JOURNALOFVIROLOGY,JUlY1973, p.108-113
Copyright O 1973 American Society for Microbiology
Vol.12,No.1
Printed in U.S.A.
Lack of
a
Close
Relationship Between Three
Strains of Human
Rhinoviruses
as
Determined
by Their RNA Sequences'
FAY H. YIN, K. LONBERG-HOLM, AND S. P. CHAN
CentralResearchDepartment,Experimental Station, E. I. Du Pont de Nemours and Company, Wilmington, Delaware
19898,
andBionetics ResearchLaboratories,
Bethesda,
Maryland
20014Received forpublication9March 1973
The possible genomic homologies between three serotypesofhuman
rhinovi-ruses (HRV 1A, HRV 2, and HRV 14)wereinvestigated. Firstweconfirmed that
theseviruseswereunrelatedby the criterion of the absence ofcommonantigenic
determinantsonthe surfaces of thenative virions,asdetected by
cross-neutrali-zation ofcomplementfixation. RNA-RNA hybridizationwasthen examined with
purified, highly radioactive, double-stranded, replicative-form RNA andexcess
single-stranded virion RNA. Single-stranded RNA showed 100% homology with the minusstrand from thereplicative-form RNA of thesametypeof virus. HRV 1A, HRV 2, and HRV 14 showed low intertypic homologies; these were not
significantly greaterthanthose found between therhinoviruses and
poliovirus,
whichwereused as anegative control. The immunological relationship and the
RNA homologybetweenHRV1A andHRV 1Bwerealsoexamined by the above
techniques. It was confirmed that HRV 1A and HRV 1B share some surface
determinants and itwasalso foundthat HRV1B RNA shares 70%homology with
HRV 1A RNA.
Rhinoviruses have been
categorized
as asub-group of
the
picornaviruses because
oftheir
acid
lability and their relatively
high buoyant
den-sity
inCsCl (for
reviews see9, 19,20). Most
ofthe
everincreasing numbers of
recognized
human
rhinovirus(HRV)
types(5, 6)
are,by
definition, unrelated
tothe others
by
neutrali-zation
with
antisera. In atleast
one case,however, there
is a commoncomplement fixing
antigen
(7). There
is,
however, little
informa-tion uponwhich
tobuild
anyconclusion about
the
degree
ofgenetic
relatedness
amongthe
human rhinoviruses.
This
investigation
is a partof
anongoing
efforttomake adetailed molecular
comparison
of a few
selected
strains ofhuman
rhinoviruses (10, 12, 14, 15). Native virions of HRV1A,
HRV 2, and HRV 14 werepermitted
to react withhigh
titer antiserawhich wereproduced against
the native virions ofthese types, and the results confirm the absence of
intertypic
native anti-genic determinantsby both
the criteria ofcrossneutralization
andby
complement
fixation. We have then examined thedegree
ofRNA: RNAIThis paperis contribution no. 2014 ofthe Central
Re-search Department ofE. I. DuPont de Nemoursand Com-pany.
homology between these selected
viruses. Inboth the immunochemical and hybridization
studies, HRV 1B served
as a positive controlsince
it isalready known
tobe related
toHRV
1A.
Polio
2served
as a negativecontrol.
Aspecific
serological relatedness between
dena-tured
virions ofHRV
1Aand
HRV 2 hasbeen
discovered and
we willdiscuss this
in thefollowing article (13).
MATERIALS AND METHODS
Virus, viruspreparations,andpurification.The
sources and propagation of HRV 1A (strain 2060), HRV2(strainHGP), HRV14(strain 1059), andpolio type 2 (strain P712-Ch2ab) have already been de-scribed (10, 12). HRV 1B(strainB-632) wasobtained
from the Reference Reagent Service oftheNational InstituteofAllergy and Infectious Diseases (NIAID) (Bethesda, Md.) andwas
plaque-purified
and propa-gated by the same procedures. The identityofeachstrain ofvirus wasconfirmed by neutralization tests with standard reference serum obtained from the NIAID.
Allhighly purifiedvirionpreparationswerelabeled with3H-uridine,
3H-amino
acids,"4C-amino
acids,or32P-phosphatetofacilitate the processofpurification (10, 12). Thenumberof virionparticles per milliliter
in thedialyzed pools was calculated bythe method
108
on November 10, 2019 by guest
http://jvi.asm.org/
LACK OF
already described (10).
Antiserum. Antisera against highly purified vi-rions were prepared in young female rabbits. After CsCl-gradient centrifugation, virions were diluted with nine volumes of cold water and samples of 1 ml were either used directly or frozen at -70 C. In the firstinjections, 1 ml of antigen wasmixed with 0.5 ml of CompleteFreunds Adjuvant (Difco) and portions of the mixture were injected intoall four foot pads. At 2-to 4-week intervals, additional samples of antigen were thawedand given in the ear vein. Typically, 0.2 to 0.4 absorbance units (260 nm) ofvirions were used perinjection.The rabbits were exsanguinated 1 week afterthe 4th or 5th injection.
Neutralization. An appropriate amount of virus wasdiluted 100-fold into antiserum in medium con-taining 5% heat inactivated fetal calf serum. This mixture was kept for 1 h at room temperature and then diluted 10- and 100-fold. These dilutions were used forplaque assay accordingtothe methodalready described (10). The neutralization titers of the anti-sera are the dilutions producinga90% reduction in plaques.
Complement fixation tests. The method was es-sentially that of Chan et al. (4). This employed sheep erythrocytes, rabbit anti-sheep erythrocyte hemoly-sin, and guinea pig complement (Microbiological Associates, Bethesda, Md.). Complement was ti-trated daily in serial twofold dilution steps. The highest dilution which caused100%lysis of sensitized blood (an equal mixture of 0.5%sheep-red-blood cells andhemolysin) wasused in eachtest.
Complement fixation titrations were made in
block-checkerboard patterns. Purified viruses and antisera were diluted serially with Veronal buffered saline in Kahn tubes. Samples of 0.025 ml each of virus, antiserum, andcomplement were deposited in microtiter plates (Microbiological Associates, Be-thesda, Md.), agitated, sealed, and kept at 4 C overnight. The next morning, 0.05 ml of sensitized bloodwasadded to each well and this was incubated
at37C for1h. Theplates were centrifuged at 200 x g for 5 min inanI.E.C. centrifuge with the aid of special adapters (Microbiological Associates, Bethesda, Md.). Thepatternsobtained resembled thosealready described byothers for the complement fixation by crude poliovirus (11, 17). The +4 (0-10%hemolysis) endpointwaschosen ineachcase (17).
VirionRNA. Nonradioactive virionswerepartially purified through the sucrose-gradient centrifugation step, but omitting CsCl-gradient centrifugation, as employed in the purificationofradioactive virus (see above). In the cases of HRV 1A or HRV 1B, the
sucrosegradients contained1MNaCl and 0.02 M pH
8.1Tris-chloridebuffer. The virions were visualized in
the sucrose gradients by light scattering and the appropriate fractions were dilutedfivefold in 0.02 M pH 7.5 Tris-chloride buffer and sedimented in the ultracentrifugeat175,000 x gfor1.5hat 4 C. At this stage, thepelleted virions were impure but the major fraction of theRNA is probably within virions.
The pelleted virus from 20 or more roller bottles was frozen in 1.5 ml 0.2% sodium dodecyl sulfate (SDS) 0.02M pH 7.5 Tris-chloride, 0.002 M EDTA,
pH 7.5. The pellet wasthawed and dispersed with the aid of ultrasonic treatment (15 s withthe Biosonic III ultrasonic generator, Bronwill Scientific, Rochester, N.Y.). RNA was extracted for 90 s with phenol at
45 C; the phenol phase was reextracted with the buffer used above and the combined aqueous phases were reextracted with phenol. The RNA was pre-cipitated from the final aqueous phase by addition of 0.04volumes of2MNaCl and three volumes of etha-nol(overnight, -20). The RNAwaspelleted ina low-speedcentrifuge andwasredissolvedin 1.0ml0.2M NaCl containing 0.01 MpH7.5Tris-chloride buffer. It was then reprecipitated with ethanolat -20 C. The pelletwaswashed witha cold7:3mixture of ethanol and buffer saline and then dissolved in 2.25 x SSC (SSC, 0.15MNaCl, 0.015 M sodium citrate, pH 7.0). It was assumed that the RNA hadan Eiv257 of250.
Typical preparations contained 200 ug of RNA perml, and the ratio ofadsorptionat257 nm tothatat230nm
was2.3.
Preparation of labeled double-stranded RNA. Monolayer cultures of HeLa cellswereinfectedat34.5
C with rhinovirusesat amultiplicityof50
PFU/cell
inthe presence of 5
,g
ofactinomycin D per ml. After4h, 50
MCi
of 3H-uridine per ml was added and the incubation was continued for12h.The cellswerethen pelleted from the medium by low-speed centrifuga-tionandwerestored frozenat -70C. The frozen cells werethawed andsuspended in buffer containing 1%SDS and0.01Msodium acetate,pH5,at adensityof 2 x 107 cells per ml. The suspension was extracted three timeswithequal volumes ofphenolat60C for 6 minand the RNA in the aqueousphasewas precipi-tated twice with ethanol. Single-stranded RNAwas
differentiallyprecipitatedwith1MNaClaccordingto
the method ofBishop and Koch (1). Thesupernatant fluid containing soluble RNA and double-stranded RNA (ds-RNA) was precipitated with alcohol and redissolved inbuffer containing 0.1 MNaCl,0.002M sodiumphosphate,0.001MEDTA, and 0.5% butanol, pH 7.2. RNA from approximately 2 x 101 cells in 2 ml ofbufferwasappliedtoandseparated on a column of sepharose2B (2.7by35cm)at roomtemperaturewith theabove bufferat aflowrateof 15ml per haccording
to the method of
Oberg
and Philipson (16). The effluentwascollectedandmonitored forradioactivity
andoptical absorbanceat260 nm.Hybridization of RNA. Hybridization tests were
performed according toa modification of the method ofYoungetal. (23). Ten
Mliters
ofds-RNAcontaining less than 10 ng and over 1,000 counts/minute was mixed with 90Mlitersofdimethyl sulfoxide (DMSO) and5Mliters
of yeastt-RNA (1 mg/ml).Themixture was denatured at 67 C for 20 min and was then quicklycooled inanicebath. Forannealing, 1.2 ml of2.25 x SSC and 10 Mliters of unlabeled single-strandedvirion RNAwere then added. The mixture was heated again to 67 C for 1 h and then slowly cooledto roomtemperature. RNaseAand RNase
Ti
(Worthington Biochemical Corp., Freehold, N.J.)wereadded to the finalconcentrations of 40
Ag/ml
and60units/mland the mixturewasincubatedat 37C for
30 min. It was assumed that trichloroacetic acid-insolublecountsrepresented hybridized RNA.
109
12,1973
on November 10, 2019 by guest
http://jvi.asm.org/
YIN, LONBERG-HOLM, AND CHAN
RESULTS
Lack of
commonantigenic determinants
onthe
native
virions
of three
typesof
rhinoviruses. The anti-virion
seraproduced
asdescribed
inthe Materials and Methods section
have higher potency
than the usual typing
seraand
wereable
toneutralize
90% ofhomolytic
virus at a
dilution of
up to 33,000(HRV 1A),
43,000
(HRV 2),
or14,500(HRV 14). These
serashowed
noheterotypic neutralization within
this
group atconcentrations
1,000-fold
greaterthan those
atthe
homologous end point. HRV
1B, however, was
90%
neutralized
by
a480-fold
dilution of
anti-HRV 1A
serum.Further
attempts weremade
todetect
hetero-typic
antigenic
determinants
onthe surface
of native virionsby complement
fixation.The
+4 end point (17) waschosen
foreach virus-serum
pair, and
for purposes ofrepresentation, the
dilution
ofthe antiserum
atthis
point
wasmultiplied
by the dilution of the virus. This
product
wasthen
multiplied by
1010and
then
normalized
by dividing
itby
the
number of
particles
permilliliter
inthe
undiluted
virussample. The
titersfor
all virus-antisera pairs
have
been represented
inthis
mannerand
are given inTable
1which gives the results of both
heterotypic and homotypic
tests.Except
forthe
anti-HRV 1A and HRV 1B
pair, the native
virions
fix
complement
only
with
homotypic
sera.
Characterization
of
labeled
double-stranded RNA.
Atypical
profile
ofthe
separa-tion
of3H-labeled cell-associated rhinovirus
RNA
on asepharose 2B column is shown in
Fig.
1.
The
peak
at48ml,
which is the void
volume,
contained ds-RNA. It
usually
contained less
than
1.0,ug
ofRNA
perml with
105to5 x105
counts per min per ,g.
This material
eluted
inthe
void volume
again when
rechromatographed
on
sepharose 2B and
italso
migrated during
gel-electrophoresis
as asingle peak
which
coin-cided with the
replicative
form
ofrhinovirus
RNA,
aspreviously
reported by
Yin
and
Knight
(22).
The
double-stranded
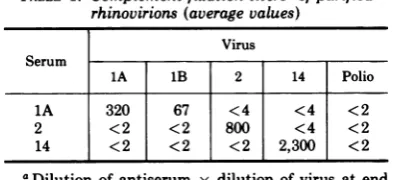
RNA was 95 to 100%TABLE 1. Complementfixationtitersaofpurified rhinovirions(averagevalues)
Virus Serum
1A 1B 2 14 Polio
1A 320 67 <4 <4 <2 2 <2 <2 800 <4 <2
14 <2 <2 <2 2,300 <2
aDilution ofantiserum x dilution of virus atend
pointx1010dividedbyvirusparticlesperml.
C.PM.
O.D.
40 80 120 160 200 240 280 ml
FIG. 1. Isolation of double-stranded RNA. HRV 2 RNA was labeled with 3H-uridine, extracted from infected cells, and fractionated with1MNaCI.RNA from 2 x 108 cells was applied to a sepharose 2B column. The radioactivity of the effluent was deter-mined as counts per minute per 0.1 ml, and optical ab-sorbance was determined at 260 nm.
resistant
todigestion by
pancreatic RNase Aand RNase
Ti
under
the conditions described
inMaterials and
Methods, but
wasabout 10%
resistant
after first
being denatured
in DMSOand then
being annealed
inthe
presence ofadded
yeastt-RNA,
asalso described. In the
absence
ofadded
yeastRNA,
alarger and
variable fraction became RNase
resistant.The
fact that the
radioactive RNA became
RNase sensitive afterdenaturation-annealing indicates
that
itsconcentration
during annealing is
toolow for
most ofthe
separated strands
tofind
each other. The
reason for a 10%residual
resistance is not
entirely clear
but this residual
portion
was shown tobe RNA since
it wascompletely
digested by
RNase
ifdiluted
in water instead of 2.25 x SSC. DNase also did notdigest the
resistantfraction and
it wascom-pletely
digested
by
0.5M
NaOH. Without
annealing (i.e.,
withrapid
cooling), the
RNase-resistant counts were
found
tobe
7 to8%. Thereplicative
form ofpoliovirus
RNA hasalso been
found
tocontainsuch
anRNase-resistant frac-tion after denaturationand
annealing under
similar conditions
(personal
communication,N. A.Young).
Hybridization of
homologous RNA.
La-beled
HRV 2ds-RNA
washybridized
with
increasing amounts of
single-stranded
virion110
J. VIROL.on November 10, 2019 by guest
http://jvi.asm.org/
[image:3.495.270.456.61.291.2] [image:3.495.64.262.558.648.2]RNA. The percentage of hybridization was
calculated by (i) subtractingthe
RNase-resist-antcounts ofthedenatured ds-RNA alone as a
background from all other samples, and (ii)
assumingthatthe RNase-resistantcountsofthe
undenatured ds-RNA were equal to 100%
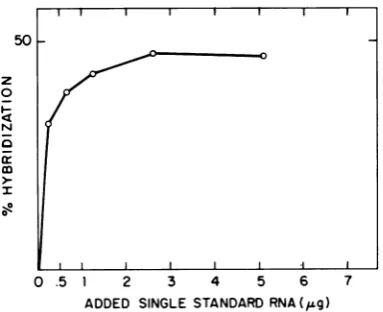
an-nealing. The hybridization increased with in-creasing quantities ofadded virion RNA (Fig.
2). When more than 2.5 usg of single-stranded RNA was added, the percentage of hybridiza-tion approached the theoretical value of 50% (i.e., all labeled minus strands were
hybri-dized). InthecaseofHRV lAand HRV14, the dependency of hybridization upon added ho-mologousvirionRNAwaspreciselythesameas
shown for HRV 2 inFig. 2.Themean deviation
forreplicate sampleswasalso foundtobe -4-2%. Lack of hybridization of heterologous RNA. Attempts were madeto anneal
heterolo-gous virionRNA withds-RNA. In these
experi-ments,4 Mgof virion RNAwasusuallyusedper
sample, buttwotothree times thisquantitywas
also employed in a few cases. Homologous
single-strandedRNAwasalsousedasapositive
control andsingle-strandedpoliovirusRNAas a
negativecontrol. The resultsareshown inTable
2. Rhinovirus iB, which is known to be
im-munologically related to HRV 1A, was also
included in orderto test the sensitivity ofthe
technique.
The results of Table 2 show that
single-stranded RNAs are able to hybridize with
ap-proximately 50% of the homologous ds-RNA. Except in the case of HRV lA and HRV iB, heterologous HRV single-stranded RNAs show
no more than about 2% hybridization ofHRV
50
z
0
N
a
I'
0 .5 1 2 3 4 5 6 7
ADDED SINGLE STANDARDRNA(,~g)
FIG. 2. Dependence ofthe extentofhybridization
upon the concentration ofunlabeled single-stranded
RNA. Double-stranded RNA (1,120 counts per min persample) was hybridizedwithincreasingamounts
ofvirion RNA. Thepercentage ofhybridization was
[image:4.495.241.437.87.300.2]calculatedas described in Results.
TABLE 2.
Hybridization of
HRVRNAsDouble-stranded Single-stranded Hybridization
labeled RNA unlabeled RNA (%
lA lA (3)a 52.3
lB (1) 36.3
2 (2) 1.3
14 (2) 0.9
polio
(3) 0.8lB lB (1) 51.0
lA (4) 34.8
polio
(3) 1.02 2 (2) 46.0
14 (2) 0.0
lA
(2) 1.5polio
(3) 0.114 14 (2) 49.0
lA
(1) 0.02 (2) 2.0
_____________ polio (3) 0.0
aParentheses indicatenumber of
experiments
per-formed to obtain average value of percentage ofhybridization.
1A,
HRV2,
orHRV 14ds-RNA;
this isprobably
not
significantly
greater
than thehybridization
produced
by poliovirus
RNA. In the case of HRVlA
and HRViB,
eithersingle-stranded
RNA can
hybridize
about 35% of thecomple-mentary
ds-RNA.DISCUSSION
The three
serotypes
of human rhinoviruses chosen forthisstudy
represent
two members ofone
cell-receptor family (HRV
2 and HRV1A)
and one member of
another
family (HRV 14)
(12).
They
alsodiffer,
each from theother,
inbuoyant density
in CsCl and in the size of theirconstituent
polypeptides
but havesimilar sedi-mentation coefficients(10)
(Korant
andLon-berg-Holm, unpublished
data).
Although
these virusesdo differ
fromeach other
by
other than
serological
criteria,
it is notknown
whether
they
may
be
considered
asrepresentative
examples
for
the
large
HRV
family.
It has
already
been
established that the
strains
of
HRV
designated
astypes
1A, 2,
and
14 differ in theirneutralizing
determinants.Typ-ing
sera aregenerally
ofrelatively
lowpotency
and itwasof interesttoreexamine the
question
of
cross-neutralization
with therelatively highly
potent
serum which wasproduced against
highly purified
virions. Thisserum wasable
toneutralize
90%ofhomotypic
virusinfectivity
ata
dilution
ofgreater
than 104.Our
resultsconfirmed
that the rhinovirustypes
investi--I
VOL.
12,
1973on November 10, 2019 by guest
http://jvi.asm.org/
[image:4.495.40.232.454.610.2]YIN,LONBERG-HOLM,AND CHAN
gated did
notshare neutralizing determinants.
However, anti-HRV
1A serumshowed about
1 to 2% of itshomotypic reactivity
againstHRV
1B
and
thisconfirmed that HRV
1Aand HRV
1B
maybe
considered
assubtypes. The possible
existence of
non-neutralizing determinants
wasalso examined
by complement
fixation testsand
the data
of Table 1 showthatthere
was nosignificant
reactionbetween heterotypic
pairs of virionsand
sera.There
was,however,
a signifi-cantreactionbetween HRV 1A
serumand HRV
1B virions, as
expected
onthe basis
of theneutralization data. This
reaction isalso
rela-tively
moreprominent
(67/320)
than
the
corre-sponding
crossneutralization (480/33,000).
It
washoped that RNA:RNA hybridization
would detect
asignificant
homology
between
atleast
twoof
the three
serotypesexamined.
Other
groups of viruses
have shown such
relationships.
For
example, the RNAs
ofthe three
serotypes ofpoliovirus share
about
25 to 33% sequencehomology although the
virionsdo
notshare
appreciable neutralizing determinants (23).
Three strains of foot-and-mouth disease virus
also share RNA
homology (8)
aswell
ascertainneutralizing
antigens (2;G. E. Cottral,
1972.Bull. Off.
Int.Epiz.,
inpress).
Six
strains ofinfluenza A share
70% RNA homology and
influenza
Aand B share
20% homology (18).
In the
caseof
hybridization between
homolo-gous.
HRV
RNAs, for
example
HRV
2ds-RNA
and HRV
2virion
RNA
asshown
inFig.
2, half
the
label
ofthe ds-RNA
could be annealed
tononradioactive
virusRNA and this
waspresum-ably the minus strands. This
simply
confirms
that
the
virionscontained
only plus
strands,
and hence
50%
hybridization
indicates
100%homology. The sensitivity
ofour testfor RNA
homology
wasverified
by
the
hybridization
found with RNAs
of HRV1A and HRV
1B.
There
isabout
35%homology using
either
pair
of
single-stranded
and ds-RNAs and this
indi-cates
that about
70%of the
genomes arecom-posed of homologous
sequences.The
results
ofhybridization
tests madewith
various
pairs
ofds- and
single-stranded
RNAs are shown inTable
2. It canbe
seen thatheterologous pairs
donotproduce
morethan 2%hybridization,
afigure
which isprobably
notsignificantly
greater than the0 to 1%observed
with
poliovirus plus
strand RNA. Two percenthybridization would
indicate that 4% of the genome ishomologous,
under
the conditionsused
inthe
test. This mustthen be considered anupperlimit
forthespecific
intertypic
hybrid-ization of HRV 2
and
1A. Since
the rhinovirus genome has amolecular
weight
ofabout
2.5 x 106(3,
14),
it cancode
forabout
2.5 x105
molecular weight
units of protein.Four
percent ofthis
represents 104molecular weight
units orroughly
onepolypeptide
atthe
most, if all homology resides in a single portion of the genome.It is
also possible that hybridization is
not sensitiveenough
todetect genetic homology
between
certainrelated polypeptides because
of misfitsproduced
by degeneracy
inthe
geneticcode
(21)and
because
ofgeneticdrift. In this
regard, it has
recently been
found that the messengerRNA of human
hemoglobin
cannothybridize effectively with DNA complementary
to
rabbit
hemoglobin
message,despite the fact
that
human and rabbit
hemoglobin have
verysimilar
structures(S.
Packman, H. Aviv, J.
Ross, D. Swan, and P. Leder; private
communi-cation).
The
possibility still
existsthat
there
could be
common
polypeptide
sequenceswhich
cannotbe detected
atthe surface
ofHRV
virionsby
cross-neutralization
orby complement fixation
tests.
Some
commonantigenic
determinants
have indeed been discovered between denatured
HRV
1A and HRV 2virions, and
this isthe
subject
ofthe
following
paper(13).
ACKNOWLEDGMENTS
We thank R. Z. Lockart for continued support and encouragement.Lynn Magee and Virginia Kiloren provided experttechnical assistance.
LITERATURE CITED
1. Bishop, J. M., and G. Koch. 1966. Purification and characterization of polio-induced infectious double-stranded ribonucleic acid. J. Biol. Chem. 242:1736-1743.
2. Brown, F., B. Cartwright, and J. F. E. Newman. 1964. Further studies of the early antibody in the sera of cattle and guinea pigs infected with foot-and-mouth disease virus. J. Immunol. 93:397-402.
3. Brown, F., J. F. E. Newman, and E. J. Stott. 1970. Molecular weight of rhinovirus ribonucleic acid. J. Gen. Virol. 8:145-148.
4. Chan, S.P., R. D. Maca, P. H. Levine, and R. C. Ting. 1971.Immunological studies of human breast cancer. I. Serum reactivity againstalymphoid cell line (Belev) derived from a breast cancer patient as detected by
complement fixation test. J. Nat. Cancer Inst. 47:511-517.
5. Collaborative Report. 1967. Rhinoviruses, a numbering system.Nature (London) 213:761-763.
6. Collaborative Report. 1971. A collaborative report: rhino-viruses-extension ofthenumbering system. Virology 43:524-526.
7. Dans, P. E., B. R. Forsyth, and R. M. Chanock. 1966. Density of infectious virus and complement-fixing antigen of two rhinovirus strains. J. Bacteriol.
91:1605-1611.
8. Dietzschold, B., 0. R. Kaaden, T. Tokui, and H. 0. Bohm. 1971. Polynucleotide sequence homologies among the RNAsoffoot-and-mouth disease virus types A, Cand0.J.Gen.Virol. 13:1-7.
9. Hamre, D. 1968. Rhinoviruses, p. 1-88. In J. L. Melnick (ed.), Monographs in Virology, vol. 1. S. Karger, New
York.
112 J. VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
10. Korant, B. D., K. Lonberg-Holm, J. Noble, and J. T. Stasny.1972. Naturally occurringandartificially
pro-duced components of three rhinoviruses. Virology 48:71-86.
11. LeBouvier, G. L. 1955. The modification ofpoliovirus antigens by heat and ultraviolet light. Lancet
2:1013-1016.
12. Lonberg-Holm, K., and B. D. Korant. 1972. Early in-teraction of rhinoviruses with host cell. J. Virol.
9:29-40.
13. Lonberg-Holm, K., and F. H. Yin. 1973. Antigenic determinants of infective and inactivated virustype2. J. Virol.12:114-123.
14. Nair, C. N., and K. K. Lonberg-Holm. 1971.Infectivity and sedimentation of rhinovirus ribonucleic acid. J.
Virol.7:278-280.
15. Noble, J., and K. Lonberg-Holm. 1973. Interactions of
components of rhinoviruses with host cells. Virology 51:270-278.
16. Oberg, B., and L. Philipson. 1969. Replication of
polio-virusRNA studied bygelfiltration and
electrophore-sis. Eur. J.Biochem. 11:305-315.
17. Schmidt, N. J., and E. H. Lennette. 1956. Modification of the homotypic specificity ofpoliomyelitis comple-ment fixing antigens by heat. J. Exp. Med.
104:99-120.
18. Scholtissek,C., and R. Rott. 1969. Hybridization studies withinfluenzavirusRNA. Virology 39:400-407. 19. Stott, E. J., and R. A. Killington. 1972.Rhinoviruses.
Annu. Rev.Microbiol.26:503-524.
20. Tyrrell, D.A.J. 1968. Rhinoviruses,p.67-124.In S. Gard, C. Haullauer, and K. T.Meyer (ed.), Virology
Mono-graphs, vol.2.Springer-Verlag, New York.
21. Watson,J. D.1970.Molecular biologyofthegene.W. A.
Benjamin, Inc., New York.
22. Yin, F.H.,andE.Knight,Jr. 1972.In vivo and in vitro
synthesisofhuman rhinovirustype2ribonucleicacid.
J. Virol. 10:93-98.
23. Young, N. A., B. H. Hoyer, and M. A. Martin. 1968. Polynucleotide sequence homologies among
polio-viruses.Proc. Nat.Acad.Sci.U.S.A.61:548-555.