Bacteriophage
Interference
in
Bacillus
subtilis 168
R. E. YASBIN, A. T. GANESAN,' ANDF. E. YOUNG
Departments ofMicrobiology, Pathology, and Radiation BiologyandBiophysics, University ofRochester
Schoolof Medicine and Dentistry, Rochester,New York14642
Received forpublication 14November 1973
StrainsofBacillus subtilislysogenic fortemperatebacteriophage SP02 inhibit
thedevelopment of bacteriophage01.After infection by bacteriophage 41,DNA
and RNA synthesis inthelysogenic hostterminates, culminatingin cell death. Bacteriophage SP02 also prevents the production of bacteriophage 0105.
Mechanisms for these twotypes ofbacteriophage interference are discussed.
Successful infection by bacteriophage
re-quires attachment, injection of its nucleic acid intoabacterium, transcription, translation and replication of bacteriophage genetic
informa-tion, as well as release of infectious progeny.
Many factors may cause abortionofthe infec-tion such as alteration of growth media,
pro-phageinterference, and preinfectionofthe bac-teria with identicalordifferent bacteriophages, or both (1, 2, 8). In Bacillus subtilis, sporula-tionprofoundly influencesviralreplication. For example, the genetic information of bacterio-phage
oe
cannot be transcribed in sporulating cells(19,20) andbacteriophages02and015caninfect but not lyse cultures of B. subtilis 168, whereas theyarecapable of lysing nonsporulat-ingmutants (14, 15).Abortive infectioncanalso be the resultoflysogenic conversionorprophage interference(1,2).Prophage interferencecanbe accomplished byarestriction andmodification
system (6) with alteration ofthe cell envelope (16, 21)orby the repressionand/orinhibitionof
bacterial enzymes (18, 25). InEscherichia coli, bacteriophage lambda kills bacteria lysogenic
forbacteriophageP2without producingprogeny
phage (18, 25, 26), whereas lambda lysogens exclude the successfuldevelopmentof bacterio-phages T2, T4, and T6 that carry the rII
mutation (2, 12). The ability ofa prophage to
subsequently interfere withthedevelopmentof
other bacteriophages is often dependent on
interactionsbetween the bacterial hostand the
residentprophage.Specifically, temperate bac-teriophage co will prevent bacteriophages T2
and T4 from successfully infecting lysogenic
strains ofE. coliWand E. coliWs, butnotE. coli K-12 (21). In addition, the inability of
bacteriophage SP10to growundernormal
con-'Present address: Department of Genetics, Stanford Uni-versitySchool of Medicine, Stanford, California94305.
ditions in certain strains ofB.subtilis also has been attributed to prophage interference (10, 11).
0105 (isolated by Reilly) andSP02 (isolated byOkubo) are twotemperatebacteriophagesof
B. subtilis which have approximately thesame molecular weight and thesamemorphology (3,
5) andsimilarly decrease the levelsof
transfor-mation in lysogenic cultures (28-30). However, thesetwobacteriophages (i) areheteroimmune (4), (ii) havedifferent receptorsites, (iii)
inte-grate at differentsites on the bacillus
chromo-some (13, 23, 27), (iv) show about 10%
homol-ogy in DNA-DNA hybridization studies (7), and (v) differ in their requirements for host DNA polymerases (24). Our studies of inter-ference by bacteriophages 4105 and SP02 on infection with nine bacillus bacteriophages have revealed that bacteriophage SP02 in-hibits the virulent cycle of bacteriophage 41. Additionally, this report will show that bac-teriophage 0105 is depressed in its
develop-ment by SP02 when both bacteriophages are
inalyticcycleofinfection.
MATERIALS AND METHODS Strains and methods of propagation. Strain BR151 (carrying Iys-3, trpC2, metB10) is an auxo-trophic derivative of B. subtilis 168, originally ob-tained from B.Reilly. BR151(0105), BR151 (SPO2), and BR151 (4105, SPO2) are lysogenic strains of
BR151 carrying the prophages of 0105, SPO2, and both 4105 and SPO2, respectively. B. subtilis HSR
wasobtained from B. Reilly.
Thebacteriophages used inthis study are listed in Table1along with their method of propagation. All of thebacteriophages werepropagated on strainBR151 except for X29 whichwas grown on B. subtilis HSR. Three methods of propagation were employed. (A) The temperatebacteriophageswereinducedfromthe lysogenic cultures by the methods previously de-scribed (29). (B) Most of the phage lysates were 916
on November 10, 2019 by guest
http://jvi.asm.org/
TABLE 1. Bacteriophages used and methodof propagation
Bacteriophage Method of Indicator Mediautilized" Source
propagationa strain
0105 ... A BR151 M agar, M broth J. Hoch
40105cl-z B BR151 Magar, M broth S. Zahler
SP02 A BR151 M agar, M broth W.Romig
SPO2cl-2 B BR151 Magar, M broth S. Zahler
4e B BR151 Magar,Penassaybrothc C.Yehle
)1... B BR151 Magar,Penassaybroth B.Reilly
425 B BR151 Magar, Penassay broth B.Reilly
4)29 B HSR TBAB, Penassay broth B.Reilly
SP82 ... B BR151 Magar, Penassay broth M. Green
SPOl ... B BR151 Magar,Penassaybroth K.Bott
SPP1 ... B, C BR151 Magar,Mbroth T.Trautner
aProceduresdiscussed inMaterials and Methods.
bMedia aredescribed in Materials and Methods.
cPenassaybrothsupplementedwith 0.5%glucoseand0.1%yeastextractatthe time of inoculation. prepared by the addition ofphage to exponentially
growing cultures of bacteria (optical density of 50 Klettunitson aKlett-Summersoncolorimeter, filter
no. 66) at an MOI of 1 at37 C. (C) Bacteriophage
SPP1 was collected from agar plates of bacteria
having approximately 1,000 plaques by placing the topsoft agarinPenassay broth (2ml/plate), mixing
and then centrifugingat 1,200 rpminaSorvalltype
SPcentrifuge. Exceptforbacteriophage029,allofthe
phageswereassayed inasemisolidoverlayof modified
M agar on modified M agar plates as previously
described (29). Bacteriophage 429 was assayed in a
semisolid overlay of 1.0% Tryptone, 0.8% NaCl,
0.6%agar,and1.0%glucoseonTBABagarplates.
Measurement of RNA and DNA synthesis. A
20-MCiamountof[3H Juridine(New England Nuclear
Corp., Boston, Mass., 42.1 Ci/mmol) or 3 uCi of
[3HIthymidine(New EnglandNuclear Corp., Boston,
Mass.,6.7Ci/m mole)wasaddedto2mlof
exponen-tiallygrowing cells (opticaldensity of50Klettunits,
Klett-Summerson spectrophotometer,filterno.66) in
Penassay Broth(Difco antibioticmedium3).Samples of0.3 mlweretransferredatvarious timesinto 1.0 ml of cold 10% trichloroacetic acid. After 20 min of
incubation the samples were poured onto filters
(HAWP 025, HA, 0.45gmporesize; MilliporeCorp.
Bedford, Mass.) that hadbeenpreviously soaked in 0.2Msodiumpyrophosphate.Thesampletubeswere
washed three times with 3 ml of 0.015 M NaCl
containing1.5mMsodium citrate and0.014%
trichlo-roacetic acid, and the washes were pouredoverthe
filters. The filters were dried, placed in 10 ml of
Aquasol (Nuclear Chicago, Chicago, Ill.), and the
radioactivitywasdetermined inaTri-Carb
scintilla-tion counter.
RESULTS
Effect of lysogeny on bacteriophage
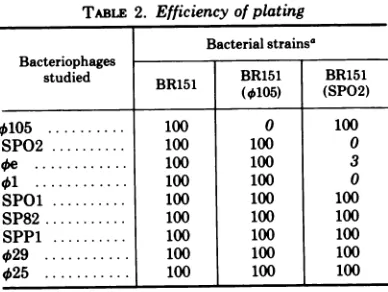
infection. Nine bacteriophages were used to infectB.subtilisBR151and its lysogenic deriv-atives. As expected, bacteriophages 0105 and SPO2 did notsuccessfully infect strains BR151
TABLE 2. Efficiency ofplating
Bacterialstrainsa Bacteriophages
studied BR151 BRI51 BR161
B51 (4405) (SP02)
4$105 .100 0 100
SPO2 ... 100 100 0
4e .100 100 3
41 .100 100 0
SPOl ... 100 100 100
SP82 ... 100 100 100
SPP1 ... 100 100 100
4)29
.100 100 100025 .100 100 100
aThepercentage of the bacteriophagesplating on strainsBR151 (SP02) orBR151 (4105) as compared
with BR151.
(0105)
and BR151 (SPO2),respectively
(Table
2).Thisinability toinfectthelysogenicstrains is presumably attributable to the presence of
repressor products
produced
by
the resident prophages. Strains lysogenic forbacteriophage
SPO2 were found toseverely
restrict bacterio-phage4e
and to totally prevent thedevelop-ment of bacteriophage
41.
The results with bacteriophage4e
are similar tothosereported
by Rettenmier andHemphill (22).
Thepro-phageSPO2 mediated interference of bacterio-phage
4)1
isnotrelatedtotheinability
of41
to attach to the cell wall of alysogen
of SPO2 (manuscriptinpreparation). Lysogenydoesnot appear to interfere with infectionby
SPOl,
SP82, SPP1,029,
or425.
Todetermineifbacterialdeath resultedfrom aninfectionofstrainBR151
(SPO2)
by
bacteri-ophage41,
thenumberofcolony
forming
units (CFU)was determinedprior
toand afterinfec-tion at an MOI of 20. The results
(Table
3)
917
on November 10, 2019 by guest
http://jvi.asm.org/
[image:2.494.246.440.253.399.2]clearlyshowthat bacteriophage 41 infection is lethal to bacteria lysogenic for SPO2. Control experiments involved infecting BR151 (SPO2) withSPO2c1-2(aclear plaquemutantofSPO2)
at an MOI of 20. In thiscase there was little if anycell death due to bacteriophage infection. These results clearly indicate that bacterio-phages 41 and SPO2arepreventedfrom devel-opinginstrains of B. subtilislysogenic for SPO2 by different mechanisms.
Co-infection ofB. subtilis with bacterio-phages 41 and SPO2. Exponentially growing culturesofstrainBR151wereinfected simulta-neously with bacteriophages 41 and SPO2. These infected cultures completely lysed
ap-proximately 60 min after infection. However, the number of bacteriophage produced was
severely decreased comparedtothesingle infec-tions(Table4). These resultswerealsoobtained whenbacteriophageSPO2c1-2wasused instead of SPO2. This reduction in the numbers of
[image:3.494.269.466.332.592.2]progeny bacteriophage could be the resultofa depressor effect type ofphenomenon (1). Mu-tual exclusion and depressoreffect phenomena have been previously identified in B. subtilis
TABLE 3. Infection ofBR151 (SP02) with bacteriophages 01andSPO2a
Infecting After infection bacteriophages CFU/ml Survival(%)
401
6.0x 105 0.9SPO2c1-2 6.1x107 71.8
aBacterial strain BR151
(SPO2)
at 8.5 x 107 colony [image:3.494.53.254.333.580.2]formingunits/ml (CFU/ml) was infected with either SPO2c1-2 or )1 (MOI = 20), and cultures were incubatedat37 C for 15 min.
TABLE 4. Infection ofBR151 withbacteriophages 4)1andSPO2a
Bacteriophages Bacteriophage progeny (PFU/ml)
added SPO2 01
01 ... NDb 1.1x 101i
SPO2 ... 5.2 x10' ND
SPO2 + 01 ... 1.0 x 108 1.2 x 107
SPO2c1-2 ... 4.7 x 109 ND SPO2c1-2 + )1 ... 1.0 x 108 1.8 x 107
aBacterialstrain BR151 (1.2 x 108 colony forming
unitsperml)wasinfected withbacteriophages 4)1and SPO2atanMOIof 20. Fifteenminutes after infection, the cultureswerecentrifuged at 1,200 rpm in a Sorvall type SP centrifuge, resuspended in M media, and incubated untillysis.
'ND,Nonedetermined.
with bacteriophages SPO2 (17). Although
si-multaneous infection ofstrain BR151 with bac-teriophages 41 and SPO2 result in decreased bacteriophage progeny, the interactionis
differ-ent than that observed when strain BR151 (SPO2) is infected by bacteriophage
01.
The former represents bacterial lysis with thepro-duction of bacteriophage, whereas the latter results in the death of the bacteria without production ofanybacteriophage orcell lysis.
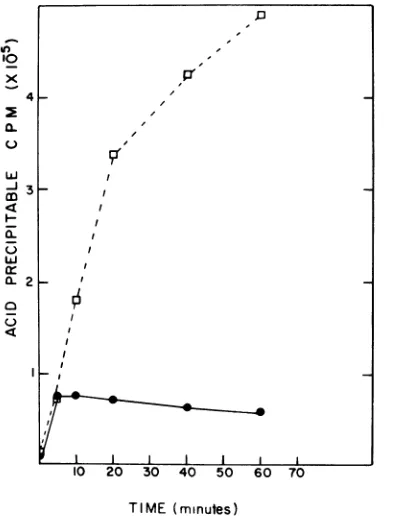
Macromolecular synthesis in 41 infected lysogens of SPO2. A determination of DNA and RNAsynthesiswasmadeonSPO2 lysogens infected with bacteriophage 01. Strain BR151 (SPO2) was infected with 4)1 (MOI of 20) simultaneously with the additionofradioactive uridineorthymidine. The resultsinFig. 1and2
demonstrate that shortly after infection, the synthesis of both RNA and DNA was termi-nated. The abrupt cessation of RNA and DNA synthesis was notobserved when nonlysogenic cultures were infected with 01. Additionally, preincubation of theSPO2 lysogenin 200 ugof
chloramphenicol per ml for 5 min priortothe addition of bacteriophage 01 preventsthe shut
In
x
4
a.
C.)
ui3
I-3
m3
a:
w
O-2 a
10 20 30 40 50 60 70
TIME (minutes)
FIG. 1. Incorporation of [3H]uridine into acid-precipitable counts in growing cultures of strains
BR151 (SPO2) (0) andBR151 (SPO2)infected with
41atanMOI of20(0)asdescribed inMaterials and Methods.
Pr
_ ,~~~~~~~~~~~
~r
I
I
I
I
on November 10, 2019 by guest
http://jvi.asm.org/
[image:3.494.63.255.347.413.2]"4
10
x
a-u3
LL J
m
I-a-2
u
a-0
< I
5 10 15 20 25 30 35 40 45
[image:4.494.43.243.58.322.2]TIME (minutes)
FIG. 2. Incorporation of [3H]thymidine into
acid-precipitable counts in growing cultures of strains
BR151 (SPO2) (0) and BR151 (SPO2) infected with 411atanMOIof20(0)asdescribed inMaterials and
Methods.
off of DNA synthesis (unpublished
observa-tion). This couldbeinterpretedas anindication that some protein is made either by bacterio-phage (1 orby the lysogen after infection with bacteriophage that adversely affects both
host andbacteriophage DNA synthesis. Effect ofbacteriophage SPO2 on develop-ment of bacteriophage 0105. Bacteriophage
0105 plates with equal efficiency on bacterial
strains BR151 and BR151 (SP02) (Table 2). However, we observed that after the induction (with mitomycin C) of bacterial strain BR151
(0105, SPO2) 100- to 1,000-fold more bacterio-phage SPO2were producedthanbacteriophage
0105 (Table 5). The two bacteriophages were distinguished byuse ofBR151, BR151 (0105),
and BR151 (SPO2) as indicator strains. Addi-tionally, plaques formed on strainBR151 after
induction of strain BRi51 (0105, SPO2) were picked andtested for theirabilitytolysestrains BR151 ((105) and BR151 (SPO2). Of the 208 plaques tested, 206 contained only bacterio-phage SPO2,whereas 2wereonlybacteriophage (105. Furthermore, simultaneous infection of BR151 by bacteriophages SPO2c1-2 and (105cl-z (aclearplaque mutantof 4105) again resulted in 100 times the number of
phage SPO2 released, compared to
bacterio-phage (105 (Table6). These results are similar to those obtained by L. B. Boice (personal communication).
A shorter eclipse period for bacteriophage SPO2 than for 0105 might explain the results seen in a mixed infection. Bacterial strains
BR151 (SPO2), BR151
(0105),
and BR151 (SPO2, 0105) were induced with mitomycin C to establish one step growth curves for thesebacteriophages.Under the conditions utilized in ourlaboratory, bacteriophage SPO2 does have a
shorter latent period than does bacteriophage
(105 (Fig. 3). Bacteriophage SPO2 begins to be released from the induced lysogen approxi-mately 20 min before bacteriophage X105 is
released. The latent period is the same in the
single anddouble lysogen.
DISCUSSION
Prophage interference. The establishment of lysogeny can result in vast physiological
changesinbacteria(2, 9). Amongthesechanges are the production of restriction enzymes (6),
the alterationofcell envelope (16, 21), and the interference with the biosynthesis and activity of enzymes (18, 25,26). Becauseofthese physio-logical changes, lysogenic cultures often are
unableto support the propagation of
bacterio-TABLE 5. Induction of BR151 (X105,SP02)a
Bacteriophagereleased
Timeafter (PFU/ml)
induction (min)
SPO2 +105
15 1.1 x 10' 9.0 x10' 120b 4.3x 109 6.1 x106
aStrain BR151 (0105,
SPO2)
was induced with mitomycinCasdescribedinMaterials and Methods. [image:4.494.246.441.378.599.2]bComplete lysishad occurredby 120min.
TABLE 6. Infection ofBR151 with bacteriophages SPO2cl-2 and4105c1-z'
Bacteriophage progeny Bacteriophages (PFU/ml)
added
SPO2 0105
SPO2c1-2 2.5 x 10' NDb
4105c1-z NDb 8.3 x 10'
SPO2cl-2+ 4105cl-z 2.1 x10' 2.3 x 107
aBacterialstrain BR151 was infected with
bacteri-ophages SPO2cl-2 and/or0105c1-zat anMOI of 20 as
describedinTable 4. Thecultureswereincubated at 37C untillysis.
bND, None determined.
g1
(C
919
on November 10, 2019 by guest
http://jvi.asm.org/
[image:4.494.245.440.509.597.2]06.I 0
105~.-,,,~~~~~~00,,,,, 0
w~~~~~~~~~
a~~~~~~~~~~~~~~~5a
30 60 goo
MINTE AFTER INUTIO
0
C06
5 3
10 10
30 60 90 120
MINUTES AFTER INDUCTION
FIG. 3. Induction of lysogenic cultures with
mi-tomycin C as described in Materials and Methods.
After addition of mitomycin C, sampleswereremoved
from strain BR151 (0105, SPO2), centrifuged in a
Brinkmann 3,200centrifuge (8,000 xg) for1min, and
the number of PFU in the supematant fluid was
determined.
phages. In this report we have identified two typesofprophage interference that exist in B. subtilis. Bacteria lysogenic for bacteriophage
SPO2 severely restrict virulent bacteriophage
4e, whereas these same lysogenic bacteria
to-tallypreventthedevelopment of virulent bacte-riophage )1 (Table 2). The type ofinteraction
that exists between bacteriophage 41 and
pro-phage SPO2appearstoresemble theinability of
temperature bacteriophage lambda to success-fully infect strains of E. coli lysogenic for P2
(18). This prophage interference has been shown to be the result ofan interaction of the old+ gene product of the prophage P2 and the three gene products of the bacteriophage lambda (gamma+, delta+, and red+a orred+)
(26, 31, 32). When both of the bacteriophages are wildtype they interact to causecell death withoutlysis (26). This phenomenon appearsto be the result ofa requirement for the recB, C
enzymefor survival in P2 lysogens and the fact that lambda infection inactivates this enzyme (25, 26, 31, 32). Infection of a B. subtilis lysogenic for SPO2 with bacteriophage 41 simi-larly results in cell death without lysis (Table 3), withaconcomitanttermination of DNAand
RNA synthesis (Fig. 1 and 2). Presently, it cannot be determined if the termination of DNA and RNA synthesis in bacteriophage 41
infected strains of BR151 (SP02) is
synchro-nous or asynchronous. However, Fig. 1 and 2 suggestthe cessation of RNAsynthesis precedes cessation of DNA synthesis. Determination of
the mechanismofprophage interference which
inhibits thedevelopmentofbacteriophage41 in
bacterial cultures lysogenic for SP02 will
de-pend on the identification of those genes con-trolling this prophageinterference system.
Effectof bacteriophage SPO2on bacterio-phage 4105. As shown in Table 2, bacterial cultures lysogenic for either bacteriophages 0105 or SP02 can be successfully infected by either SP02 or 0105, respectively. However, bacteriophage SP02seemstoinhibit the release
of bacteriophage 0105 in simultaneous mixed
infectionsorinduced cultures (Tables 5and6). Thisdepressionofthe numberofbacteriophage 46105 producedin amixed infectioncould be the resultof a shorter latentperiod for SP02than
0105 (Fig. 3). On the other hand, this
depres-sioncould be theresultofbacteriophage SP02 interfering with the development of
bacterio-phage 4105. It has already been shown that bacteriophage 0105 is more dependent on the bacterial DNA polymerase than is bacterio-phage SPO2 (24). Conceivably, during the de-velopmentofbacteriophage SPO2,hostenzyme systemswhichareessentialforthedevelopment ofbacteriophage4)105could be inhibited. Inany case, an important problem has arisen by the finding that both bacteriophages 0105 and SPO2 cannot develop in the same bacterium. Rutberg and co-workers attempted to
demon-strate that genetic recombination or com-plementation did not occur between bacterio-phages 0105 and SPO2 (24). However, their experiments were carried out by infecting bac-teria with various mutants of bacteriophage 0105 and a particular strain ofbacteriophage SPO2, and then by scoring the number of
bacteriophage 4105 produced under restrictive conditions. Becausethey chose to look at only
the number of 0105 bacteriophages produced, their conclusion that genetic recombination or
complementation did not exist between these
two temperate bacteriophages should be
re-examined. Chow et al. have recently reported
14% homology between the DNA of
bacterio-phages
4105
and SPO2, and have concluded that this homology might represent the genescoding for antigen specificity (7). This conclu-sioniscurrentlybeingtestedby performing the appropriate genetic recombination and
on November 10, 2019 by guest
http://jvi.asm.org/
plementation studies with bacteriophages
p105
and SPO2.ACKNOWLEDGMENTS
This study was aidedby grant VC-27-Jfromthe American Cancer Society and Public Health Service grant 5-TOI-GM-00592, from the National Institute of General Medical Sciences. R.E.Y., a predoctoral trainee, completed a portion ofthis study during a Presidential Fellowship of the American SocietyforMicrobiology in the laboratory of A.T.G.
Weareindebted to G. A. Wilson and D. Figurski for their critical evaluation of thedata.
LITERATURE CITED
1. Adams, M. H. 1959. Bacteriophages. Interscience Pub-lishers, Inc., New York.
2. Bertani, G. 1958. Lysogeny. Advan. Virus Res.
5:151-193.
3. Birdsell, D. C.,G. M. Hathaway, and L. Rutberg. 1969. Characterization of temperate bacillusbacteriophage
4105.J. Virol. 4:264-270.
4. Boice, L. B. 1969. Evidence that Bacillus subtilis bacteri-ophageSP02istemperateand heteroimmune to bacte-riophage4105.J. Virol.4:47-49.
5. Boice, L. B., F. A. Eiserling, and W. R. Romig. 1969. StructureofBacillussubtilis phageSP02and its DNA; similarity of Bacillussubtilis phages SPO1,4105,and SPP1. Biochem.Biophys. Res. Commun. 34:398-403. 6. Boyer, H. W. 1971. DNA restriction and modification
mechanisms in bacteria. Ann. Rev. Microbiol. 25:153-176.
7. Chow, L. T., L. B. Boice, and N. Davidson. 1972.Mapof thepartialsequencehomologybetween DNA molecules ofBacillus subtilisbacteriophagesSP02and0105.J. Mol. Biol.68:391-400.
8. Delbruck, M. 1945. Interference between bacterial vi-ruses. III.The mutual exclusion effect and the
depres-soreffect. J.Bacteriol. 50:151-170.
9. Echols, H. 1972. Developmentalpathways for the
temper-atephage: lysisvslysogeny. Ann.Rev. Genet.
6:157-190.
10. Goldberg,I. D., and T. Bryan. 1968. Productive infection of Bacillus subtilis 168, with bacteriophage SP-10, dependent upon inducing treatments. J. Virol. 2:805-812.
11. Gwinn,D.D.,and W. D. Lawton. 1968. Alteration of host specificityin Bacillussubtilis. Bacteriol.Rev. 32:297-301.
12. Howard, B. D. 1967. Phage lambdamutantsdeficientin
rII exclusion. Science 158:1588-1589.
13. Inselburg, J. W.,T.Eremenko-Volpe, L.Greenwald,W. L.Meadow,and J. Marmur.1969.Physicalandgenetic mapping of theSP02prophageonthe chromosomeof
Bacillus subtilis 168. J. Virol. 3:627-628.
14. Ito, J., G.Mildner, and J.Spizizen. 1971.Earlyblocked asporgenousmutantsofBacillus subtilis168.I.
Isola-tionand characterization ofmutantsresistantto
an-tibioticsproduced bysporulatingBacillussubtilis168. Mol. Gen. Genet. 112:104-109.
15. Ito, J., and J. Spizizen. 1971. Abortive infection of
sporulating Bacillus subtilis 168by02bacteriophage. J. Virol. 7:515-523.
16. Jonasson, J., L. Rutberg, and F. E. Young. 1969. Lyso-genic conversion in Bacillus amyloliquefaciens H af-fecting viraladsorption. J. Virol. 4:309-310.
17. Kolenbrander, P. E., H. E. Hempfill, and H. R. Whiteley. 1973. Mixed infections of Bacillus subtilis involving bacteriophageSP02-cl. J. Virol. 11:25-34.
18. Lindahl, G., G. Sironi, H. Bialy, and R. Calendar. 1970. Bacteriophage lambda; abortive infection of bacteria lysogenic for phageP2. Proc. Nat. Acad. Sci. U.S.A. 66:587-594.
19. Losick, R., R. G. Shorenstein, and A. L. Sonenshein. 1970. Structuralalteration of RNA polymerase during sporulation. Nature (London) 227:910-913.
20. Losick, R., and A. L. Sonenshein. 1969. Change in the template specificity of RNA polymerase during sporu-lation of Bacillus subtilis. Nature (London)224:35-37. 21. Pizer, L. I., H. S. Smith, M. Miovic, and L. Pylkas. 1968.
Effect ofprophagewon the propagation of bacterio-phagesT2and T4. J. Virol. 2:1339-1345.
22. Rettenmier,C. W., and H. E. Hemphill. 1973. Prophage-mediated interference affecting the development of Bacillussubtilis bacteriophage4e.J. Virol.11:372-377.
23. Rutberg,L. 1969.Mapping of a temperatebacteriophage active onBacillus subtilis. J. Virol. 3:38-44. 24. Rutberg, L., R. W.Armentrout, and J. Jonasson. 1972.
UnrelatedAless of temperate Bacillus subtilis bacterio-phagesSP02and0105.J. Virol. 9:732-737.
25. Sakaki, Y., A. E. Karu, S. Linn, and H. Echols. 1973. Purification andproperties of they-protein specified bybacteriophage A: an inhibitorofthe host Rec BC recombination enzyme. Proc. Nat. Acad. Sci. U.S.A. 70:2215-2219.
26. Sironi, G., H. Bialy, H. A. Lozeron, and R. Calendar. 1971.Bacteriophage P2: interaction withphage lambda and with recombination deficient bacteria. Virology 46:387-396.
27. Smith, I., and H. Smith. 1973. Location of the SP02 attachment site and thebryamycinresistance marker on the Bacillus subtilis chromosome. J. Bacteriol. 114:1138-1142.
28.Yasbin, R. E., M. J. Tevethia, G. A. Wilson, and F. E. Young 1973. Analysis of steps in transformation and transfection in transformation-defective mutantsand lysogenic strains of Bacillus subtilis, p. 3-26. In L. Archer (ed.), Bacterial transformations. Academic PressInc., New York.
29. Yasbin, R. E., G. A. Wilson, and F. E. Young. 1973. Transformation and transfection inlysogenicstrains of Bacillus subtilis168.J. Bacteriol. 113:540-548. 30. Yasbin, R.E.,and F. E.Young. 1972. The influence of
temperatebacteriophageX105ontransformation and transfection in Bacillus subtilis. Biochem. Biophys. Res.Commun.47:365-371.
31. Zissler, J.,E. Signer,and F.Schaefer.1971.The role of recombination in growth of bacteriophage lambda. I. The gamma gene, p. 455-468. In A. O. Hershey (ed.),Thebacteriophagelambda. ColdSpringHarbor Monograph Series,ColdSpring Harbor,N.Y.
32. Zissler,J., E.Signer, and F.Schaefer, 1971. Therole of
recombination ingrowthofbacteriophagelambda. II. Inhibition ofgrowthby prophage P2,p.469-475.In A. 0. Hershey (ed.), The bacteriophage lambda, Cold SpringHarborMonographSeries,ColdSpring Harbor,
N.Y.
on November 10, 2019 by guest
http://jvi.asm.org/
![FIG. 2.precipitableBR151411 at Incorporation of [3H]thymidine into acid-countsin growing cultures of strains (SPO2) (0) and BR151 (SPO2) infected with an MOI of 20 (0) as described in Materials andMethods.](https://thumb-us.123doks.com/thumbv2/123dok_us/1585656.111218/4.494.43.243.58.322/precipitablebr-incorporation-thymidine-countsin-infected-described-materials-andmethods.webp)