrease in a22
expres-sion:
(i)
the HSV-1virionsentercells,butthey donotreachthe nucleus; (ii) decapsidation of HSV-1 virions is not
properly achieved; (iii)the HSV-1 DNA isproperly located
in thenucleus anddecapsidates well, butthe RNA
polymer-ase does not recognize the promoters of immediate-early
genes; and
(iv) immediate-early
HSV-1 mRNAs arerapidlydegraded. At present, we have no convincing evidence to
conclude anyoneofthesepossibilities. Ifpossibilitiesiii and ivare true, since neither thetranscription ofcellulargenes
northestabilityof cellular mRNAsseems tobeaffected,we
have to propose specific mechanisms acting on viral DNA
transcriptionor viral mRNAstabilityorbothtoexplainthe
absence ofimmediate-earlyviral mRNA in cells treated with
TNFplus IFN--y.
Much effort has been directed to control HSV-1 and
HSV-2 infections in humans by chemotherapy. In fact,
either IFN aloneorincombinationwithantiherpes drugshas been describedas apotential inhibitorof HSVinfections (16,
24). The finding that TNF acts in synergy with HuIFN-y
on November 10, 2019 by guest
http://jvi.asm.org/
[image:5.612.74.303.74.179.2] [image:5.612.99.287.528.677.2]opens the possibility of using these two agents in combina-tion against certain HSV-1 diseases.
ACKNOWLEDGMENTS
The excellenttechnicalassistance of M.Chorrois acknowledged. Wethank G. R. Adolf for his gift ofrHuTNF-a andrHuIF-y, N. Finter for his gift ofHuIFN-(Ly)-a, and E. Tabards for supplying plasmids.
E.F.istheholder ofaFPIfellowship. Comite Conjunto Hispano-Norteamericano, CAICYT, CSIC, andFIS are acknowledged for financialsupport.
LITERATURE CITED
1. Beutler, B., D. Greenwald, J. D. Hulmes, M. Chang, Y. C. E. Pan, J. Mathison, R. Ulevitch, and A. Cerami. 1985.Identityof tumor necrosis factor and the macrophage-secreted factor cachectin. Nature(London) 316:552-554.
2. Blin, N., and D. W. Stafford. 1976. Isolation of high-molecular weightDNA.Nucleic Acids Res. 3:2303.
3. Cayley,P.J., J. A. Davies, K. McCullagh, andI.M. Kerr.1984. Activation of the ppp(A2'p)nA system in interferon-treated, herpes simplex virus-infected cells and evidence for novel inhibitors of the ppp(A2'p)nA-dependent RNase. Eur. J. Bio-chem. 143:165-174.
4. Chatterjee, S., E. Hunter,andR.Whitley.1985. Effectofcloned humaninterferons on protein synthesis andmorphogenesis of herpessimplex virus. J.Virol. 56:419-425.
5. Domke,I., P.Straub, H. Jacobsen, H. Kirchner, and A. Paret. 1985. Inhibition of replication of herpes simplex virus inmouse macrophagesby interferons.J. Gen. Virol. 66:2231-2236. 6. Domke-Opitz, I., P. Straub, and H. Kirschner. 1986. Effect of
interferononreplicationofherpessimplexvirus types 1 and 2 in human macrophages. J. Virol.60:37-42.
7. Favoloro, J., R. Trisman, and R. Kamen. 1980. Transcription maps of polyoma virus-specificRNA: analysis by two-dimen-sional nucleaseS1 gel mapping. Methods Enzymol. 65:718-749. 8. Feduchi, E., and L. Carrasco. 1987. Adenovirus infection re-verses theantiviral state induced by human interferon. FEBS Lett. 214:153-157.
9. Feduchi, E., M. Esteban, and L. Carrasco. 1988. Reovirustype 3synthesizes proteins in interferon-treated HeLacellswithout reversing the antiviral state.Virology164:420-426.
10. Laemmli, U. K.1970.Cleavage of structural proteins duringthe assemblyof the head ofthebacteriophageT4. Nature(London) 227:680-685.
11. Lebleu, B., and J. Content. 1982. Mechanisms ofinterferon action: biochemical and genetic approaches, p. 47-94. In I. Gresser(ed.), Interferon4.Academic Press, Inc.,NewYork. 12. Maniatis, T., E. F. Fritsch, and J. Sambrook. 1982. Molecular
cloning: alaboratory manual. ColdSpring Harbor Laboratory, ColdSpring Harbor, N.Y.
13. Mestan, J., W. Digel, S. Mittnacht, H. Hillen, D. Blohm, A. Moller,H.Jacobsen, and H. Kirchner.1986. Antiviral effects of recombinanttumour necrosis factor in vitro. Nature (London) 323:816-819.
14. Mistchenko, A. S., and R. Falcoff. 1987. Recombinant human
interferon-yinhibits adenovirusmultiplicationin vitro. J. Gen. Virol. 68:941-944.
15. Mittnacht, S., P. Straub, H. Kirchner, and H. Jacobsen. 1988. Interferon inhibits onset of herpes simplex virus immediate-earlytranscription. Virology 164:201-210.
16. Moran, D. M., E. R. Kern, and J. C.Overall.1985. Synergism betweenrecombinanthumaninterferon andnucleosideantiviral agentsagainstherpessimplex virus: examinationwith an auto-matedmicrotiter plateassay.J. Infect. Dis. 151:1116-1122. 17. Munoz,A., and L. Carrasco. 1984. Formation of noninfective
herpesvirus particles in cultured cells treated with human inter-feron.J.Gen. Virol. 65:1069-1078.
18. Mufioz, A., and L. Carrasco. 1984. Action ofhuman lympho-blastoid interferononHeLacellsinfected withRNA-containing animal viruses.J. Gen. Virol.65:377-390.
19. Munoz,A., and L. Carrasco.1987.Effects of interferononviral replication and on the cytopathic effects induced by animal viruses, p. 147-188. In L. Carrasco (ed.), Mechanisms of viral toxicityinanimal cells. CRC Press, Inc., Boca Raton, Fla. 20. Oberman, F., and A. Panet. 1988.Inhibition of transcription of
herpessimplex virus immediate earlygenesininterferon-treated humancells.J. Gen. Virol.69:1167-1177.
21. Old, L. J. 1985. Tumor necrosis factor (TNF). Science 230: 630-632.
22. Paez, E., and M. Esteban. 1984.Resistance of vacciniavirus to interferonis relatedto aninterference phenomenon betweenthe virus and theinterferonsystem. Virology134:12-28.
23. Shalaby, M. R., B. B. Aggarwal, E. Rinderknecht, L. P. Suekerky,B. S.Finkle, and M. A. Palladino. 1985.Activation of human polymorphonuclear neutrophil functions by interferon--y
andtumornecrosisfactor. J. Immunol. 135:2069-2073.
24. Stanwick, T. L., R. F. Schinazi, D. E. Campbell, and A. J. Nahmias. 1981. Combined antiviral effect of interferon and acycloviron herpes simplex virus types 1and2. Antimicrob. Agents Chemother. 19:672-674.
25. Straub,P., J. Donke, H. Kirchner, H. Jacobsen, and A. Panet. 1986. Synthesis of herpes simplex virus proteins and nucleic acids in interferon-treated macrophages. Virology 150:411-418. 26. Sugarman,B.J., B. B.Aggarwal, P. E. Hass, I. S. Figari, M. A. Palladino, and H. M.Shepard.1985.Recombinanthumantumor
necrosisfactor-a: effectsonproliferation of normal and trans-formed cellsinvitro. Science 230:943-945.
27. Thomas, S. P. 1983. Hybridization of denatured RNA trans-ferred or dotted to nitrocellulose paper. Methods Enzymol. 100:255-266.
28. Vilcek, J., V. J. Palombella,D.Henriksen-Destefano, C. Swen-son,R. Feinman, M.Hirai,and M.Tsujimoto. 1986.Fibroblast growth enhancing activity of tumor necrosis factor and its relationshiptootherpolypeptide growth factors.J. Exp. Med. 163:632-643.
29. Wagner, E. R. 1985. IndividualHSVtranscripts: characteriza-tionofspecificgenes, p. 45-104. In B. Roizman(ed.), Herpes-viruses,vol. 3. PlenumPublishing Corp.,NewYork.
30. Wong, G. H. W., and D. V. Goeddel. 1986. Tumor necrosis factors a and ,B inhibit virus replication and synergize with interferons. Nature(London)323:819-822.