Copyright © 1999, American Society for Microbiology. All Rights Reserved.
A Cell Line-Based Neutralization Assay for Primary Human
Immunodeficiency Virus Type 1 Isolates That Use either the
CCR5 or the CXCR4 Coreceptor
ALEXANDRA TRKOLA,1,2JAMIE MATTHEWS,1CYNTHIA GORDON,1,3TOM KETAS,1
ANDJOHN P. MOORE1,2*
The Aaron Diamond AIDS Research Center,1The Rockefeller University,2and Department of Pathology, New York University School of Medicine,3New York, New York
Received 14 June 1999/Accepted 6 August 1999
We describe here a cell line-based assay for the evaluation of human immunodeficiency virus type 1 (HIV-1) neutralization. The assay is based on CEM.NKR cells, transfected to express the HIV-1 coreceptor CCR5 to supplement the endogenous expression of CD4 and the CXCR4 coreceptor. The resulting CEM.NKR-CCR5 cells efficiently replicate primary HIV-1 isolates of both R5 and X4 phenotypes. A comparison of the CEM.NKR-CCR5 cells with mitogen-activated peripheral blood mononuclear cells (PBMC) in neutralization assays with sera from HIV-1-infected individuals or specific anti-HIV-1 monoclonal antibodies shows that the sensitivity of HIV-1 neutralization is similar in the two cell types. The CEM.NKR-CCR5 cell assay, however, is more convenient to perform and eliminates the donor-to-donor variation in HIV-1 replication efficiency, which is one of the principal drawbacks of the PBMC-based neutralization assay. We suggest that this new assay is suitable for the general measurement of HIV-1 neutralization by antibodies.
The development of an effective human immunodeficiency virus type 1 (HIV-1) vaccine is a major scientific priority (7, 23, 24, 32). It is a reasonable hypothesis that an effective vaccine will require the induction of robust humoral immunity, as well as a strong cellular response (7, 22, 23, 33). The most impor-tant component of the humoral immune response is virus-neutralizing antibodies, which reduce or eliminate the infec-tivity of cell-free virus (5–7, 42, 46, 53). Passive immunization studies in hu-PBL-SCID mice and in macaques indicate that, to be protective, neutralizing antibodies must be present at a concentration sufficient to cause virtually complete (⬎99%) neutralization in vitro (20, 37, 56, 61). This provides an impor-tant target at which vaccine designers must aim, but to gauge progress, it is necessary to be able to accurately and reliably quantitate the extent of HIV-1 neutralization (44).
At present, the generally accepted standard assay for HIV-1 neutralization is one based on the measurement of virus rep-lication in mitogen-stimulated human peripheral blood mono-nuclear cells (PBMC), commonly called the PBMC blast assay (11, 42, 77). Most of the virus production in these cultures derives from activated CD4⫹ T lymphocytes, the same cells
that are responsible for ⬎99% of HIV-1 replication in vivo (54). Furthermore, CD4⫹T lymphoblasts express both CCR5
and CXCR4 (4, 31, 39, 58, 74), the two most important core-ceptors used by primary HIV-1 isolates for replication in CD4⫹T cells and macrophages (1, 9, 12, 15, 17–19, 75, 76).
This allows the PBMC blast assay to be used with both CCR5-using macrophage-tropic isolates (R5 viruses) and CXCR4-using T-cell line-tropic isolates (X4 or R5X4 viruses) (3, 30, 41, 66). The PBMC blast assay has some drawbacks, however. First, it is not user-friendly in that considerable effort and expense are required to isolate PBMC, culture them in the presence of HIV-1, and determine the viral end point (usually the measurement of supernatant p24 antigen content by
im-munoassay). A second concern is that it takes 4 to 10 days to generate an end point, because of the kinetics of virus repli-cation and spread throughout the culture. Thirdly, donor-to-donor variation in PBMC can affect interassay performance (42).
It would be desirable to create a neutralization assay that has many of the properties of the PBMC blast assay, while improv-ing its performance and eliminatimprov-ing its drawbacks. The use of immortalized cell lines would, in principle, be advantageous. Until fairly recently, this was infeasible because only a subset of HIV-1 strains replicated in immortalized CD4⫹ T-cell lines,
since, with a few exceptions (31, 34), almost all such lines express adequate levels of CXCR4 but little or no CCR5 (31). Furthermore, passage of X4 or R5X4 primary isolates on cell lines leads to the selection of T-cell line-adapted (TCLA) strains, which are abnormally sensitive to neutralization (5–7, 42, 43, 45, 46, 53, 62, 64, 69, 72, 77). Together, these factors provide a major skew to neutralization assays based on T-cell lines. However, the cloning of CCR5 (and CXCR4) has opened up new possibilities for the development of cell line-based neutralization assays suitable for use with HIV-1 isolates of different phenotypes, as defined by coreceptor usage pat-terns (3).
The following parameters influence the creation of a cell line-based neutralization assay. The cell line must be human, since postentry restrictions on HIV-1 replication mean that unacceptably high inocula must be applied to nonprimate cells, even if they express transfected human CD4 and coreceptors, and since there is no obvious reason to select a monkey cell line over a human one. The line must express both CCR5 and CXCR4 (and of course CD4) at levels broadly comparable with those of activated PBMC, to permit the replication of HIV-1 strains of diverse phenotypes (29, 31, 55). The assay should have a rapid yet simple end point that allows the con-venient processing of multiple samples. Ideally, viral spread in culture would be minimized, so that the assay has many of the characteristics of a focal-infectivity assay, even if the end point does not involve the laborious counting of multiple plaques. * Corresponding author. Mailing address: The Aaron Diamond
AIDS Research Center, 455 First Ave., New York, NY 10021. Phone: (212) 725-0018. Fax: (212) 725-1126. E-mail: [email protected].
8966
on November 9, 2019 by guest
http://jvi.asm.org/
Despite the need for a speedy end point, the viral inoculum should not be much higher than 100 50% tissue culture infec-tive doses (TCID50), since higher inocula are proportionately difficult to neutralize by antibodies (77).
Taking the above factors into consideration, we have created and evaluated a CCR5-expressing variant of the CEM.NKR cell line, a human line that naturally expresses both CD4 and CXCR4 (25).
MATERIALS AND METHODS
Antibodies, sera, cell lines, and viral isolates.Monoclonal antibodies (MAbs) 2F5 (50) and 2G12 (68) were gifts from Hermann Katinger (University of Agriculture, Institute of Applied Microbiology, Vienna, Austria). The CD4-IgG2 molecule was provided by Paul Maddon of Progenics Pharmaceuticals Inc., Tarrytown, N.Y. (2). HeLa-CD4-CCR5 cells (also called HeLa-MAGI-CCR5 cells) were provided by Michael Emerman (Fred Hutchinson Cancer Center, Seattle, Wash.) (26, 70). GHOST-CD4-CXCR4 and GHOST-CD4-CCR5 cells were a gift from Dan Littman (New York University School of Medicine, New York, N.Y.) (66). CEM.NKR cells were obtained from the National Institute of Allergy and Infectious Diseases (NIAID) Reagent Repository (25).
The sources of the R5 viruses JR-FL and SF162, the R5X4 viruses DH123, 92US593, 2076 cl3, C7/86, SF2, and C17, and the X4 viruses 2075, 2044, ACH-H9, and NL4-3 have all been described previously (66–68). Isolates C17, SF2, ACH-H9, and NL4-3 are TCLA viruses; the others are primary viruses. Patient sera and autologous HIV-1 isolates were provided by Marty Markowitz from The Aaron Diamond AIDS Research Center patient cohort (36, 52). All five patients were treated with highly active antiretroviral therapy involving three or four drugs of which at least one was a protease inhibitor. All individuals maintained viral loads below 50 except patient no. 12, who was intermittently adherent to therapy. This patient is described in more detail by Ortiz et al. (52). Serum samples denoted by the no. 1 (after the patient number) were taken before the onset of therapy, and samples denoted by the no. 2 and above (after the patient number) were obtained sequentially during the treatment course. We could detect no influence on HIV-1 replication of any antiviral drug carried over in the serum samples (see Results and Table 3). Patients no. 33 and 245 were chron-ically infected with HIV-1, and patients no. 12, 904, and 1308 were acutely infected at the time of onset of therapy. All four patient isolates used only the CCR5 coreceptor.
Production of CEM.NKR cells stably expressing CCR5.The⌿2 ecotropic-and the PA12 amphotropic-packaging cell lines used in the production of CCR5-containing retrovirus particles were provided by David Kabat (University of Portland, Portland, Oreg.) (35, 38). These adherent cells were maintained in Dulbecco’s modified Eagle’s medium supplemented with 10% fetal calf serum (FCS), glutamine, and antibiotics. The pSFF retroviral vector containing the CCR5 gene (20g; also a gift from David Kabat) was transfected, by the calcium phosphate method, into a 1:1 coculture of ⌿2 and PA12 cells, at a total concentration of 2⫻106cells per 10-cm2dish (28). The cultures were passaged
every 3 to 4 days, with daily monitoring of CCR5 expression by fluorescence-activated cell sorting (FACS), using anti-CCR5 MAb 2D7 (Pharmingen Inc.) (73). On day 26 posttransfection, when CCR5 expression had peaked, the culture medium was removed from the cells and replaced with 10 ml of RPMI 1640 medium supplemented with 10% FCS, glutamine, and antibiotics (CEM.NKR-CCR5 cell culture medium) and containing 2⫻106CEM.NKR cells. Four days
later, the culture medium and the CEM.NKR cells were removed and trans-ferred to new tissue culture flasks. Any residual⌿2 or PA12 cells were removed, by adherence, during passaging of the CEM.NKR suspension cells. The CCR5-positive CEM.NKR cells (5⫻106) were subjected to two rounds of selection by
labeling with MAb 2D7 (2g/ml) and subsequent capture onto goat anti-mouse immunoglobulin G MicroBeads (Miltenyi Biotec Inc.). The purity of the selected CEM.NKR-CCR5 cells was verified by FACS by using the 2D7 MAb, as outlined above.
The CEM.NKR-CCR5 cells have been deposited in the NIAID Reagent Repository and may be obtained from there on request.
FACS analysis of CCR5, CXCR4, and CD4 expression.GHOST-CD4-CCR5 and -CXCR4 cells, HeLa-CD4-CCR5 cells, or CEM.NKR-CCR5 cells were analyzed on day 3 after their last passage for their expression of CCR5, CXCR4, and CD4. To do this, 3⫻105cells were washed with phosphate-buffered saline
(PBS) containing 1% bovine serum albumin and 0.05% sodium azide (staining buffer) and then incubated for 10 min at room temperature with the phyco-erythrin-labeled anti-CCR5 MAb 2D7 (Pharmingen Inc.), the phycoerythrin-labeled anti-CXCR4 MAb 12G5 (Pharmingen Inc.), the fluorescein isothiocya-nate-labeled anti-CD4 MAb SK3 (Becton Dickinson Inc.), or with appropriately labeled isotype-matched control MAbs (Becton Dickinson Inc.). The cells were washed three times with buffer, resuspended in 50l of PBS, fixed with 200l of PBS containing 1% formaldehyde, and analyzed on a FACScan instrument. The median fluorescent intensity values were derived by using CellQuest soft-ware.
HIV-1 infection of CEM.NKR-CCR5 cells and determination of tissue culture infectious dose.The CEM.NKR-CCR5 cells were maintained on a precise
pas-sage regimen (1:10 split, twice a week), because we noticed that fluctuations in their CCR5 expression and HIV-1 infectibility were dependent upon when after the day of passage the cells were used (data not shown). To prepare cells for infection assays, they were split 1:3 on the day of passage and then infected on the following day. The cells were adjusted to 2⫻105per ml in CEM.NKR-CCR5
cell culture medium and then seeded (50l) into wells of a 96-well flat-bottom microplate. HIV-1 infection was initiated by adding 100 TCID50of virus in a
volume of 100l. To maximize the extent of infection with R5 viruses, 50l of a Polybrene solution in culture medium (final Polybrene concentration, 15g/ ml) was also added. After incubation for 6 days, the supernatant medium (50l) was assayed for HIV-1 p24 antigen production by using an in-house immunoas-say, described previously (47, 68).
For determination of the TCID50values for each HIV-1 stock, 100-l aliquots
of serial dilutions of the virus preparation were used to infect the cells. The production of the p24 antigen was monitored on day 9 postinfection, and the TCID50value was calculated by using the method of Reed and Muench (59).
HIV-1 infection of HeLa-CD4-CCR5 cells.HeLa-CD4-CCR5 cells at 104per
ml in assay medium (Dulbecco’s modified Eagle’s medium supplemented with 10% FCS, glutamine, and antibiotics) were seeded into wells of a 96-well plate 1 day prior to the experiment. The cells were infected with 100 TCID50of HIV-1
in the presence or absence of 20g of Polybrene per ml. On day 4 postinfection, the cells were harvested, lysed, and analyzed for-galactosidase activity by using the Galacto-Light system (Tropix), according to the manufacturer’s instructions.
PBMC stimulation.PBMC were isolated from healthy blood donors by Ficoll-Hypaque centrifugation and then adjusted to 4⫻106per ml in RPMI 1640
medium containing 10% FCS, 100 U of interleukin 2 (a gift from Hoffmann-La Roche, Nutley, N.J.) per ml, glutamine, and antibiotics (PBMC culture medium). The cell suspensions were then divided into three equal parts and stimulated either with 5g of phytohemagglutinin (PHA) per ml, 0.5g of PHA per ml, or surface-immobilized anti-CD3 MAb TR66 (a gift from Charles Mackay, Millen-nium Inc., Cambridge, Mass.). To immobilize this MAb, culture flasks (175 cm2)
were coated overnight with MAb at 2g/ml in 12 ml of PBS. After 72 h, equal numbers of PBMC stimulated by one of these three methods were combined (referred to hereafter as three-way-stimulated PBMC) and then used in infection and neutralization assays. We have found that this triple stimulation method for PBMC activation increases the consistency with which the cells can be infected by HIV-1, with a reduction in interdonor variation (data not shown).
Infection of PBMC with HIV-1 and determination of TCID.The cell density of three-way-stimulated PBMC was adjusted to 2⫻106per ml in culture medium,
and 100l of this suspension was seeded into wells of a 96-well flat-bottom plate. The cells were infected with 100 TCID50of HIV-1 in a volume of 100l, the
cultures were incubated for 7 days postinfection, and the supernatant medium (50l) was assayed for HIV-1 p24 antigen production. Determination of the TCID50of each HIV-1 stock was performed essentially as described above. Every
virus stock that we used was titered on PBMC. The TCID50values recorded in
the Results section were derived from PBMC titrations, unless otherwise stated.
Neutralization assay with CEM.NKR-CCR5 cells.The HIV-1 inoculum was adjusted to contain approximately 1,000 to 4,000 TCID50/ml in assay medium.
Aliquots (60l) were incubated with serial dilutions of test MAbs or patient sera (60l) for 1 h at 37°C. The calculated inhibitory concentrations refer to the concentrations of the sera and antibodies in this preincubation mixture. A 100-l aliquot of the preincubation mixture was then transferred to 96-well flat-bottom microplates containing 104cells in 50l of culture medium, followed by 50l of
Polybrene-containing culture medium (final Polybrene concentration, 15g/ml). The total volume of the infection mixture was 200l, and the final concentration of virus after all dilutions was 250 to 1,000 TCID50/ml.
The infected cultures were incubated for 3 days. On day 3 postinfection, 24-well plates were prepared containing 2⫻104uninfected CEM.NKR-CCR5
cells in 1 ml of culture medium without Polybrene. Then cell samples (150l) were transferred from each well of the 96-well plate to a well of the 24-well plate containing the uninfected cells. Care was taken to thoroughly resuspend the infected cells before transfer, to ensure that equal amounts of cells were sampled from each well. The cultures in the 24-well plates were washed three times to remove residual sera and virus. By transferring cultures to the 24-well plates for the washout procedure, cell loss during this step was reduced and better assay-to-assay reproducibility was achieved. After the serum washout was completed, the cells were resuspended in 1 ml of culture medium containing 10g of Polybrene and incubated for 3 more days. On day 6 postinfection, the superna-tant medium (200l) was assayed for the HIV-1 p24 antigen. The production of the p24 antigen in the absence of serum was designated as 100%, and the ratios of p24 production in serum-containing cultures were calculated relative to this value. The reciprocal serum dilutions causing 50, 70, and 90% reduction in p24 production (the 50% infective dose [ID50], ID70, and ID90values) were
deter-mined by linear regression analysis. If the appropriate degree of inhibition was not achieved at the lowest antibody or serum dilution (e.g., 1:50), a value of ⬍1:50 was recorded. When determining the mean values derived from several independent experiments, a value marked less than or equal indicated that in one or more of these experiments no neutralization was achieved at the lowest serum dilution. In these cases, the nonneutralizing value was arbitrarily set as equal to the lowest serum dilution tested, and the calculated mean titer was reported as less than or equal to the recorded dilution. A similar procedure was used in
on November 9, 2019 by guest
http://jvi.asm.org/
experiments involving MAbs, except that the mean values are denoted as greater than or equal to the recorded concentration.
Neutralization assay with PBMC.The cell density of three-way-stimulated PBMC was adjusted to 1.25⫻106per ml in culture medium, and then 800l was
seeded into wells of a 24-well plate. The virus inoculum was adjusted to 1,000 to 4,000 TCID50/ml in culture medium and 110-l aliquots were incubated with
serial dilutions of MAbs or sera (110l) for 1 h at 37°C. The calculated inhibitory concentrations again refer to the antibody concentrations in this preincubation mixture. A 200-l aliquot of the preincubation mixture was then transferred to 24-well plates containing the stimulated PBMC, so that the total volume of the infection mixture was now 1 ml. The final concentration of virus in the cultures, after all dilutions, was therefore 100 to 400 TCID50/ml. The cultures were
washed three times at 16 h postinfection to remove free virus. The cells were resuspended in 1 ml of culture medium and then incubated for 4 days before assaying 200l of the supernatant medium for the HIV-1 p24 antigen. If virus production in the cultures had not peaked on day 4 (based on previous experi-ence with the relevant isolate), the cultures were fed with 500l of medium and reanalyzed for p24 production on subsequent days. The extent of neutralization was calculated and recorded as outlined above for work with the CEM.NKR-CCR5 cells, except that if the appropriate degree of inhibition was not achieved at the lowest serum dilution (1:10), a value ofⱕ1:10 was recorded.
RESULTS
Preparation and characterization of the CEM.NKR-CCR5
cell line.For use in neutralization assays with HIV-1 isolates of
multiple phenotypes, we decided to create a derivative of the CEM.NKR cell line that stably expressed CCR5 (CEM.NKR-CCR5 cells). Among the reasons for this choice are that CEM.NKR cells are efficient producers of HIV-1, they grow well in suspension cultures, and they express both CD4 and CXCR4 endogenously. Preliminary studies by FACS indicated that the levels of CD4 and CXCR4 expression on the parental CEM.NKR cells were comparable to those on activated PBMC (data not shown).
Retroviral vectors were used to stably introduce CCR5 into the CEM.NKR cells, as described in Materials and Methods, to make CEM.NKR-CCR5 cells (35, 38). We then evaluated the extent of CD4, CXCR4, and CCR5 expression on these cells (Fig. 1). For comparison, we measured the expression levels of these receptors on activated PBMC, HeLa-CD4-CCR5 cells (also known as HeLa-MAGI-HeLa-CD4-CCR5 cells),
GHOST-CD4-CXCR4 cells, and GHOST-CD4-CCR5 cells. Activated PBMC are the basis of the standard PBMC blast assay for HIV-1 neutralization, whereas the various GHOST and HeLa cell lines have all been used in neutralization assays (42, 66). The GHOST-CD4-CCR5 and HeLa-CD4-CCR5 cells express CXCR4 endogenously, although at very low levels in the case of the GHOST-CD4-CCR5 cells, and have been en-gineered to express CD4 and CCR5; the GHOST-CD4-CXCR4 cells were engineered to express higher amounts of CXCR4 than were expressed endogenously (42, 66).
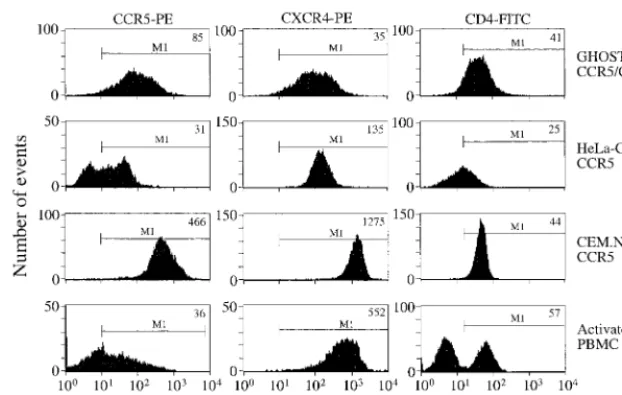
We performed FACS analyses to monitor the expression of the CD4, CCR5, and CXCR4 receptors on a variety of cell lines and PBMC (Fig. 1). Although FACS analyses of the type performed here do not provide an indication of the ab-solute number of receptor molecules on the various cell types, useful semiquantitative information can be obtained. The CEM.NKR-CCR5 cells were found to express both CD4 and CXCR4 at high levels that appear to be comparable with those found on activated PBMC, although obviously not all of the PBMC are CD4⫹ cells. In contrast, HeLa-CD4-CCR5 cells
express relatively low amounts of CD4 (Fig. 1). The level of CD4 expression on the CD4-CCR5 and GHOST-CD4-CXCR4 cells was comparable to that on PBMC (Fig. 1; not shown for CD4-CXCR4 cells). The GHOST-CD4-CCR5, GHOST-CD4-CXCR4, and HeLa-CD4-CCR5 cell lines are not clonal, as judged by their variation in receptor expression (Fig. 1). The expression of endogenous CXCR4 can clearly be detected on both the HeLa-CD4-CCR5 and the GHOST-CD4-CXCR4 cells, but at lower levels than are on activated PBMC or the CEM.NKR-CCR5 cells (Fig. 1).
The extent of CCR5 expression on the CEM.NKR-CCR5 cells was much higher than on the other two cell lines (Fig. 1). Only a fraction of the CD4⫹T cells in PBMC preparations
[image:3.612.146.457.72.269.2]stained for CCR5 after the 3 days of stimulation that are the standard conditions for their activation prior to use in neutral-ization assays, as found previously (4, 39, 58). At this time, the extent of CCR5 expression, judged by median fluorescence FIG. 1. Expression levels of CD4, CXCR4, and CCR5 on CEM.NKR-CCR5 cells. The cells were analyzed by FACS for the surface expression of CCR5, CXCR4, and CD4 with MAbs 2D7, 12G5, and SK3, respectively. Row 1, GHOST-CCR5 cells or GHOST-CXCR4 cells, as appropriate (CD4 levels were similar on the two cell lines; the GHOST-CCR5 cells are depicted); row 2, HeLa-CD4-CCR5 cells; row 3, CEM.NKR-CCR5 cells; row 4, activated PBMC (stimulated for 3 days with 5g of PHA per ml). The CCR5 and CXCR4 expression levels for PBMC are gated for the CD4⫹cell population only. The results shown are representative of one of between two and five independent experiments. The median fluorescence intensity values are recorded in the top right-hand corner of each histogram. Gate M1 depicts the positive fluorescence relative to the background staining level observed when an isotype-matched control antibody was used. PE, phycoerythrin; FITC, fluorescein isothiocyanate.
on November 9, 2019 by guest
http://jvi.asm.org/
intensity values, on the activated PBMC was approximately an order of magnitude less than that on the CEM.NKR-CCR5 cells (Fig. 1). Although the quantitative aspects of these mea-surements should not be overinterpreted, they do suggest that there are significant differences in the extent of CCR5 expres-sion on these two cell types.
Although CCR5 expression is generally stable in the CEM.NKR-CCR5 cells, we have noticed that there can be some fluctuations in the level of CCR5 staining on these cells and in their infectibility by HIV-1 when they are used too soon after initial culturing from frozen stocks (data not shown). We therefore performed infection and neutralization assays on
thawed CEM.NKR-CCR5 cells only after at least 2 weeks of continuous maintenance on a strict passage regimen (see Ma-terials and Methods).
Infection of CEM.NKR-CCR5 cells with HIV-1 isolates.We
[image:4.612.148.459.74.248.2]selected CEM.NKR-CCR5 cells with high CCR5 expression for detailed evaluation of their susceptibility to HIV-1 infec-tion (Fig. 2; Tables 1 and 2). To achieve sufficient infecinfec-tion of the CEM.NKR-CCR5 cells by R5 isolates, we found it neces-sary to add Polybrene to the cultures. We had noticed during initial evaluation of the CEM.NKR-CCR5 cells that high con-centrations of Polybrene enhanced their infection by R5 iso-lates but inhibited the infectibility of X4 viruses. These effects FIG. 2. Infectibility of CEM.NKR-CCR5 cells by HIV-1 isolates of different phenotypes. Stocks of HIV-1 JR-FL (F), SF162 (E), DH123 (‚), C17 (Œ), and NL4-3
(■) virus titers were determined on CEM.NKR-CCR5 cells in the presence of 10g of Polybrene per ml. HIV-1 replication, determined by p24 antigen production, was measured on day 9 postinfection and is recorded as the mean⫾standard deviation of four replicate cultures. JR-FL and SF162 are R5 viruses; DH123 is an R5X4 virus; and C17 and NL4-3 are X4 viruses.
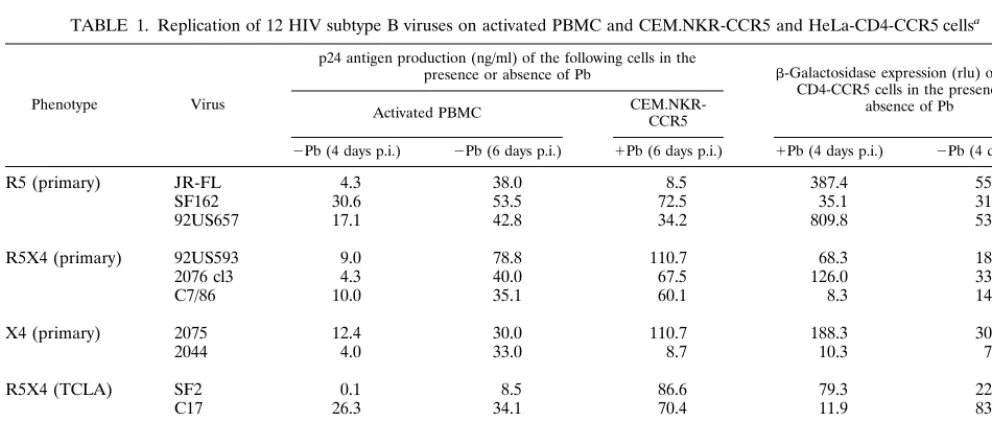
TABLE 1. Replication of 12 HIV subtype B viruses on activated PBMC and CEM.NKR-CCR5 and HeLa-CD4-CCR5 cellsa
Phenotype Virus
p24 antigen production (ng/ml) of the following cells in the
presence or absence of Pb -Galactosidase expression (rlu) of HeLa-CD4-CCR5 cells in the presence or
absence of Pb Activated PBMC CEM.NKR-CCR5
⫺Pb (4 days p.i.) ⫺Pb (6 days p.i.) ⫹Pb (6 days p.i.) ⫹Pb (4 days p.i.) ⫺Pb (4 days p.i.)
R5 (primary) JR-FL 4.3 38.0 8.5 387.4 55.8
SF162 30.6 53.5 72.5 35.1 31.6
92US657 17.1 42.8 34.2 809.8 53.0
R5X4 (primary) 92US593 9.0 78.8 110.7 68.3 18.7
2076 cl3 4.3 40.0 67.5 126.0 33.9
C7/86 10.0 35.1 60.1 8.3 14.7
X4 (primary) 2075 12.4 30.0 110.7 188.3 30.2
2044 4.0 33.0 8.7 10.3 7.5
R5X4 (TCLA) SF2 0.1 8.5 86.6 79.3 22.6
C17 26.3 34.1 70.4 11.9 83.0
X4 (TCLA) NL4-3 3.6 24.8 17.4 6.0 35.9
ACH-H9 21.2 41.2 87.3 119.6 68.0
aInfections were carried out by using 100 TCID
50of each virus stock per culture, the TCID50values having been previously determined by titration on PBMC. The
infections of PBMC (three-way stimulated) were performed in the absence of Polybrene (Pb), whereas 15g of Polybrene per ml was added for infections of the CEM.NKR-CCR5 cells. The infections of the HeLa-CD4-CCR5 cells were performed both in the presence (⫹) and absence (⫺) of 20g of Polybrene per ml. The outcome of the infections of PBMC and CEM.NKR-CCR5 cells were determined by measuring p24 antigen production on days 4 and 6 postinfection (p.i.). The infection of the HeLa-CD4-CCR5 cells was monitored by measuring-galactosidase expression on day 4, expressed as relative light units (rlu). The values shown are representative of one from two to five independent experiments of similar design.
on November 9, 2019 by guest
http://jvi.asm.org/
[image:4.612.53.549.468.679.2]of Polybrene were not unique to CEM.NKR-CCR5 cells but were also observed in studies of GHOST-CD4, U87-CD4, and HeLa-CD4 cells expressing the relevant coreceptor (data not shown).
We therefore varied the concentrations of Polybrene to identify one that would permit the efficient replication of both R5 and X4 HIV-1 isolates in the CEM.NKR-CCR5 cells. A Polybrene concentration of 10 or 15 g/ml was found to be optimal for all HIV-1 isolates tested, the optimal concentration being dependent on whether the assays were carried out in 24-well plates (10g of Polybrene per ml) or 96-well plates (15 g of Polybrene per ml). At these Polybrene concentrations, there is still some inhibition of X4 virus replication, but the use of a fixed concentration does permit the evaluation of all HIV-1 isolates by a standard protocol, without predetermina-tion of their coreceptor usage. The efficient replicapredetermina-tion of R5, R5X4, and X4 isolates on the CEM.NKR-CCR5 cells under standard conditions is shown in Fig. 2. In contrast, we could find no single Polybrene concentration that allowed efficient infection of HeLa-CD4 and GHOST-CD4 cells by all HIV-1 isolates; instead, it was necessary to perform a Polybrene ti-tration for each isolate to identify the optimal concenti-tration for that isolate, which complicated the development of a stan-dard protocol.
To study the infectibility of the CEM.NKR-CCR5 cells in more detail, we compared the ability of 12 HIV-1 subtype B isolates of different phenotypes to infect CEM.NKR-CCR5 cells, HeLa-CD4-CCR5 cells, and activated PBMC (Table 1). All the infections were set up with a viral inoculum of 100 TCID50per well, which is equivalent to 500 TCID50per ml. The infections were performed with 15g of Polybrene per ml for the CEM.NKR-CCR5 cells, with or without 20 g of Po-lybrene per ml for the HeLa-CD4-CCR5 cells, and with no Polybrene for the PBMC (Table 1). In general, HIV-1 repli-cation in the HeLa-CD4-CCR5 cells was poor compared with replication in the PBMC and CEM.NKR-CCR5 cells, which performed comparably, as monitored by -galactosidase and p24 antigen production. With many isolates, the amount of the p24 antigen released from the HeLa-CD4-CCR5 cells was too low to be detected after 4 days of culture, i.e., it was⬍10 pg/ml (data not shown).
The calculated TCID50values for the five HIV-1 isolates whose titers were determined on CEM.NKR-CCR5 cells were compared with those derived from the same isolate titers on PBMC (Table 2). The replication of both the R5 viruses was very similar on the two cell types, whereas R5X4 and X4 virus replication was reduced by 10- to 100-fold on the CEM.NKR-CCR5 cells, probably because of the inhibitory effects of Po-lybrene. However, all the isolates grew to high titers on these
cells, judged by the extent of p24 antigen release, which was usually well in excess of 10 ng/ml (Fig. 2). This provides a good dynamic range for neutralization assays. In contrast, there was considerable variation in the ability of the various isolates to infect the HeLa-CD4-CCR5 cells, and several of the isolates were only minimally infectious (data not shown). We did not routinely measure p24 antigen production from the HeLa-CD4-CCR5 cells, only the extent of activation of the -galac-tosidase reporter gene. But in many cases the extent of re-porter gene activation was inadequate to provide a workable dynamic range for neutralization assays. Overall, no correla-tion was observed between the infectibility of the various test isolates for the HeLa-CD4-CCR5 cells and either PBMC or the CEM.NKR-CCR5 cells (data not shown).
Comparison of CEM.NKR-CCR5 cells with PBMC in
neu-tralization assays. We next evaluated the suitability of the
CEM.NKR-CCR5 cells for use in neutralization assays. To do this, we used an assay protocol for the CEM.NKR-CCR5 cells that was based as closely as possible on our PBMC-based neutralization assay. We then compared neutralization titers obtained from assays of the same virus and HIV-1-positive serum samples on both cell types (Table 3). Longitudinal se-rum samples from five HIV-1-infected individuals were tested for their abilities to neutralize the autologous isolates. The highest serum dilution tested on the CEM.NKR-CCR5 cells was 1:50, whereas with PBMC it was 1:10. Human sera or plasma could not be tested at dilutions of less than 1:50 with the CEM.NKR-CCR5 cells because cytostatic effects were of-ten detectable when serum or plasma samples from uninfected people were used at lower dilutions. Such cytostatic effects can masquerade as neutralization, because moribund cells produce less HIV-1 than healthy cells.
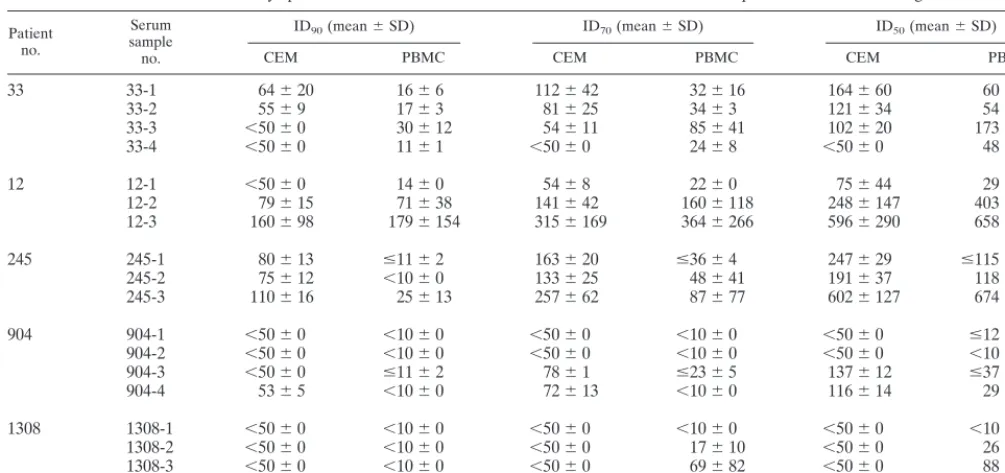
For samples from HIV-1-infected patients no. 33, 12, 245, and 904, the neutralization data obtained with CEM.NKR-CCR5 cells and PBMC were very similar. Thus, with patient no. 12, the neutralization titers obtained in both assays were almost identical, with only minor exceptions. With serum sam-ple no. 12-1, a low level of neutralization was found in the PBMC assay (ID90of 1:14) but not in the CEM.NKR-CCR5 assay (ID90 ⬎ 1:50) because high concentrations in serum could not be used with the CEM.NKR-CCR5 cells. For sam-ples from no. 1308, a marginal level of autologous neutraliza-tion was measurable in the PBMC assay (for two sera, an ID70 value was attainable) but not in the CEM.NKR-CCR5 cell assay. Of note, the virus isolate from no. 1308 replicates ex-ceptionally well in the CEM.NKR-CCR5 cells, which might reduce its sensitivity to neutralization in these cells. In contrast, with several other serum samples (e.g., no. 33-1, 33-2, 245-1, 245-2, and 245-3), more potent neutralization was observed in the CEM.NKR-CCR5 cell assay than in the PBMC assay. Overall, however, the differences in neutralization titers ob-tained in the two assays were only minor (0.6- to 7.2-fold).
Of note is that, although all of the test subjects were receiv-ing antiviral therapy, we could detect no effect of any carried-over drugs on HIV-1 replication in vitro. Thus, for all four cases, the serum sample from the second time point studied (denoted by the no. 2) caused no more inhibition of viral replication than the first sample, which was taken before ther-apy was initiated (denoted by the no. 1). We therefore con-clude that, for these samples and under the assay conditions we used, the influence of any residual drugs in serum samples is negligible.
[image:5.612.53.293.91.183.2]We next tested two anti-HIV-1 MAbs, 2F5 and 2G12, and the CD4-IgG2 molecule for their neutralization of HIV-1 JR-FL (R5 primary) and HIV-1 NL4-3 (X4 TCLA) in the CEM.NKR-CCR5 cell and PBMC assays. The neutralization TABLE 2. Determination of HIV-1 infectious titers on
CEM.NKR-CCR5 cells and PBMC
Virusa Phenotype
TCID50/mlb TCID50ratio of
PBMC/ CEM.NKR-CCR5
cells Activated
PBMC CEM.NKR-CCR5cells
JR-FL R5 104.15 104.14 1
SF162 R5 105.89 105.01 8
C17 R5X4 106.07 104.66 26
DH123 R5X4 104.67 103.44 17
NL4-3 X4 106.07 104.14 85
aThe virus stock titers were determined on PHA-stimulated PBMC and on
CEM.NKR-CCR5 cells (Fig. 2).
bThe TCID
50values were determined as described in Materials and Methods.
on November 9, 2019 by guest
http://jvi.asm.org/
titers obtained in the two assays were virtually identical in all cases (Table 4).
DISCUSSION
Our aim was to create a cell line suitable for use in HIV-1 neutralization assays that would retain the most desirable fea-tures of the current standard neutralization assay based on activated PBMC, while eliminating some of its drawbacks. Chief among the limitations of the PBMC assay are donor-to-donor variability in the kinetics or extent of HIV-1 replication and the cost, labor, and general inconvenience associated with the preparation of sufficient activated PBMC for use in large-scale assays. An immortalized cell line would be simpler and cheaper than PBMC and would allow much-needed standard-ization. But to be worthwhile, it is necessary for a cell line to express both the major HIV-1 coreceptors, CCR5 and CXCR4, to permit its use with primary isolates of different phenotypes. The cell line must also replicate HIV-1 with an efficiency compa-rable with that of activated PBMC, since otherwise skews will be introduced as a result of diminished viral throughput; obviously, the less virus that replicates, the easier it is to neutralize (77).
We chose CEM.NKR cells as the basis for creating a cell
line-based neutralization assay, since our experience is that this cell line efficiently replicates HIV-1. The CEM.NKR cells are also of lymphoid origin, which may help minimize differences between these cells and PBMC in the virus-cell attachment stage of HIV-1 infection, when there can be cell type-depen-dent effects (40). The ability of CEM.NKR cells to grow in suspension facilitates their maintenance. Furthermore, the en-dogenous expression of CD4 and CXCR4 on the CEM.NKR cells obviated the need to introduce these HIV-1 receptors by transfection. An additional factor was that CEM.NKR cells are excellent targets for antibody-mediated cellular cytotoxicity, so the introduction of CCR5 into them would create a target cell line suitable for antibody-mediated cellular cytotoxicity studies with primary R5 and X4 isolates (25).
To introduce the CCR5 coreceptor into the CEM.NKR cells, we used a retroviral gene transduction technique because of the efficiency of this method (35, 38). We selected from among the initial CCR5 transfectants of CEM.NKR cells sub-clones that expressed levels of CCR5 that were comparable with or greater than those found on activated CD4⫹T cells. If
[image:6.612.52.555.82.319.2]CCR5 levels are too low, this can impair the efficiency of HIV-1 entry, especially when CD4 expression is limiting (29, 55). Such factors can skew neutralization assays because an TABLE 3. Neutralization assays performed on CEM.NKR-CCR5 cells and PBMC with HIV-1-positive sera and autologous isolatesa
Patient no.
Serum sample no.
ID90(mean⫾SD) ID70(mean⫾SD) ID50(mean⫾SD)
CEM PBMC CEM PBMC CEM PBMC
33 33-1 64⫾20 16⫾6 112⫾42 32⫾16 164⫾60 60⫾42
33-2 55⫾9 17⫾3 81⫾25 34⫾3 121⫾34 54⫾8
33-3 ⬍50⫾0 30⫾12 54⫾11 85⫾41 102⫾20 173⫾22
33-4 ⬍50⫾0 11⫾1 ⬍50⫾0 24⫾8 ⬍50⫾0 48⫾2
12 12-1 ⬍50⫾0 14⫾0 54⫾8 22⫾0 75⫾44 29⫾12
12-2 79⫾15 71⫾38 141⫾42 160⫾118 248⫾147 403⫾419
12-3 160⫾98 179⫾154 315⫾169 364⫾266 596⫾290 658⫾307
245 245-1 80⫾13 ⱕ11⫾2 163⫾20 ⱕ36⫾4 247⫾29 ⱕ115⫾29
245-2 75⫾12 ⬍10⫾0 133⫾25 48⫾41 191⫾37 118⫾108
245-3 110⫾16 25⫾13 257⫾62 87⫾77 602⫾127 674⫾506
904 904-1 ⬍50⫾0 ⬍10⫾0 ⬍50⫾0 ⬍10⫾0 ⬍50⫾0 ⱕ12⫾3
904-2 ⬍50⫾0 ⬍10⫾0 ⬍50⫾0 ⬍10⫾0 ⬍50⫾0 ⬍10⫾0
904-3 ⬍50⫾0 ⱕ11⫾2 78⫾1 ⱕ23⫾5 137⫾12 ⱕ37⫾2
904-4 53⫾5 ⬍10⫾0 72⫾13 ⬍10⫾0 116⫾14 29⫾9
1308 1308-1 ⬍50⫾0 ⬍10⫾0 ⬍50⫾0 ⬍10⫾0 ⬍50⫾0 ⬍10⫾0
1308-2 ⬍50⫾0 ⬍10⫾0 ⬍50⫾0 17⫾10 ⬍50⫾0 26⫾23
1308-3 ⬍50⫾0 ⬍10⫾0 ⬍50⫾0 69⫾82 ⬍50⫾0 88⫾97
1308-4 ⬍50⫾0 ⬍10⫾0 ⬍50⫾0 ⬍10⫾0 ⬍50⫾0 70⫾85
aThe values shown are from two to four independent experiments. CEM, CEM.NKR-CCR5 cells.
TABLE 4. Neutralization assays performed on CEM.NKR-CCR5 cells and PBMC with MAbsa
Virus MAb ID90(mean⫾SD) ID70(mean⫾SD) ID50(mean⫾SD)
CEM PBMC CEM PBMC CEM PBMC
JR-FL 2F5 ⬎50⫾0.0 ⬎50⫾0.0 ⱖ41⫾10.9 46⫾6.9 23⫾11.6 35⫾23
2G12 1.8⫾0.9 0.9⫾0.7 0.7⫾0.2 0.1⫾0.0 0.4⫾0.2 0.1⫾0.0
CD4-IgG2 ⱕ1⫾0.7 1.3⫾0.8 0.5⫾0.3 0.5⫾0.3 0.3⫾0.2 0.2⫾0.1
NL4-3 2F5 19⫾11.8 ⱖ45⫾9.0 7.7⫾1.2 ⱖ36⫾24 5.1⫾1.4 28⫾22
2G12 1.7⫾1.4 0.3⫾0.1 0.5⫾0.5 ⬍0.1⫾0.0 0.3⫾0.2 ⬍0.1⫾0.0
CD4-IgG2 0.5⫾0.3 0.3⫾0.2 0.3⫾0.3 0.2⫾0.1 0.2⫾0.3 0.1⫾0.2
aThe values shown are from two to four independent experiments. CEM, CEM.NKR-CCR5 cells.
on November 9, 2019 by guest
http://jvi.asm.org/
[image:6.612.56.551.620.719.2]unacceptably high virus inoculum has to be used to generate a productive infection.
The CEM.NKR-CCR5 cells that we generated are stable, grow well, and are good producers of HIV-1. There are, how-ever, limitations to their use. Thus, it is necessary to use the polycation Polybrene to facilitate the infection of the CEM.NKR-CCR5 cells by R5 HIV-1 isolates. This is not un-usual; it has long been known that Polybrene increases the efficiency of retroviral infection of cell lines in culture (8, 27, 65). The reasons are not fully understood, but they may involve the overcoming of electrostatic repulsions between viruses and the anionic extracellular matrix (63), thus facilitating the virus-cell attachment stage, which is often the limiting factor in retroviral infectivity (10, 48, 51, 57, 71). The use of Polybrene does, however, reduce the infectivity of the CEM.NKR-CCR5 cells by X4 HIV-1 isolates to some extent, something which we have found also to occur with other cell lines (data not shown). We suspect that Polybrene inhibits the replication of X4 iso-lates because it partially masks the CXCR4 coreceptor, which has a strongly anionic surface charge. The binding of anti-CXCR4 MAb 12G5 to CEM.NKR-CCR5 cells is reduced in the presence of Polybrene (data not shown), and several other polycations are known to inhibit the binding of 12G5 to CXCR4 (13, 14, 49, 60).
There may be differences in the attachment properties of R5 and X4 isolates to the surfaces of human cells (40); presum-ably, X4 isolates are less dependent than R5 isolates on Poly-brene to overcome electrostatic repulsive forces. Notwith-standing these various effects of Polybrene, we were able to identify a Polybrene concentration (10 to 15g/ml, depending on the microplate well size) that permitted efficient infection of the CEM.NKR-CCR5 cells with both R5 and X4 isolates. We have not yet evaluated how this assay performs with simian immunodeficiency virus strains, but we would expect that a useful assay of simian immunodeficiency virus neutralization could be readily established with the CEM.NKR-CCR5 cells. When the CEM.NKR-CCR5 cell HIV-1 neutralization assay was compared with the standard PBMC-based assay, it per-formed comparably well. Thus, only minor variations in neu-tralization titers on the two cell types were identified. That there is no significant difference in the inherent sensitivities of the two assays was expected because the efficiency of HIV-1 neutralization is determined predominantly by the virus-anti-body interaction and not by the identity of the target cell, provided that the efficiency of HIV-1 replication is comparable (30, 41, 66, 77). More extensive evaluations of the CEM.NKR-CCR5 cell line may reveal differences in the performance of these cells and PBMC in neutralization assays with specific HIV-1 isolates or specific antibody-virus combinations. The potential for such variations under certain circumstances ap-plies to all engineered cell lines.
The principal limitation of the CEM.NKR-CCR5 cell assay in terms of its sensitivity is that serum or plasma dilutions of ⬍1:50 cannot be used, because of cytostatic effects of serum or plasma components that are apparent at higher concentra-tions; the CEM.NKR-CCR5 cells appear to be slightly more sensitive than PBMC to such factors. Thus, weakly neutralizing sera or plasmas might be undetected in the CEM.NKR-CCR5 cell assay. This problem is, however, outweighed by the general advantages of the CEM.NKR-CCR5 cell assay, in our view.
At present, the end point in the CEM.NKR-CCR5 cell neu-tralization assay involves the detection of extracellular virus production by conventional means, which can be expensive. We used a p24 antigen assay to detect virus production, but other methods, such as a reverse transcriptase assay, would also be suitable. We are presently making a variant of the
CEM.NKR-CCR5 cell line that expresses the luciferase pro-tein as a reporter of HIV-1 entry and integration, which po-tentially allows a more rapid, cheaper, and simpler detection of HIV-1 infection than can be achieved by measuring extracel-lular virus production. We have opted to pursue a luciferase reporter end point over other alternatives, such as the green fluorescent protein (GFP) or-galactosidase reporter systems. Although the detection of GFP fluorescence can be very sen-sitive, allowing a rapid neutralization assay end point (16, 21, 66), FACS is not a procedure that lends itself to the routine processing of up to hundreds of samples, as can be needed in clinical studies of HIV-1 neutralization. We chose not to pur-sue our initial studies with coreceptor-expressing GHOST cell lines, since the effort required to obtain neutralization titers was greater and more expensive than the use of PBMC, with-out any compensating increase in assay sensitivity (66). It is possible that fluorimeters could be used instead of FACS to detect GFP fluorescence, but our experience is that the use of a bulk fluorimetric end point offers no significant advantage over the detection of luciferase luminescence by a luminom-eter.
It is notable that the sensitivity of primary isolate neutral-ization in the CEM.NKR-CCR5 cells is comparable to what is measured in mitogen-stimulated PBMC. For several years, it was considered by some researchers that the failure of gp120 subunit vaccines to generate antibodies capable of neutralizing primary isolates at significant titers was the fault of the stan-dard PBMC-based neutralization assay and not of the limited immunogenicity of the vaccines (78). Here, we show that a cell line-based neutralization assay is no more or less sensitive than the standard PBMC-based assay, although it is certainly more convenient. This is consistent with the conclusion that the efficiency of HIV-1 neutralization is far more dependent on the virus-antibody interaction than the virus-cell interaction (30, 41, 66). The CEM.NKR-CCR5 cells we describe here should therefore be useful for routine evaluations of HIV-1 neutral-ization in the setting of clinical trials and vaccine-related stud-ies. The cells have been deposited in the NIAID Reagent Repository, where they may be obtained.
ACKNOWLEDGMENTS
We are very grateful to David Kabat, Mike Emerman, and Dan Littman for providing cell lines, to Hermann Katinger and Paul Mad-don for MAbs and CD4-IgG2, and to Marty Markowitz for clinical samples. We thank David Montefiori for helpful discussions and Si-mon Monard for advice and assistance with FACS procedures.
This work was supported by NIH grants R37 AI36082, RO1 AI41420, and RO1 HL59735. A.T. is a Fellow of the Austrian Program for Advanced Research and Technology; J.P.M. is an Elizabeth Glaser Scientist of the Pediatric AIDS Foundation.
REFERENCES
1.Alkhatib, G., C. Combadiere, C. C. Broder, Y. Feng, P. E. Kennedy, P. M. Murphy, and E. A. Berger.1996. CC CKR5: a RANTES, MIP-1␣, MIP-1 receptor as a fusion cofactor for macrophage-tropic HIV-1. Science272:
1955–1958.
2.Allaway, G. P., K. L. Davis-Bruno, G. A. Beaudry, E. B. Garcia, E. L. Wong, A. M. Ryder, K. W. Hasel, M.-C. Gauduin, R. A. Koup, J. S. McDougal, and P. J. Maddon.1995. Expression and characterization of CD4IgG2, a novel heterotetramer which neutralizes primary HIV-1 isolates. AIDS Res. Hum. Retroviruses11:533–540.
3.Berger, E. A., R. W. Doms, E.-M. Fenyo¨, B. T. M. Korber, D. R. Littman, J. P. Moore, Q. J. Sattentau, H. Schuitemaker, J. Sodroski, and R. A. Weiss.1998. A new classification for HIV-1. Nature391:240.
4.Bleul, C. C., L. Wu, J. A. Hoxie, T. A. Springer, and C. R. Mackay.1997. The HIV coreceptors CXCR4 and CCR5 are differentially expressed and regu-lated on human T lymphocytes. Proc. Natl. Acad. Sci. USA94:1925–1930. 5.Burton, D. R.1997. A vaccine for HIV type 1: the antibody perspective. Proc.
Natl. Acad. Sci. USA94:10018–10023.
6.Burton, D. R., and D. C. Montefiori.1997. The antibody response in HIV-1
on November 9, 2019 by guest
http://jvi.asm.org/
infection. AIDS11(Suppl. A):587–598.
7.Burton, D. R., and J. P. Moore.1998. Why do we not have an HIV vaccine and how can we make one? Nat. Med.4:495–498.
8.Castro, B., C. Weiss, L. Wiviott, and J. Levy.1988. Optimal conditions for recovery of the human immunodeficiency virus from peripheral blood to mononuclear cells. J. Clin. Microbiol.26:2371–2376.
9.Choe, H., M. Farzan, Y. Sun, N. Sullivan, B. Rollins, P. D. Ponath, L. Wu, C. R. Mackay, G. LaRosa, W. Newman, N. Gerard, G. Gerard, and J. Sodroski.1996. The -chemokine receptors CCR3 and CCR5 facilitate infection by primary HIV-1 isolates. Cell85:1135–1148.
10. Chuck, A. S., and B. O. Palsson.1996. Consistent and high rates of gene transfer can be obtained using flow-through transduction over a wide range of retroviral titers. Hum. Gene Ther.7:743–750.
11. Daar, E. S., X. L. Li, T. Moudgil, and D. D. Ho.1990. High concentrations of recombinant soluble CD4 are required to neutralize primary human immunodeficiency virus type 1 isolates. Proc. Natl. Acad. Sci. USA87:6574– 6578.
12. Deng, H., D. Unutmaz, V. N. Kewalramani, and D. R. Littman.1997. Ex-pression cloning of new receptors used by simian and human immunodefi-ciency viruses. Nature388:296–300.
13. Donzella, G. A., D. Schols, S. W. Lin, J. A. Este´, K. A. Nagashima, P. J. Maddon, G. P. Allaway, T. P. Sakmar, G. Henson, E. De Clercq, and J. P. Moore.1998. AMD3100, a small molecule inhibitor of HIV-1 entry via the CXCR4 co-receptor. Nat. Med.4:72–77.
14. Doranz, B. J., K. Grovit-Ferbas, M. P. Sharron, S.-H. Mao, M. B. Goetz, E. S. Daar, R. W. Doms, and W. A. O’Brien.1997. A small-molecule inhibitor directed against the chemokine receptor CXCR4 prevents its use as an HIV-1 coreceptor. J. Exp. Med.186:1395–1400.
15. Doranz, B. J., J. Rucker, Y. Yi, R. J. Smyth, M. Samson, S. Peiper, M. Parmentier, R. G. Collman, and R. W. Doms.1996. A dual-tropic, primary HIV-1 isolate that uses fusin and the -chemokine receptors CKR-5, CKR-2b as fusion cofactors. Cell85:1149–1159.
16. Dorsky, D. I., M. Wells, and R. D. Harrington.1996. Detection of HIV-1 infection with a green fluorescent protein reporter system. J. Acquired Im-mune Defic. Syndr.13:308–313.
17. Dragic, T., V. Litwin, G. P. Allaway, S. R. Martin, Y. Huang, K. A. Na-gashima, C. Cayanan, P. J. Maddon, R. A. Koup, J. P. Moore, and W. A. Paxton.1996. HIV-1 entry into CD4⫹cells is mediated by the chemokine receptor CC-CKR-5. Nature381:667–673.
18. Edinger, A. L., T. L. Hoffman, M. Sharron, B. Lee, B. O’Dowd, and R. W. Doms.1998. Use of GPR1, GPR15, and STRL33 as co-receptors by diverse human immunodeficiency virus type 1 and simian immunodeficiency virus envelope proteins. Virology249:367–378.
19. Feng, Y., C. C. Broder, P. E. Kennedy, and E. A. Berger.1996. HIV-1 entry cofactor: functional cDNA cloning of a seven-transmembrane G protein coupled receptor. Science272:872–877.
20. Gauduin, M.-C., P. W. Parren, R. Weir, C. F. Barbas, D. R. Burton, and R. A. Koup.1997. Passive immunization with a human monoclonal antibody pro-tects hu-PBL-SCID mice against challenge by primary isolates of HIV-1. Nat. Med.3:1389–1393.
21. Gervaix, A., D. West, L. M. Leoni, D. D. Richman, and F. Wong-Staal.1997. A new reporter cell line to monitor HIV infection and drug susceptibility in vitro. Proc. Natl. Acad. Sci. USA94:4653–4658.
22. Haynes, B. F., G. Pantaleo, and A. S. Fauci.1996. Towards an understanding of the correlates of protective immunity to HIV infection. Science271:324– 328.
23. Heilman, C. A., and D. Baltimore.1998. HIV vaccines—where are we going? Nat. Med.4:532–534.
24. Hilleman, M. R.1998. A simplified vaccinologist’s vaccinology and the pur-suit of a vaccine against AIDS. Vaccine16:778–793.
25. Howell, D. N., P. E. Andreotti, J. R. Dawson, and P. Cresswell.1985. Natural killing target antigens as inducers of interferon: studies with an immuno-selected, natural killing-resistant human T lymphoblastoid cell line. J. Im-munol.134:971–976.
26. Kimpton, J., and M. Emerman.1992. Detection of replication-competent and pseudotyped human immunodeficiency virus with a sensitive cell line on the basis of activation of an integrated beta-galactosidase gene. J. Virol.
66:3026–3031.
27. Konopka, K., L. Stamatatos, C. Larsen, B. Davis, and N. Duzgunes.1991. Enhancement of human immunodeficiency virus type 1 infection by cationic liposomes: the role of CD4, serum and liposome-cell interactions. J. Gen. Virol.72:2685–2696.
28. Kozak, S. L., and D. Kabat.1990. Ping-pong amplification of a retroviral vector achieves high-level gene expression: human growth hormone produc-tion. J. Virol.64:3500–3508.
29. Kozak, S. L., E. J. Platt, N. Madani, F. E. Ferro, Jr., K. Peden, and D. Kabat.
1997. CD4, CXCR-4, and CCR-5 dependencies for infection by primary patient and laboratory-adapted isolates of human immunodeficiency virus type 1. J. Virol.71:873–882.
30. LaCasse, R. A., K. E. Follis, T. Moudgil, M. Trahey, J. M. Binley, V. Planelles, S. Zolla-Pazner, and J. H. Nunberg.1998. Coreceptor utilization by human immunodeficiency virus type 1 is not a primary determinant of
neutralization sensitivity. J. Virol.72:2491–2495.
31. Lee, B., M. Sharron, L. J. Montaner, D. Weissman, and R. W. Doms.1999. Quantification of CD4, CCR5 and CXCR4 levels on lymphocyte subsets, dendritic cells, and differently conditioned monocyte derived macrophages. Proc. Natl. Acad. Sci. USA96:5215–5220.
32. Letvin, N. L.1998. Progress in the development of an HIV-1 vaccine. Science
280:1875–1880.
33. Letvin, N. L.1998. What immunity can protect against HIV infection? J. Clin. Investig.102:1643–1644.
34. Lusso, P., F. Cocchi, C. Balotta, P. D. Markham, A. Louie, P. Farci, R. Pal, R. C. Gallo, and M. S. Reitz, Jr.1995. Growth of macrophage-tropic and primary human immunodeficiency virus type 1 (HIV-1) isolates in a unique CD4⫹T-cell clone (PM1): failure to downregulate CD4 and to interfere with cell-line-tropic HIV-1. J. Virol.69:3712–3720.
35. Mann, R., R. C. Mulligan, and D. Baltimore.1983. Construction of a retro-virus packaging mutant and its use to produce helper-free defective retrovi-rus. Cell33:153–159.
36. Markowitz, M., M. Vesanen, K. Tenner-Racz, Y. Cao, J. M. Binley, A. Talal, A. Hurley, X. Jin, M. R. Chaudhry, M. Yaman, M. Heath-Chiozzi, J. M. Leonard, J. P. Moore, P. Racz, D. F. Nixon, and D. D. Ho.1999. The effect of commencing combination antiretroviral therapy soon after human immu-nodeficiency virus type 1 infection on virus replication and antiviral immune responses. J. Infect. Dis.179:525–537.
37. Mascola, J. R., M. G. Lewis, G. Stiegler, D. Harris, T. C. VanCott, D. Hayes, M. K. Louder, C. R. Brown, C. V. Sapan, S. S. Frankel, Y. Lu, M. L. Robb, H. Katinger, and D. L. Birx.1999. Protection of macaques against patho-genic simian/human immunodeficiency virus 89.6PD by passive transfer of neutralizing antibodies. J. Virol.73:4009–4018.
38. Miller, A. D., M. F. Law, and I. M. Verma.1985. Generation of helper-free amphotropic retroviruses that transduce a dominant-acting, methotrexate-resistant dihydrofolate reductase gene. Mol. Cell. Biol.5:431–437. 39. Mo, H., S. Monard, H. Pollack, J. Ip, G. Rochford, L. Wu, J. Hoxie, W.
Borkowsky, D. D. Ho, and J. P. Moore.1998. Expression patterns of the HIV type 1 co-receptors CCR5 and CXCR4 on CD4⫹T-cells and monocytes from cord and adult blood. AIDS Res. Hum. Retroviruses14:607–618. 40. Mondor, I., S. Ugolini, and Q. J. Sattentau.1998. Human immunodeficiency
virus type 1 attachment to HeLa CD4 cells is CD4 independent and gp120 dependent and requires cell surface heparans. J. Virol.72:3623–3634. 41. Montefiori, D. C., R. G. Collman, T. R. Fouts, J. Y. Zhou, M. Bilska, J. A.
Hoxie, J. P. Moore, and D. P. Bolognesi.1998. Evidence that antibody-mediated neutralization of human immunodeficiency virus type 1 by sera from infected individuals is independent of coreceptor usage. J. Virol.72:
1886–1893.
42. Montefiori, D. C., and T. G. Evans.1999. Toward an HIV type 1 vaccine that generates potent, broadly cross-reactive neutralizing antibodies. AIDS Res. Hum. Retroviruses15:689–698.
43. Moog, C., H. J. A. Fleury, I. Pellegrin, A. Kirn, and A. M. Aubertin.1997. Autologous and heterologous neutralizing antibody responses following ini-tial seroconversion in human immunodeficiency virus type 1-infected indi-viduals. J. Virol.71:3734–3741.
44. Moore, J. P., and D. R. Burton.1999. HIV-1 neutralizing antibodies: how full is the bottle? Nat. Med.5:142–144.
45. Moore, J. P., Y. Cao, L. Qing, Q. J. Sattentau, J. Pyati, R. Koduri, J. Robinson, C. F. Barbas III, D. R. Burton, and D. D. Ho.1995. Primary isolates of human immunodeficiency virus type 1 are relatively resistant to neutralization by monoclonal antibodies to gp120 and their neutralization is not predicted by studies with monomeric gp120. J. Virol.69:101–109. 46. Moore, J. P., and D. D. Ho.1995. HIV-1 neutralization: the consequences of
viral adaptation to growth on transformed T cells. AIDS9(Suppl. A):S117– S136.
47. Moore, J. P., J. A. McKeating, R. A. Weiss, and Q. J. Sattentau.1990. Dissociation of gp120 from HIV-1 virions induced by soluble CD4. Science
250:1139–1142.
48. Morgan, J. R., J. M. LeDoux, R. G. Snow, R. G. Tompkins, and M. L. Yarmush.1995. Retrovirus infection: effect of time and target cell number. J. Virol.69:6994–7000.
49. Murakami, T., T. Nakajima, Y. Koyanagi, K. Tachibana, N. Fujii, H. Tama-mura, N. Yoshida, M. Waki, A. Matsumoto, O. Yoshie, T. Kishimoto, N. Yamamoto, and T. Nagasawa.1997. A small molecule CXCR4 inhibitor that blocks T cell line-tropic HIV-1 infection. J. Exp. Med.186:1389–1393. 50. Muster, T., R. Guinea, A. Trkola, M. Purtscher, A. Klima, F. Steindl, P.
Palese, and H. Katinger.1994. Cross-neutralizing antibodies against diver-gent human immunodeficiency virus type 1 isolates induced by the gp41 sequence ELDKWAS. J. Virol.68:4031–4034.
51. Orloff, G. M., S. L. Orloff, M. S. Kennedy, P. J. Maddon, and J. S. McDou-gal.1991. Penetration of CD4 T cells by HIV-1. The CD4 receptor does not internalize with HIV, and CD4-related signal transduction events are not required for entry. J. Immunol.146:2578–2587.
52. Ortiz, G. M., D. F. Nixon, A. Trkola, J. Binley, X. Jin, S. Bonhoeffer, P. J. Kuebler, S. M. Donahoe, M.-A. Demoitie, W. M. Kakimoto, T. Ketas, B. Clas, L. Zhang, Y. Cao, A. Hurley, J. P. Moore, D. D. Ho, and M. Markowitz.
1999. Temporary containment of HIV-1 replication following
on November 9, 2019 by guest
http://jvi.asm.org/
tion of HAART is associated with strong virus-specific immune responses. J. Clin. Investig.104:R13–R18.
53. Parren, P. W. H. I., J. P. Moore, D. R. Burton, and Q. J. Sattentau.1999. The neutralizing antibody response to HIV-1: viral evasion and escape from humoral immunity. AIDS13(Suppl. A):S137–S162.
54.Perelson, A., A. U. Neumann, M. Markowitz, J. Leonard, and D. D. Ho.1996. HIV-1 dynamics in vivo: virion clearance rate, infected cell life-span, and viral generation time. Science271:1582–1586.
55.Platt, E. J., N. Madiani, S. L. Kozak, and D. Kabat.1997. Infectious prop-erties of human immunodeficiency virus type 1 mutants with distinct affinities for the CD4 receptor. J. Virol.71:883–890.
56.Poignard, P., R. Sabbe, G. R. Picchio, M. Wang, R. J. Gulizia, H. Katinger, P. W. H. I. Parren, D. E. Mosier, and D. R. Burton.1999. Neutralizing antibodies have limited effects on the control of established HIV-1 infection in vivo. Immunity10:431–438.
57.Poret, C. D., K. V. Lukacs, G. Box, Y. Takeuchi, and M. K. L. Collins.1998. Cationic liposomes enhance the rate of transduction by a recombinant ret-roviral vector in vitro and in vivo. J. Virol.72:4832–4840.
58.Rabin, R. L., M. K. Park, F. Liao, R. Swofford, D. Stephany, and J. M. Farber.1999. Chemokine receptor responses on T cells are achieved through regulation of both receptor expression and signaling. J. Immunol.162:3840– 3850.
59.Reed, L. J., and H. Muench.1938. A simple method of estimating fifty per cent endpoints. Am. J. Hyg.27:493–497.
60.Schols, D., S. Struyf, J. Van Damme, J. A. Este´, G. Henson, and E. De Clerq.
1997. Inhibition of T-tropic HIV strains by selective antagonization of the chemokine receptor CXCR4. J. Exp. Med.186:1383–1388.
61.Shibata, R., T. Igarashi, N. Haigwood, A. Buckler-White, R. Ogert, W. Ross, R. Willey, M. W. Cho, and M. A. Martin.1999. Neutralizing antibody di-rected against the HIV-1 envelope glycoprotein can completely block HIV 1/SIV chimeric virus infections of macaque monkeys. Nat. Med.5:204–210. 62.Spenlehauer, C., S. Saragosti, H. J. A. Fleury, A. Kirn, A.-M. Aubertin, and C. Moog.1998. Study of the V3 loop as a target epitope for antibodies involved in the neutralization of primary isolates versus T-cell-line-adapted strains of human immunodeficiency virus type 1. J. Virol.72:9855–9864. 63.Springer, T. A.1990. Adhesion receptors of the immune system. Nature
346:425–434.
64.Sullivan, N., Y. Sun, J. Li, W. Hofmann, and J. Sodroski.1995. Replicative function and neutralization sensitivity of envelope glycoprotein from primary and T-cell line-passaged human immunodeficiency virus type isolates. J. Vi-rol.69:4413–4422.
65. Toyoshima, K., and P. Vogt.1969. Enhancement and inhibition of avian sarcoma virus by polycations and polyanions. Virology38:414–426. 66. Trkola, A., T. Ketas, V. N. Kewalramani, F. Endorf, J. M. Binley, H.
Katinger, J. Robinson, D. R. Littman, and J. P. Moore.1998. Neutralization sensitivity of human immunodeficiency virus type 1 primary isolates to an-tibodies and CD4-based reagents is independent of coreceptor usage. J. Vi-rol.72:1876–1885.
67. Trkola, A., W. A. Paxton, S. P. Monard, J. A. Hoxie, M. A. Siani, D. A. Thompson, L. Wu, C. R. Mackay, R. Horuk, and J. P. Moore.1998. Genetic subtype-independent inhibition of human immunodeficiency virus type 1 replication by CC and CXC chemokines. J. Virol.72:396–404.
68. Trkola, A., A. B. Pomales, H. Yuan, B. Korber, P. J. Maddon, G. P. Allaway, H. Katinger, C. F. Barbas III, D. R. Burton, D. D. Ho, and J. P. Moore.1995. Cross-clade neutralization of primary isolates of human immunodeficiency virus type 1 by human monoclonal antibodies and tetrameric CD4-IgG. J. Virol.69:6609–6617.
69. Van Cott, T. C., V. R. Polonis, L. D. Loomis, N. L. Michael, P. L. Nara, and D. L. Birx.1995. Differential role of V3-specific antibodies in neutralization assays involving primary and laboratory adapted isolates of HIV-1. AIDS Res. Hum. Retroviruses11:1379–1392.
70. Vodicka, M. A., W. C. Goh, L. I. Wu, M. E. Rogel, S. R. Bartz, V. L. Schweickart, C. J. Raport, and M. Emerman.1997. Indicator cell lines for detection of primary strains of human and simian immunodeficiency viruses. Virology233:193–198.
71. Wang, H., R. Paul, R. E. Burgeson, D. R. Keene, and D. Kabat.1991. Plasma membrane receptors for ecotropic murine retroviruses require a limiting accessory factor. J. Virol.65:6468–6477.
72. Wrin, T., T. P. Loh, J. C. Vennari, H. Schuitemaker, and J. H. Nunberg.
1995. Adaptation to growth in the H9 cell line renders a human immuno-deficiency virus type 1 sensitive to neutralization by vaccine sera. J. Virol.
69:39–48.
73. Wu, L., G. LaRosa, N. Kassam, C. J. Gordon, H. Heath, N. Ruffing, H. Chen, J. Humblias, M. Samson, M. Parmentier, J. P. Moore, and C. R. Mackay.
1997. Interaction of chemokine receptor CCR5 with its ligands: multiple domains for HIV-1 gp120 binding and a single domain for chemokine bind-ing. J. Exp. Med.186:1373–1381.
74. Wu, L., W. A. Paxton, N. Kassam, N. Ruffing, J. B. Rottman, N. Sullivan, H. Choe, J. Sodroski, W. Newman, R. A. Koup, and C. Mackay.1997. CCR5 levels and expression pattern correlate with infectability by macrophage tropic HIV-1, in vitro. J. Exp. Med.185:1681–1691.
75. Zhang, Y.-J., T. Dragic, Y. Cao, L. Kostrikis, D. S. Kwon, D. R. Littman, V. N. Kewalramani, and J. P. Moore.1998. Use of coreceptors other than CCR5 by non-syncytium-inducing adult and pediatric isolates of human immunodeficiency virus type 1 is rare in vitro. J. Virol.72:9337–9344. 76. Zhang, Y.-J., and J. P. Moore.1999. Will multiple coreceptors need to be
targeted by inhibitors of human immunodeficiency virus type 1? J. Virol.
73:3443–3448.
77. Zhou, J. Y., and D. C. Montefiori.1997. Antibody-mediated neutralization of primary isolates of human immunodeficiency virus type 1 in peripheral blood mononuclear cells is not affected by the initial activation state of the cells. J. Virol.71:2512–2517.
78. Zolla-Pazner, S., and S. Sharpe.1996. A resting cell assay for improved detection of antibody-mediated neutralization of HIV type 1 primary iso-lates. AIDS Res. Hum. Retroviruses11:1449–1458.