0022-538X/85/050515-10$02.00/0
Copyright C 1985,American Society forMicrobiology
Isolation and Structural
Characterization of Cap-Binding Proteins
from Poliovirus-Infected HeLa Cells
KEVIN A. W. LEE, ISAAC EDERY, AND NAHUM SONENBERG*
Department of Biochemistry and McGill Cancer Center, McGill University, Montreal, Quebec, Canada H3G 1Y6 Received 2 August 1984/Accepted 8 February 1985
Inpoliovirus-infected HeLa cells, poliovirus RNA is translatedattimes whencellularmRNA translationis strongly inhibited. It is thought that this translational control mechanism is mediated by inactivation of a cap-binding protein complex (comprising polypeptidesof 24[24-kilodaltoncap-binding protein], 50,and -220 kilodaltons). Thiscomplexcanrestorethe translation ofcapped mRNAs in extracts frompoliovirus-infected cells. We havepreviouslyshown that thevirally induced defect prevents interaction betweencaprecognition factorsand mRNA.Here,weshowthatthecap-binding protein complex (andnotthe 24-kilodaltoncap-binding protein)hasactivitythatrestoresthecap-specific mRNA-proteininteraction when added to initiation factors frompoliovirus-infected cells. Thus, theactivitythatrestores thecap-specificmRNA-proteininteractionand that whichrestoresthe translation of cappedmRNAs inextractsfrom poliovirus-infected cells, copurify.The results also indicate, byanalternativeassay,that the cap-bindingprotein complex istheonlyfactor inactivated by poliovirus. We also purified cap-binding proteins from uninfected andpoliovirus-infected HeLa cells. By various criteria, the 24-kilodalton cap-binding protein is not structurally modified as a result of infection. However,the220-kilodaltonpolypeptideof thecap-binding protein complexisapparentlycleavedbyaputative viral (orinduced) protease. Byin vivo labelingandm7GDPaffinity chromatography, we isolated amodified cap-binding protein complex frompoliovirus-infected cells, containing proteolytic cleavage fragments of the 220-kilodalton polypeptide.
The cap structurem7GpppX(m)isfoundatthe 5' terminus
of almost all eucaryotic mRNAs, picornaviral and some plant viralRNAs being notable exceptions(27). Many stud-ies have indicated that the cap structure facilitates 40S
ribosome attachment to mRNA during initiation of transla-tion (2, 27), and it was anticipated that this function is
mediated by acap-specific mRNA-protein interaction.
By chemical cross-linking to 3H-labeled oxidized capped viral mRNAs, it has been shown that polypeptides of 24, 50, and 80 kilodaltons (kDa) present in crude initiation factors (IF) fromrabbitreticulocytes and severalother mammalian sources (12, 19, 28; K. A. W. Lee and N. Sonenberg, unpublished data) specifically interact with the cap struc-ture. The
identity
oftwoofthesepolypeptide
isknown: the24-kDa polypeptide corresponds to the
24,000-molecular-weight(24K)cap-binding protein (CBP) (28, 29; see below), and the50-kDapolypeptide correspondstoeIF-4Abasedon thefactthat itcanbeimmunoprecipitatedwith a monoclonal
antibody againsteIF-4A(6) and that purified eIF-4A canbe
cross-linked to mRNA with characteristics similar to those of the 50-kDapolypeptide (6, 10). Theidentityof the 80-kDa
polypeptide is not established, but several results strongly suggest that it is eIF-4B. It was shown that this factor can
specifically cross-linkto5'oxidized reovirus mRNA only in the presence of
ATP-Mg2+
(6, 10) as demonstrated for the 80-kDapolypeptideinpreparations of crude IF. In addition,cross-linking of eIF-4B requiresthe presence ofother initi-ation factors (eIF-4A [10] and CBP complex [6]).
Polypeptideswith affinityfor the cap structure have been
purified from rabbit reticulocytes by
m7GDP
affinity chro-matography. Originally, a 24K cap-binding protein(24K-CBP, CBPI, or eIF-4E) was purified by Sonenberg et al.
(30), and subsequently a high-molecular-weight complex
comprisingthe 24K-CBP andmajor polypeptides of 50 and
* Correspondingauthor.
220 kDawaspurifiedby several groupsindependently(6,11, 31). This complex is referred to as CBP II, eIF-4F, orthe
CBPcomplex (throughoutthispaper). The 50-kDa
polypep-tide is very similarto eIF-4A asdetermined by
two-dimen-sional gel analysis (11), peptide map analysis (6), and im-munoreactivity (6). Furthermore, both polypeptidesexhibit
similar mRNA cross-linking characteristics, and we will therefore refer to this polypeptide as eIF-4A. Experiments
with purified factors have shown that the CBP complex,
eIF-4B, and mRNAaresufficienttoreconstitutethe
cap-spe-cific mRNA-protein interaction observed by the chemical
cross-linking assay when crude initiation factors are used (6). These results suggest thatinteraction of the CBP com-plex and eIF-4B with the capstructure somehowfacilitates attachmentof 40S ribosomal subunits to capped mRNAs.
Poliovirus infection ofHeLa cells results in a rapid and
apparently quantitative inhibition of cellularprotein synthe-sis, such that viral RNA is almost exclusively selectedfor translation (1, 8). It was shown that crude IF from
polio-virus-infected cells could stimulate translation ofpoliovirus RNAin vitro buthadnosucheffectontranslation of cellular mRNAs(15). Consequently, various groups wereledto ask which particular initiation factorwas inactivated by polio-virus.Using different approaches, Helentjaris etal. (16)and Rose et al. (25) obtained evidence that eIF-3 and eIF-4B, respectively, were inactivated.
Theseapparentlyconflicting observationswere soon tobe reconciled. On discovery that poliovirus RNA is naturally uncapped (17, 22), it was an attractive hypothesis that inactivation of some form of CBP actually explains the shut-offofhost protein synthesis (33). In accord with this
idea, Tahara et al. (31) purified a protein complex (-8 to 10S) by m7GDP affinity chromatography which comprised
major polypeptides of24 (24K-CBP) 50, 94, and -220 kDa and which could restore translation ofcapped mRNAs in extractsfrompoliovirus-infected cells. This latteractivity is 515
on November 10, 2019 by guest
http://jvi.asm.org/
referred to as restoring activity. Recently, we have de-scribed a CBP complex (the CBP complex [6]) comprising
the 24K-CBP and polypeptides of 50 (eIF-4A) and -220
kDa,which also has restoringactivity(7). The24K-CBPcan
also be detected in preparations ofeIF-3andeIF-4B (29), as canother CBPcomplex components (9, 11), thus mostlikely explaining theeffects previously attributed to these factors
(16, 25). By a different approach (19), we analyzed CBPs
after poliovirus infection, using the chemical cross-linking assay, and showed that thecap-binding activity ofthe 24-,
50-, and80-kDacap-specific polypeptides was almost
com-pletely abolished after infection. In contrast tothis, Hansen and Ehrenfeld (12), by using the cross-linking assay,
re-ported no change in theamount ofthe 24-kDa polypeptide but did find that the 24-kDa polypeptide no longer
cosedi-ments with eIF-3 after poliovirus infection (13). These results suggested a modification to CBP which possibly prevents afunctional association between eIF-3and CBP.
Taken together, these results engender the belief that someform ofCBPis indeed inactivatedbypoliovirus,and a reportfrom Etchison et al. (9) pointed to the likely
mecha-nism. Using antisera against a 220-kDa polypeptide (P220) present inpreparations ofeIF-3 (underconditions in which
therestoring activity fractionates with eIF-3), these authors
showed that P220 is degraded in poliovirus-infected cells. Theanti-P220antibody also recognizes the -220-kDa
poly-peptide oftheCBPcomplex(9),andsoitwasproposedthat
proteolysis ofP220 by a poliovirus-dependent protease re-sults inthe shut-off ofhost protein synthesis.
Here we show that addition ofthe CBP complex to IF
from poliovirus-infected HeLa cells (I-IF) can restore the
interaction betweenthe 80-kDapolypeptide(present inI-IF) and the cap structure, asassayed bythechemical cross-link-ing technique. The 24K-CBP has no suchactivity, strongly suggestingthatrestoration ofthe80-kDacap-bindingactivity
and restoration ofcapped mRNAfunction in extractsfrom poliovirus-infected cellsare due to the same activity. In an attempt to demonstrate directly the defect in the CBP complex caused by
poliovirus,
we isolated CBPs from uninfected and poliovirus-infected HeLa cells. By various criteria, the 24K-CBP is unaltered bypoliovirus infection,
whereas the CBP complex is structurally modified. By in
vivo labelingof cells andsubsequent
m7GDP
affinity purifi-cation ofCBPs, we obtained aCBP complex from poliovi-rus-infectedHeLacellswhich containsproteolytic cleavage fragments ofP220.MATERIALSAND METHODS
Cells and virus. Mouse L-929cells and HeLaS3 cellswere grown insuspension in 10% calfserum. Infection ofLcells with reovirus type 3 (Dearing strain; 10 PFU per cell) and
virus purification were performed as previously described
(3). Infection of HeLa cells withpoliovirustype 1(Mahoney strain; 10 to 20 PFU percell, except where otherwise
indi-cated) was bythe method of Rose et al. (25).
Preparation of
methyl-3H-labeled
oxidized reovirusmRNA. Synthesisofmethyl-3H-labeled
reovirus mRNA to aspecific activityof -8 x 104cpm/,ugwithviral cores in thepresenceofS-adenosylmethionine
(specific activity,
70Ci/mmol;
New EnglandNuclearCorp.;1Ci = 3.7x1010
Bq) andperiodateoxidation were bythe method of Muthukrishnan et al. (21).
Preparation of crude protein synthesis initiation factors.
Preparation of rabbit reticulocyte
lysate,
high-salt wash of ribosomes (as a source of initiation factors), and subfraction-ation to a 0 to 40% ammonium sulfate fraction were as described by Schreier and Staehelin (26). Preparation ofHeLa cell extracts and crude initiation factors was by the
method of Lee and Sonenberg (19).
Cross-linkingof mRNAtoprotein synthesisfactors.
methyl-3H-labeled oxidized reovirus mRNA was incubated with
crude IForpurifiedCBPsorboth(asdescribed in thefigure legends) for 10 minat 30°C essentiallyas described before
(19), followed by addition of NaBH3CN
(freshly prepared
solution; AldrichChemicalCo.,
Inc.)overnight
and RNase A to digest the mRNA. The samples were resolved onsodium dodecyl sulfate
(SDS)-polyacrylamide
gels, and la-beled bandsweredetectedby fluorography.
Allcross-linking
incubationswerecarriedoutin thepresenceofATP-Mg2'
as previously described (19).Purification of CBPs. Purification of rabbit
reticulocyte
CBP
complex
wasessentially
as describedby Edery
et al.(6). A 0 to 40% ammonium sulfate fraction of ribosomal
high-salt wash was layered on 12-ml, 10 to 35% linear
sucrose
gradients
in buffer A (20 mM HEPES[N-2-hydroxyethylpiperazine-N'-2-ethanesulfonic acid] [pH 7.5],
0.2 mM
EDTA,
0.5 mMphenylmethylsulfonyl fluoride,
and 7 mM3-mercaptoethanol) containing
0.5 M KCI. Centrifu-gation wasfor24 h at 38,000rpm in an SW40rotor at4°C.
The tophalf ofthe
gradient, excluding
thefast-sedimenting
eIF-3
(>1OS),
waspooled
anddialyzed against
buffer Acontaining
0.1 M KCI and 10%glycerol.
Thedialyzed
material was then loadeddirectly
onto anm7GDP-agarose
affinity
column(6,9)
equilibrated
inbufferAcontaining
0.1 M KCI and 10%glycerol.
Nonspecifically
boundproteins
were elutedby washing
the column in 50 ml ofbuffer
Acontaining
0.1M KCIand10%glycerol,
followedby
4mlof100 ,IMGTP in thesamebuffer.
Cap-specific proteins
were thenelutedwith75 ,uM m7GTPinbufferAcontaining
0.1 MKCIand 10%
glycerol.
Rabbit
reticulocyte
24K-CBP waspurified
fromthe S100 fractionby
a modification oftheprocedure
ofTaharaetal.(31). S100 was mixed with DEAE-cellulose (3 volumes of S100and 1volumeof swollen
DEAE-cellulose)
equilibrated
in low-column buffer
(LCB;
20 mMTris[pH 7.5],
0.2 mMEDTA,
7 mMP-mercaptoethanol)
containing
80 mM KCI.The 24K-CBP binds to
DEAE-cellulose
underthese condi-tions. The resin was then washedextensively
with LCBcontaining
80 mM KCl to remove excesshemoglobin,
fol-lowedby
batchelution ofbound material with LCBcontain-ing
250 mM KCl.The eluate was thenconcentratedby
0to 50% ammonium sulfatefractionation,
theprecipitate
wasdialyzed
against
LCBcontaining
200 mMKCl,
and thedialyzed
material was diluted twofold beforebeing
loaded ontothem7GDP
affinity
column.Purification of CBP
complex
from HeLa cells was very similarto thepurification
ofrabbit CBPcomplex,
with theexception
that the crude IFs were not fractionatedby
ammonium sulfate. For each
purification,
not less than 20 litersoflog-phase
HeLacellsat acelldensity
of5x 105cells per mlwasusedtoprepare crude IF. In the caseofinfectedlysates,
infection was verifiedby
variouscriteria,
e.g., the presence of viralantigens by
immunoblotting
and the trans-lationalspecificity (capped
versusnaturally
uncapped
mRNAtranslation)
of cell extractsin in vitro translation.Purification of24K-CBPfrom HeLacellswasachieved
by
passing
totalpost-ribosomal
supernatant(S100)
over them7GDP affinity column,
followedby
elution as describedabove for the CBP
complex.
Purification of CBP complex from in vivo-labeled HeLa cells.HeLacells
(13
mlat4 x105
cellsperml)
werepelleted
andsuspended
in 8 mlof methionine-free mediumcontaining
20%
dialyzed
fetalcalfserumand 200,uCi
of[35S]methionine
on November 10, 2019 by guest
http://jvi.asm.org/
per ml (.1,000Ci/mmol, New England Nuclear). Labeling
was for6 h,after which thecells were split equally in two.
Halfwere infected with poliovirus inavolumeof400
[LI
at a multiplicity of infection of 50 PFU per cell, and the other half were mock infected. The conditions of adsorption andinfection were as described by Rose et al. (25). At 3 h
postinfection, cells werepelletedandsuspendedin 180
p1
of lysis buffer containing 150 mMNaCl, 20 mM Tris (pH 7.5), 0.5% Nonidet P-40, and 2 mMphenylmethylsulfonyl
fluo-ride. The suspension was adjusted to 600 mM potassiumacetateandthenlefttostironice for 30min.CrudeIF(-200
,ug)from uninfectedcells wasthen addedascarrier,and the
cell extracts were sedimented through a linear 10 to 30%
sucrose gradient in buffer A containing 0.5 M potassium
acetate in an SW50.1 rotor at 39,000 rpmfor 15 h. Catalase (llS) was run on a separate gradient, and the material migrating slowerthancatalase inthegradientwaspooledfor
m7GDP affinity chromatography. Thepooled fractions were dilutedwithwater to afinalpotassiumacetateconcentration of100 mM, and this material was loaded
directly
onto theaffinity column. The columnwasthen washed with 50 ml of LCB containing 100 mM KCI andthen with 20 ml of100
F.M
GDP in LCBcontaining 100 mMKCl.Thefirst 1 mlof GDP eluate wascollected. Theaffinity
resinwasthen transferredto an Eppendorf tube, and the
cap-specific polypeptides
were batch eluted with 100
p.M m7GDP
in LCB containing100 mM KCl.
Preparation of polyclonal antisera
against sheep
CBP complex was carried outby
the method of Vaitukaitis(34).
The CBP complex was purified from sheep erythrocytes under the protocol described for the
purification
of the24K-CBP from rabbit reticulocyte S100 (see above). CBP complex (20
p.g)
in LCBcontaining
500 mMKCI wasmixedwith 1.2 volumes ofcomplete Freund adjuvant. This
mate-rialwasinjectedintradermally intotheback ofarabbitinca. 20differentspots. Fourmonths
later,
therabbitwasboosted subcutaneously with 20p.g
ofantigen injected
in threedifferent places inthe back. One week
later,
the rabbit was bled through the earand serum wasprepared.
Immunoblot analysis (Western
blotting)
wasperformed
essentially as described byEdery
et al. (6).Polypeptides
were resolved on a10% SDS polyacrylamide
gel
and trans-ferredtonitrocellulose paper, and the blotwasincubatedinTris-buffered saline (pH 7.5)
containing
1% bovine serum albumin for 1 h. This was followedby incubationovernight
inTris-buffered saline
containing
1% bovine serum albumin and the anti-CBP complexantibody.
Blots were washed inTris-buffered saline andthenwereincubated with
peroxidase-conjugatedgoatanti-rabbitimmunoglobulin G;
immunoreac-tive species were visualized by color development with diaminobenzidene (32). Antisera
against
P220(prepared
as described above) were diluted 1:330 in Tris-buffered salinecontaining 1% bovine serumalbuminbefore incubation with nitrocellulose blots.
RESULTS
Taharaetal.(31)and,morerecently, Ederyetal.(7)have shown that the activity that restores translation ofcapped
mRNAs in extracts from
poliovirus-infected
HeLa cells(referred to as restoring activity) copurifies with the CBP
complex but does not reside in the 24K-CBP (the 24-kDa subunitoftheCBPcomplex).Usingadifferentapproach,we analyzedCBPs in IF from poliovirus-infectedcells(I-IF) by
thechemicalcross-linkingassay and detected reduced levels ofthe 24-, 50-,and 80-kDapolypeptides compared with the levelsdetected in IF from uninfected cells (U-IF) (19).
1 2 3 4 5 6 7 8
i5.
111
80K.ab - - -- MO
A44
. AM -l _f _f AMd_
5OKt_--I _ ~~ a~_ft_ Vm_
24K
0
0
9 10 11 12
I
_ As
_m
_4b
.'P-f +- + - + + -4 - 4
FIG. 1. Effects of purified CBPs on chemical
cross-linking
of crude IFfrompoliovirus-infectedcells.CrudeIF,purified CBPs,or mixtures ofthe two wereincubated undercross-linkingconditions with 3H-oxidizedreovirusmRNAasdescribed in thetext. Labeled polypeptideswere then resolvedon 10%SDS-polyacrylamide gels
and autoradiographed. Lanes 1 and 2 contained -100 ,ug of crude U-IF;lanes3and4, -100,ug of crude U-IFplus1.5 ,ug(containing
-0.3 ,ug of24K-CBP)of rabbitreticulocyteCBP
complex;
lanes 5 and 6, -100 ,ug ofcrudeI-IF;lanes7and8, -100 ,ugof crudeI-IF plus1.5 ,ug(containing-0.3 ,ug of24K-CBP)of rabbitreticulocyte CBPcomplex;lanes 9and10, 2,ugof rabbitreticulocyte24K-CBP fromthe S100fraction;lanes 11 and12, -100 ,ugof crudeI-IFand 2,g of rabbit reticulocyte 24K-CBP. m7GDP (0.67 mM) was includedorexcludedas indicated below thefigure.We wanted to test the
hypothesis
that the activitiesrequired
to restorecapped
mRNAfunction inextractsfrompoliovirus-infected
cells and for the interaction of thecap-specific polypeptides
with the cap structure(as
assayed by
cross-linking)
are identical and reside in the CBPcomplex.
Tothis
end,
weassayed
theability
ofpurified
CBPcomplex
to restore
cap-specific cross-linking
ofthedifferentcap-spe-cific
polypeptides.
We havepreviously
demonstrated thatcross-linking
of the CBPcomplex by
itself,
in the presence or absenceofATP-Mg2+,
results incap-specific
cross-link-ing
of the 24K-CBPonly
(6;
the samepreparation
of CBPcomplex
was used in theseexperiments).
Addition ofpuri-fied eIF-4B to the CBP
complex (in
the presence ofATP-Mg2+)
results incap-specific cross-linking
of eIF-4A and eIF-4Bin additiontothe24K-CBP(6).
These resultsstrongly
suggest that the 80-kDa
polypeptide
thatcanbecross-linkedincrude IF is eIF-4B.
Figure
1(lane
1)
shows thecross-link-ing
profile
ofa total IFpreparation
from uninfected HeLa cells.Cross-linking
of severalpolypeptides
is inhibitedby
the addition of m7GDP (compare lane 2 with lane
1),
aspreviously reported by
us(19),
andincludesthe24-, 50-,
and80-kDa
polypeptides.
The 24-kDapolypeptide
indicated in thefigure
isactually
adoubletwhichcorresponds
to the 26-and28-kDacap-specific
polypeptides
described in reference 19. We alsopreviously
reported
specific
cross-linking
ofapolypeptide
of -32 kDa in U-IF(19),
whichcanalsobeseen inFig. 1,
lane 1. The amount of thispolypeptide
varies in differentpreparations,
however,
and is oftencompletely
absent. Its
significance,
if any, is therefore notclear. Addi-tionof the CBPcomplex
to U-IFhad nostimulatory
effect oncross-linking
of the 80-kDacap-specific
polypeptide
on November 10, 2019 by guest
http://jvi.asm.org/
[image:3.612.322.555.77.271.2]present in the IF
preparation,
whereas there was a small increase in the amount of cross-linked 24- and 50-kDacap-specific polypeptides (twofold,
asdeterminedby
densi-tometryof the labeled bands;compare lanes 3 and 1 inFig.1).
Cross-linking
ofI-IF resulted in a very small amount ofspecific cross-linking
of the 24-kDapolypeptide only,
aspreviously reported
(19;Fig.
1, lanes 5 and 6). However,addition of theCBP
complex
toI-IFrestored thecross-link-ing profile
to that observed when U-IF was used alone(compare lanes 7 and 1 in
Fig.
1; the24-, 50-,
and 80-kDacap-specific polypeptides
areindicatedby
arrowheadstotheright
oflane8).
Itisnotpossible
totell from thecross-linking
in lane 7whether thecross-linked 24-and 50-kDa
polypep-tides arecontributed
by
theI-IFortheCBPcomplex, since both the 24- and 50-kDapolypeptides
are present in both fractions.However,
it is clear that theactivity required
forthe
cap-specific cross-linking
of the 80-kDapolypeptide
ispresentinthe added CBP
complex
andislacking
intheI-IFpreparation.
Since the 80-kDapolypeptide
is not presentin theCBPcomplex
(6),theonly
interpretation
ofthisexperi-mentis that
(i)
the80-kDapolypeptide (probably
eIF-4B) is notinactivated inpoliovirus-infected
cellsasassayed bythecross-linking
assay, in accord with earlier reports (5, 16),and (ii) it cannot cross-link to mRNA in I-IF because the
CBP
complex
is inactivated.Ourresultsalsoindicate(based
onanassayother than restoration of
capped
mRNAfunctionin extracts from
poliovirus-infected
cells) that the CBPcomplex
is theonly
initiation factor inactivatedduring
poliovirus
infection. It was alsoimportant
to assay theability
ofpurified
24K-CBPto restorethecap-specific
cross-linking
of the 80-kDapolypeptide
in I-IF inlight
of datashowing
that itmight
berequired
for eIF-4Bcross-linking
(10).
Thecross-linking
ofthe24K-CBPiscompletely
sensi-tive tom7GDP
(compare lanes 9 and 10 inFig.
1).Addition
of 24K-CBP toI-IFresulted in
specific cross-linking
ofthispolypeptide,
albeit at a somewhat reduced level (compare lanes12 and 10),presumably
duetocompetition
forlabeledoxidized mRNA from the vast excess of
non-cap-specific
polypeptides
present in the I-IF. Addition ofthe24K-CBP,
however,
did not enhance thecross-linking
of any otherpolypeptide
in the I-IFpreparation (Fig.
1,lanes 11 and 12).Thus,
theability
ofthe CBPcomplex
but notthe 24K-CBP to restorethecross-linking
of the 80-kDapolypeptide
(prob-ably eIF-4B)
is consistent withprevious
resultsdemonstrat-ing
that the CBPcomplex
isabsolutely
essential for thecap-specific cross-linking
ofeIF-4B(6),
and indicates that thiscomplex
is inactivatedinpoliovirus-infected cells,
con-sistent with earlier reports(7, 9, 31).
In view of the fact that the 24K-CBP by itself does not restore
cap-specific cross-linking
upon addition toI-IF, we wantedtofind outwhytheamountof24K-CBP detected bythe
cross-linking
assay isgreatly
reduced in I-IFcompared
with thatdetected in U-IF. Itwas also
important
toaddress thisquestion
inlight
ofaprevious report from Hansen and Ehrenfeld(12)
that theamount of 24K-CBP detectedbythecross-linking
assay in IFwas notreduced as aconsequence ofpoliovirus
infection. Since the CBPcomplexseems tobe the factor that isinactivated,
we tested theidea thatcross-linking
of the24K-CBPtomRNAis moreefficient when it is partof theCBPcomplex
than when it is in the free form. If this is true, then a defect in the CBP complex mightindirectly
affect thecross-linking
of the 24K-CBP. Weperformed cross-linking
experimentswithpurified24K-CBP and CBPcomplex (containing
an approximately equal amount of24K-CBP). Figure
2A shows cross-linking of 24K-CBP which iscompletelym7GDP
sensitive(lanes1and2). It should be noted that ca. eightfold less 24K-CBP was used herecompared with the amount used in Fig. 1, lanes 9 and10, and that the exposure time is different. Thisexplains
the substantial difference in the amount of cross-linked 24K-CBP observed in the two cases. When the CBP com-plex was used, there was a much higher amount of m7GDP-sensitive cross-linked24K-CBP (comparelanes 3 and 4 with lanes 1 and 2 in Fig. 2A; note that the autoradiogram is overexposed to show the cross-linked polypeptide in lane 1). Figure 2B shows Coomassie blue staining of the samples
used for the cross-linking experiments. m7GDP
affinity-pu-rified rabbitreticulocyte CBP fromribosomeswas run on a sucrose gradient in 0.5 M KCI for resolution of the free 24K-CBP and the CBP complex which otherwise copurify
on the cap affinity column. The gel shows a section of the
gradient, withlane 1 towards thetop of the gradient. Lanes 1,2,and 3 inFig. 2representcontiguousgradient fractions. Lane 1 (Fig. 2B) shows a Coomassie blue stain of the material used for cross-linking analysis in Fig. 2A lanes 1 and 2, indicating the presence ofthe24K-CBPand a small amount of the 50-kDa polypeptide. Lane 3 (Fig. 2B) is a Coomassie blue stain of the CBP complex used for
cross-linking in Fig. 2A lanes 3 and 4, showing ca. twofold more 24K-CBP than in lane 1 (Fig. 2B) and the othermajor CBP
complex polypeptides (50 and -220 kDa [6]). The presence of the 50-kDa polypeptide (eIF-4A, in lane 1) sedimenting more slowlythan the purified CBPcomplex (lanes 2 and 3)
probably means that it tends to dissociate from the CBP
complex to a slight degree during centrifugation under
high-salt(0.5 M KCI) conditions. Itshould benoted that the
A
1 2 3 4
B 12 3
-P220
-elF-4A
24k-CBP-m7GDP:
-24k-CBP
Ew.-.
- +
FIG. 2. Autoradiograph of cross-linked 24K-CBP as the free polypeptide or as part ofthe CBP complex and Coomassie blue staining of the two forms of CBP. (A) Cross-linking, SDS-poly-acrylamide gel analysis, andautoradiography were asdescribed in the text. Lanes 1 and 2contained -0.25 ,ug of24K-CBP;lanes 3 and
4contained CBPcomplex containing-0.5 ,ugof 24K-CBP. m7GDP (0.67 mM) was included in lanes 2 and 4 as indicated below the figure. (B) SDS-polyacrylamide gel analysis of purified rabbit reti-culocyte CBP sedimented through a 0.5 M KCIsucrosegradient. Rabbit reticulocyte CBP purified from ribosomes was sedimented througha10to30% linear sucrosegradientinLCBcontaining0.5 M KCItoresolve the CBPcomplexfrom thefree 24K-CBP. Samples from across the gradientwere then resolved on a 10% SDS-poly-acrylamide gel and stained by Coomassie blue. A section ofthe gradientisshown;sedimentationwasfrom lefttoright (i.e.,lane 1 is toward the topofthegradient). Materialshown in lane 1(30,ll of thefraction fromthegradient)wasusedforcross-linking analysisin A,lanes 1 and 2. Materialshown in lane 3(30p.lof the fraction from thegradient)wasusedforcross-linking analysisinA, lanes 3 and 4.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:4.612.318.553.386.553.2]amounts of 24K-CBP used in these experiments fall in the linear range for the cross-linking assay (data not shown). Thus, it is clear from these data that the cross-linking efficiency of the 24K-CBP is considerably higher when it is part of the CBPcomplex. This result most likely relates to the reduced level ofcross-linked24K-CBPobserved in I-IF, in which case the CBPcomplex isinactivated and indicates that the activity required to stimulate the cross-linking of 24K-CBP is impaired. We have also examined the transla-tional restoring activity of thefractionsshown in Fig. 2B and have found that it correlates with the presence of the 220-kDa polypeptide (data notshown), consistent with pre-vious observations (7, 31). Insummary,efficient cap-specific cross-linking of the 24K-CBP to mRNA and translational restoringactivity are bothdependent on the CBP complex. The results of Etchinson et al. (9), which indicate that poliovirus causes proteolysis of the 220-kDa polypeptide (P220) of the CBP complex, provide the first evidence of a particular structural defect in the CBP complex. Again, though, as in allpreviousattempts tocharacterize the defect in CBP caused by poliovirus, the approach was indirect. In an attempt to examine directly the abundance, structure,
and subcellular distribution of CBPs after poliovirus infec-tion, wepurifiedthemfromuninfectedorpoliovirus-infected
cells, using the m7GDPaffinity chromatography technique.
Hansen et al. (14) were able to detect the 24K-CBP by chemical cross-linking in the S100 fraction ofHeLa cells, and wehave purifiedhomogeneous 24K-CBPfrom the S100 fraction of rabbitreticulocytes.Consequently,we attempted topurify the 24K-CBPfrom theS100 fraction obtained from
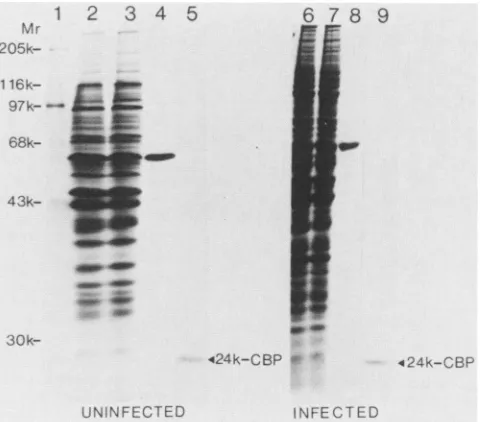
equal amounts ofuninfected or infected cells; Fig. 3 shows anSDS-polyacrylamide gelanalysisofthepurified fractions.
Severalassays wereused toverify that theinfectedfractions used as starting material for the purification were actually infected, e.g., mRNA (capped versus naturally uncapped) specificity of the corresponding cell extracts in translation
and the presence ofviral antigens byimmunoblotting. Most
of the polypeptides present in the S100 fraction are not retained during passage through the m7GDP-coupled resin (e.g.,compare lanes 2 and 3 inFig. 3, which are the load and
flow-through, respectively, from uninfected material). Elu-tion with 100 ,uM GDP shows a single polypeptide of Mr
-60,000 that either has affinity for the GDP moiety of the affinity columnor, less likely, associates with the 24K-CBP
via aGDP-sensitive interaction (Fig. 3, lane 4). The amount
and size of this polypeptide are not affected by poliovirus
infection (compare lanes 4 and 8 in Fig. 3) Elution with 100 ,uM m7GDP yielded homogenous 24K-CBP (lane 5), which
comigrates with the 24K-CBP of rabbit reticulocytes (data notshown). Again, neithertheabundancenor the size of this
polypeptide was altered after poliovirus infection (compare
lanes 5and 9 in Fig. 3).
In light of some speculation that the 24K-CBP becomes phosphorylated during poliovirus infection (18), and to ex-amine the possibility that it undergoes some other kind of
covalent modification, we performed two-dimensional gel analysis (isoelectric focusing in the first dimension and
SDS-polyacrylamide gel electrophoresis in the second
di-mension) of the m7GDP affinity-purified protein from unin-fected and infected cells. Figure 4 shows Coomassie blue staining of the two-dimensional gels. It is clear that the
polypeptidesfromuninfectedand infectedcellscomigrate in both dimensions (Fig. 4C, mixture of24K-CBP from unin-fected andinfected cells) and that there is only one species
with a slightly acidic isoelectric point of -6.5. Thus, polio-virus infection has no effect on the size, abundance, or net
Mr
205k-1 205k-
16k-
97k-
68k-
43k-1 2 3 4 5
-sop
30k-UNINFECTED
6 78
2_C
,424k-CBP
#-9
424k-C3BP
INFECTED
FIG. 3. Purification of 24K-CBPfrom the S100 fraction of unin-fected and poliovirus-inunin-fected cells. m7GDP affinity chromatogra-phy was performed as described in the text. Purified fractions were resolved on a 10% SDS-polyacrylamide gel followed by Coomassie blue staining. Lane 1, 5 ,ul of molecular weight standards, 1 mg of protein per ml (Sigma); lane 2, -100 ,ug of material from uninfected cells loaded onto the m7GDP column; lane 3, -100 p.g of flow-through from uninfected material; lane 4, 40 ,ul (from a total of 1 ml) of GDP eluate from uninfected cells; lane 5, 50p.l (from a total of 500 ,ul)of m7GDP eluate obtained from uninfectedcells; lane 6, -200,ug of material from poliovirus-infected cells loaded onto the m'GDP column; lane 7,-200,ug of flow-through from infected cells; lane 8, 40 ,ul (from a total of 1 ml) of GDP eluate from infected cells; lane 9, 50 p.l (from a total of 500p.l) of m7GDP eluate from infected cells. Note that the total amount of protein loaded on theaffinity column was the same for uninfected and infected cells.
charge of the 24K-CBP isolated from the S100 fraction; neither does it impair its ability to bind to a capanalog, since it can be retained by and specifically elutedfrom the m7GDP
affinitycolumn. It was also important to examine the charge of the 24K-CBP associated with ribosomes in relation toits distribution between the free polypeptide and the CBP complex and hence in relation to the restoring activity. We performed these experiments with CBPisolatedfrom rabbit reticulocytes, since more manageable amounts of material are obtained from this source. Analysis of ribosomal 24K-CBP from rabbit reticulocytes (either asfree24K-CBPor as partof the CBP complex) showed the presence of two major isoelectric variants as previously reported (30). There were, however, noobvious differences in therelative abundanceof these forms when those associated with the CBP complex were compared with those isolated as the free 24-kDa polypeptide (data not shown). We have not performed this analysis with HeLa ribosomal 24K-CBP, and therefore we cannot exclude the possibility that it behaves differently.
However, since the activity that can restore translation of capped mRNAs in extracts from poliovirus-infected cells is associated with the CBP complex and is absent from the 24K-CBP isolated from ribosomes, it seems clear from the latter results that the restoring activity is not related to a particularisoelectric variant of the 24K-CBP.
Therabbitreticulocyte CBPcomplex is definedassuchby several criteria: (i) co-elution of the different polypeptides from them7GDP affinity column; (ii) co-elution and stability
on November 10, 2019 by guest
http://jvi.asm.org/
[image:5.612.322.562.74.285.2]520 LEE, EDERY, AND SONENBERG
IEF
WSDS
A
4
24K
CBP
UNINFECTED
B
previously shown (6) andsomedegradation productofP220, identified as such by tryptic peptide mapping (data not shown) and the presence ofcommonantigenicdeterminants (9). Figure 5, lane 2 shows the GDP eluate obtained when material from uninfected cells was loaded on the m7GDP column. Again, as was the case forS100 fractions, a single polypeptide of Mr -60,000was eluted. This is presumably the same polypeptide as that obtained from the S100 frac-tions (Fig. 3, lane 4). Figure 5, lane 3 shows the m7GDP eluate obtained from uninfected cells. The 24-, 50-, and -220-kDa polypeptides comigrate with their reticulocyte counterparts,althoughthe -220-kDapolypeptideis asmear, presumably duetoproteolysis. In addition, therearebands of -60 and 70 kDa and other minor bands. The 60-kDa polypeptide isnotassociated with the CBPcomplexsince it can be completely removed by extensive washing of the affinity column with GDP before elution with m7GDP. The 70-kDa polypeptide is specifically eluted with m7GDP and may therefore correspond to the -70-kDapolypeptide pre-viously described in preparations of CBP II (11). These results show that the CBP complex from HeLa cells is
INFECTED
1
2
3
~
s-205k-*~
e.FF---116k
-- -9k7
MIXING
FIG. 4. Two-dimensional gelanalysis of 24K-CBP from uninfect-edandpoliovirus-infected cells.Samples of m7GDPaffinity-purified 24K-CBP(from lanes5or9 inFig. 3)wereresolvedon two-dimen-sionalgels accordingtoO'Farrell (23), followed by Coomassie blue staining. (A)0.5 ,ugof24K-CBP from uninfected cells(U-24K-CBP); (B)0.5
p.g
of 24K-CBPfrompoliovirus-infected cells(I-24K-CBP); (C) mixture of-0.3 ,ugof U-24K-CBPand -0.3 ,ugofI-24K-CBP.elF-4A- o_m
toseveralconventionalpurification stepsincluding gel filtra-tion; and(iii) cosedimentation ofcomponents of thepurified CBP complex in sucrose gradients containing 0.5 M KCI. Further indication that the 24-, 50-, and 220-kDa polypep-tides are complexed together comes from the facts that (i)
the 24-kDa polypeptide is the only polypeptide which, by itself, interacts with cap structures asassayed by chemical cross-linking (6), and (ii) the purified 50-kDa polypeptide (eIF-4A) doesnot bind to the m7GDP column(unpublished data). Thus, although the CBPcomplexhas notbeen rigor-ouslycharacterizedstoichiometricallyorbiophysically,there is good reasonto believe that it represents a homogeneous biological entity.
The CBPcomplexwas previously purifiedfrom the
high-salt washof rabbitreticulocyteribosomesbym7GDPaffinity chromatography (6, 9, 31),andweusedasimilarprotocolto purify it from HeLa cells, with the exception that the IF
were not fractionated with ammonium sulfate for technical convenience (see above). Figure 5 shows SDS-polyacryl-amidegel analysisof thepurified fractions. Lane 1 contains
asampleof rabbitreticulocyteCBPcomplex, showing major bands of 24(24K-CBP), 50 (eIF-4A), and -220kDa(P220)as
-3k30k
24K-CBP- C
0.C
o ro
E
I _4J I
o) a
0 r,(
L
4.-U
FIG. 5. Purification of CBP complex from the ribosomal salt
wash obtained from uninfectedand poliovirus-infectedcells.
Frac-tionswere purifiedon the m7GDPaffinity column,asdescribed in
the text,and resolvedon a10%SDS-polyacrylamide gel,followed
by silverstaining (20).Lane1, 0.5,ugof CBPcomplexfrom rabbit
reticulocyte ribosomes; lane2,40 ptl(fromatotal of4ml) of GDP
eluatefrom uninfectedcells;lane3, 30,u1 (from atotal of 1ml) of m7GDP eluate from uninfectedcells;lane4,40 p.1(fromatotal of4 ml)of GDP eluate frompoliovirus-infected cells;lane5,30 ,u1 (from
atotal of 1 ml)of m7GDP eluate frompoliovirus-infectedcells.
C
P220-Mr
4
5
" r .. ..7
C68kk
-43k
J.VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:6.612.81.274.73.419.2] [image:6.612.317.556.310.631.2]structurally
very similar to the rabbitreticulocyte
CBPcomplex,
a result which accords with thehigh
degree of conservation ofprotein synthesis
initiation factors betweenrabbits and humans(4).
Purification of CBP from the ribosomal high-salt wash
obtained from
poliovirus-infected
HeLa cellsyielded
dis-tinctly
differentresults(Fig. 5,
lanes 4 and 5).The samples were runon adifferent gel from that shown inlanes 1 to 3; [image:7.612.318.560.76.305.2]the
corresponding
molecularweights
are indicated in thefigure.
Lane 4showsthe GDPeluate obtainedfrom infectedcells, showing again
a -60-kDapolypeptide.
The amount and sizeof thispolypeptide
again were not changed due topoliovirus infection (compare lanes 4 and 2). There is also
staining
oneither side ofthe -60-kDapolypeptide
that isan artifact ofthe silverstaining procedure
(20) and does not representpurified polypeptides. Figure
5, lane 5 shows them7GDPeluateobtained from
poliovirus-infected
cells. There was nochange
in eithertheamount orthe sizeofthe24-kDapolypeptide,
whereas theremaining m7GDP-specific
bands weredistinctly
different.First,
the -220- and 50-kDa(elF-4A)
polypeptides
are almostcompletely
absent.Second,
therearetwonewbandsof-130kDathatarenotpresent in thepreparation
from uninfectedcells(compare lanes5 and3in
Fig.
5). Although
it should be borne in mind that silverstaining
ofpolypeptides
is notnecessarily quantitative,
it does appear that the amountofthe -130-kDapolypeptides
is
significantly
less thanthe amountof24-kDapolypeptide,
particularly
on amolar basis. The presence of 24K-CBPin them7GDP
eluate obtained from infected cells serves as a useful internal control and arguesstrongly against
nonspe-cific loss ofthe other CBP
complex
polypeptides (50
and -220kDa). Furthermore,
theobservationthathomogeneous
24K-CBP is obtainedby m7GDP affinity purification
ofthepostribosomal
supernatantfrominfected cells indicatesthat theCBPcomplex
isnotmerely
redistributed inthe infectedcell such that it no
longer
associateswith ribosomes.Thus,
a reasonable
interpretation
ofthese resultsis that theCBPcomplex
is modified afterpoliovirus
infection. Thesignifi-canceof the -130-kDa
polypeptides
will be addressed later inlight
of the results shown inFig.
6andother data.The
purification
oflarge
amounts of the CBP complex fromHeLa cells isa somewhat cumbersome andtime-con-suming activity, yielding only
ca. 50 ,ug ofCBPcomplex
from1010 log-phase
HeLacells(20
liters ofcells at 5 x 105 cells perml).
There are inaddition many steps between thecell harvest and the m7GDP
affinity purification,
possibly
contributing
to artifactualdisintegration
ofthe native CBPcomplex
as it exists in the cell.Consequently,
we decided to label mock- andpoliovirus-infected
cells with[35SJmethionine
and attempt to isolate the CBPcomplex
by
a fasterprotocol.
Cells were labeled for 6 h with[35S]methionine.
At the end of thistime,
the cells were dividedequally
in two; halfwere infected withpoliovirus
and the other half were mock-infected. To monitor the
infectionwe
performed
amock-labeling experiment
in which[35S]methionine
was added at 2.5 hpostinfection
toprevi-ously
unlabeledcells,
in the presence and absenceofpolio-virus. The in vivo
labeling
pattern observed between 2.5 and 3 hpostinfection
(at which time the cell extracts wereprepared)
confirmedthat shut-off of cellularprotein
synthe-siswas
complete
and thatvirus-specific proteins
werebeing
synthesized (data
notshown).
Figure
6 shows the results of thepurification
of[35S]methionine-labeled
CBPfromuninfectedandpoliovirus-infected HeLa cells. Elution of the
m7GDP
columnwith100,uM
GDPyielded
asingle polypeptide
ofMr-60,000 (lane 1,2 3 4 Mr 5 6 7 8
am
-220k-pw
_w-a
-9 -P220
wa
-r
--W-60k
-50k
-24k
u u
-24k-CBP
u u
FIG. 6. Purification of in vivo-labeled CBP complex from unin-fected and poliovirus-infected HeLacells. Fractions werepurified on anm7GDPaffinitycolumnasdescribed in the text, resolvedona 10% SDS-polyacrylamide gel, and autoradiographed. Lane 1,40,ul (fromatotalof1 ml) of GDP eluate obtained from uninfected cells; lane2,40,u (from atotal of 1 ml) of GDP eluate from poliovirus-infectedcells; lane 3,50pI(fromatotalof 500pI)ofm7GDP eluate from uninfectedcells;lane4,50p.1(fromatotalof 500p1)ofm7GDP eluate from poliovirus-infected cells. Polyclonal antiserum against theCBPcomplexwasusedto probeextractsfrom uninfected and poliovirus-infected HeLa cells for P220 related antigens. S10 ex-tracts wereresolvedon a10% SDS-polyacrylamide gel, followed by Western blottingasdescribed inthe text. Thefigure shows immuno-reactivespeciesinlanes 5to9. Lanes5and 6, 150pugofprotein from differentS10extractsfrom uninfectedcells; lanes7and8,150p.g of protein from different S10extracts from poliovirus-infected cells; lane 9,5pugof rabbit reticulocyte CBP complex.
uninfected). This is presumably the same polypeptide ob-served when unlabeled materialwasusedfor the purification of 24K-CBP from the S100 fraction (e.g., Fig. 3, lane 4).
Again, the amount and size of this polypeptide were not
affectedby poliovirusinfection (compare lane 2withlane1). Lane 3 shows the m7GDP eluate obtained from uninfected material(includingthe 60Kpolypeptide whichisparticularly
abundant and isnotcompletelywashedoffduringtheelution
with GDP in this experiment). The material eluted has
polypeptides comigrating with the 24-, 50-, and 220-kDa
polypeptidesof rabbit reticulocyteCBP complex(indicated
by molecularweight to the right oflane 4). However, we havenoevidenceatpresenttoprovethat the 24-and 50-kDa
polypeptides are indeed the 24K-CBPand eIF-4A,
respec-tively. The other bands (Mr = 35,000 and 70,000, etc.) are either relatedto theCBPcomplex-since theyare
specific-allyeluted with m7GDP(compare lanes 3 and 1)-or, alter-natively, are other proteins which bind specifically to the column(thecriterion forspecificity beingelution with m7GDP andnotwithGDP).Since the loadontothecolumncontains totalsoluble cellproteinand alsoproteins solubilizedby0.5 MKCI and 0.5% NonidetP-40, it is
likely
thatatleastsome of the additional polypeptides are not related to the CBPcomplex. For example, likely candidates are
putative
nu-clearCBPsaspreviously reported
(24). Anotherpossibility
on November 10, 2019 by guest
http://jvi.asm.org/
is that the additional polypeptides are loosely associated
with the CBP complex but are lost during the purification protocols
previously
employed (6, 11, 31).Itshould benoted that the relativelabeling
intensities of the 24-, 50-, and -220-kDa polypeptides are not equal. This might be ac-countedfor in partbythe size of thepolypeptides (assuming an average methionine content for eachpolypeptide)
but may also reflect differentratesofentry ofthe newly synthe-sized components into the CBP complex. Inaddition,
the relativelabeling
intensities of the 24-, 50-, and 220-kDapolypeptides
vary among differentpreparations
fromunin-fected cells
(unpublished
data). Whenmaterial from poliovi-rus-infected cells was loaded onto the column, the m7GDPeluate obtained was
distinctly
different from that obtained from uninfected material(Fig.
6, lane 4). There was nochange
in either the abundance or the size ofthe 24- and 50-kDapolypeptides,
whereas in contrast there was no detectable -220-kDapolypeptide.
Instead, therewereaddi-tional bands ofMr -130,000 (indicatedby arrows)thatwere
completely
absent from the m7GDP eluate obtained from uninfected cells (compare lane 4 withlane 3 in Fig. 6). Thesize and abundance of all the other bands present in the
m7GDP
eluate are also not affected by poliovirus infection (comparelane 4 with lane 3).These results demonstrate that P220 is cleaved by a
putative
viral (orinduced) protease, but that thepresumedcleavage products (-130-kDa polypeptides)
are stillretained andcanbespecifically
eluted fromthem7GDP affinity
resin. This suggests that the presumed cleavage products remain associatedwithacap-binding
component(mostprobablythe24K-CBP)
intheform ofamodified CBP complex. Itseems to usextremelyunlikely,
althoughadmittedly
notprecluded,that the
cleavage
products derivedfromP220 would have acryptic m7GDP binding
site and thus bind directly to them7GDP affinity
resin. The difference in the amounts of the -130-kDa cleavage products obtained in Fig. 6 (lane 4) ascompared
withFig.
5 (lane 5) suggest that the putative modified CBPcomplex
is not stable to the purification protocolemployed
fortheexperiments in Fig. 5. This may alsoexplain
theabsenceof the 50-kDapolypeptideinFig.5. Theexistenceofproteolytic
cleavagefragments
of P220 ofMr
-130,000
accords withtheoriginal
observation ofEtch-ison et al. (9), who demonstrated the appearance of such
polypeptides
in crude HeLa cell extracts afterpoliovirus
infection. In this case, thecleavage
products observed arethought
to berelated toP220oftheCBPcomplex byvirtueofcommon
antigenicity.
Wehaverecently
raisedpolyclonal
antibodies to the
purified
sheep CBP complex which bindstrongly
tothe 24- and 220-kDapolypeptides
of rabbit CBPcomplex
(Fig.
6, lane 9).Consequently,
we probed HeLacell extracts with this antiserum and obtained results very
similar tothose of Etchisonet al. (9). In
Fig.
6, lanes 5 and6, differentextracts from uninfected
cells,
and in lanes 7 and8,
different extracts frompoliovirus-infected cells,
wereprobed
with theanti-CBPcomplexantiserum. The antiserum does not react with the HeLacell 24K-CBP (lanes 5to 8).However,
the antiserum clearly reacts with the -220-kDapolypeptide
presentinextractsfrom uninfected cells(lanes
S and 6), and this antigen comigrates with P220 of the rabbit CBPcomplex
(compare lane 5 with lane 9). There is no detectable 220-kDapolypeptide in extractsfrompoliovirus-infected HeLa
cells,
whereas thereare putative degradationproducts
OfMr
-130,000(Fig. 6, lanes7and8,indicatedby arrows). These latter bands comigrate with the cleavage products present in the putative modified CBP complex isolatedbym7GDP
affinity chromatographyfrompoliovirus-infected cells (compare lane 7 or 8 with lane 4), strongly
suggesting thatthey areidentical. DISCUSSION
Analysis of eucaryotic mRNA CBPs by the chemical cross-linking assay indicatesacomplex interactionprimarily
between polypeptides of 24, 50, and 80 kDa, ATP, and mRNA. This interaction is presumed to facilitate 40S ribosomal subunit attachment to cellular mRNA during translation and is very discretely prevented uponpoliovirus infection of HeLa cells, resulting in shut-off of cellular protein synthesis. Recently, Edery et al. (6) have demon-strated that the cap-specific mRNA-protein interaction ob-served between crude IF and mRNA can bereconstituted by using the CBP complex (containing eIF-4A as a subunit), eIF-4B, and mRNA. These factors appear to interact with mRNA in close concert, since the m7GDP-sensitive cross-linking of eIF-4A as part of the CBP complex is strictly dependent on eIF-4B and the cross-linking of eIF-4B is likewise dependent on the CBP complex. This idea accords with the simultaneous loss of the cross-linking ability of all thecap-specific polypeptides afterpoliovirus infection (19), againconsistent withaclosefunctionalrelationshipbetween them.Thus, the virally induced lesion in the CBP complex is probably sufficient by itselftopreventinteraction of eIF-4A and the 80-kDapolypeptide (probably eIF-4B) with the cap structureandconsequentlyblocks40Sribosomeattachment to cellular mRNAs. The CBP complex has activity which restores the specific cross-linking profile when added to IF
from infected cells. This activity is not present in the
24K-CBPand thuscopurifieswith thetranslationalrestoring activity, strongly suggesting that the two activities are identical. The fact that eIF-4A and eIF-4B are neither
structurally modified (5) norfunctionally impaired(16) after
poliovirus infection is consistent with ourobservation that the 80-kDa polypeptide (probably eIF-4B) is present and active in IF from infected cells, at least as assayed by chemical cross-linkingto mRNA in the presenceof exoge-nous CBPcomplex.
The 24K-CBP from infected cells, either as the free
polypeptide or as part of the putative modified CBP com-plex, can recognize the cap structure, as indicated by the fact that it canbe purified by m7GDPaffinity
chromatogra-phy (Fig. 3 and 5). However, the amount of cross-linked 24K-CBP in I-IFis considerablylower than that from U-IF. Inlight ofourfindingthat thecross-linkingof the 24K-CBP from rabbitreticulocytesisgreatly enhanced when it is part of the CBPcomplexascomparedwith the freepolypeptide,
it seems likely that the 24K-CBP in the putative modified CBPcomplexfrompoliovirus-infectedcells behaves like the free 24-kDapolypeptideintermsofcross-linkingtomRNA. This again points to asignificant role for P220 in mediating the interaction between the 24K-CBP and mRNA.
We have presented direct eivdence that the 24K-CBP is notstructurallymodified afterpoliovirusinfection. Further-more, the subcellular distribution of the 24K-CBP is not changed. In contrast, the native CBP complex cannot be
purifiedfrom anyfraction obtained frompoliovirus-infected
cells. These resultsdemonstratedirectlythatthe native CBP
complex is modified by poliovirus infection. The in vivo
labeling experimentsindicate thata modified CBPcomplex
exists in infected cellswhich contains theproteolytic cleav-ageproducts ofP220, and possibly eIF-4A. Similar results are obtained for purification of unlabeled ribosomal CBP from infectedcells, although in thiscaseeIF-4Aisdefinitely
on November 10, 2019 by guest
http://jvi.asm.org/
absent andthe amountof the-130-kDa
polypeptides
seemssignificantly
reduced.Thus,
theexact structure of the mod-ified CBPcomplex
is uncertain. In otherexperiments,
we have obtained the24K-CBPinfree form from ribosomalhighsalt washofinfected
cells,
which suggests that the modified CBPcomplex
is unstable. Inaddition,
the eIF-4A compo-nent of the CBPcomplex
isapparently
not asstrongly
associated withthe CBPcomplex
asthe other components, as indicatedby
the observation that a small amount of eIF-4A dissociates from the CBPcomplex
underhigh-salt
conditions(Fig. 2B).
Thismight
wellexplain
the lack of eIF-4A in theribosomal CBP isolated frominfectedcells,
inwhich caseP220is
cleaved, possibly resulting
indecreasedaffinity
of eIF-4Afor othercomplexed
components. In any event, theresultspresented
here suggest thatanintactP220 is essential for CBPcomplex
function.First,
efficientcap-specific cross-linking
of the24K-CBP, eIF-4A,
and eIF-4B isdependent
on the CBPcomplex
and does not occur afterpoliovirus
infection.Second, restoring activity
islikewise a propertyofthe CBPcomplex
andcorrelates with the pres-enceof P220asopposed
tothe24K-CBPoreIF-4A.Whether or not the association of the modified CBPcomplex
withribosomes from infected cells reflects an involvement in translation of
poliovirus
RNA remains to be determined.Indeed,
the mechanismby
whichpoliovirus
RNA initiatestranslation is
something
ofamystery, both in terms ofanypossible
roleofCBP(s) (modified
orotherwise)
andconcern-ing
which structural features of the viral messenger allow efficienttranslation intheabsence ofthe cap structure.The
question concerning
therelationship
between thevirus-dependent
protease that cleaves P220 and thepoliovi-rus
replicative
cycle
nowchallenges.
Onepossibility
is that theactivity
that processes thepoliovirus
primary cleavage
products
(3C) is alsoresponsible
forcleavage
of P220.However,
this appearsunlikely,
inlight
ofour recent data thatantibodies directedagainst poliovirus
protein
3C donot inhibitcleavage
of P220invitro(K.
A. W.Lee,
IsaacEdery,
R.
Hanecak,
E.Wimmer,
and N.Sonenberg,
submitted forpublication). Therefore,
theremight
be another viral prote-ase involved in thiscleavage
or induction of a cellularfunction, possibly
one which is involved inregulation
ofprotein synthesis
in a broader sense. Theavailability
of mutants derived from infectious clonedpoliovirus
DNA should aid inapproaching
theseproblems.ACKNOWLEDGMENTS
We extendourthankstoChristiane BabinandSheelin Howard for assistancein preparationof viralRNA.
This researchwassupported bygrantsfrom the Medical Research Council and the NationalCancer Institute of CanadatoN.S. N.S. is therecipientofaTerryFoxCancerResearchScientist awardof the National Cancer Institute of Canada. K.A.W.L. and I.E. are
predoctoralfellows oftheCancer Research SocietyofMontreal.
LITERATURE CITED
1. Baltimore,D.1969. Thereplicationofpicornaviruses,p. 101-176. In H. B. Levy (ed.), The biochemistry of viruses. Marcel Dekker, Inc., NewYork.
2. Banerjee, A. K. 1980. 5'-Terminal cap structurein eucaryotic messengerribonucleic acids. Bacteriol.Rev. 44:175-205. 3. Banerjee,A.K.,and A.J.Shatkin.1970.Transcriptionin vitro
byreovirus-associated ribonucleicacid-dependent polymerase. J. Virol. 6:1-11.
4. Brown-Luedi,M.L.,L.J.Meyer,S.C.Milburn,P.M.-P.Yan, S. Corbett, and J. W. B. Hershey. 1982. Protein synthesis initiation factors from human HeLa cells and rabbit reticulo-cytesaresimilar:comparisonofproteinstructure,activities and immunologicalproperties. Biochemistry21:4202-4206.
5. Duncan, R., D. Etchison, andJ. W.B. Hershey. 1983. Protein synthesis initiation factors eIF-4A and eIF-4Bare notalteredby poliovirus infection of HeLa cells. J. Biol. Chem. 258:7236-7239. 6. Edery, I., M. Humbelin, A. Darveau, K. A. W. Lee, S.Milburn, J. W. B. Hershey, H. Trachsel, and N. Sonenberg. 1983. Involve-mentof eukaryotic initiation factor 4A in the cap recognition process. J. Biol. Chem.258:11398-11403.
7. Edery, I., K. A. W. Lee, and N. Sonenberg. 1984. Functional characterizationofeukaryotic mRNA cap binding protein
com-plex: effects on translation of capped and naturally uncapped RNAs. Biochemistry 23:2456-2462.
8. Ehrenfeld, E. 1982. Poliovirus-induced inhibition of host cell protein synthesis.Cell 28:435-436.
9. Etchison, D., S. C. Milburn, I. Edery, N. Sonenberg, and J. W. B. Hershey. 1982. Inhibition of HeLa cellprotein synthe-sisfollowingpoliovirus infection correlates with the proteolysis of a 220,000-dalton polypeptide associated with eucaryotic initiation factor 3 and acapbinding protein complex. J. Biol. Chem. 257:14806-14810.
10. Grifo, J. A., S. M. Tahara, J. P. Leis, M. A. Morgan, A. J. Shatkin, and W. C. Merrick. 1982. Characterization of eukaryo-tic initiation factor4A, a protein involved in theATP-dependent bindingofglobinmRNA.J. Biol. Chem. 257:5246-5252. 11. Grifo, J. A., S. M. Tahara, M. A. Morgan, A.J. Shatkin, and
W. C. Merrick. 1983. New initiator factoractivity required for globinmRNA translation. J. Biol. Chem.258:5804-5810. 12. Hansen, J.,and E.Ehrenfeld.1981.Presence of the cap-binding
protein in initiation factor preparations frompoliovirus-infected HeLa cells. J. Virol.38:438-445.
13. Hansen, J., D. Etchison, J. W. B. Hershey, and E. Ehrenfeld. 1982. Association ofcap-binding proteinwitheucaryotic initia-tion factor 3in initiation factorpreparationsfromuninfected and poliovirus-infectedHeLa cells. J. Virol. 42:200-207.
14. Hansen, J. L., D. 0. Etchison, J. W. B. Hershey, and E. Ehrenfeld. 1982. Localization ofcap-bindingprotein in subcel-lularfractions of HeLacells. Mol. Cell. Biol. 2:1639-1643. 15. Helentjaris, T., and E. Ehrenfeld. 1978. Control of protein
synthesis in extracts from poliovirus-infected cells. I. mRNA discriminationby crude initiationfactors. J. Virol. 26:510-521. 16. Helentjaris, T.,E.Ehrenfeld,M. L.Brown-Luedi, andJ. W. B. Hershey. 1979. Alterations in initiation factor activity from poliovirus-infectedHeLa cells.J. Biol. Chem. 254:10973-10978. 17. Hewlett,M.J., J.K.Rose,and D.Baltimore.1976. 5'-Terminal
structureofpoliovirus polyribosomalRNA ispUp. Proc.Natl. Acad. Sci. U.S.A. 73:327-330.
18. James, A. L., and D. R. Tershak. 1981. Protein phosphoryla-tionsinpoliovirus infected cells. Can. J. Microbiol. 27:28-35. 19. Lee, K. A. W., and N. Sonenberg. 1982. Inactivation ofcap
binding proteins accompanies the shut off of host protein synthesis by poliovirus. Proc. Natl. Acad. Sci. U.S.A. 79:3447-3451.
20. Merril, C. R., D. Goldman, and M. L.Van Keuren. 1984. Gel proteinstains: silver stain. Methods Enzymol. 104:441-447. 21. Muthukrishnan, S., M. Morgan, A. K. Banerjee, and A. J.
Shatkin. 1976.Influenceof5'terminalm7G and2'-O-methylated residues on messengerribonucleic acid binding to ribosomes. Biochemistry 15:5761-5768.
22. Nomoto, A., Y. F. Lee,and E. Wimmer. 1976. The 5' endof poliovirus mRNAisnotcapped with m7G(5')ppp(5')Np. Proc. Natl.Acad. Sci. U.S.A.73:375-380.
23. O'Farrell, P. H. 1975. High resolution two-dimensional elec-trophoresisofproteins.J. Biol. Chem.250:4007-4021. 24. Patzelt, E., D. Blaas, and E. Kuechler. 1983. CAP binding
proteins associated with the nucleus. Nucleic Acids Res. 11:5821-5835.
25. Rose, J. K., H. Trachsel, K. Leong, and D. Baltimore. 1978. Inhibition of translationby poliovirus: inactivationofaspecific initiation factor. Proc. Natl.Acad.Sci. U.S.A. 75:2732-2736. 26. Schreier,M.H.,and T.Staehelin. 1973.Initiation ofmammalian
protein synthesis: the importance of ribosome and initiation factorqualityfor theefficiencyof in vitro systems. J.Mol.Biol. 73:329-349.
27. Shatkin, A. J. 1976. Capping of eukaryotic mRNA. Cell
on November 10, 2019 by guest
http://jvi.asm.org/
9:645-653.
28. Sonenberg, N. 1981. ATP/Mg2 -dependent cross-linking ofcap
binding proteins to the 5' end of eucaryotic mRNA. Nucleic Acids Res. 9:1643-1656.
29. Sonenberg, N., M. A. Morgan, W. C. Merrick, and A. J. Shatkin. 1978. Apolypeptide ineukaryoticinitiation factors that crosslinks specifically to the 5'-terminal cap in mRNA. Proc.
Natl. Acad. Sci. U.S.A.75:4843-4847.
30. Sonenberg, N., K. M. Rupprecht, S. M. Hecht, and A. J. Shatkin. 1979.EukaryoticmRNAcapbindingprotein:
purifica-tion by affinitychromatographyonsepharose coupledm7GDP.
Proc. Natl. Acad. Sci. U.S.A.76:4345-4349.
31. Tahara, S., M. A. Morgan, and A. J. Shatkin. 1981.Twoforms ofpurified m7G-cap binding protein with different effects on
cappedmRNAtranslation inextracts ofuninfected and polio-virus-infected HeLa cells. J. Biol. Chem. 256:7691-7694. 32. Towbin, H., T. Staehelin, and J. Gordon. 1979. Electrophoretic
transfer ofproteins from polyacrylamide gelsto nitrocellulose sheets: procedures and some applications. Proc. Natl. Acad.
Sci. U.S.A. 76:4350-4354.
33. Trachsel,H.,N.Sonenberg,A.J.Shatkin,J. K. Rose, K.Leong, J. E. Bergmann,J.Gordon, andD.Baltimore.1980. Purification ofafactor thatrestores translation of VSV mRNA inextracts
from poliovirus-infected HeLa cells. Proc. Natl. Acad. Sci. U.S.A. 77:770-774.
34. Vaitukaitis, J. L.1981. Production of anti-sera with small doses of immunogen: multiple intradermal injections. Methods Enzymol. 73:46-52.