Vol.64, No. 7 JOURNAL OF VIROLOGY,
JUlY
1990, P.3545-35500022-538X/90/073545-06$02.00/0
CopyrightX) 1990,American Society for Microbiology
Sequence and
Functional Differences
between Schmidt-Ruppin
D
and
Schmidt-Ruppin
A
Strains of
pp6Ov-src
SITA
REDDY,t DIANNE MAZZU,t
DENNIS MAHAN, ANDDAVID SHALLOWAY*Department of Molecular and Cell Biology, The Pennsylvania State University, University Park, Pennsylvania 16802
Received5January 1990/Accepted18April1990
We show that
Schmidt-Ruppin
D pp6ov-src kinase activity is reduced byamutation previouslyshowntobeassociated with
Schmidt-Ruppin
A pp6Ov-srctemperaturesensitivity and that its reduced transformingactivityis associated with
aconformational
change in the SH3 region. The evolutionary relationship ofseven v-srcstrains
wasstudied
byusing parsimony analysis.At
least
six
different v-src strains are commonly used,
often
with
the assumption
that
their biological effects are
equivalent. With
the
exception of two recently isolated
strains
(6), these
strains probably derive from the original
Rous tumor no. 1 isolate (19, 34). All v-src proteins
con-tain
the same
replacement for the carboxyl terminus of
pp60-csrc
but, because of rapid
retroviral mutation (3),
have diverged at multiple isolated residues during their
evolution
through
laboratory
passaging (2, 4, 15, 18, 28,
31-33).
Reduced transforming activities of SRD v-src relative to
SRA-SF v-src. We
discovered
that
chimeric
v-srclc-src
plas-mids
expressing Schmidt-Ruppin D (SRD) amino region
coding
sequences
combined
with c-src
carboxyl region
se-quences
had significantly lower focus-forming activity, when
tested by transfection into NIH 3T3 cells, than similar
constructs
containing Schmidt-Ruppin
A-San Francisco
(SRA-SF)
sequences
(Table
1
[26]). This
effect
wasevident
with
plasmids
in
which either Rous sarcoma
virus
(RSV)
long
terminal
repeats
(LTRs)
for low-level
expression
orMoloney murine leukemia virus (MoMLV) LTRs
for
high-level
expression
were
used.
Nonchimeric SRD
v-srchad less
activity
than
SRA-SF v-src when tested with RSV LTRs but
not
when tested with
MoMLV LTRs
(Table
1).
(This
is
consistent with
ourearlier demonstration
that
transfection
assays
with weak
promoters
often
provide
moresensitive
tests
for functional differences
than assays with strong
promoters
[26].) We
conclude
that
structural differences
between
SRD and SRA-SF
v-srcmodulate
their
transform-ing
activities.
SRD
v-srcsequence.
To
identify
the
mutations
responsible
for
these
functional
differences,
wesequenced
the SRD
v-srcgene
(Fig. 1).
We
found
that the SRD
and SRA-SF
(2,
15)
v-src
sequences
differ
in
nine
nucleotides, which
result in
five amino acids changes,
atpositions 62, 124, 318,
368,
and
461.
pp6ov(SRD)-src
kinase
activity
is reduced
primarily
by
its
Val-461-*Met
mutation.
The presence of Met-461 in SRD
v-src
rather than
Val-461
(present
in
SRA-SF
v-src)
wasof
*
Corresponding
author.t
Presentaddress: Centerfor CancerResearch, MIT,
Cambridge,
MA02139.tPresent address:
University
of Nevada School ofMedicine,
Reno,NV89510.
particular interest since this mutation is required, but not
sufficient,
for the
induction of
temperature
sensitivity in
three mutants
of SRA-New
York
(SRA-NY) v-src (8, 21). To
study the effect of this mutation alone, we
constructed
plasmid
pMvsrcAD
expressing
a
SRA-SF mutant
having
aVal-461-*Met
substitution
[pp6ov(sRA-SF)-src(M46l)]
and
plas-mid pMvsrcDA
expressing
aMet-461-*Val SRD v-src
mu-tant[pp6Ov(sRD)-src(V461)I
(Fig. 2). The relative
specific
activ-ities of
the
v-srcvariants
weremeasured with
the exogenous
substrate
(acid-denatured
enolase)
immune
complex
kinase
assay
(Table 1). Similar results
wereobtained in
experiments
in
which either monoclonal
antibody
EC10 (22) or 327 (16),
which binds different epitopes,
wasused.
pp6OV(sRD)-src
had
about
threefold
lower
specific activity
than
pp60vtSRASF)src;
analysis of
pp60v(SRA-SF)-src(M461)
and
pp60v-src(SRD)-src(V461)
showed
that this
reduction
wasassociated with Met-461
alone. Garber
etal.
(5) have
previously reported
that
Val-461-*Met mutation
does
notdecrease the kinase
activity
of SRA-NY
pp60v-src.
The
difference
from
ourresults
might
be due
tothe
useof different types of kinase assays
ordue
tothe
additional amino acid differences (at residues 124
and
301) between the SRA-NY and SRA-SF
pp6Ov-src
strains.
We suspect that the
equal focus-forming
activities
of
pMvsrcAD
and
pMvsrcDA
reflect lack of
sensitivity of
the
focus-forming
assay
with MoMLV
LTRplasmids (26)
and
that differences between the
pp6ov(SRA-SF)-src(M46l)
and
pp6ov(SRD)-src(V461)
focus-forming activities
would be
ob-served if RSV LTR
expression plasmids
wereused
(cf.
the
focus-forming
activities of
the MoMLV and RSV LTR
v-srcplasmids pMvsrcD, pMvsrc, psrcll,
and
pRvsrcA).
Altered
pp6Ov(SRD)-src
V-8 protease
cleavage
pattern.
Partial
digestion of
either 32P-labeled
immunoprecipitated pp6Oc-src
pp6Ov(sRA-sF)-c
.orpp60v(5RA
SF)-src(M461)
withStaphylo-coccus aureus
V8
protease
yielded
34-kilodalton
(kDa)
ami-no-terminal
Vi
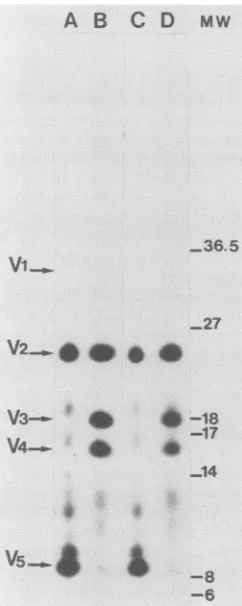
and
26-kDa
carboxyl-terminal
V2
fragments
and
18-and 16-kDa V3 and
V4amino-terminal
subfragments
(Fig.
3).
However,
analtered pattern
wasobserved
for
pp60_v(SRD)-src
andpp6Ov(sRD)-src(v46l):
this
contained
re-duced
amountsof the V3 and V4
fragments
and
anovel
9.5-kDa V5
fragment
that
wasonly
weakly
present
in the
pp6Ov(sRA-sF)-src
digests.
The
V3,
V4,
and
V5
fragments
of
both
pp6Ov(SRD)-src
andpp60v(SRA-SF)-src
contained only
phosphoserine,
while all the
V2fragments
contained
only
phosphotyrosine
(data
notshown).
The
pp6Ov(SRD)-src
and
3545
on November 10, 2019 by guest
http://jvi.asm.org/
3546 NOTES J. VIROL.
D ACTCTGCTGGTGGCCTCGCGTACCACTGTGGCCAAGCGGTAGCTGGAACGTGCAGCCGACCACC
A G G
D 1 MET GlySer SerLys Ser Lys Pro LysAsp Pro Ser GlnArg Arg Arg Ser Leu GLuPro ProAsp Ser Thr His His Gly Gly Phe Pro D 1 ATG GGG AGT AGC MG AGC MG CCT MG GAC CCC AGC CAG CGC CGG CGC AGC CTG GAG CCA CCC GAC AGC ACC CAC CAC GGG GGA TTC CCA
D 31 Ala Ser GLn ThrProAsnLysThr AlaAla ProAspThr HisArg Thr Pro SerArgSer PheGlyThrVal Ala Thr GluPro Lys Leu D 91 GCC TCGCAGACC CCC AAC MG ACA GCA GCC CCC GAC ACG CAC CGC ACC CCC AGC CGC TCC TTC GGG ACC GTG GCC ACC GAG CCCAAG CTC
c GlyGLy
D 61 Phe Glu AspPheAsnThr SerAspThrVal Thr Ser ProGlnArg Ala GLyAla LeuALa GLy GlyVat Thr Thr PheVal Ala LeuTyr D 181 TTC GAG GAC TTC AAC ACT TCT GAC ACC GTT ACG TCG CCG CAG CGT GCC GGG GCA CTG GCT GGC GGC GTC ACC ACT TTC GTG GCT CTC TAC
A G
A Gly
c Arg Thr Asp
D 91 Asp Tyr Glu Ser Trp lieGtu ThrAspLeuSer Phe Lys Lys Gly Glu ArgLeuGln Ile Val Asn Asn ThrGlu Gly Asn Trp Trp Leu D 271 GAC TAC GAG TCCTGG ATT GAA ACG GAC TTG TCC TTCMG AMGGA GAACGC CTG CAG ATT GTC AACAAC ACG GM GGT AAC TGG TGG CTG
A G
c Leu
D 121 Ala HisSer ValThr Thr Gly Gln Thr Gly Tyr Ile Pro Ser AsnTyr Val Ala Pro SerAspSer Ile GinAla Glu GluTrp Tyr Phe
b 361 GCT CAT TCC GTG ACT ACA GGACAG ACG GGC TAC ATC CCC AGT AAC TAT GTC GCG CCC TCA GAC TCC ATC CAG GCT GAA GAG TGG TAC TTT
A C C
A Leu
D 151 Gly Lys Ile ThrArg Arg Glu SerGluArg Leu Leu Leu Asn Pro GluAsn ProArg Gly Thr Phe LeuValArg Glu Ser GluThr Thr D 451 GGGMGATC ACT CGT CGG GAG TCC GAG CGG CTG CTG CTCMC CCC GAA MC CCC CGG GGA ACC TTC TTG GTC CGG GAG AGC GAG ACG ACA D 181 LysGly Ala Tyr Cys Leu Ser VaL Ser AspPhe Asp AsnAla Lys GlyLeu AsnVal Lys His Tyr Lys Ile Arg Lys LeuAspSerGly
D 541 MA GGT GCC TAT TGC CTC TCC GTT TCT GAC TTT GACMC GCCMG GGGCTC MT GTG MG CAC TAC AAG ATC CGC AAG CTG GAC AGC GGC D 211 Gly PheTyr Ile ThrSer ArgThr GlnPhe Ser Ser LeuGln Gln Leu Val Ala Tyr Tyr Ser LysHisAla AspGly LeuCys HisArg
D 631 GGC TTC TAC ATC ACC TCA CGC ACA CAG TTC AGC AGC CTGCAG CAG CTG GTG GCC TAC TAC TCC AAA CAT GCT GAT GGC TTG TGC CAC CGC D 241 Leu ThrAsnVal Cys ProThrSer Lys Pro GlnThr GlnGly LeuAla Lys Asp Ala Trp Glu Ile Pro Arg Glu Ser Leu Arg Leu Glu D 721 CTG ACCMC GTC TGC CCC ACG TCCMG CCC CAG ACC CAG GGA CTC GCCMG GAC GCG TGG GMATC CCC CGG GAG TCG CTG CGG CTG GAG D 271 Vat Lys LeuGly GlnGlyCysPhe GlyGlu Val Trp Met Gly Thr Trp AsnGlyThr ThrArgVal Ala Ile Lys Thr LeuLys Pro Gly D 811 GTG MGCTG GGG CAG GGC TGC TTT GGA GAG GTC TGG ATG GGG ACC TGGMC GGC ACC ACC AGA GTG GCC ATA AAG ACT CTG MG CCC GGC
c Asn Arg
D 301 Thr Met Ser ProGluAla Phe LeuGln GluAla Gln Val Met Lys Lys Leu GlnHis Glu Lys Leu Val Gln Leu TyrAla Val Val Ser D 901 ACC ATG TCC CCG GAG GCC TTC CTG CAGGMGCC CAA GTG ATGMG MGCTC CAG CAT GAGMG CTG GTT CAA CTG TAC GCA GTC GTG TCG
A G G
A Arg
c Thr
D 331 GluGlu Pro lieTyr lieVal lieGlu Tyr Met Ser Lys GlySer Leu Leu AspPhe Leu Lys Gly GluMet GlyLys Tyr Leu Arg Leu D 991 GMGAG CCC ATC TAC ATC GTC ATT GAG TAC ATG AGCMG GGG AGC CTC CTG GAT TTC CTGMG GGA GAG ATG GGCMG TAC CTG CGG CTG
c Ala
D 361 Pro GlnLeuVal Asp MetAlaAspGin lieAla SerGlyMetAla TyrVal Glu Arg Met Asn Tyr Vat HisArg Asp Leu ArgAla Ala
D 1081 CCACAG CTC GTT GAT ATG GCT GAT CAG ATT GCA TCC GGC ATG GCC TAT GTG GAG AGG ATGMC TAC GTG CAC CGA GAC CTG CGG GCG GCC
A C
A Ala
D 391 Asn IleLeuVal GlyGlu Asn Leu Val Cys Lys Val Ala AspPheGly LeuAlaArg Leu lieGlu Asp Asn Glu Tyr Thr Ala Arg Gln
D 1171 MCATC CTG GTG GGGGAG MC CTGGTG TGCMG GTG GCT GAC TTT GGG CTG GCA CGC CTC ATC GAG GACMC GAG TAC ACA GCA CGG CM
D 421 Gly Ala LysPhe Pro IleLys TrpThrAla ProGluAla Ala Leu Tyr GlyArg Phe Thr Ile Lys Ser AspVat Trp Ser Phe Gly Ile
D 1261 GGT GCC AAG TTC CCC ATCMG TGG ACAGCC CCC GAG GCA GCC CTC TAT GGC CGG TTC ACC ATC MG TCG GAT GTC TGG TCC TTC GGC ATC
c Val Val Gln
D 451 Leu LeuThr Glu LeuThr Thr Lys GlyArg Met Pro Tyr ProGlyMet GlyAsnGlyGlu Val Leu Asp ArgVal GluArg Gly Tyr Arg D 1351 CTG CTG ACT GAG CTG ACC ACC MG GGCCGG ATG CCA TAC CCA GGG ATG GGC MC GGG GAG GTG CTGGAC CGG GTG GAG AGG GGC TAC CGC
A G
A Val
c Lys
D 481 Met Pro Cys Pro Pro Glu Cys Pro Glu Ser Leu His Asp Leu Met CysGln Cys Trp Arg Arg Asp ProGlu GluArg Pro Thr PheGlu
D 1441 ATG CCC TGC CCG CCC GAG TGC CCC GAG TCG CTG CAT GAC CTT ATG TGC CAG TGC TGGCGGAGG GAC CCT GAG GAG CGG CCC ACT TTT GAG
A C
c Phe Glu Asp Tyr Phe Thr Ser Thr Glu Pro GlnTyrGlnProGlyGluAsnLeu
D 511 Tyr Leu GlnAla Gln Leu Leu ProAla CysVal Leu Glu Val Ala Glu
D 1531 TAC CTG CAG GCC CAG CTG CTC CCT GCT TGT GTG TTG GAG GTC GCT GAGTAGTGCGCGAGCAAAATTTAAGCTACAACMGGCAAGGCTTGGCCGACAATTGCA
A T A
FIG. 1. SRDv-srcsequence. Thenucleotide sequence reported here will appear in the GenBank and EMBL data bases under accession number M33292. The coding sequence begins at position 1. Nucleotides and amino acids that are different in SRA-SF v-src are shown in lines marked A. Amino acids that are different in
pp60-src
are shown in lines marked c. The sequence was determined by Maxam-Gilbert sequencing (17) of fragments fromasubcloneof SRDv-srcphageXSR-RSV-D-5 (29). Allregions were sequenced at least twice and, in most regions, onbothDNAstrands.on November 10, 2019 by guest
http://jvi.asm.org/
[image:2.612.96.533.94.645.2]NOTES 3547
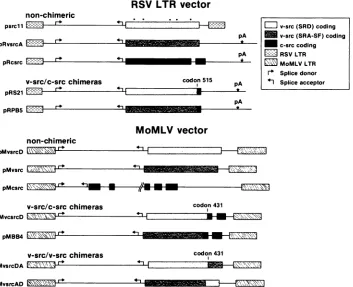
non-chimeric
pIrr1
psrcl11 A= L pRvsrcA
pRcsrc
v-src/c-src
_
chimeras-*
pRS21 Lt--=
RSV LTR vector
pA
pA
codon 515
pA
pA
pRPB5
non-chimeric
pMvsrcD r
pMvsrc
MoMLV
vector
I _H__
_\_NN~, 4,
pMcsrc \'r* I"
v-src/c-src chimeras
pMvcsrcD *
pMBB4
v-src/v-src chimeras
pMvsrcDA E _
codon 431
_'
codon 431
.,.,________________________
pMvsrcAD F-EX 7\'
FIG. 2. src expression plasmids. Only eucaryotic regions are shown. LTRs and coding regions are boxed. Coding sequences are from chickenc-src, SRD v-src, orSRA-SF v-src and are indicated by hatching. Dots above the psrcll coding sequence indicate the positions of amino acid differences between SRD and SRA-SF v-src. pA marks the positions of simian virus 40 early region polyadenylation sites. Recombinations were made at eitherBglI (codon 431) orPstI (codon 515) sites. psrcll (35), pMvsrc, and pMcsrc (9), pRvsrcA, pRcsrc, pRS21, pRPB5, and pMBB4 (26) have been described. Fragments from these plasmids were cross-ligated to create the other plasmids. The pMvsrc, pRvsrcA, pRPB5, and pMBB4 coding sequences were derived from plasmids donated by H. Hanafusa.
pp6oV(SRA-SF)-src
hadidentical tryptic
phosphopeptide
maps(data
notshown).
The
release
of
asmall
amountof V5 fragment from
pp60v(SRA-SF)-src
andcomparison of the SRD and SRA-SF
sequences
show
thatthe SRD
V5 cleavage site does
notresult from the
introduction of
anadditional
glutamic acid
residue.
Thus,
it islikely that
accelerated V5 cleavage
results
from
amutation-induced conformational change
in the
pp60v(SRD)-src
amino region. This
conformational
change
couldexplain
thereduced
transforming activity of
pMvcsrcD,
the v/c-src chimeracontaining
SRD-specific
amino acids at
positions 62, 124, 318, and 368, relative
topMBB4,
the related SRA-SFchimera
(Fig.
2 and Table1).
This reduced
transforming activity is
notdue
toreduced
specific
kinaseactivity
(Table 1)
butmight
reflect alteredinteraction(s)
with cellularproteins resulting
from amuta-tion-inducedconformational
change.
TheV5 cleavage
siteis located near theboundary
of the srchomology
3(SH3)
region, indicating
that the conformation ofat least part of thisregion
is altered.Furthermore,
oneof thetwomutationsflankingthe novelsite, Leu-124(SRA-SF)-* Val(SRD),lies
withintheSH3
region.
It haspreviously
beensuggested
that the SH3region
governs interactions ofpp6Osrc
and cellularproteins (8, 10, 23, 24).
The functional difference between SRD and SRA-SF v-src mayprovide
anexample
of acorrelated effect on both
pp6Osrc
structure and function mediatedby
thisregion.
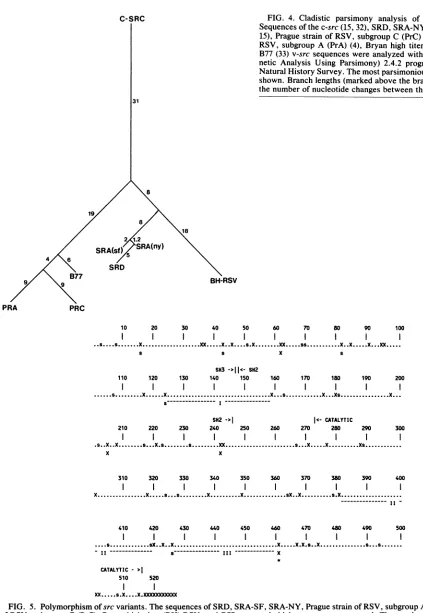
Parsimony
andpolymorphism
analysis.Figure
4shows
anevolutionary
map ofseven v-srccoding
sequencesderived
fromcladistic
parsimony analysis
(which
generates themapcontaining
the minimal numberofnucleotide
changes [20]).
The
consistency
index is96%
(i.e., the analysis
iscompli-cated
by
<4% of back orparallel mutations).
The map is consistent with the limitedinformation
that is availabledescribing
theevolutionary
historiesof the srcvariants(12,
27,
34)
andprovides
aguide
to compareexperiments
with different v-srcstrains.Thelocations of the
polymorphic
sitesareshown inFig.
5.Except
fortheAsn-301---
>ThrandThr-338->Ile substitutions(the
latter alone can activate the transformingpotential
ofpp6Oc-src [11])
and thecarboxyl-terminal substitutions,
noneof the
coding
mutations relative to c-src are found in all of the v-src variants. This is consistent with theassumption
thatmostof the
point
mutationsaroseindependently
atlater stages of viral evolution.Although
the smallsample
sizeprecludes
calculation ofstatistically
significant
measures, the survival oflarge
re-gions
without mutation suggests thatthey
are necessaryfor
activity.
The threelargest
nonpolymorphic
amino acidsequences
(33
to 37 residues inlength)
include amino acids 125 to 158(region I),
382 to 419(region
II), and 427 to460
(region III) (Fig. 5). Region
Ioverlaps
the SH3 andSH2 domains. While
deleting
amino acids 112 to 142 does notdestroy
Prague-A
v-srctransforming activity (25),
= v-src (SRD) coding
M
v-src(SRA-SF) codingmc-src coding
=
RSV LTR=
MoMLV LTRFe Splice donor
*I Splice acceptor
-3
rlp
q'i-3
r- -1 m0-1
=
... m
VOL.64, 1990
I.- -.6
r-al ^
(( .__
on November 10, 2019 by guest
http://jvi.asm.org/
[image:3.612.134.487.79.366.2]TABLE 1. src
activity
Specifickinaseactivityc'
Plasmid Expressed gene" formingRelative
focus-activity Monoclonal EC10 Avg
antibody327 El v
RSV LTR src plasmids Nonchimeric
psrcll v(SRD)-src 0.008 x 1.3
pRvsrcA v(SRA-SF)-src 0.03 x_ 1.1
pRcsrc c-src <10-6
v-srclc-srcchimeras
pRS21 v(SRD)/c-src (515) <10-6
pRPB5 v(SRA-SF)/c-src(515) 0.002 x 1.6
MoMLV LTR srcplasmids Nonchimeric
pMvsrcD v(SRD)-src 1.1 x 1.4 0.2 0.4 0.3 x 1.4
pMvsrc v(SRA-SF)-src 1.0x 1.3 1 1 1 x 1.2
pMcsrc c-src 0.02 x. 2.1 0.1 0.1 0.1 x 1.4
v-srclc-srcchimeras
pMvcsrcD v(SRD)/c-src(431) 0.2 x 1.6 0.9 1.3 1.0x 1.2
pMBB4 v(SRA)/c-src(431) 1.1 x 1.4 1.4 1.3 1.3 x 1.2
v-srclv-srcchimeras
pMvsrcDA v(SRD)-src(V461) 0.9 x 1.4 1.1 1.2 1.1 x 1.1
pMvsrcAD v(SRA-SF)-src(M461) 0.9_x 1.5 0.4 0.2 0.3 x. 1.4
a V461,
Met-461-aVal
mutation. M461,Val-461-'Met
mutation.Numbers inparentheses indicatethe codonpositionsatwhich thechimeric recombinationsweremade.
bNIH3T3 cells weretransfected (5x 105cellsper 60-mmplate) with0.002 ,ug(forsrcexpression plasmidscontainingtheMoMLVLTRs)or0.1ILg(forsrc
plasmids containing RSVLTRs) ofthe indicated plasmidsand20
jig
ofsonicated calfthymuscarrierDNA aspreviously described (9).Pilot experiments showedthattheseamountsinduced foci in thelinearresponse range of 0 to 100fociper plate. DNAs werelinearized bycleavageat auniqueNruIin theirprocaryotic regionspriortotransfection. Relativefocus-forming activities aregeometricaverages of a minimum of threeexperiments withduplicate plates. Valuesare normalized to theaveragefocus-formingactivityofpMvsrc (1.4x105fociper pmol). Thehigheroverallfocus-forming activities ofthe MoMLV LTRplasmids
resultfromtheirinductionofhigherlevels of srcexpression (26).
'Massculturesofsrcexpressercellswerecoselectedin 400,ug of G418(GIBCO Laboratories)per ml aftercotransfecting 5x 105 cells per 60-mmplate with
5 ,ugoftheindicated srcexpression plasmid andneoexpression plasmidpSV2neo(30) in a10:1molarratio. G418-resistantcolonies(50to 100)were pooledand
grownforeach massculture. Activitiesof theindicatedsrcproteinsweremeasuredin theimmune complexkinaseassayby usingeither monoclonal antibody
327 (2 to 3 experiments; average value shown) or EC10 (1 experiment) with acid-denatured rabbit muscle enolase as substrate. The proteins were
immunoprecipitated fromthe mass cultures after 18 hof metaboliclabelingwith[35Slmethionine and were thenincubatedwith[y-32PIATPand substrateas
previously described(13). Specific kinase activities werecalculated fromthe amounts of
[35S1pp60'r'
and[32P]enolase
in theexcised bands (determined by double-channel scintillation counting)and thespecific activitiesof metaboliclabeling(13). Thegeometric averages ofallexperimentswithbothantibodiesare shown.A
B
C
D
Mw.36.5
deletions that include the 3' part of
region I (e.g., 142
to169
[1],
143
to173
[25],
or112
to196
[25])
destroy
ordi-minish
v-srctransforming activity
in
accord with the
hypo-thesis that
atleast this
portion of region I has been
selec-tively
conserved.
Regions
II and III lie
within
the central
core
of the
catalytic domain,
which has
the greatest
fre-quency
of
highly
conserved
residues
(these
regions
include
subdomains VII and VIII described in
reference
7).
Numer-ous
studies have shown
that
mutation of
amino
acids
within these
regions
caneliminate v-src kinase and
trans-forming
activities
(reviewed
in reference
23).
It is
interesting
that the Val 461- Met
mutation is
atthe
border
of
region
III.
None of the other
v-srcpolymorphisms
occur atamino
acids
that
arehighly
conserved amongst the
protein kinases
(7).
*- * *_11
V4--.O.
[image:4.612.57.560.64.276.2]Vs-- .&
FIG. 3. Cleveland
digests
of SRDandSRA-SFpp60src
proteins.pp60src
proteins
wereimmunoprecipitated
withanexcessofmono-clonal
antibody
EC10from cells labeledwith[32P]orthophosphate
for 16h,partially digested
withStaphylococcus
aureusV8 protease,and
analyzed by
12.5% sodiumdodecyl sulfate-polyacrylamide
gelelectrophoresis
andautoradiography
aspreviously
described (13).Lanes A
through
D containdigests
ofpp60v(sRD)-src
(A),pp60v(SRA-SF)-src
(B),
pp6Ov(SRD)-src(V461)
(C),
andpp60v(SRA-SF)-src(M461)
(D)
derivedfromcoselectedmassculturesof NIH 3T3cells thatweretransfected withsrcexpression plasmids
(Fig.
2)
andpSV2neo (30).
Thepositions
of the 34-kDa aminofragment (V1),
26-kDacarboxyl
fragment (V2),
18-kDa amino-terminalfragment
(V3),
16-kDaamino-terminalfragment (V4),
and novelamino-terminal9.5-kDaV5
fragment
areindicated. The posi-tionsofmolecularweight
standards areindicatedin kilodaltons in theMw
column.3548 -8
-6
on November 10, 2019 by guest
http://jvi.asm.org/
[image:4.612.120.241.415.718.2]NOTES 3549
C-SRC
FIG.
4. Cladisticparsimony
analysis
of seven v-src strains.Sequences of thec-src(15,32), SRD,SRA-NY (18, 31),SRA-SF (2,
15), Prague strain of RSV, subgroup C (PrC)(28), Prague strain of
RSV, subgroup A (PrA) (4), Bryan high titer (BH-RSV) (18), and
B77 (33)v-src sequences were analyzed with thePAUP (Phyloge-netic Analysis Using Parsimony) 2.4.2 program from the Illinois
Natural HistorySurvey. Themostparsimoniousevolutionarytreeis shown. Branch lengths(marked above thebranches)correspondto
thenumberofnucleotide changes between the different srcgenes. 31
8
81
18 2 1.2
SRA(s SRA(ny)
4
SRD
B77 BH-RSV
PRA PRC
10 20 30 40 50 60 70 80 90 100
II
...X...XX... ....X...S.X..X...X...S...X...X X...X
S S X S
SH3 ->||<- SH2
110 120 130 140 150 160 170 180 190 200
...s...X...X... X Xs...X...
-- I
---SH2
-I
||c- CATALYTIC210 220 230 240 250 260 270 280 290 300
.X. s. ...s...s.. X...X... . ....Xs...
X
310 320 330 340 350 360 370 380 390 400
X...X.s...s..X.X ..sX..X s.X.
410 420 430 440 450 460 470 480 490 500
. sX..X..X.... X.X.s. ....s..Xs
- I--- s--- III--- X
CATALYTIC
>1
510 520
l
l
XX s.X X.XXXXODX)O(X
FIG. 5. Polymorphism ofsrcvariants.Thesequences of SRD, SRA-SF, SRA-NY, PraguestrainofRSV, subgroupA(PrA), Praguestrain
of RSV, subgroup C (PrC), Bryan high titer(BH)-RSV, and B77 v-src andchicken c-src are compared. The numbers identify amino acid
positions. Dots represent amino acids thatareconserved in all variants. Silent mutations are indicatedbys;amino acid substitutions are
indicated by X. Multiple symbols represent multiple mutations within the same codon in different variants. With the exception ofthe
mutationsat301,338,and the carboxylterminus,noneofthesubstitutionsrelativetoc-srcareconservedinallv-srcstrains.Thewigglylines
and Roman numerals identify the threelargestnonpolymorphicregions.The boundariesof theconservedcatalytic, SH2(14),andSH3 regions
aremarked. The positionof Met-461 is marked withanasterisk.
VOL. 64, 1990
on November 10, 2019 by guest
http://jvi.asm.org/
[image:5.612.56.479.61.674.2]3550 NOTES
We areindebtedto T. Whittamfor cladisticparsimony analysis andhelpfuldiscussions; K.Lynch, S. Haggerty,and R. F.Frisque for assistance in sequencing; H. Hanafusa, G.M. Cooper, and A. D.Zelenetzfor plasmids;and J. S.Bruggeand S. J. Parsonsfor antibodies.
This workwas supported by Public HealthService grants RO1-CA32317 andRO1-CA47333 fromthe National InstitutesofHealth andanRCDA (K04-CA01139)toD.S.
LITERATURE CITED
1. Cross, F. R., E. A.Garber, and H. Hanafusa. 1985. N-terminal deletion inRoussarcomavirus
p60src:
effectsontyrosine
kinase andbiological activities andonrecombinationintissue culture withthecellularsrc gene. Mol. Cell.Biol. 5:2789-2795. 2. Czernilofsky, A. P., A. D. Levinson, H. E. Varmus, J. M.Bishop, E. Tischer, and H. Goodman. 1983. Correctionstothe nucleotide sequence of the src gene ofRous sarcoma virus. Nature(London) 301:736-738.
3. Dougherty, J. P., and H. M. Temin. 1986.High mutationrateof aspleennecrosis virus-based retrovirusvector.Mol. Cell. Biol. 6:4387-4395.
4. Fincham, V. J., and J. A. Wyke. 1986.Locationof temperature-sensitive transformation mutations and back mutations in the Roussarcoma virus src gene. J.Virol.58:694-699.
5. Garber, E. A., B.J. Mayer, R. Jove, and H. Hanafusa. 1987. Analysis ofp60vsrc mutantscarrying lesions involved in tem-peraturesensitivity. J. Virol. 61:354-360.
6. Hagino-Yamagishi, K., S. Ikawa, S. Kawai, H. Hihara, T. Yamamoto, and K. Toyoshima. 1984. Characterization oftwo strains ofavian sarcoma virus isolated from avian lymphatic leukosis virus-induced sarcomas.Virology 137:266-275. 7. Hanks, S. K., A. M. Quinn, and T. Hunter. 1988. The protein
kinase family:conservedfeatures and deduced phylogeny ofthe catalytic domains. Science241:42-52.
8. Hirai, H., and H. E. Varmus. 1990. Site-derivedmutagenesis of the SH2- and SH3-coding domains of c-src produces varied phenotypes, including oncogenic activation of
p60csrc.
Mol. Cell. Biol. 10:1307-1318.9. Johnson, P. J., P. M. Coussens, A. V. Danko, and D. Shalloway. 1985. Overexpression ofpp6Oc-src can induce focus formation without complete transformation of NIH 3T3cells. Mol. Cell. Biol.5:1073-1083.
10. Jove, R., and H. Hanafusa. 1987. Cell transformation by the viralsrconcogene. Annu. Rev. Cell Biol. 3:31-56.
11. Kato,J.-Y., T. Takeya, C. Grandori, H.Iba,J.B. Levy, and H. Hanafusa. 1986. Amino acid substitutions sufficient toconvert thenontransforming
p60csrc
protein to a transforming protein. Mol. Cell.Biol.6:4155-4160.12. Klement, V., and J. Svoboda. 1963. Induction of tumors in Syrian hamstersby two variants of Rous sarcoma virus. Folia Biol. (Prague) 9:181-186.
13. Kmiecik, T. E., and D.Shalloway.1987.Activationand suppres-sion ofpp60c-Src transformingability by mutation of its primary sites of tyrosine phosphorylation. Cell49:65-73.
14. Koch, C. A., M. Moran,I. Sadowski, and T. Pawson. 1989. The commonsrchomology region2domain of cytoplasmic signaling proteins isapositive effectorof v-fps tyrosine kinase function. Mol. Cell. Biol. 9:4131-4140.
15. Levy,J. B., H. Iba, and H. Hanafusa. 1986. Activation of the transformingpotential of
p60csrc
by asingle amino-acid change. Proc.Natl. Acad. Sci. USA83:4228-4232.16. Lipsich, L. A., A. J. Lewis, and J. S. Brugge. 1983. Isolation of monoclonalantibodies that recognize the transforming proteins
of aviansarcomaviruses. J. Virol. 48:352-360.
17. Maxam,A.,andW.Gilbert. 1980.Sequencingend labeledDNA withbase
specific
chemical cleavages. MethodsEnzymol. 65: 499-560.18. Mayer, B.J.,R.Jove,J.F.Krane,F. Poirier,G.Calothy,and H. Hanafusa. 1986. Genetic lesions involved in
temperature
sensitivityof thesrcgeneproductsoffour Roussarcomavirus
mutants.J.Virol. 60:858-867.
19. Morgan, H. R. 1964. Origin of Rous sarcoma strains. Natl. Cancer Inst.Monogr. 17:392-393.
20. Nei,M. 1987. Molecularevolutionary genetics. Columbia
Uni-versity
Press, NewYork.21. Nishizawa, M., B. J. Mayer, T. Takeya, K. Yamamoto, K. Toyoshima,H.Hanafusa,and S.Kawai. 1985. Two
independent
mutations arerequired for temperature-sensitive cell transfor-mationbyaRous sarcomavirustemperature-sensitive mutant. J.Virol. 56:743-749.
22. Parsons,S.J.,D.J. McCarley,C. M.Ely,D. C.
Benjamin,
and J.T. Parsons. 1984. Monoclonal antibodies to Rous sarcomaviruspp6Osrcreactwithenzymaticallyactive cellularpp6Osc of avianand mammalianorigin.J. Virol.51:272-282.
23. Parsons, J. T., and M. J. Weber.1989. Genetics of src: structure andfunctional organizationof aprotein tyrosinekinase. Curr. Top.Microbiol. Immunol. 147:79-120.
24. Pawson, T. 1988.Non-catalytic domains of cytoplasmic protein-tyrosine kinases: regulatory elements in signal transduction. Oncogene3:491-495.
25. Raymond, V.W.,andJ.T.Parsons. 1987. Identification ofan
aminoterminal domainrequiredfor thetransforming activityof the Rous sarcoma virus src protein.Virology160:400-410. 26. Reddy, S.,P. Yaciuk, T. E.Kmiecik, P. M. Coussens, and D.
Shalloway. 1988. v-src mutations outside the carboxyl-coding region are not sufficient to fully activate transformation by pp60c-src in NIH 3T3 cells. Mol. Cell. Biol. 8:704-712. 27. Schmidt-Ruppin, K. H. 1964. Heterotransplantation of Rous
sarcomaand Rous sarcoma virusto mammals. Oncologia 17: 247-272.
28. Schwartz, D. E., R. Tizard, and W. Gilbert. 1983. Nucleotide sequenceofRous sarcomavirus. Cell 32:853-869.
29.
Shalloway,
D., A. D. Zelenetz, and G. M. Cooper. 1981. Molec-ularcloning and characterization of the chickengene homolo-gous to the transforming gene of Rous sarcoma virus. Cell 24:531-541.30. Southern,P.J., and P.Berg. 1982. Transformation of mamma-lian cells to antibiotic resistance with abacterial gene under control of theSV40 early regionpromoter. J.Mol. Appl.Genet. 1:327-341.
31. Takeya, T., and H. Hanafusa. 1982. DNA sequence of the viral and cellular src geneofchicken.II.Comparisonof the src genes of two strains of avian sarcoma virus and of the cellular homolog. J. Virol. 44:12-18.
32. Takeya, T., and H. Hanafusa. 1983. Structure and sequenceof the cellular gene homologous to the RSV src gene and the mechanism for generating the transforming virus. Cell 32: 881-890.
33. Verderame, M.F., J. M. Kaplan, and H. E. Varmus. 1989. A mutation in v-srcthat removes asingleconserved residue in the SH-2 domain of pp6O-src restricts transformation in a host-dependent manner. J. Virol. 63:338-348.
34. Wang, L.-H., and H. Hanafusa. 1988. Avian sarcoma viruses. VirusRes.9:159-203.
35. Yaciuk, P., and D. Shalloway. 1986. Features of the pp6Ov-src carboxyl terminus that are required for transformation. Mol. Cell. Biol. 6:2807-2819.
J. VIROL.