JOURNALOFVIROLOGY,Apr. 1992,p. 2564-2569 0022-538X/92/042564-06$02.00/0
CopyrightC1992, American Society for Microbiology
Human
and
Mouse Mx
Proteins Inhibit Different
Steps of the
Influenza Virus Multiplication Cycle
JOVANPAVLOVIC,l* OTTO HALLER,2 ANDPETER STAEHELI2InstituteforImmunology and Virology, Universityof Zurich, CH-8028 Zurich, Switzerland,1and
Department of Virology, University of
Freiburg,
D-7800Freiburg,
Germany2Received 15 October 1991/Accepted13 January 1992
Human MxA and mouse Mxl are interferon-induced proteins capable of inhibiting the multiplication of
influenza virus. MxAprotein is localized in the cytoplasm, whereas Mxl protein accumulates in the nucleus.
Taking advantage ofstablytransfected cell lines thatconstitutively express eitherMxAorMxl protein,we
examined thesteps atwhich these proteins block influenza A viruses.Ininfectedcells expressing MxA protein,
all viral mRNAs synthesized as aresultofprimarytranscription in the nucleus by the virion-associated RNA
polymeraseaccumulatedtonormal levels.Theseprimaxyviral transcriptswerepolyadenylated,wereactive in
directing viral protein synthesisinvitro, andappearedtobe
efficiently
transportedtothe cell cytoplasm. Yetviral protein synthesis and genome amplification were strongly inhibited, suggesting that MxA protein
interferedwith either intracytoplasmictransportof viral mRNAs, viral protein synthesis, ortranslocationof
newly synthesized viral proteins tothe cell nucleus. However, in infected cells expressing Mxl protein, the
concentrations ofthe longestprimarytranscripts encodingthe threeinfluenzavirus polymerase proteins PB1,
PB2, and PAwereatleast50-foldreduced.Accumulationof the shorterprimarytranscripts encoding the other
viral proteinswasalso inhibited buttoalesserextent.Theseresults demonstrate that themouseMxlprotein
interferes with primary transcription of influenza virus in the nucleus, whereas the human MxA protein
inhibits a subsequentstepthatpresumablytakes placeinthe cytoplasmof infected cells.
Influenza virus multiplication is highly sensitive to the antiviral action of interferon (IFN), which is mediated by IFN-inducedproteins. Despitenumerousefforts, the precise step at which influenza virus multiplication is blocked in IFN-treated cells has beendifficulttoestablish, both inhibi-tion of virus transcription and inhibition of viral mRNA translation being reported (3, 26). Influenza virus is a seg-mentednegative-strand RNAvirus witha complex
replica-tion strategy (for a review, see reference 14). The virus penetrates the host cell via receptor-mediated endocytosis afterwhich membranefusiontakes place (forareview, see reference 17). After uncoating, the individual viral
nucleo-capsids migrate rapidly to the cell nucleus (18), where the
parental viral genome is transcribed into mRNAs by the
virion-associatedRNA-dependent RNApolymerase, a pro-cessknownasprimary transcription. Influenza virus mRNA
synthesis is dependent on mRNA caps that are recruited from cellular pre-mRNAs (25). Genome replication also takes place in the nucleus of infected cells (28) and is initiated after translation of the primaryviral mRNAs and transport of newly synthesized viral proteins to the cell nucleus (14). Viral RNAs of negative polarity complexed
with viralproteinsareexported tothecytoplasm and
even-tually packaged.Thematurevirions bud fromplasma mem-branes of the host cell.
In mice, the antiviral state induced by IFNs against
influenza virus is controlledbythe hostgene,Mxl (8, 9, 29).
The IFN-inducedAMxgeneproduct (11) isanuclearprotein (7)that is sufficienttoconfer selective resistancetoinfluenza
virus (23, 24, 32). Comparing IFN-treated cellscarryingor
lackingfunctionalAMxgenes,weand others havepreviously shown that Mxl doesnotinterfere with host cellpenetration
oruncoatingof influenza virus(10, 20). Furthermore,
trans-*Correspondingauthor.
portofparentalinfluenza virusnucleocapsidstothe host cell nucleus was not inhibited by Mxl (4). However, further analysis of the precise step inhibited by Mxl produced conflictingresults. Krug and coworkers (15) reported that
Mxl inhibited influenza virus mRNA synthesis in IFN-treated cells. Others concluded from similar studies that Mxl inhibited translation of influenzavirus mRNAs(20).
MxA is an IFN-induced cytoplasmic protein of humans
that confers resistance to influenza virus and vesicular stomatitis virus (VSV) (1, 23, 24, 31). We recentlyshowed that MxA inhibits mRNA synthesis of VSV (33), but the mode of MxA action toward influenza virus remains unclear. To determine the influenza virus multiplication steps
blockedbyhuman MxA andmouseMxl proteins, wetook
advantageofpermanentlytransfected 3T3 cells thatexpress high levels of the different Mx proteins in a constitutive manner (24). We show that mouse Mxl interfered with influenza virus mRNA synthesis. Human MxA inhibits a step that follows primary transcription butprecedes
ampli-fication of the influenza virusgenome.
Infected cellsexpressingMxlorMxAcontain reduced levels of influenza virusproteins.Wepreviouslydemonstrated that permanently transfected Swiss 3T3 cells constitutively
ex-pressinghuman MxAormouseMxlprotein acquired resis-tance toinfluenza virus infection. Incontrast,cells
express-ing human MxB protein remained fully susceptible to
influenza virus (24). We havenowdetermined the levels of influenza virus proteins in infected cells expressing the different Mx proteins. Monolayers of several independent
cell clonesexpressingMxl, MxA, orMxBprotein (12, 23a, 24)were infected with 3 PFU ofinfluenza A virus FPV-B (13)percell for 30minat25°C (24).Thevirus inoculumwas removedby two washings with phosphate-buffered saline,
and the cells were further incubated in growth medium at 37°C. At 5 h postinfection, the cells were harvested and proteinextractswereprepared by lysingthe cells inprotein
2564
Vol. 66,No. 4
on November 9, 2019 by guest
http://jvi.asm.org/
NOTES 2565
neo Mxl MxA
1 CO N D C U) U)
o ne r-t1o c
kDa C\N CO - CM C CULU) U)
200 -a
116-*
97
7-MxB m
r6
(,6
1-d UncnC_ -HA
66 _
45 -w
--HAl
_
____
~~~~.
2 3 4 5 6 7 8 9 10 1l 1213
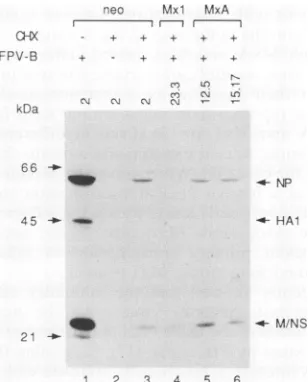
FIG. 1. Western blot analysis of influenza virus proteins in
infected3T3 cells constitutively expressing different Mx proteins. Confluent monolayers ofpermanently transfected Swiss 3T3cells wereinfected with 3 PFU of influenzavirusFPV-Bpercell, and cell extracts were prepared at 5 h postinfection. Infected cell clones expressing eithernoMxproteins (lanes1and2), Mxl (lanes 3to6), MxA(lanes 7to10),orMxB (lanes 11 and 12)wereimmunostained
withamouseantiserum directed against influenza virusA/Turkey/
England/63. Extract of uninfected cells servedas acontrol (lane 13). Thebandscorresponding to theviral proteins HA, NP, HAl, M, andNSareindicated. Therelative positions of marker proteinsare depictedatthe left.
gel samplebuffer(16). Samplesof theseproteinextracts(100 ,ugof protein per lane) were electrophoresed through 12%
polyacrylamide gels (16), and the levels of viral proteins
wereassessed by the Western immunoblotting technique (1), using a serumofICR mice immunized with influenza virus
A/Turkey/England/63, which recognizes the viral proteins NP,HA, M,andNS.Infectedcells expressing MxlorMxA
contained muchlowerlevels of HA, HAl, and NP than did infected control cellslacking Mx proteinsorcells expressing MxB (Fig. 1). Curiously, ratherstrong M/NS signals were detected inmostcell clonesirrespective ofthepresence or absence of Mx proteins, except for cell clone 10.2, which expressedveryhigh levels of Mxl(12) and which contained very low levelsof all influenza virus proteins, including M
andNS (Fig. 1).
Immunostaining of the Western blot with a monoclonal
antibodyto neuraminidase (NA) showed that accumulation
of this influenza virus protein was strongly reduced in all clonal cell linesexpressing Mxlor MxA (data notshown). Similarly, specific polyclonal antisera revealed that the
lev-elsof the influenza virus polymerase proteins PA and PB1 were much lower in cells expressing Mxl or MxA than in controlcells lacking Mxproteins (data notshown).
Influenza virus mRNA synthesisis inhibited bymouseMxl but not by human MxA protein. Amplificationofthe influ-enzavirusgenomeininfected host cells isdependenton de novo synthesis of viral proteins. In cells with blocked
protein synthesis, theviral multiplication cycle is arrested afterprimarytranscription, which iscatalyzed by the RNA
polymeraseassociatedwith theparental virions. Therefore, the extent of primary transcription can be determined by
monitoring the accumulation of viral mRNAs in infected cells treated withthe protein synthesis inhibitor cyclohexi-mide (CHX).
Wesimultaneously measured the accumulationof
primary
viraltranscripts(Fig.2A) andtotal viral RNAs(Fig. 2B)ina panelof influenzavirus-infected cellclonesexpressingeither MxlorMxA.Twoparallelcultureswerepreparedfrom each clonal cell line. One culturewas treatedwith50 p,g of CHXpermlfor 45 min before infectionwith influenza virus. This
concentration of CHX, which caused minimal cytotoxic
effects and inhibitedproteinsynthesisof these cell clonesby more than98% (33), wasmaintainedin the culture medium
throughout theexperiment.Thesecond culturewaskeptin medium lacking CHX. Both cultures were infected with 3 PFUof influenza virus FPV-B per cell. At 3 h
postinfection,
the CHX-treated cultures were harvested. The infected cultures kept in CHX-free medium were harvested at 4 hpostinfection. Total cellular RNA was prepared from the cultures according to the procedure of Chomczynski and
Sacchi (6), and samples were separated in 1.2% agarose-formaldehyde gels and subjected to Northern (RNA) blot
analysis (32). To distinguish between viral
primary
tran-scripts and genomic RNA from the parental virion, we hybridized the blots with radiolabeled in vitro-transcribed negative-senseRNAprobesderivedfromsevenof theeight influenza virus segments (Fig. 2A). The cDNAfragments
coding forHA, NP, M, and NS ofinfluenza virus
A/FPV/
R/34 were derived from plasmids pFPV-HA, pFPV-NP,pFPV-M, and pFPV-NS (kindly provided by C. Scholt-issek).The cDNAfragmentscodingfor the threepolymerase proteins PB1, PB2,and PAofinfluenzavirus
A/PR/8/34
werederived from plasmids pAPR102, pAPR206, and pAPR303 (36). All cDNA fragments were inserted into the multiple cloningsite of thevectorpBS(-)KS(Stratagene). Negative-sense RNA transcriptswere synthesized invitro, usingT3 RNA polymerase
(Stratagene)
and[(t-32P]UTP.
Total viral RNAfromindividualviral genes(Fig. 2B)wereprobedwithnick-translated influenzavirus cDNAfragments. The
inten-sityof thehybridization signalswasquantitated by densito-metricscanningof theautoradiograms.
Probesderived from the NA gene of influenza virus strains
A/FPV/Rostock/34 andAlParrot/Ulster/73 (kindly provided by C. Scholtissek) yielded only very weak hybridization signals with RNA from infected control cells (results not shown). Monitoring of primary NA mRNA synthesis was
thereforenotfeasible.
Ininfected cells expressing mouse Mxl protein, the
pri-mary transcript levels of the three polymerase genes PB1, PB2, and PAwere at least 50 times lower than in infected controlcellslackingMxproteins (Fig.2A, lanes 3 to6).The
primary transcript levels of the HA and NP genes were reduced about15-fold, whereasthe levels of theMand NS
primarytranscripts were only slightly reduced (3- and 2-fold,
respectively). As expected from the results and from the viral protein analysis presented above, total viral RNA
synthesis was strongly inhibited in cells expressing Mxl
protein (Fig. 2B). In contrast, infected cells expressing human MxAprotein contained high levels of primary PB1, PB2,PA, HA, and NP transcripts,comparable to the levels insusceptible control cells (Fig. 2A, lanes 7to10). Surpris-ingly, the levels of the M and NS primary transcripts were
evenhigher (about two- and fourfold, respectively) in cells
expressingMxAthan in cells lacking Mx proteins.
Simulta-neousmonitoring of total viral RNAs (Fig. 2B) showed that all four clonal cell linesexpressingMxAaccumulated much less total viralRNAthan didsusceptible control cells. This result demonstrated that despite unhindered primary tran-scription, influenza virus could not replicate in
MxA-ex-pressing cells. Cells expressing MxA protein contained VOL.66, 1992
on November 9, 2019 by guest
http://jvi.asm.org/
[image:2.612.66.287.73.257.2]2566 NOTES
A
M\A
..,A-
-<'C- ~ C. ~. 1)
-C- r' nS.i CL..
FIB 1-0 *@
@-D
PB?- a.
A -_ a
IHA
-B -e
PBr
-* e O i PA
-.-* * * PA - _
o
40is
*0
HA4FA
-11,NP - 3't *@.@@ NP
,NaA
-NB
"""j§h-
'
NS_
UObSUSWUWU
e 3 t - 'I
2 3 4 5 6 7 8 9 10
FIG. 2. Accumulationofinfluenza virus transcripts in infected cells expressing mouseMxlorhuman MxAprotein.Two parallelcultures of the indicated3T3 cellclonesexpressing either no Mx proteins (lanes 1 and 2),Mxl(lanes3to 6), orMxA(lanes 7 to 10)wereinfectedwith 3PFU of influenzavirus FPV-B per cell. To measure primary viral genome transcription (A), infection was carried out for 3 h in thepresence of50jigofCHXperml.To measureoverallviral RNA synthesis (B), infection was carried out for 4 h in theabsence ofCHX.TotalRNA wasprepared fromeach culture; samples were separated in 1.2% agarose-formaldehyde gels and subjected toNorthernblot analysis. For the primary transcriptionexperiments,15-jgsamples of the different RNAs were loaded into each lane, and the Northern blots were hybridized with radiolabeled, negative-sense RNA probes derived from the indicated influenza virus genes. The blots were exposed to X-ray films for 72 h at-70°C,using intensifying screens. For overall viral RNA analysis,5-jLgsamples of the different RNAs were loaded into eachlane, and the Northern blotswerehybridized with radiolabeled cDNA probes of the indicated influenza virus genes. The blots were exposedto X-rayfilms for2h(M andNS), 4 h (HA and NP), or 8 h(PB1, PB2,and PA) at room temperature.
significantly less viral RNA than did cells expressing Mxl
protein (Fig. 2B). This effect is due to the fact that
MxA-expressing cell clones arevery stable and represent more
homogeneous cell populationsthan is true for
Mxl-express-ing cell clones, which always contain a low percentage of
cellsexpressinglittle or noMxlprotein.
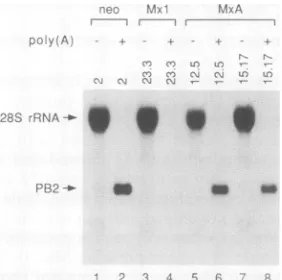
Primary viral transcripts from cells expressing MxA are polyadenylated and can direct protein synthesis in vitro.
Unhindered accumulation of primary viral transcripts in
cells expressing MxA did not necessarily mean that these RNAs were functional. We therefore tested whether the
primaryviral RNAs fromMxA-expressing cells were
poly-adenylated.Total cell RNAs from the experiment shown in
Fig. 2Awere fractionated into the poly(A)+ and
poly(A)-fractions by oligo(dT)-cellulose column chromatography. Northern blots containing these RNAs were probed with
radiolabeled, negative-sense RNA transcribed in vitro from influenza virus PB2 cDNA. The PB2 primary transcripts fromsusceptiblecontrol cells werefound almost exclusively in the poly(A)+ fraction (Fig. 3, lanes 1 and 2). The PB2
primary transcripts from Mx.A-expressing cells were also present in the poly(A)+ fraction (Fig. 3, lanes 5 to 8),
indicatingthat they contained a normal poly(A) tail.
Simi-larly,the small amounts of PB2 primary transcripts detected in Mxl-expressing cellswere found in the poly(A)+ RNA
r n r --..._
CM CMx M-A
P28y.A- 9 9 . S
28S
rRQNA_
PB[?- fl m
-O 3 4 6
FIG. 3. EffectofMxAonthepolyadenylationof influenza virus primary transcripts. The indicated cell clones expressing no Mx proteins (lanes1and2),Mxl (lanes3and4),orMxA(lanes5 to8) weretreated with 50 jigofCHX per mland infectedfor 3 hwith3 PFU of influenza virus FPV-B percell. Total RNAwasprepared from each culture, and 40-jig samples were fractionated by oli-go(dT)-cellulose chromatography. The poly(A)- RNA fractions (lanes 1, 3, 5,and7)andthepoly(A)+ RNAfractions(lanes 2, 4, 6, and 8) were separated in a 1.2% agarose-formaldehyde gel and subjected to Northern blot analysis. Radiolabeled negative-sense PB2 RNA wasusedas ahybridization probe.Thepositionsof PB2 mRNAandcross-hybridizing28S rRNAareindicated.
J. VIROL.
on November 9, 2019 by guest
http://jvi.asm.org/
[image:3.612.165.477.70.355.2] [image:3.612.372.513.474.614.2]neo Mxl MxA
I fml I I
CHX - + + + + +
FPV-B + - + + + +
neo MxA
C N C N
kDa
NP
-_-66
-0-45 -_
- NP
-HA1
21
-_-M/NS
2 3 4 5 6
FIG. 4. Effect ofMxA on the in vitro translation capacity of influenza virus primary transcripts. The indicated cell clones ex-pressingnoMxproteins (lanes1to3), Mx1 (lane 4),orMxA(lanes
5and 6)weretreated with 50jigofCHX permland infected for 3 h with 3 PFU of influenza virus FPV-B percell. Total RNAwas prepared from each culture and fractionatedby oligo(dT)-cellulose chromatography. Samples (0.5 ,ug) ofpoly(A)+ RNA were
trans-lated invitro, usingrabbitreticulocyte lysateand [35S]methionine. Poly(A)+ RNAs from infected control cellsnot treated with CHX
(lane 1)and from uninfected cells(lane 2)wereincludedascontrols. The in vitro-translated viralproteinswereimmunoprecipitatedwith
a mouse antiserum to influenza virus A/Turkey/England/63. The
bandscorrespondingtotheviralproteinsNP, HAl, M,andNSare
indicated. The relativepositionsof markerproteins aredepictedat
the left.
fraction(Fig. 3,lanes 3 and4).Thestrongsignalobserved in all
poly(A)-
RNAfractions isdueto cross-hybridization of28SrRNA with the PB2 RNAprobe.
Wenextexaminedwhether the viral mRNAs could direct
protein synthesis in vitro. Samples of0.5 jig of poly(A)+
RNAs were translated in vitro, using rabbit reticulocyte lysate (Promega) in the presence of [3S]methionine. Viral
proteinswerethen immunoprecipitated (19) with an antise-rum of mice immunized with inactivated influenza virus
A/Turkey/England/63. The immunoprecipitated translation products were analyzed on a 10% polyacrylamide-sodium
dodecyl sulfate gel (30). Similar levels of radiolabeled NP were detected irrespective ofwhether the templates were mRNAs from CHX-treated control cells (Fig. 4, lane 3) or mRNAs from CHX-treated cells expressing MxA (Fig. 4,
lanes 5 and 6). The M and NS proteins were produced at
slightly higherlevels withtemplatesofMxA-expressingcells than withtemplatesofcontrol cells.Onlyverylowlevels of NP protein were observed when poly(A)+ RNA of Mxl-expressing cells was used as the template (Fig. 4, lane 4),
whereas lowbutsignificant amountsof M andNS proteins
were synthesized from such RNA. Thus, the relative
amounts of viral proteins synthesized in vitro reflected the
relativeamountsof thecorresponding primary transcripts in
the different celllines, suggesting that the primaryinfluenza
virustranscripts fromMxA-expressing cells could be
trans-lated normally.
Primaryinfluenza viral transcripts accumulate in the
cyto-plasmofMxA-expressingcells. Inpermissive cells, influenza
virus mRNAs are efficiently transported to the cell
[image:4.612.105.259.73.264.2]cyto-1 2 3 4
FIG. 5. Intracellulardistributionofinfluenza virusprimary tran-scripts in infected cells expressing MxA protein. CHX-treated control cells (neo, clone 2; lanes 1 and 2) and CHX-treated cells expressingMxA(MxA, clone5.5.3;lanes3 and4)wereinfected for 3 h with3 PFU ofinfluenza virus FPV-Bpercell.Samples(15 ,ug)
of RNAs isolated from the cytoplasmic (C; lanes 1 and 3) and nuclear(N; lanes2and4) fractionsweresubjectedtoNorthernblot analysis. Radiolabeled negative-sense NP RNA was used as a
hybridization probe. Theposition of theNP RNAisindicated.
plasm (28). To testwhether MxA affected the transport of viral mRNAs, we analyzed the intracellular distribution of primary influenza virustranscriptsin infected cells express-ing MxA. Susceptible control cells and MxA-expressing cellswereinfected with3 PFU of influenza virus FPV-B per cell in the presence of CHX. At 3 h postinfection, total RNAs were prepared from the cytoplasmic and nuclear fractions of each culture, and the RNAswere analyzed by the Northern blottingtechnique. For the isolation of
cyto-plasmic and nuclear RNAs, the cells were lysed in lysis
buffer(10 mMTris-hydrochloride[pH7.5],10 mMNaCl,1.5 mMMgCl2, 0.5%NonidetP-40)for 5 minonice. The nuclei
weresedimented for 5 minat1,500 xg,andthe supernatant was designated the cytoplasmic fraction. The nuclei were
washed once in lysis buffer. Sodium dodecyl sulfate and EDTA (final concentrations of 0.4% and 10 mM,
respec-tively)were addedto bothfractions, and nuclear and
cyto-plasmic RNAs were isolated as previously described (22, 32).Thepurityof the nuclear andcytoplasmic fractionswas
monitored as described by Shapiro and coworkers (28). Radiolabelednegative-senseNPRNAwasusedas a hybrid-ization probe. The primary NP transcripts were detected
exclusivelyin thecytoplasmic fractionsof bothcontrol cells
and cellsexpressing MxA(Fig. 5). Reprobing of the
mem-brane with radiolabeledpositive-sensePB2 RNAyieldedno signals, indicatingthat the sensitivity ofthis assaywas not high enoughtodetect thegenomicRNAof theparentalvirus
(datanotshown).
Conclusions. We have recently shown that constitutive
expressionofhuman MxA andmouseMxlproteininstably
transfectedmouse3T3 cellsconferredresistancetoinfluenza virusinfection(24). Taking advantage of these cell lines,we
nowexamined which steps of theinfluenzavirus multiplica-tion cycle were blocked by these two proteins known to
accumulate indifferentsubcellular compartments. We show here that the nuclear mouse Mxl protein inhibited viral mRNA synthesis, whereas the cytoplasmic human MxA protein blocked amplification of the viral genome without
affectingprimary transcription.
Previous studies aimed at elucidating the influenza virus
multiplication stepsblocked by Mxl protein in IFN-treated cells generatedconflictingresults. UsingIFN-induced
on November 9, 2019 by guest
http://jvi.asm.org/
[image:4.612.397.483.74.198.2]2568 NOTES
rophages of Mx- and
Mx+
mouse strains, an initial study presented evidence suggesting that the Mx block was at the level of translation of viral mRNAs (20). In contrast, subse-quent findings with IFN-induced mouse embryo fibroblasts indicated that Mxl inhibited virus multiplication at the level of primary RNA synthesis (15). The differences, although difficult to reconcile, could have been due to the different cell types that were used or to additional IFN effects superim-posed on the Mx effect. The present study with transfected 3T3 cells constitutively expressing Mx proteins clearly indi-cates thatMx1
protein indeed inhibits the multiplication of influenza virus at the level of primary transcription. Accu-mulation of the primary transcripts of individual influenza virus genes was not uniformely inhibited by Mx1 protein. The degree of inhibition correlated with the length of the genomic segments. The levels of the longest primary tran-scripts coding forPB1, PB2, and PA were reduced at least 50-fold in infected cells expressing Mx1,whereas the levels of the shortest primary transcripts coding for M and NS were reduced at best 2- to 3-fold (Fig. 2A).Mxl protein interferes with an important step of viral multiplication, since the production of polymerase mRNAs in the cell nucleus and subsequent synthesis of polymerase proteins are absolutely essential for amplification of the viral genome. Our experiments do not allow us to distinguish between decreased stability of primary transcripts and re-duced synthesis of primary transcripts. However, the ob-served reduction of viral RNA synthesis in the presence of
Mx1 (15) argues in favor of an inhibition of the transcrip-tional activity of the viral polymerase byMx1. The fact that the extent of the inhibition of RNA accumulation correlated with segment length (Fig. 2A) (15) suggests that chain elongation, rather than initiation, is the target ofMxl action. Premature chain termination is expected to preferentially
affect the transcripts of longer genes, and incomplete RNA transcripts lacking apoly(A) tail are usually very unstable. Interestingly, MxAprotein has a similar inhibitory effect on transcription of the VSV genome. We found that in
VSV-infected cells expressingMxAprotein, the relative levels of the longprimary transcriptswere reduced muchmore
dra-matically than were levels of the short transcripts (33).
In contrast to Mxl, the human MxA protein has no inhibitory effect on primary transcription of influenza virus. The primary transcripts of influenza virus accumulating in infected cells expressing MxA protein were polyadenylated and able to direct protein synthesis in vitro. Furthermore,
these RNAs appeared to be efficiently transported to the cytoplasm. The precise step at which influenza virus is blocked byMxA protein remains undefined. However, our results indicate that MxA protein interfered either with access of the viral mRNA to the protein synthesis machin-ery, with viral protein synthesis, with transport of newly synthesized viral proteins back to the cell nucleus, or with viral genome replication. Because MxA-expressing cells readily synthesized influenza virus proteins after transfec-tion with plasmid vectors encoding influenza virus proteins (21a), protein synthesis is presumably not theMxA-sensitive step. Also, viral RNA replication appears to be anunlikely target of MxA action, sinceMxA is a cytoplasmic protein and thus presumably blocks a multiplication step occurring in the cytoplasm. In the light of recent findings that Mx proteins are homologous to a family ofproteinsinvolved in microtubule-dependent intracellular protein transport (2, 5, 21, 27, 34, 35), it is possible that MxA protein interferes either with the intracytoplasmic transport of viral RNAs to
ribosomes or with translocation of newly synthesizedviral proteins from thecytoplasm to the nucleus.
Although MxA and Mxl inhibit different steps of the influenza virusmultiplication cycle, the two proteins could still exert their function by a common mechanism. The
differences reported here might simply stem from the fact that MxA and Mxl are localized in different subcellular compartments. Recent experiments indicate that thismight indeed be the case (37).When moved to the cell nucleus with the help of a foreign nuclear translocation signal, human MxA proteinretained its activityagainst influenza virus. But unlike the cytoplasmic MxA protein, the nuclear form of MxA blocked primary transcription of influenza virus,
thereby mimickingmouse Mxlprotein.
We recently showed that the inhibitory effect of Mxl
protein against influenza virus co-uld be neutralized by overexpression of the threeviralpolymeraseproteinsandto
a lesser extent by PB2 alone (12), suggesting that Mxl can
physically interact orfunctionally competewith components of theinfluenza viruspolymerasecomplex, therebyblocking
its normal function. It remains to be determined whether overexpression of proteins of the influenza virus RNA polymerase complex can alsoneutralize theinhibitoryeffect of themutant MxA protein in the nucleus and thewild-type
MxAprotein in thecytoplasm.
We thank C. Scholtissek for providing plasmids carrying avian influenza virus cDNAs, J. Lindenmann, L. Bazzigher, F. Pitossi,
andT.Zurcherforhelpfuldiscussions,andD. Bucher for excellent technicalassistance.
This work was supported by grants from the Swiss National ScienceFoundation andbytheCantonofZurich.
REFERENCES
1. Aebi, M., J. Fah, N. Hurt, C. E. Samuel, D. Thomis, L. Bazzigher, J. Pavlovic, 0. Haller, and P. Staeheli. 1989. cDNA structures andregulation oftwointerferon-induced human Mx proteins. Mol. Cell. Biol. 9:5062-5072.
2. Arnheiter, H., and E. Meier. 1990. Mx proteins: antiviral proteins by chance orbynecessity. NewBiol. 2:851-857. 3. Bean, W. J., andR. W. Simpson. 1973. Primarytranscriptionof
theinfluenza genome in permissive cells. Virology 56:646-651. 4. Broni, B.,I.Julkunen, J. H.Condra,M.-E.Davies,M.J.Berry, andR. M. Krug. 1990. Parental influenza virionnucleocapsids are efficiently transported into the nuclei of murine cells ex-pressing the nuclear interferon-induced Mx protein. J. Virol. 64:6335-6340.
5. Chen, M. S., R. A.Obar, C. C. Schroeder,T. W.Austin, C. A. Poodry, S. C. Wadsworth, and R. B. Vallee. 1991. Multiple forms ofdynamin are encoded byshibire, aDrosophila gene involved in endocytosis. Nature(London) 351:583-586. 6. Chomczynski, P., and N. Sacchi. 1987. Single-step method of
RNAisolation byacidguanidinium thiocyanate-phenol-chloro-formextraction. Anal. Biochem. 162:156-159.
7. Dreiding, P., P. Staeheli, and 0. Haller. 1985. Interferon-induced protein Mx accumulates in nuclei of mouse cells expressing resistance to influenza viruses. Virology 140:192-196.
8. Haller, 0. 1981. Inborne resistance of mice to orthomyxovi-ruses. Curr. Top. Microbiol. Immunol. 92:25-52.
9. Haller, O., H. Arnheiter, J.Lindenmann, and I. Gresser. 1980. Host geneinfluences sensitivitytointerferon actionselectively forinfluenzavirus. Nature (London)283:660-662.
10. Horisberger, M. A.,0.Haller,and H.Arnheiter. 1980. Interfer-on-dependentgenetic resistancetoinfluenza virusin mice:virus replication inmacrophagesis inhibitedat anearly step.J. Gen. Virol.50:205-210.
11. Horisberger, M.A., P. Staeheli,and0. Haller. 1983. Interferon induces a unique protein in mouse cells bearing a gene for resistance to influenza virus. Proc. Natl. Acad. Sci. USA J. VIROL.
on November 9, 2019 by guest
http://jvi.asm.org/
80:1910-1914.
12. Huang, T., J. Pavlovic, P. Staeheli, and M. Krystal. Submitted forpublication.
13. Israel, A. 1979. Preliminary characterisation of the particles from productive and abortive infections of L cells by fowl plague virus. Ann. Microbiol. (Inst. Pasteur) 130B:85-100. 14. Krug, R. M., F. V. Alonso-Caplen, I. Julkunen, and M. G.
Katze. 1989. Expression and replication of the influenza virus
genome, p.89-152. In R.M.Krug (ed.), The influenza viruses.
PlenumPublishingCorp., New York.
15. Krug, R. M., M. Shaw, B. Broni, G. Shapiro, and0. Haller.
1985. Inhibition of influenza viral mRNA synthesis in cells expressing the interferon-induced Mx gene product. J. Virol. 56:201-206.
16. Laemmli, U. K. 1970. Cleavage of structural proteins during the assembly of the head ofbacteriophage T4. Nature (London) 227:680-685.
17. Marsh, M., and A. Helenius. 1989. Virusentryintoanimal cells. Adv. Virus Res.36:107-151.
18. Martin, K., and A. Helenius. 1991. Transport of incoming influenzavirus nucleocapsids into the nucleus. J. Virol. 65:232-244.
19. Meier, E., J. Fah, M. S. Grob, R. End, P. Staeheli, and 0.
Haller. 1988. A family of interferon-inducedMx-related mRNAs encodescytoplasmic and nuclear proteins inratcells.J. Virol. 62:2386-2393.
20. Meyer, T., and M. A. Horisberger. 1984. Combined action of
mouse aand 13 interferons in influenza virus-infected
macro-phages carrying the resistancegene Mx. J. Virol. 49:709-716. 21. Obar, R. A., C. A. Collins, J. A. Hammarback, H. S. Shpetner,
and R. B. Vallee. 1990. Molecularcloning of the microtubule-associated mechanochemical enzymedynamin reveals
homol-ogywitha newfamily of GTP-binding proteins. Nature
(Lon-don) 347:256-261.
21a.Pavlovic, J. Unpublisheddata.
22. Pavlovic, J., E. Banz, and R. W. Parish. 1989. The effects of transcriptiononthe nucleosomestructureof fourDictyostelium
genes.NucleicAcids Res.17:2315-2332.
23. Pavlovic, J., and P. Staeheli. 1991. Theantiviral potentials ofMx
proteins.J. Interferon Res.11:215-219. 23a.Pavlovic, J., and T. Zurcher.Unpublished data.
24. Pavlovic, J., T. Zurcher, 0. Haller, and P. Staeheli. 1990.
Resistance to influenza virus and vesicular stomatitis virus
conferred by expression of human MxA protein. J. Virol.
64:3370-3375.
25. Plotch, S. J., M. Bouloy,I.Ulmanen, and R. M. Krug. 1981. A
uniquecap(m7GpppXm)-dependent influenza virion endonucle-asecleavescappedRNAstogeneratetheprimers that initiate viral RNAtranscription. Cell 23:847-858.
26. Repik, P., A. Flamand, and D. H. L. Bishop. 1974. Effect of interferon upon the primary and secondary transcription of vesicular stomatitis and influenza viruses. J. Virol. 14:1169-1178.
27. Rothman, J. H.,C. K. Raymond, T. Gilbert, P. J.O'Hara,and T. H. Stevens. 1990. Aputative GTPbinding protein homolo-gous to interferon-inducibleMx proteins performsanessential functionin yeastprotein sorting. Cell 61:1063-1074.
28. Shapiro,G.I., T.Gurney,Jr., and R. M. Krug. 1987.Influenza virus gene expression: control mechanisms at early and late times of infection and nuclear-cytoplasmictransport of virus-specificRNAs. J.Virol. 61:764-773.
29. Staeheli, P.1990. Interferon-induced proteinsand theantiviral state.Adv. VirusRes.38:147-200.
30. Staeheli, P., P. Dreiding,0. Haller, andJ.Lindenmann. 1985. Polyclonal and monoclonal antibodies tothe interferon-induc-ible proteinMxofinfluenza virus-resistant mice. J.Biol.Chem. 260:1821-1825.
31. Staeheli, P., and 0. Haller. 1985. Interferon-induced human protein with homologytoproteinMxofinfluenza virus-resistant mice.Mol. Cell. Biol. 5:2150-2153.
32. Staeheli, P., 0.Haller, W. Boll, J.Lindenmann, andC. Weiss-mann. 1986. Mx protein: constitutive expression in 3T3 cells transformed with clonedMxcDNAconfers selective resistance toinfluenza virus. Cell 44:147-158.
33. Staeheli, P., and J. Pavlovic. 1991. Inhibition of vesicular stomatitis virus messenger RNA synthesis by human MxA protein. J. Virol. 65:4498-4501.
34. Van derBliek,A. M., and E. M.Meyerowitz. 1991. Dynamin-likeprotein encoded by theDrosophila shibiregeneassociated with vesicular traffic.Nature(London)351:411-414.
35. Yeh, E., R.Driscoll,M.Coltera,A. Olins,and K. Bloom.1991. Adynamin-like protein encoded by theyeastsporulationgene SPO15.Nature(London) 349:713-714.
36. Young, J. F., U. Desselberger, P. Graves, P. Palese, A. Schatz-man, and M. Rosenberg. 1983. Cloning and expression of influenza virusgenes, p. 129-138. In W. G. Laver (ed.), The origin of pandemic influenza viruses. Elsevier Science Publish-ing Co., Inc., New York.
37. Ziurcher, T., J. Pavlovic,and P.Staeheli. Mechanism of human MxAprotein action: variants with changed antiviral properties. EMBOJ., inpress.